
Organic Inputs Positively Alter the Bacteriome of Post-Agricultural Soils
by
 , , , , and
, , , , and
Tadeusz Malewski
1,
Piotr Borowik
2,
Patrycja Golińska
3,
Adam Okorski
4,
Ireneusz Olejarski
5 and
Tomasz Oszako
2,* 1
Museum and Institute of Zoology, Polish Academy of Science, ul. Wilcza 64, 00-679 Warszawa, Poland
2
Department of Forest Protection, Forest Research Institute, ul. Braci Leśnej 3, 05-090 Sękocin Stary, Poland
3
Department of Microbiology, Nicolaus Copernicus University, ul. Lwowska 1, 87-100 Toruń, Poland
4
Department of Entomology, Phytopathology and Molecular Diagnostics, Faculty of Agriculture and Forestry, University of Warmia and Mazury in Olsztyn, Pl. Łódzki 5, 10-727 Olsztyn, Poland
5
Department of Ecology, Forest Research Institute, ul. Braci Leśnej 3, 05-090 Sękocin Stary, Poland
*
Author to whom correspondence should be addressed.
Forests 2023, 14(9), 1711; https://doi.org/10.3390/f14091711
Submission received: 26 July 2023
/
Revised: 21 August 2023
/
Accepted: 22 August 2023
/
Published: 24 August 2023
(This article belongs to the Special Issue Advances in Detection and Identification of Insect Pests and Pathogens)
Abstract
:Agriculture can degrade soils and reduce microbial diversity. The reduction in microbial diversity of degraded soils is due to their long-term agricultural use. In most cases, such areas are afforested but rarely succeed in converting them into first-generation pine forests without adequately revitalizing the soils and restoring the natural relationships characteristic of forest habitats. This is possible thanks to the positive changes in soil biodiversity. To facilitate and enhance this phenomenon in the present experiment, various forms of organic matter were added to the soil: pine bark compost scattered on the soil surface (BCS) or placed under the roots of the seedlings at planting (BCR), woody debris (WW) and sawdust (S), while the control plot was not treated. The studies started in 2001 on two experimental plots, a poorer one in Bielsk and a more fertile one in Czarne Człuchowskie (eastern and northern Poland, respectively). The following year, 2-year-old pine seedlings (Pinus sylvestris L.) were planted on the plots. After 20 years, the physicochemical properties of the soil and its microbial composition were determined and compared with the control. The results encourage the use of organic matter for established pine forest crops on post-agricultural land for revitalization: C, N, and P content increased both in the organic layer and in the topsoil (up to 40 cm), where most fine roots are located. The total content of exchangeable base cations (Ca, Mg, K) and the sorption capacity of the soils (a measure of the ion binding capacity of the soil) also improved. The genetic analyses carried out using the molecular method (NGS) showed positive changes in the composition of the soil microbiome. Compared to poorer soil conditions in richer habitats, the number of taxa increases when organic matter is added, leading to significant qualitative changes in the bacteriome. The addition of organic material from the forest had a positive effect on the bacterial communities, which in turn accelerated the changes in the diversity of bacteriomes characteristic of agricultural soils and brought them closer to the forest ecosystem. The organic horizon was restored, and at the same time, the biodiversity of the soil microbiome increased, which is important for the health and sustainability of pine stands on formerly agricultural land.
1. Introduction
To increase forest cover and reduce the negative impacts of climate change on a global scale [1], both natural regeneration and silvicultural assumptions need to be implemented by growing forests on low-value land (including post-industrial land) and post-agricultural land exhausted by long-term use [2]. The long-term use of soils in agriculture (restricted crop rotation or monocultures) leads to their gradual degradation, especially in terms of biodiversity. This happens through intensive fertilization and the use of pesticides (especially insecticides, fungicides, and herbicides). As a result, these soils become less productive, until eventually agricultural production is no longer viable and such areas are designated for afforestation.
The most common forest-forming species that can cope with such conditions are pines and birches in temperate forests. However, they are susceptible to a root fungus of the genus Heterobasidion, which causes root and butt rot disease. The spores of these fungi are ubiquitous and benefit from the fact that there are no natural counterparts (mainly fungi as in forest ecosystems) on agricultural land. Bacteria, on the other hand, thrive in agricultural soils and are a very important group of microorganisms as they are responsible for biogeochemical cycling or decomposition and mineralization processes; most of them are also antagonists of fungi.
Our previous studies [2,3] showed that wood waste (residues after felling) can be successfully used as organic material to restore symbiotic mycorrhizal communities, as they contain spores of fungi that were abundant in the forest ecosystem [4]. Wood substrates not only restored organic content but also increased the amount of fungi antagonistic to pine pathogens.
The soil rhizosphere regulates processes supported by microbes (e.g., nutrient fixation and metabolism), and microorganisms in the rhizosphere are of great importance in the context of sustainable agriculture and environmental sustainability [5,6], as well as forest ecosystems. The positive effects of plant growth-promoting rhizobacteria (PGPR), as well as ectomycorrhizal and arbuscular mycorrhizal fungi on plants, suggest that they can substitute chemical fertilizers [7]. Soil microbes contribute to a variety of ecosystem functions, such as the breakdown of organic matter, symbiosis with tree roots, and their protection against pathogens. Plant root symbionts act as important regulators of plant productivity and are responsible for the acquisition of limiting nutrients, such as nitrogen (N) or phosphorus [8], especially in nutrient-poor forest ecosystems [9].
Until now, fungal communities in agricultural soils have been better studied by our team, but now, by using powerful molecular biology techniques (DNA analysis), we have the opportunity to better understand their bacteriome. The species composition and complex ecological relationships of soils, in which bacteria have important functions, are determined by physical, chemical, and biological factors, such as soil texture and chemical composition, as well as pH, precipitation, and temperature. In addition, there are biotic factors, such as the interaction with plant root secretions, the absence of mycorrhizal fungi in the soil after agricultural use, and litter as organic material undergoing decomposition (due to annual plowing) [10].
The occurrence of certain bacterial taxa with characteristic properties in the soil depends on the specific management of the soil [11]. Soil bacteria directly or indirectly promote plant growth, influence processes of vegetation succession, and can significantly affect ecosystem development [12,13]. The microbial diversity of forest soils depends on the content of organic matter and total nitrogen. An important parameter affecting the structure and function of the soil bacteriome is the ratio of carbon to nitrogen and the content of lignin or organic acids [14]. Coniferous forests have lower carbon and nitrogen content in the soil than deciduous forests, leading to higher biodiversity index values in deciduous forests [15].
The few studies dealing with the bacteriome of forest soils indicate that both the species composition of the bacteriome and the ecological indices used in scientific studies to assess microbial diversity are highly dependent on the tree species present in the habitat [16]. The species composition of bacterial communities is not only shaped by the vegetation in a particular forest habitat, but conversely, changes in bacterial populations also influence the composition and diversity of the vegetation [17].
According to Weng et al. [17], soil microorganisms, including bacteria, play a crucial role in rehabilitation and succession processes. In a study by Weng et al. [17], it was shown that soil bacterial diversity (expressed by the Shannon index) correlates positively with soil carbon and nitrogen content. Therefore, the process of restoring the correct ratio of soil microorganisms can be regulated by using organic matter as soil amendment [18], which could be particularly important for restoring degraded soils.
In view of this hypothesis, the addition of organic matter to planted pines should bring soil processes closer to those in forest ecosystems. The aim of the present study was, therefore, to use organic material as an additive for degraded soils after agricultural use in order to restore the natural bacteriome characteristic of forest soils.
2. Materials and Methods
In autumn 2000, the soil was prepared for afforestation on quite fertile mixed forest sites in the forest district (FD) of Czarne Człuchowskie (hereafter, Czarne) and Bielsk (a poorer site, and a dry and fresh coniferous forest). (Figure 1). In order to restore the organic level, various organic materials were spread in a quantity of approximately 1.5 m³/ha on trial plots of 0.3 ha each for subsequent revitalization of the soil (Figure 2). The following substrates were applied separately to each experimental plot: bark compost on the soil surface (BCS), wood waste (WW), bark compost on the roots (BCR) and sawdust (S). No treatments were applied in the control plot (C). The addition of each type of organic material was repeated in three experimental plots, which were spatially randomized to exclude random effects of soil conditions.
In the FD Czarne, BCS, BCR, WW, and S were applied to the entire soil surface (as mulch), creating an organic layer of about 5 cm (Figure 2). In the Bielsk FD plot, organic material (BCS, WW, and S) was only spread in the rows before planting, while bark compost was added at a rate of about 5 liters of BCR per plant at a depth of about 20 cm below the roots during planting. In the spring of 2001, the plots were planted with two-year-old pine seedlings, Pinus sylvestris L.
The compost came from the commercial processing of pine bark in the local forest nursery, the sawdust came from the local carpenter’s workshop, which is mainly involved in the processing of pine wood, and the residues from logging were collected by machines when the trees in the area were felled. The idea was that the application method should be simple, the organic material should be widely available, and everyone should be able to repeat the treatments on a larger scale, with the advantage of increasing the biodiversity of the soil.
2.1. Preparation of Soil Samples for the Analysis of Physicochemical Properties and DNA Extraction
In 2021, the regeneration of the organic level and the changes in the physical and chemical properties of the soil in all the designated experimental plots (compared to the control) were assessed, and the microbiological composition of the soil collected near the tree trunks (1–1.5 m) was determined using the next-generation sequencing (NGS) method. Cylinders of 100 cm3 were filled with soil (containing also fine roots) from different layers: organic and mineral 0–20 cm and 20–40 cm. The soil was then prepared (sieved) in the laboratory of the Forest Research Institute (IBL) for the analysis of macronutrient content: C, N, and P, and for DNA extraction. The content of exchangeable cations, which are adsorbed by the sorption complex of the soil and can be exchanged with cations in the soil solution under certain conditions, was also determined. The sorption complex, which comprises soil components that can bind ions, particles, and suspended matter, as well as vapors, gases, and microorganisms to their surface, was determined using the soil sorption method.
The following physicochemical analyses of soil samples were carried out:
- pH in H2O and KCl potentiometry;
- Cog high-temperature combustion with TCD detection;
- Nog high-temperature combustion with TCD detection;
- Egner-Riehm P2O5;
- Ca, K, Mg, Na exchangeable in 1 M ammonium acetate extract by ICP-OES atomic emission spectrometry;
- Hh (hydrolytic acidity) modified caps.
The sum of the exchangeable bases is the difference between the amount of HCl taken for analysis and the amount of this acid remaining in the solution after the exchangeable cations have been displaced from the sorption complex. The exchangeable bases in soils depend mainly on the humus and clay mineral content, which shape the sorption properties, and on the calcium carbonate content. This is the sum of the basic cations taken up by the soil, Ca, Mg, K, Na, expressed in [cmol(+)/kg]. The calculation of the properties of the sorption complex: Sz (total alkali content) = Ca + K + Mg + Na, Th (total sorption capacity) = Sz + Hh, Vh (degree of saturation of the sorption complex with alkali) = (Sz:Th)100%.
To determine the physical properties, nine soil samples were taken in 100 cm3 cylinders from the mineral layers 0–20 cm and 20–40 cm for each experimental variant. They were then thoroughly mixed to obtain a composite sample for each treatment from which the soils were taken for physicochemical analyses. Determinations made on the collected samples include volume density in the natural system of soil dry matter (g/cm3 and actual moisture content by drying at 105 °C to a constant weight (g/100 cm3).
For the metagenomic analyses, following the organic layer, 24 soil cylinders (100 cm3) were taken from a depth of about 5 cm from the Bielsk and Czarne Człuchowskie locations. Each sample was collected in three replicates, each from a depth of about 5 cm (a total of 72 samples). The soil samples were thoroughly mixed and 1 g was taken for DNA extraction. Details of the DNA extraction were described in an earlier paper [3].
2.2. Library Preparation for NGS
Multiplex amplicon libraries were prepared using the two-step PCR protocol described by Caporaso et al. [19]. This procedure consists of a double PCR amplification. The first PCR uses amplicon-specific primers, including an Illumina adapter overhang (amplicon PCR), and the second, cycle-limited PCR is used to include Illumina index adapters for multiplexing (index PCR). Amplification of the V3–V4 regions of the 16S rRNA gene was performed according to the protocol described by Klindworth et al. [20]. The primer pairs used were 5′-TCGTCGGGCAGCGTCAGATGTGTATAAGACAG CCTACGGGNGGCWGCAG-3′ (forward primer), and 5′-GTCGTCGTGGCTCGGAGATGTGTATAAGACAG GACTACHVGGGTATCTAATCC-3′ (reverse primer) (Nextera-Illumina adapter overhang sequence in italics).
PCR reactions were performed using the following program on a Veriti 96-well thermal cycler (Thermo Fisher Scientific, Waltham, MA, USA): initial denaturation for 3 min at 95 °C, followed by 30 cycles of 30 s at 95 °C, 30 s at 55 °C, 30 s at 72 °C, and a final extension cycle for 5 min at 72 °C.
2.3. Processing and Analysis of Sequencing Data
The demultiplexed and barcode-free DNA sequences were analyzed using Geneious 10.2.3 software (Geneious, Auckland, New Zealand) [21]. The BBDuk plugin for trimming NGS data was used to trim NGS data [22]. The 16S biodiversity tool was then used for taxonomic analysis of the microorganisms in the samples. This tool assigned a taxonomy to each sequence and compared it to the ribosomal database project (RDP) classifier [23]. OTUs with a read count below 100 were excluded from further analysis. The sequences were deposited in the NCBI Sequence Read Archive (SRA) under the study accession numbers PRJNA980196 (Bielsk) and PRJNA980196 (Czarne Człuchowskie).
2.4. Analysis of Bacterial Biodiversity
The data analysis was carried out using Python 3.10 codes. For data analysis and visualization, libraries such as Pandas [24], Matplotlib-Venn, Matplotlib [25], Seaborn [26], SciPy [27], scikit-bio [28] were used.
The analysis of similarities was conducted using Venn diagrams, comparing all pairs of independently investigated soil variants for both experimental sites.
The similarities between the studied plots were calculated using the Jaccard index at the genus level, represented by the following formula:
where , are the number of bacterial taxons detected in plots x and y plots, respectively, and is the number of taxons detected at both sites.
Jaccard’s formula was also used to calculate the distances between plots in the hierarchical grouping procedure. In this approach, the similarities between plots are calculated and the most similar plots are grouped into clusters in a recurrent algorithm. In this case, the reciprocal of was used as the distance for selecting the grouped clusters.
3. Results
Number of Individual Taxa Compared to Studied Soil Variants
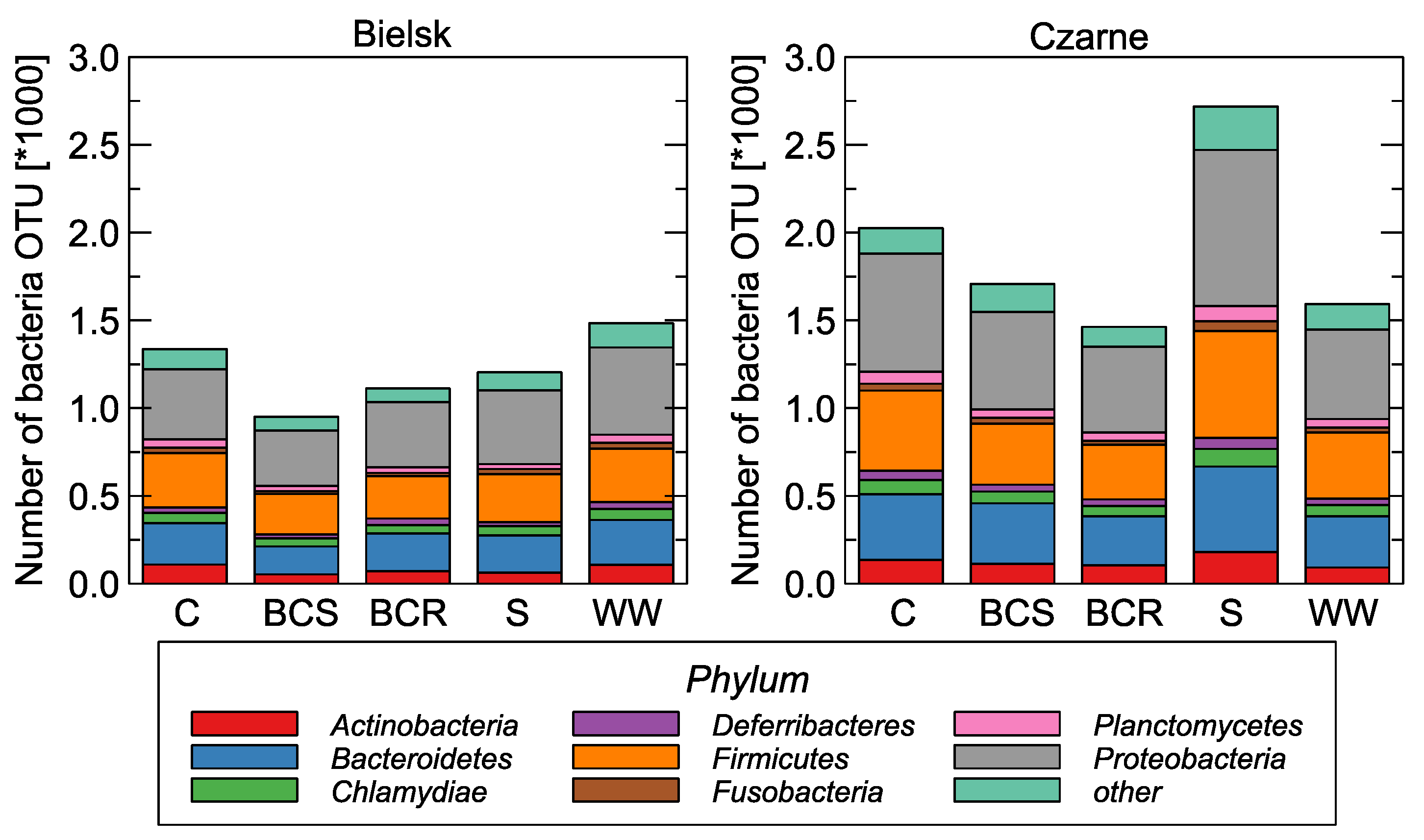
Raw sequences were preprocessed and high-quality reads were assigned to OTUs, as described [3]. The prepared analysis assigned the OTUs to 571 genera. The bacterial communities in the soil were strongly dominated by eight taxons: Actinobacteria, Bacteroidetes, Chlamydiae, Deferribacteres, Firmicutes, Fusobacteria, Planctomycetes, and Proteobacteria (Figure 3).
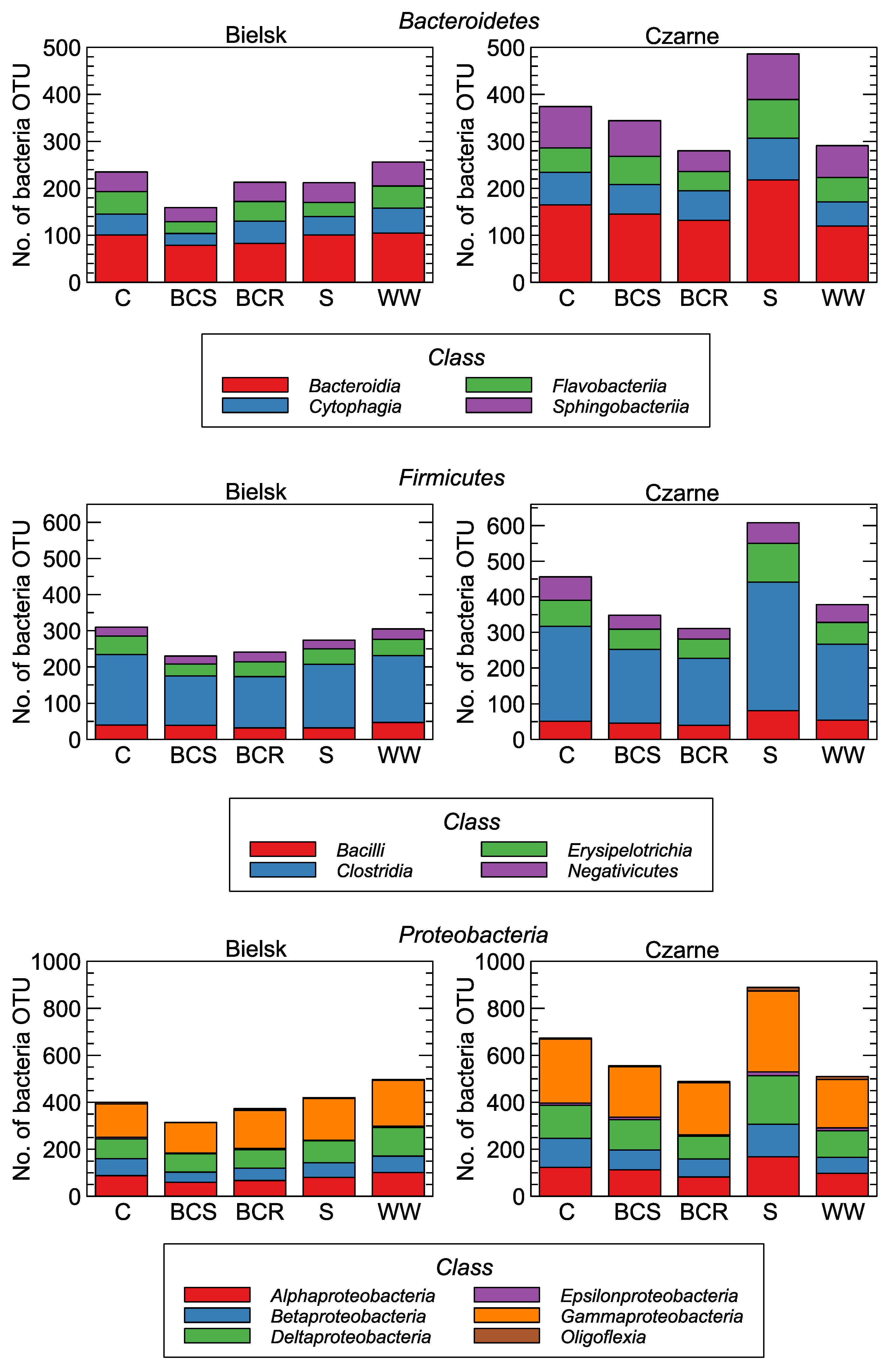
Within Bacteroidetes, four taxa were the most numerous: Bacteroidia, Cytophagia, Flavobacteria, and Sphingobacteria. Among Firmicutes, Clostridia were the most abundant, followed by Bacilli. Among Proteobacteria, Alpha-, Beta-, Delta-, and Gamma-Proteobacteria were the most abundant taxons (Figure 4).
Metagenomics analyses showed that Proteobacteria were the most abundant in the soil of control plants as well as in plots with organic material, followed by Firmicutes and Bacteroidetes. Firmicutes and Bacteroidetes responded most strongly to the supply of organic material. Significant differences in the content of these taxons were also found between Bielsk and Czarne (Figure 3).
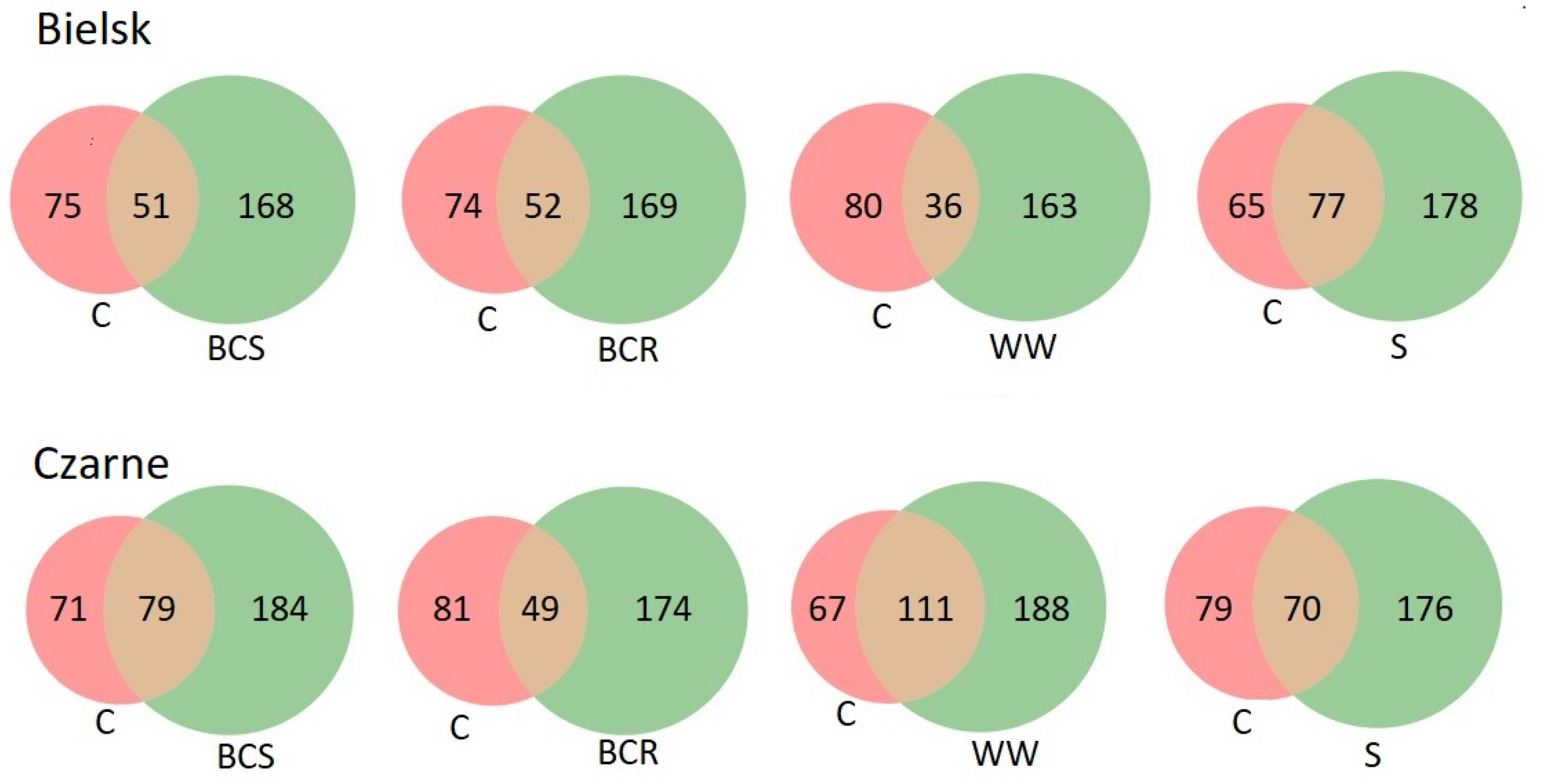
Enrichment of the soil with organic substrates led to changes in the composition of the bacteriome. After the addition of organic material in Bielsk, new genera appeared from 163 (wood waste) to 170 (sawdust). The appearance of new genera was also noted in Czarne Człuchowskie (from 174 after the addition of BCR to 188 after the addition of wood waste). The addition of organic material led not only to the appearance of new genera but also to the disappearance of a considerable number of genera present in the soil of the control plot. The number of disappeared genera varied between 65 (sawdust) and 80 (wood waste) in Bielsk and between 71 (BCS) and 81 (BCR) in Czarne Człuchowskie.
In the control plot in Bielsk, the bacteriome consisted mainly of Bacteroidetes, where 31 genera were found: Anaerophaga, Cnuella, Cruoricaptor, Fibrisoma, Fluviimonas, Gangjinia, Geofilum, Haliscomenobacter, Hydrobacter, Imtechella, Leadbetterella, Macellibacteroides, Microbacter, Mongoliitalea, Mooreia, Mucinivorans, Namhaeicola, Nubsella, Ornithobacterium, Perexilibacter, Petrimonas, Phocaeicola, Portibacter, Pseudofulvibacter, Rikenella, Rubrivirga, Rudanella, Salinirepens, Sporocytophaga, Tangfeifania, and Tannerella), 43 genera of Firmicutes: (Alkalibaculum, Allisonella, Allobaculum, Anaerospora, Butyricicoccus, Caminicella, Cellulosibacter, Dehalobacter, Desulfitibacter, Desulfonispora, Dolosicoccus, Eggerthia, Eremococcus, Ezakiella, Fastidiosipila, Fuchsiella, Gallicola, Gelria, Geothermomicrobium, Gracilibacter, Guggenheimella, Halanaerobaculum, Holdemanella, Hydrogenibacillus, Hydrogenoanaerobacterium, Ignavigranum, Johnsonella, Moryella, Negativicoccus, Oxobacter, Parvimonas, Pelospora, Peptococcus, Pilibacter, Proteiniborus, Selenihalanaerobacter, Sharpea, Soehngenia, Solobacterium, Sporanaerobacter, Syntrophothermus, Thermicanus, Tuberibacillus) and 46 genera of Proteobacteria (Amylibacter, Antarctobacter, Arsenophonus, Bilophila, Brackiella, Candidatus Carsonella, Coxiella, Craurococcus, Dehalospirillum, Desulfobacca, Desulfonatronobacter, Desulfosalsimonas, Desulfospira, Echinimonas, Endobacter, Enhygromyxa, Extensimonas, Ferrovum, Gilliamella, Halospina, Hydrogenovibrio, Isochromatium, Methylorosula, Methylosphaera, Moellerella, Nicoletella, Oligoflexus, Parablastomonas, Paralcaligenes, Parapusillimonas, Pelagibaca, Porticoccus, Pseudobacteriovorax, Psychrobium, Pyxidicoccus, Quatrionicoccus, Seminibacterium, Smithella, Spiribacter, Spongiispira, Stenoxybacter, Syntrophorhabdus, Telmatospirillum, Thermodesulforhabdus, Thiohalophilus, Woodsholea).
The addition of bark compost leads to a marked change in the bacteriome. The greatest changes occur in the Firmicutes, where 25 new genera appear (Atopostipes, Catenisphaera, Catonella, Compostibacillus, Dendrosporobacter, Dethiosulfatibacter, Eisenbergiella, Faecalibacterium, Falsibacillus, Finegoldia, Halarsenatibacter, Halocella, Halothermothrix, Lacticigenium, Mahella, Marvinbryantia, Murdochiella, Proteinivorax, Pseudoflavonifractor, Salimesophilobacter, Salinithrix, Selenihalanaerobacter, Subdoligranulum, Syntrophaceticus, Thermodesulfobium, Thermoflavimicrobium) followed by 17 new genera of Proteobacteria (Ameyamaea, Candidatus Pelagibacter, Candidatus Procabacter, Cucumibacter, Desulfoconvexum, Desulfofustis, Laribacter, Lawsonia, Morococcus, Orientia, Permianibacter, Phocoenobacter, Sandaracinus, Serpens, Succinivibrio, Thermodesulforhabdus, Thioalkalibacter) and 6 new genera of Bacteroidetes (Ichthyenterobacterium, Kriegella, Mangroviflexus, Shivajiella, Siphonobacter, Soonwooa).
The appearance of new genera after the addition of bark compost is accompanied by the disappearance of many bacterial genera. In Firmicutes the largest number of genera disappeared (13): (Abiotrophia, Alkalibaculum, Anaerosalibacter, Cellulosibacter, Desulfonispora, Eremococcus, Gallicola, Guggenheimella, Johnsonella, Oxobacter, Selenihalanaerobacter, Sharpea, Soehngenia), followed by 10 genera of Proteobacteria (Coxiella, Endobacter, Hydrogenovibrio, Paralcaligenes, Parapusillimonas, Pelagibaca, Porticoccus, Pyxidicoccus, Telmatospirillum, Thermodesulforhabdus) and 5 genera of Bacteroidetes (Fibrisoma, Gangjinia, Microbacter, Pseudofulvibacter, Rudanella).
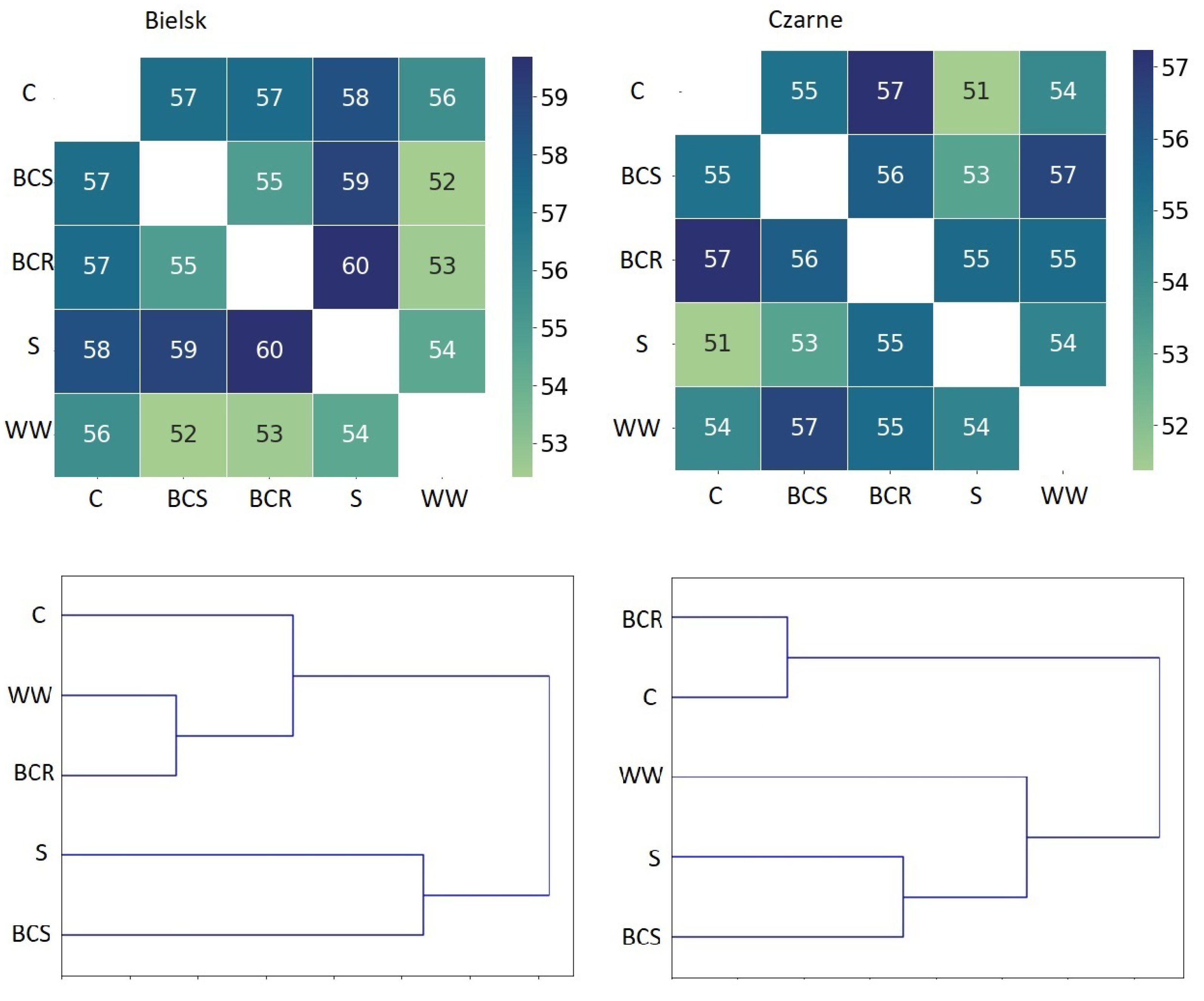
Similar changes in the bacteriome were found after the addition of other organic material in Bielsk and Czarne Człuchowskie. The addition of organic matter (BCS, BCR, WWW and S) increased the number of bacterial genera in each case compared to the control (C), and this was true for both poor and rich soil conditions Figure 5. The analysis of the similarity index showed that, at both sites, the greatest changes in the bacteriome composition occurred after the addition of sawdust and bark compost below the surface (Figure 6).
Bacterial diversity in the soils of poorer sites is lower than in richer ones, which is consistent with the idea that former agricultural land in transition to forest ecosystems should have fewer bacteria and more fungi (both groups of organisms are strong antagonists). Therefore, the addition of organic matter to (rather poor) land that is suitable for pine cultivation is beneficial. This also applies to the quality of the pine logs, because on richer soils, there are usually too many knots, which reduce the quality of the wood.
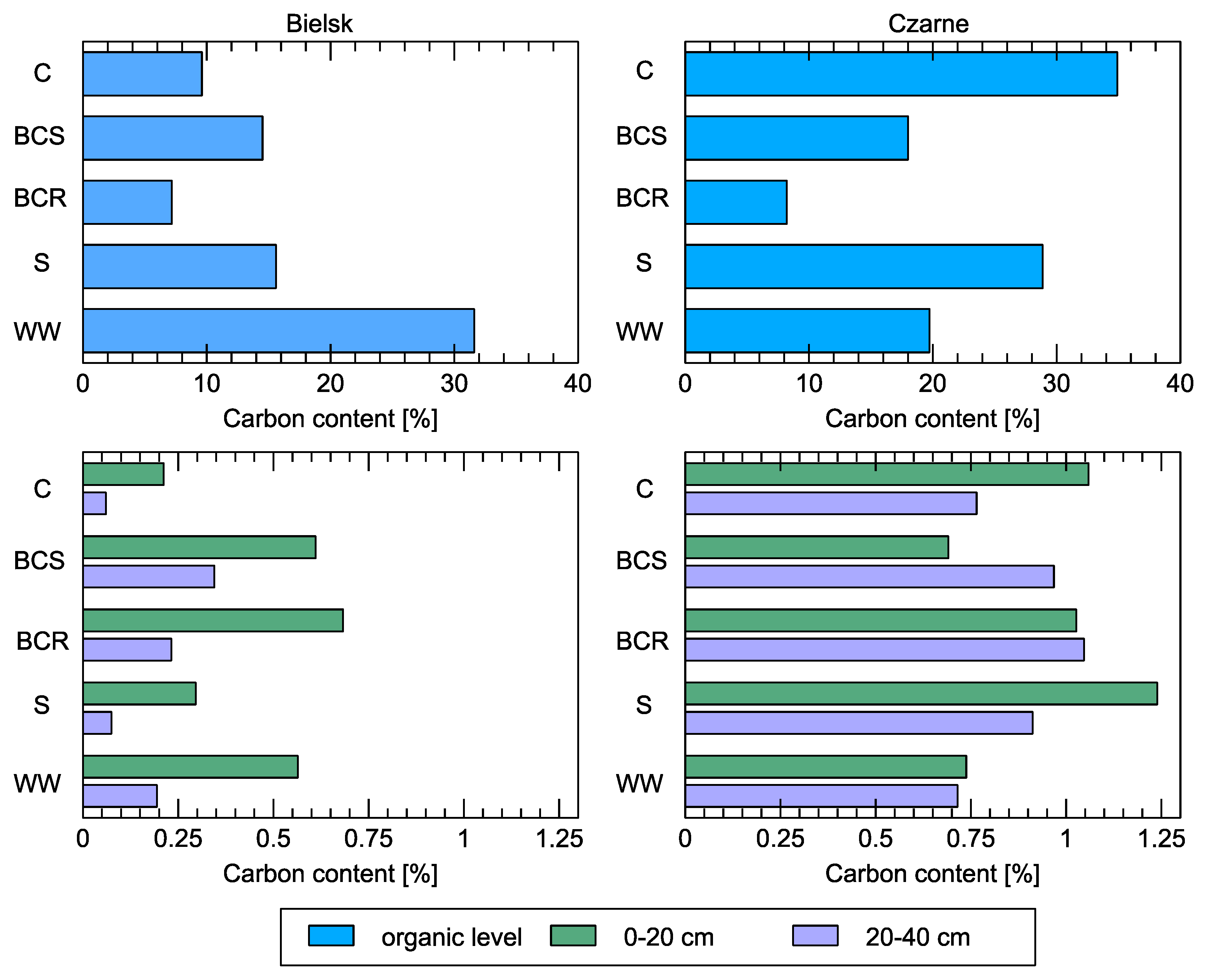
4. Soil Analysis
In the organic layer in Czarne (a more fertile habitat of the fresh mixed conifer forest), the highest carbon content (even higher than after the treatments) was found in the control (Figure 7). In Bielsk, it is exactly the opposite, but it is a poorer habitat (dry mixed conifer forest/fresh forest). The addition of organic material, especially sawdust, enriched the topsoil twice as much with carbon compared to wood residues (Table 1). The addition of compost also had a positive effect on the organic carbon content of the entire cross-section down to the investigated depth of 40 cm. On the fertile site, these differences were not large compared to the control (hence, the effect on bacterial diversity is also likely to be smaller). On the poorer site, however, the addition of organic matter was very beneficial and improved the carbon supply of the top layer (up to 20 cm). The organic carbon content in the 0–20 cm thick soil layer varied between 0.56 and 0.68% after the treatments, while it was three times lower under control conditions with only 0.21%. The organic carbon content in the 20–40 cm thick layer of the treated soil varied between 0.19 and 0.34%, while it was only 0.06% under the control conditions.
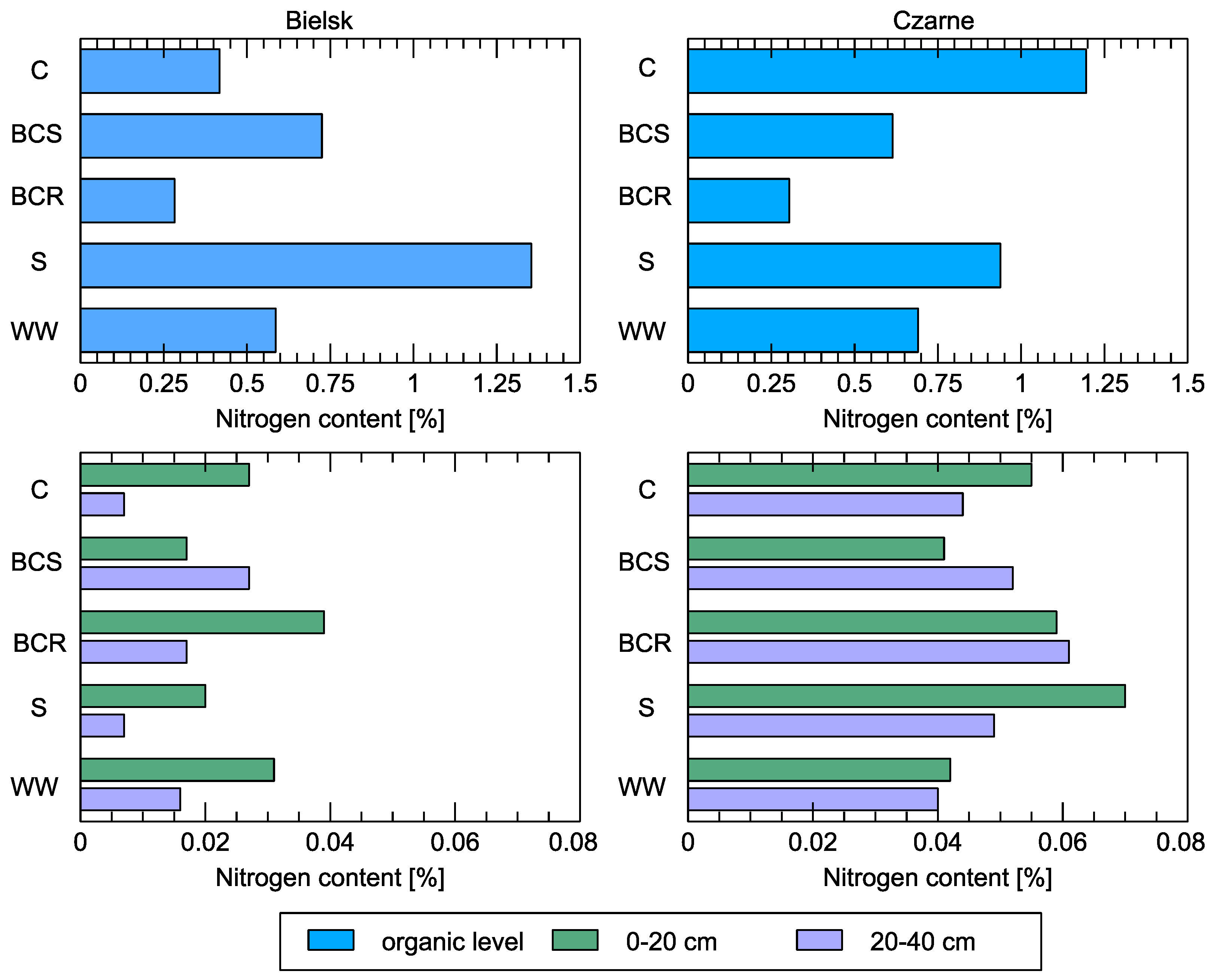
The nitrogen content of the soil also improved, but only at the worse station of the Bielsk observation plots (Figure 8). Large nitrogen contents were obtained after soil improvement with sawdust (S) in the organic layer in Bielsk and the superficial zone in Czarne (up to 20 cm depth). Application of various organic components to the plot in the Bielsk forest district resulted in soil enrichment (compared to control conditions), especially of organic carbon and nitrogen, in both the organic and mineral horizons. In the organic horizon, the organic carbon and nitrogen content was highest in the experimental plot in the Bielsk forest district (where wood waste was applied), with 31.6 and 1.35%, respectively; the organic carbon and nitrogen content was also higher in the variants with bark compost and sawdust than in the control. The nitrogen content in the 0–40 cm thick layer of the soils under control conditions was only 0.02%.
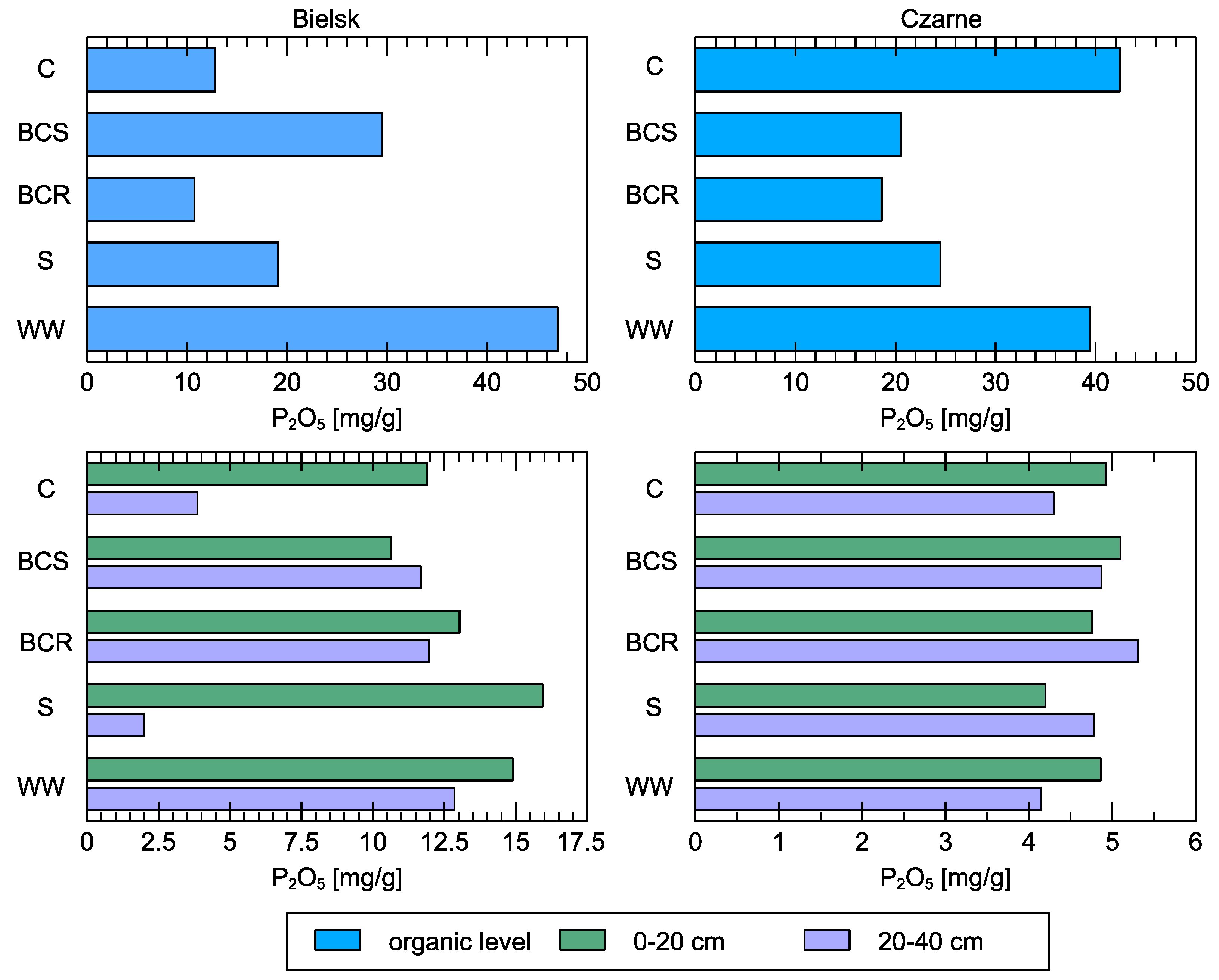
The phosphorus content of the organic horizon of the untreated soils was low in Bielsk and high in Czarne (Figure 9). Wood waste (WW) was the main source of P in the created organic layer, especially in Bielsk.
In the control (C) on the poorer site, P content also differed in the upper soil layers (0–20 cm), where about three times more of this element was present than in deeper layers (20–40 cm). Even after applying sawdust to the soil surface, the P content was many times higher in the shallower soil layers than in the deeper soil layers. In the other treatment variants in Bielsk, the differences in P content were much greater than at the more fertile site in Czarne, where the P content was at a similar level everywhere.
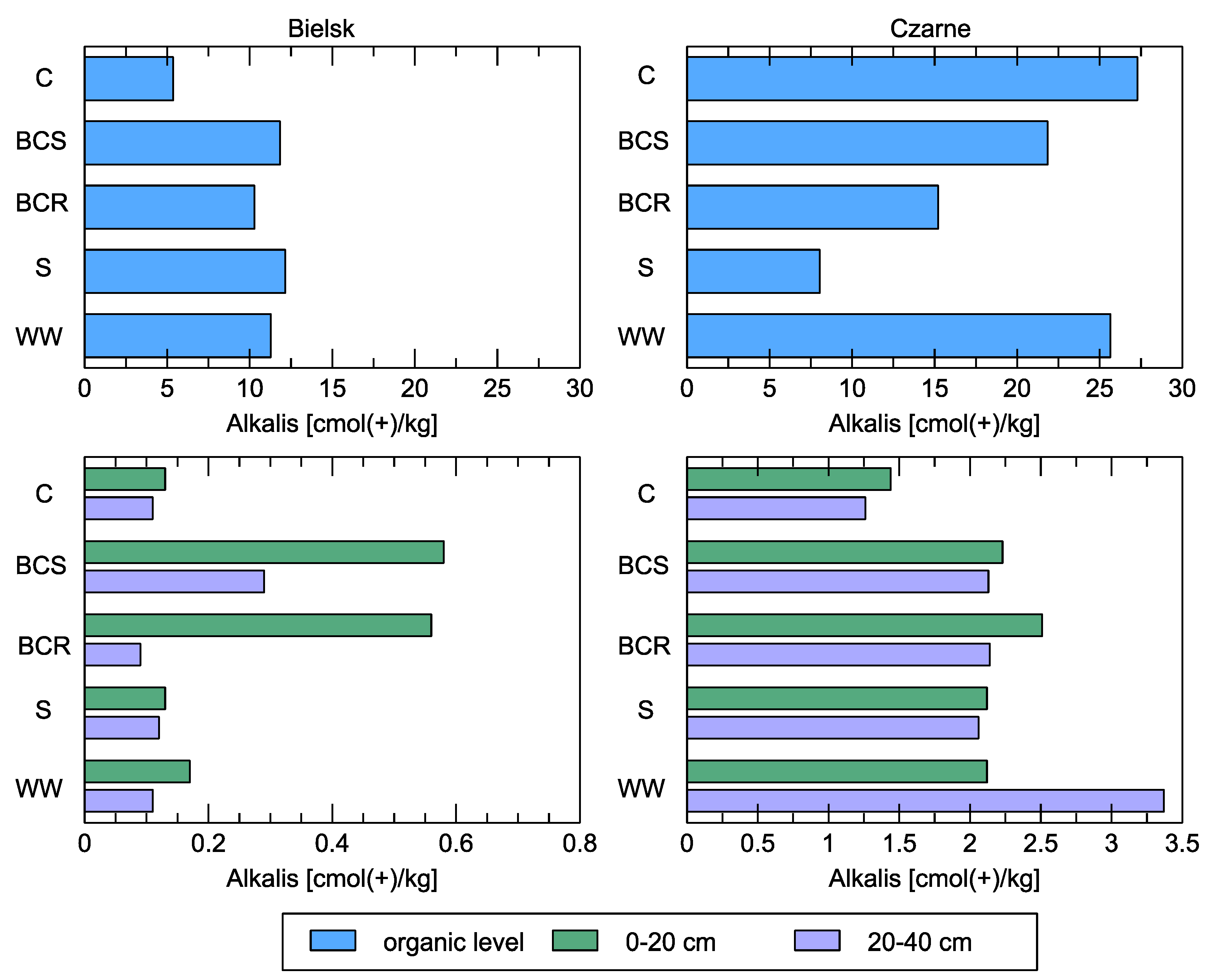
The reaction of the soil to the addition of organic matter had a positive effect on increasing the sum of exchangeable bases in the poor site in Bielsk, while no such reaction was observed in the organic layers of the rich plot in Czarne (Figure 10). In soils up to a depth of 40 cm, only one visible reaction occurred in practically all the variants studied (BCR, BCS, S, and WW). In the latter case, it was strongest at a depth of 20–40 cm and half as strong as in the control.
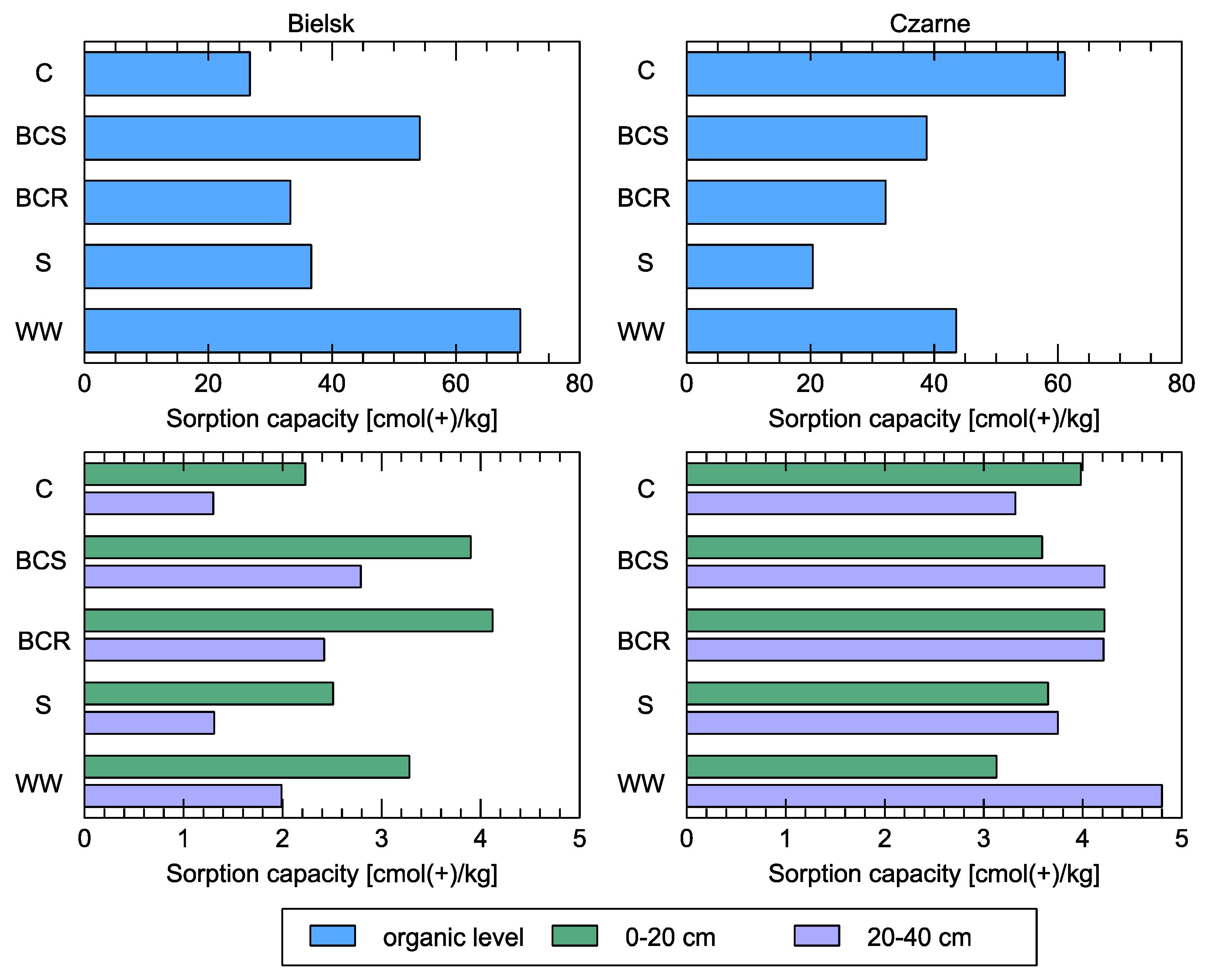
Sorption capacity in the organic layer increased at the poorer site in Bielsk (compared to the control) after the application of compost to the soil surface and wood waste (Figure 11). The improvement of soil properties in this respect also occurred at a depth of 0 to 20 cm. No significant differences were found in the more fertile soils in the Czarne forest district compared to the control.
The characteristics of the sorption complex of the soils in the poor experimental plot in the Bielsk show that the sum of bases in the organic horizons under the control conditions was 5.36 cmol (+)/kg and was twice as high as in the variants where treatments were carried out, where it ranged from 10.88 to 12.05 cmol (+)/kg, indicating an increase in the content of macroelements (Ca, K, Mg). The sorption capacity of the organic horizon under the control conditions was also lower than that of the treatment variants, at 36.72 cmol (+)/kg, and ranged from 36.36 to 70.40 cmol (+)/kg for the other variants. The sorption capacity increased after the treatments in the mineral layers (0–40 cm) of the soil and ranged from 1.86 to 3.35 cmol (+)/kg. In the control, it was lower at 1.76 cmol (+)/kg.
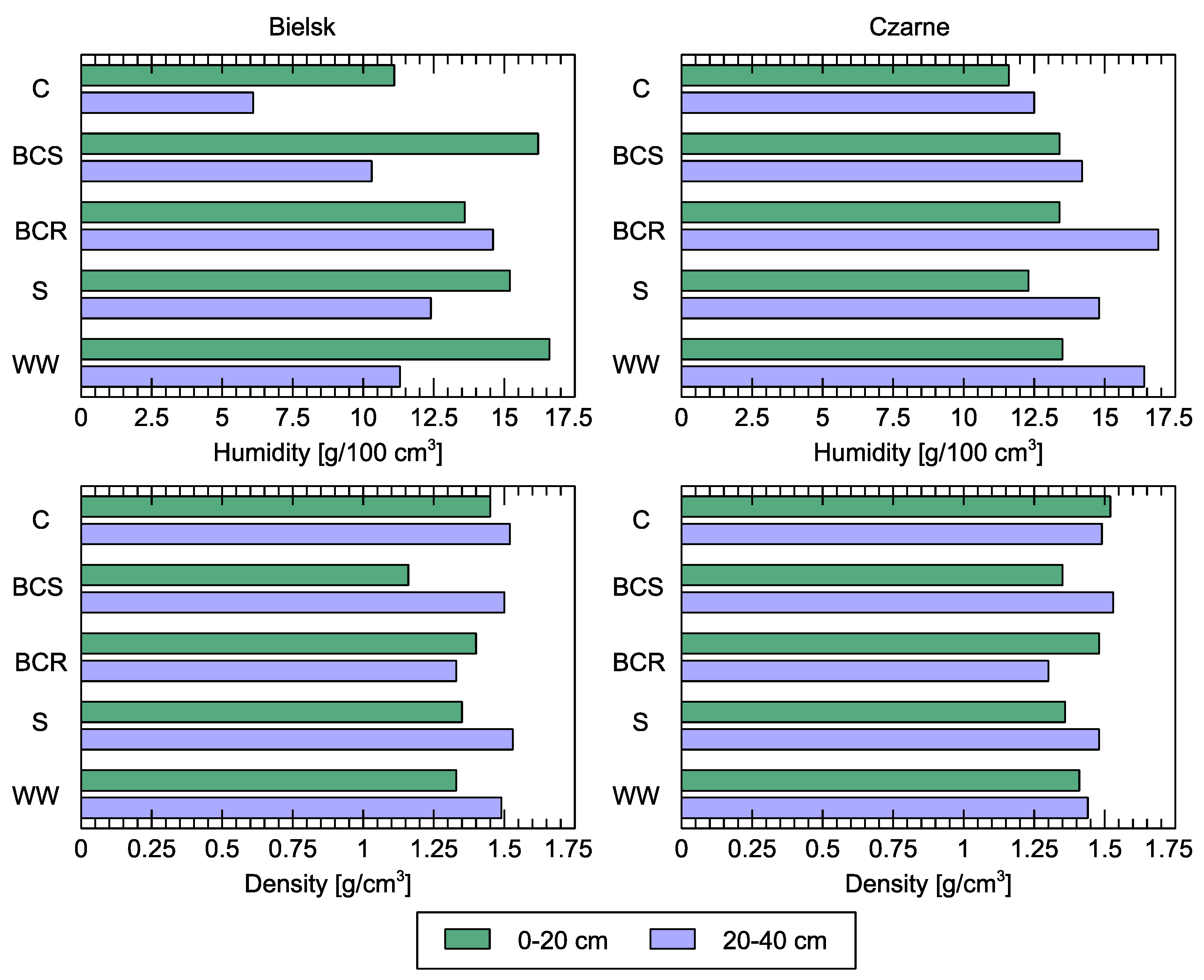
Currently, there are clear differences in soil moisture and density between plots (Figure 12). The volume density in the 0–20 cm thick soil layer of the plots in the Bielsk forest area was highest in the control variant (C) with 1.45 g/cm3. In the other variants, the volume density was lower and ranged from 1.35 (BCS variant) to 1.48 g/cm3 (BCR variant).
The average current soil moisture in the Bielsk forest district in the 0–20 cm layer was highest in the woody debris (WW) variant (16.6 g/100 cm3) and lowest in the bark compost (BCS) variant (13.6 g/100 cm3). In the control variant (C), the value of this parameter was 11.1 g/100 cm3. In the 20–40 cm thick layer, the current moisture content of the soil was lowest in the control variant (6.1 g/100 cm3), and after the treatments, it ranged from 10.3 in BCS to 14.6 g/100 cm3 in BCR.
The bulk density in the 0–20 cm thick soil layer of the plots in the experimental area in the forest district of Czarne Człuchowskie ranged from 1.35 (BCS variant) to 1.48 g/cm3 (BCR variant). In the control variant (C), the value of this parameter was 1.52 g/cm3. In the 20–40 cm thick soil layer, the volumetric density was lowest in the BCR variant with 1.30 g/cm3, while in the other variants, it ranged between 1.44 and 1.53 g/cm3 and corresponded to the control conditions with 1.49 g/cm3.
The actual moisture content of the soil in the 0–20 cm thick soil layer in the forest district of Czarne Człuchowskie was highest (13.5 g/100 cm3) in the variant with wood waste (WW). In the variants with bark compost (BCS) and (BCR), the value was 13.4 g/100 cm3, and with sawdust (S), it was 12.3 g/100 cm3. In the control variant (C), the value of this parameter was 11.6 g/100 cm3. In the 20–40 cm thick layer, the actual moisture content of the soil was lowest in the control variant at 12.5 g/100 cm3, and after the treatments, it ranged from 14.2 g/100 cm3 in BCS to 16.9 g/100 cm3 in BCR.
5. Discussion
The literature on this topic shows that the effects of fertilizers on the soil microbiome in the long-term sustainable productivity (LTSP) experimental plots persist long after the addition of fertilizers has ceased. Metagenomics analyses of bacterial and fungal communities in soils from two LTSP sites (both 25 years old) showed that fertilizer addition resulted in significant changes in both bacterial and fungal communities, while the removal of organic matter at harvest had persistent effects on the composition of fungal species in the soil ecosystem [29]. Changes in bacterial and fungal community structures and functions were also found after 25 years of water-shedding of forest soils [30]. Long-term changes in the soil microbiome have also been noted after curing insect infestations [31]. Although current soil conditions undoubtedly affect the composition of the microbiome, experiments with long-term sustained productivity have demonstrated that fertilizers can alter bacterial and fungal communities in soils not only after 25 years [29], but even after 60 years of continuous forest management of Pinus radiata [32].
Despite the fact that there is some literature on the long-term effects of fertilization on the soil microbiome, we do not insist on establishing a link between a treatment carried out twenty years ago, but rather on showing the differences between treatments. Even though the treatments carried out so long ago are no longer present in the soil, they have helped to bring the soil biodiversity closer to the forest ecosystem compared to the untreated control plots. The above statements can be supported by the current bacteriome and the physicochemical properties of the soil when comparing the treatments. Although the soil microbiome reacts within a year and depends mainly on the organic matter present, changes in the biodiversity of the forest soil could also play an important role, e.g., due to the pine bark compost or the wood residues collected after logging, both of which contain bacteria and fungi from the forest ecosystem. Both were introduced into the former agricultural soil to make the afforestation successful. Against this background, the twenty-year treatment is significant because of the turnover of organic matter, as it has set in motion natural processes that also occur during the abandonment of agricultural land and spontaneous reforestation.
5.1. Special Features of the Former Agricultural Land Compared to the Forest and the Concept of Treatment
Forest ecosystems on former agricultural land are characterized by high variability of soil conditions. The most common soil types on former agricultural land include podzol soils and rusty soils of glacial, fluvial, and dune sands. According to the classification of agricultural land for forestry, there are habitats ranging from dry and fresh coniferous forests (Bielsk) on light soils to mixed coniferous forests (Czarne Człuchowskie) and fresh forests on heavy soils. Light post-agricultural soils, such as in the Bielsk forest district, have a wider range of pH values (3.4–7.3) than forest soils (3.3–4.7) and are characterized by a wider range of nutrient contents compared to forest soils. This includes soils with higher nutrient contents than forest soils as well as soils with nutrient deficiencies as in the Bielsk plot. Heavy post-agricultural soils also have a higher pH (4.0–7.0) than forest soils (3.8–5.7) and are characterized by a wider range of nutrient contents compared to forest soils; this was the case in the Czarne Człuchowskie plot.
Afforestation of agricultural land encounters numerous difficulties, withstand death often due to infestation with pine root and butt rot disease. Silvicultural and forest protection recommendations do not take soil conditions into account. However, the chemical, physical, and biological conditions of soil are often the cause of health problems in stands on land formerly used for agriculture. Soils after agricultural use differ from forest soils primarily in the absence of an organic horizon. The organic matter accumulated on the surface of forest soils (with varying degrees of decomposition) fulfills numerous functions, such as water retention, protection of the soil from erosion, temperature fluctuations, and determination of biological, physicochemical, and physical properties. The moisture measurements in Figure 12 show the water content of the soil at the time of the summer measurement in June. Although this is a very dynamic and temporally fluctuating variable, it shows the protective function of the soil mulch compared to the control. The precipitation frequency in June is usually not very high in Poland, and at both sites, it had not rained for about two weeks before sampling. In addition, the addition of organic matter reduced the density of the soil (especially in the layer up to 20 cm, where most of the roots are located), which facilitates the absorption and storage of water during rainfall.
The decomposition of organic matter provides a constant supply of macro- and micronutrients to the mineral horizons and influences the nutritional status and vitality of individual trees. It is also a source of energy for the development of microorganisms, including mycorrhizal fungi. The agricultural soils on which crops are grown each year must be mechanically worked, intensively fertilized with mineral fertilizer, and frequently limed; pesticides and herbicides are also used. The soils destined for afforestation after agricultural use are usually completely degraded: sterilized, with a subsoil of plowshares, with an excess of nitrogen, calcium, and potassium, and a deficiency of magnesium, as in our case. The transformation of agricultural soils into forest soils is a slow process and without silvicultural interventions, it is difficult to achieve stable stands. For this reason, it is very important to determine soil conditions and implement appropriate fertilization measures before afforestation. Sustainable and stable forest ecosystems develop on forest soils whose species composition is compatible with the forest habitat type, while stands on soils that have developed after agricultural use are susceptible to threats, especially biotic threats, and this applies to both study areas. This is favored by disturbances in pH and mineral content, and by the absence of fungi antagonistic to pathogens.
On post-agricultural sites, the revitalization measures we implemented (including organic fertilization) aimed to accelerate succession processes toward forest ecosystems and contribute to the development of pine stands that are more resilient to a variety of threats. The issue of revitalizing soils on post-agricultural land has already been addressed [33,34]. Revitalization of soils with sawdust was also described by [35] in 1999, which prompted the IBL Department of Ecology to carry out investigations on established experimental plots with shredded wood waste, bark compost, and sawdust in 2001. The measures carried out were aimed at restoring the organic horizon, the importance of which IBL researchers have written about previously [36,37]. Similar results have been presented in the work of other research teams from Poland and abroad [37,38]. The prevailing opinion among scientists is that enriching afforested post-agricultural land with organic components in the form of composts, wood waste or sawdust improves the fertility of weak soils and stimulates beneficial microbial processes in the soil environment, increases the proportion of antagonists and at the same time increases the resistance of roots to pathogen infestation.
The effects of fertilizers on the soil microbiome can be long-lasting. A study of fertilizer addition to experimental plots for long-term sustainable productivity showed that the effects on bacterial and fungal communities were noticeable after 25 years [29] and even after 60 years [32]. Similarly, the effect of liming on the forest soil microbiome was also found after 25 years of treatment [30].
This paper describes the treatment of degraded agricultural soils with different types of organic matter in transition to a land use change from agriculture to forest. The same soils were sampled twenty years later to draw conclusions about the effects of the different types of treatment. The pine stand was excluded from other silvicultural treatments (e.g., early and late clearing) during these twenty years. The powerful molecular method 16S rRNA metagenomic was used to compare soil microbial communities. Twenty-year studies are rare and, therefore, particularly interesting. Of course, after such a long period of time, the question arises whether the observed effects are due to the treatment twenty years ago or to everything else that has happened since. However, we conclude that all these possible events have affected all study plots equally. A comparison of the bacterial communities in the treatment and control plots allowed us to assess the impacts of the addition of different organic matter on the biodiversity of the soil samples studied.
5.2. Relationships between Soil Fertility and the Role of Bacteria in Supporting Plant Growth
Bacteria predominate in agricultural soils, while fungi predominate in forest soils. Figure 2 and Figure 3 show the qualitative data that changed after the addition of organic material. They show how diverse the bacteria are (how many species). If the environment is unfavorable for the bacteria, many of them die out and only the most resistant species remain. If the environment was favorable, different species survived.
The revitalization of degraded soils after agricultural use facilitated nutrient uptake by planted forest seedlings, probably in part due to the beneficial bacteria that contribute to the cycling of biogenic elements after the addition of organic matter. In this way, the use of organic material helped to improve the properties of degraded soils, especially in poorer areas. Bacterial diversity decreases in a poorer habitat and increases in a more fertile habitat. In afforestation, it is important that there are fewer of them in the forest ecosystem, as they are competitors (antagonists) for fungi. In contrast to agricultural land (where there are only a few species of fungi that break down lignin or cellulose), there should be more of these fungi than bacteria in the forest. We have found that sawdust in particular promotes bacterial biodiversity. A more fertile soil binds more carbon (C) (Figure 6), and the addition of organic matter leads to its decomposition (where there is a lot of it, as in Czarne). Where there is little C (Bielsk), the addition of organic matter increases the C content (favorable for poor habitats).
The increase in C content in the organic matter (0–20 cm) is particularly striking. Fertility is also reflected in the N content (more in Czarne), and after the addition of organic matter, its decomposition and release for the plants take place. On the poor sites of Bielsk FD, it is the other way around. An exception is sawdust, which probably has to be decomposed first. The same applies to P.
Plant root symbionts act as important regulators of plant productivity and are responsible for the acquisition of limiting nutrients, such as nitrogen (N) or phosphorus [8], especially in nutrient-poor ecosystems [9]. Acidobacteria, Actinobacteria, Bacteroidetes, and Proteobacteria are the most abundant bacterial producers of carbohydrate-active enzymes involved in the C cycle [39]. In particular, members of Acidobacteria and Bacteroidetes appear to be potent degraders of biopolymers of plant and fungal origin in forest topsoil [10,40]. Our results also show that the application of BCR, BCS, WW, and S increased C, N, and P content, and Bacteroidetes responded most strongly to the addition of organic matter (Figure 3).
Boreal forests are generally considered nitrogen-limited areas [41]. Anaerobic ammonium oxidation (anammox) is a bioprocess in which NH is oxidized to N gas using nitrite (NO) as an electron acceptor. Anammox bacteria include six candidate genera, i.e., Candidatus anammoxoglobus, Ca. anammoximicrobium, Ca. brocadia, Ca. jettenia, Ca. kuenenia and Ca. scalindua, which belong to Planctomycetes [42,43]. Candidatus brocadia prefers environments with high NH and NO due to its low affinity for NH and NO [44]. In our study, Ca. brocadia and Ca. scalindua were found in all soil examined while Ca. anammoxoglobus was only found in the control soil plants. The other anammox bacterial genera were not detected.
Considering that Bacillus spp. were found mainly in the upper soil layer, we assume that they supply phosphorus to plants from the decomposition of organic matter, which stimulates plant growth. Plant growth-promoting bacteria (PGPB) are the target of intensive research studies into their efficient use as biofertilizers, biocontrol agents, and bioremediation agents, as well as numerous other applications [45]. There are examples of bacteria (Acinetobacter, Enterobacter, Providencia, and Pseudomonas) that help survive temperature changes, acidification, and mineral salt uptake [46]. We suspect that Bacillus spp. are also mainly responsible for the decomposition of organic residues. The decomposition processes take place under aerobic conditions in the superficial zones where the root systems of the trees are located. Actinobacteria are also involved in the decomposition of organic material, especially polysaccharides (such as cellulose, chitin) and fats [47]. Nitrogen in the soil stimulates the activity of soil microorganisms (bacteria of the genus Nitrosomonas), especially when the soil temperature rises and reaches or exceeds 10 degrees Celsius. The nitrification process, which leads to the complete conversion of the ammonium form into nitrate, can take only two to four weeks at higher temperatures.
We recognize that soil moisture measurements depend on many external factors, including rainfall. However, by comparing different mulches with controls, it is possible to determine whether the organic matter is helping to protect the soil surface from excessive evaporation. We observed a difference in soil density and moisture content, the latter being inversely proportional and dependent on porosity. The differences resulting from the change in porosity of the surface layers provide better infiltration and less evaporation of water after the treatments. In the first post-treatment period, neighboring observation plots differed significantly in surface color, so there was a temperature difference that led to different evaporation. After the treatments, the resulting lighter soil structure (lower density) led to more favorable porosity, so that soil air can be more easily replaced by rainwater and retained in the soil. This process of storing water in the mulch and protecting the soil from evaporation is crucial for the ability of plants to survive in dry periods compared to the control [2,3].
5.3. Role of the Microbiome in Nutrient Cycling in Soil Systems
The abundance of Bacteroidetes, Firmicutes, and Proteobacteria (Figure 3) is consistent with the bacterial communities in the organic horizon of coniferous forest soils being dominated by these taxa [48,49]. Despite extensive studies on microbial inoculations used for soil biofertilization, their effects on soil microbial community composition and functions remain poorly understood. Compost habitats harbor a large ensemble of microbes specialized in the degradation of lignocellulosic plant materials [50].
Several cellulose, hemicellulose, and lignin cellulose-degrading bacteria were isolated from compost [51]. Bacteroidetes and Chloroflexi probably played a crucial role in the degradation of hemicellulose and were specialized in the hydrolysis of their glycosidic bonds under the thermophilic conditions of the compost habitat. Our data show that Bacteroidetes are abundant taxa (Figure 3). The degradation of hemicellulose polymers at the mesophilic fermentation platform by Bacteroidetes is also confirmed [52].
Relatively low levels of Acinetobacteria, which are normally abundant in compost, were found in the bacterial communities studied. This could be due to the fact that we used bark compost in our study, whereas in the study by Zhang et al. [53] plant material served as the starting material. Members of the genus Firmicutes were most abundant in composts containing nitrogen-rich feedstock, such as animal manure and kitchen waste [54]. Lignin can be depolymerized by lignin-oxidizing enzymes produced by members of the Burkholderiaceae, Comamonadaceae, and Oxalobacteraceae families [55].
The highly active cellulolytic populations in the organic layer were clearly dominated by bacteria, in contrast to those from other regions and soil layers. These active cellulolytic populations consisted largely of Deltaproteobacteria, Gammaproteobacteria, and Planctomyces [56]. Enrichment of the soil with compost led to the appearance of cellulolytic Halocella [57,58].
Phosphorus is one of the most important elements for plant growth and nutrient cycling in soil systems. Phosphorus-solubilizing bacteria (PSB) can improve plant growth and phosphorus dissolution. It has also been reported that they can reduce the negative effects of excessive fertilizer use in agriculture and protect the soil environment [59]. The phosphate-solubilizing genera of Falsibacillus were found in all organic matter-enriched soils in Bielsk and correlated with increased soil organic matter content P2O5 (Figure 9) [60,61,62,63,64,65].
5.4. Treatments of Agricultural Soil with Organic Matter Bring It Closer to Forest Soil
Forest soils are among the most diverse microbial habitats on Earth, where bacteria are the most abundant group of microorganisms [40]. Forest soils are characterized by strong vertical stratification due to the decomposition of organic matter from litter and weathering of the mineral matrix. The decreasing content and quality of organic matter with soil depth is accompanied by a decrease in microbial biomass, respiration, and extracellular enzyme activity. The enrichment of soils with organic matter after agricultural use is necessary because of the lack of organic matter, including the lignin and hemicellulose tissues that form the structure of wood, and the roots, which are the basic substrate and energy source for the development of microorganisms, including fungi, especially saprotrophic fungi that degrade the cellulose-lignin complex [66].
Fungi, bacteria, and other microorganisms involved in the processes of creating the specific structure of forest soil give it the specific biological activity we hoped for in our study. One consequence of the special conditions of forest stands growing on formerly agricultural land is their susceptibility to biotic stress factors [67]. Mycological studies of cultivated soils have shown that they are particularly poor in fungi with antagonistic abilities against five pathogens of tree root diseases [35]. The structure and proportion of mycorrhizal symbionts in the soils after cultivation in the first years after the seedlings are established in the stand are also of crucial importance for the condition of future stands [68]. In the juvenile phase of forest tree life, great importance is attached to the creation of favorable conditions for the establishment of mycorrhizal symbiosis [69].
The process of transformation of the mycorrhizal fungal community (already present in the soils after agricultural use) into a composition that is typical of the forest environment (ectomycorrhizal fungi—ECM) has not yet been precisely timed. The study by [70] (1996) shows that an ECM community typical for a mature stand can be established on agricultural soils at the earliest 25–30 years after the end of agricultural use. Suitable bacteria (helpers) favor the formation of mycorrhizal communities. However, the succession of ECM communities in conifers planted on agricultural land or left to natural afforestation is still poorly understood. Previous research in this area has focused on the effects of tillage [71] and nitrogen fertilization [72] on changes in mycorrhizal structure, and the inoculation of pine and spruce seedlings with selected ECM fungal species [73,74,75].
There is still a lack of information on the changes in the bacterium, which is supplemented by the present study. The studies conducted in Bielsk FD (Regional Directorate of State Forests—RDSF Białystok) and in Czarne Człuchowskie Forest Inspectorate (RDSF Szczecinek) provide results describing changes in the mycobiome, [2,3]. The comparison of the studied microbiological parameters with literature data [76,77] on managed forests shows that the composition of the soil microbial community in cultures is not yet fully stabilized. The present study is a continuation of investigations on plots established in 2001.
6. Conclusions
- Bacterial diversity is lower in poorer habitats than in richer habitats. In the latter, the addition of organic material in the form of sawdust further increases bacterial diversity, which is not observed when compost and wood residues collected after felling are used.
- On poorer post-agricultural land, the application of organic matter does not lead to an increase in bacterial biodiversity in terms of the number of taxa, but a change in quality that potentially creates better-growing conditions for pines, which in turn increases their chances of survival in the first generation on post-agricultural land.
- The amount of Proteobacteria and Firmicutes, followed by Bacteroides, increases significantly after the application of sawdust over the entire area in the richer fresh forest habitat.
- The poorer habitat tends to be represented by fewer Bacteroidetes genera, while the addition of bark compost leads to significant changes in bacteria. New genera of Firmicutes appear, while other Firmicutes, Proteobacteria, and Bacteroidetes disappear.
- Restoring organic content not only improves the biological life of bacteria, especially in richer soils formerly used for agriculture, but also has a positive effect on physicochemical conditions of former agricultural soils.
- The addition of organic matter to the soil restores the damaged organic horizon, and favorably increases the proportion of carbon (C), nitrogen (N), and phosphorus (P), in the organic layer and in the reach of the finest roots of the trees.
- The use of organic material also increases the sum of primary exchangeable cations (Ca, Mg, K) and the sorption capacity of the soil, thus increasing the binding capacity of exchangeable ions in the soil.
- The addition of pine bark compost, sawdust, or wood waste leads to positive changes in the physical and physicochemical properties of the soil. Revitalizing soils with organic matter improves their moisture conditions, which not only facilitates the growth of beneficial bacteria but also nutrient uptake by the pines.
- The weakest post-agricultural soils especially need to be revitalized with organic matter, as they lack the humus typical of forest soils and are unable to provide the constant supply of nutrients necessary for the good development of the pine plantations established on them.
Author Contributions
Conceptualization, T.O. and I.O.; methodology, T.M.; software, P.B.; validation, A.O. and P.G.; formal analysis, T.M.; investigation, I.O. and T.O.; resources, T.O.; data curation, P.B., T.M. and A.O.; writing—original draft preparation, T.M., T.O., P.B. and P.G.; writing—review and editing, P.B., T.O., A.O. and P.G.; visualization, P.B. and T.M.; supervision, T.O.; project administration, I.O.; funding acquisition, T.O. All authors have read and agreed to the published version of the manuscript.
Funding
The results presented in this paper were obtained as part of a comprehensive study financed by the University of Warmia and Mazury in Olsztyn (grant no. 30.610.009.110).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sun, W.; Li, Z.; Lei, J.; Liu, X. Bacterial Communities of Forest Soils along Different Elevations: Diversity, Structure, and Functional Composition with Potential Impacts on CO2 Emission. Microorganisms 2022, 10, 766. [Google Scholar] [CrossRef] [PubMed]
- Malewski, T.; Borowik, P.; Olejarski, I.; Rutkiewicz, A.; Okorski, A.; Oszako, T. Addition of Organic Matter to Pine Plantations on Agricultural Land Positively Alters the Mycobiome of Agricultural Soils. Appl. Sci. 2023, 13, 5800. [Google Scholar] [CrossRef]
- Malewski, T.; Borowik, P.; Olejarski, I.; Berezovska, D.; Dyshko, V.; Behnke-Borowczyk, J.; Pusz, W.; Matic, S.; Oszako, T. Mycobiome of Post-Agricultural Soils 20 Years after Application of Organic Substrates and Planting of Pine Seedlings. Forests 2022, 14, 36. [Google Scholar] [CrossRef]
- Krueger, I.; Schulz, C.; Borken, W. Stocks and dynamics of soil organic carbon and coarse woody debris in three managed and unmanaged temperate forests. Eur. J. For. Res. 2017, 136, 123–137. [Google Scholar] [CrossRef]
- Adedeji, A.A.; Häggblom, M.M.; Babalola, O.O. Sustainable agriculture in Africa: Plant growth-promoting rhizobacteria (PGPR) to the rescue. Sci. Afr. 2020, 9, e00492. [Google Scholar] [CrossRef]
- Basu, A.; Prasad, P.; Das, S.N.; Kalam, S.; Sayyed, R.; Reddy, M.; El Enshasy, H. Plant growth promoting rhizobacteria (PGPR) as green bioinoculants: Recent developments, constraints, and prospects. Sustainability 2021, 13, 1140. [Google Scholar] [CrossRef]
- Nazari, M.; Smith, D.L. A PGPR-produced bacteriocin for sustainable agriculture: A review of thuricin 17 characteristics and applications. Front. Plant Sci. 2020, 11, 916. [Google Scholar] [CrossRef]
- Van Der Heijden, M.G.; Bardgett, R.D.; Van Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef]
- Franklin, O.; Näsholm, T.; Högberg, P.; Högberg, M.N. Forests trapped in nitrogen limitation–an ecological market perspective on ectomycorrhizal symbiosis. New Phytol. 2014, 203, 657–666. [Google Scholar] [CrossRef]
- Lladó, S.; López-Mondéjar, R.; Baldrian, P. Drivers of microbial community structure in forest soils. Appl. Microbiol. Biotechnol. 2018, 102, 4331–4338. [Google Scholar] [CrossRef]
- Boyarshin, K.S.; Adamova, V.V.; Wentao, Z.; Obuhova, O.Y.; Kolkova, M.V.; Nesterenko, V.A.; Bespalova, O.S.; Kluyeva, V.V.; Degtyareva, K.A.; Kurkina, Y.N.; et al. The Effect of Long-Term Agricultural Use on the Bacterial Microbiota of Chernozems of the Forest-Steppe Zone. Diversity 2023, 15, 191. [Google Scholar] [CrossRef]
- Fierer, N.; Colman, B.P.; Schimel, J.P.; Jackson, R.B. Predicting the temperature dependence of microbial respiration in soil: A continental-scale analysis. Glob. Biogeochem. Cycles 2006, 20, 3. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, X.; Zhang, M. Functional diversity changes of microbial communities along a soil aquifer for reclaimed water recharge. Fems Microbiol. Ecol. 2012, 80, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.G.; Huang, Z.L.; Cheng, R.M.; Zeng, L.X.; Xiao, W.F.; Tan, B.W. Effects of litterfall and root input on soil physical and chemical properties in Pinus massoniana plantations in Three Gorges Reservoir Area, China. Ying Yong Sheng Tai Xue Bao J. Appl. Ecol. 2012, 23, 3301–3308. [Google Scholar]
- Ze-yan, W.; Wen-xiong, L.; Zhi-fang, C.; Chang-xun, F.; Zhi-xing, Z.; Lin-kun, W.; Ming-ming, Z.; Li-hua, S. Characteristics of soil microbial community under different vegetation types in Wuyishan National Nature Reserve, East China. Yingyong Shengtai Xuebao 2013, 24, 2301–2309. [Google Scholar]
- Kaiser, K.; Wemheuer, B.; Korolkow, V.; Wemheuer, F.; Nacke, H.; Schöning, I.; Schrumpf, M.; Daniel, R. Driving forces of soil bacterial community structure, diversity, and function in temperate grasslands and forests. Sci. Rep. 2016, 6, 33696. [Google Scholar] [CrossRef] [PubMed]
- Weng, X.; Li, J.; Sui, X.; Li, M.; Yin, W.; Ma, W.; Yang, L.; Mu, L. Soil microbial functional diversity responses to different vegetation types in the Heilongjiang Zhongyangzhan Black-billed Capercaillie Nature Reserve. Ann. Microbiol. 2021, 71, 26. [Google Scholar] [CrossRef]
- Li, L.; Xu, M.; Eyakub Ali, M.; Zhang, W.; Duan, Y.; Li, D. Factors affecting soil microbial biomass and functional diversity with the application of organic amendments in three contrasting cropland soils during a field experiment. PLoS ONE 2018, 13, e0203812. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Bushnell, B.; Rood, J.; Singer, E. BBMerge – Accurate paired shotgun read merging via overlap. PLoS ONE 2017, 12, e0185056. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- The Pandas Development Team. Pandas-Dev/Pandas: Pandas. 2020. Available online: https://zenodo.org/record/8239932 (accessed on 10 June 2023).
- Hunter, J.D. Matplotlib: A 2D graphics environment. Comput. Sci. Eng. 2007, 9, 90–95. [Google Scholar] [CrossRef]
- Waskom, M.L. Seaborn: Statistical data visualization. J. Open Source Softw. 2021, 6, 3021. [Google Scholar] [CrossRef]
- Virtanen, P.; Gommers, R.; Oliphant, T.E.; Haberland, M.; Reddy, T.; Cournapeau, D.; Burovski, E.; Peterson, P.; Weckesser, W.; Bright, J.; et al. SciPy 1.0: Fundamental Algorithms for Scientific Computing in Python. Nat. Methods 2020, 17, 261–272. [Google Scholar] [CrossRef]
- Scikit-bio Phython Package. Available online: http://scikit-bio.org/docs/0.5.8/ (accessed on 15 December 2022).
- Addison, S.; Smaill, S.; Garrett, L.; Wakelin, S. Effects of forest harvest and fertiliser amendment on soil biodiversity and function can persist for decades. Soil Biol. Biochem. 2019, 135, 194–205. [Google Scholar] [CrossRef]
- Sridhar, B.; Lawrence, G.B.; Debenport, S.J.; Fahey, T.J.; Buckley, D.H.; Wilhelm, R.C.; Goodale, C.L. Watershed-scale liming reveals the short- and long-term effects of pH on the forest soil microbiome and carbon cycling. Environ. Microbiol. 2022, 24, 6184–6199. [Google Scholar] [CrossRef]
- Bell-Doyon, P.; Bellavance, V.; Bélanger, L.; Mazerolle, M.J.; Villarreal A., J.C. Bacterial, fungal, and mycorrhizal communities in the soil differ between clearcuts and insect outbreaks in the boreal forest 50 years after disturbance. For. Ecol. Manag. 2022, 523, 120493. [Google Scholar] [CrossRef]
- Addison, S.; Smaill, S.; Garrett, L.; Wakelin, S. Fertiliser use has multi-decadal effects on microbial diversity and functionality of forest soils. Appl. Soil Ecol. 2021, 163, 103964. [Google Scholar] [CrossRef]
- Sobczak, R. Teoretyczne i praktyczne aspekty zakładania upraw i prowadzenia drzewostanów na gruntach porolnych. Sylwan 1990, 134, 3–12. [Google Scholar]
- Gorzelak, A. Rola substancji organicznych w podnoszeniu produkcyjności wydm oraz słabych gruntów porolnych. Sylwan 1998, 142, 27–33. [Google Scholar]
- Sierota, Z.; Kwasna, H. Ocena mikologiczna zmian zachodzących w glebie gruntu porolnego po dodaniu trocin iglastych. Sylwan 1999, 143, 57–66. [Google Scholar]
- Olejarski, I.; Walendzik, J. Warunki wzrostu upraw leśnych w Bornem Sulinowie. Głos Lasu 2002, 8, 20–22. [Google Scholar]
- Olejarski, I. Wykorzystanie pozostałości zrębowych do nawożenia organicznego gruntów. Postępy Tech. Leśnictwie 2005, 92, 20–24. [Google Scholar]
- Małecka, M.; Hilszczańska, D. Changes in ectomycorrhizal structure of Scots pine growing on abandoned farmland soil enriched with organic substrates. Sylwan 2014, 158, 243–250. [Google Scholar]
- Žifčáková, L.; Větrovskỳ, T.; Lombard, V.; Henrissat, B.; Howe, A.; Baldrian, P. Feed in summer, rest in winter: Microbial carbon utilization in forest topsoil. Microbiome 2017, 5, 122. [Google Scholar] [CrossRef]
- Lladó, S.; López-Mondéjar, R.; Baldrian, P. Forest Soil Bacteria: Diversity, Involvement in Ecosystem Processes, and Response to Global Change. Microbiol. Mol. Biol. Rev. 2017, 81, 2. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Xu, X. Competition between roots and microorganisms for nitrogen: Mechanisms and ecological relevance. New Phytol. 2013, 198, 656–669. [Google Scholar] [CrossRef]
- Oshiki, M.; Satoh, H.; Okabe, S. Ecology and physiology of anaerobic ammonium oxidizing bacteria. Environ. Microbiol. 2016, 18, 2784–2796. [Google Scholar] [CrossRef]
- Zhang, L.; Okabe, S. Ecological niche differentiation among anammox bacteria. Water Res. 2020, 171, 115468. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Wei, N.; Wu, G. Autotrophic nitrogen removal and potential microbial interactions in anammox systems with different ammonia and organic carbon concentrations. J. Water Process. Eng. 2020, 37, 101493. [Google Scholar] [CrossRef]
- Cherif-Silini, H.; Silini, A.; Chenari Bouket, A.; Alenezi, F.N.; Luptakova, L.; Bouremani, N.; Nowakowska, J.A.; Oszako, T.; Belbahri, L. Tailoring next generation plant growth promoting microorganisms as versatile tools beyond soil desalinization: A road map towards field application. Sustainability 2021, 13, 4422. [Google Scholar] [CrossRef]
- Rabhi, N.E.H.; Cherif-Silini, H.; Silini, A.; Alenezi, F.N.; Chenari Bouket, A.; Oszako, T.; Belbahri, L. Alleviation of Salt Stress via Habitat-Adapted Symbiosis. Forests 2022, 13, 586. [Google Scholar] [CrossRef]
- Urry, L.A.; Meyers, N.; Cain, M.L.; Wasserman, S.A.; Minorsky, P.V.; Orr, R.; Da Silva, K.B.; Parkinson, A.; Lluka, L.; Chunduri, P.; et al. Campbell Biology: Australian and New Zealand Version; Pearson Australia: Melbourne, Australia, 2021. [Google Scholar]
- Baldrian, P.; Kolařík, M.; Štursová, M.; Kopecký, J.; Valášková, V.; Větrovský, T.; Žifčáková, L.; Šnajdr, J.; Rídl, J.; Vlček, Č.; et al. Active and total microbial communities in forest soil are largely different and highly stratified during decomposition. ISME J. 2011, 6, 248–258. [Google Scholar] [CrossRef]
- Uroz, S.; Ioannidis, P.; Lengelle, J.; Cébron, A.; Morin, E.; Buée, M.; Martin, F. Functional Assays and Metagenomic Analyses Reveals Differences between the Microbial Communities Inhabiting the Soil Horizons of a Norway Spruce Plantation. PLoS ONE 2013, 8, e55929. [Google Scholar] [CrossRef]
- Wang, C.; Dong, D.; Wang, H.; Müller, K.; Qin, Y.; Wang, H.; Wu, W. Metagenomic analysis of microbial consortia enriched from compost: New insights into the role of Actinobacteria in lignocellulose decomposition. Biotechnol. Biofuels 2016, 9, 22. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Guo, X.; Deng, H.; Dong, D.; Tu, Q.; Wu, W. New insights into the structure and dynamics of actinomycetal community during manure composting. Appl. Microbiol. Biotechnol. 2013, 98, 3327–3337. [Google Scholar] [CrossRef]
- Hollister, E.B.; Forrest, A.K.; Wilkinson, H.H.; Ebbole, D.J.; Tringe, S.G.; Malfatti, S.A.; Holtzapple, M.T.; Gentry, T.J. Mesophilic and thermophilic conditions select for unique but highly parallel microbial communities to perform carboxylate platform biomass conversion. PLoS ONE 2012, 7, e39689. [Google Scholar] [CrossRef]
- Zhang, X.; Zhong, Y.; Yang, S.; Zhang, W.; Xu, M.; Ma, A.; Zhuang, G.; Chen, G.; Liu, W. Diversity and dynamics of the microbial community on decomposing wheat straw during mushroom compost production. Bioresour. Technol. 2014, 170, 183–195. [Google Scholar] [CrossRef]
- Zhou, G.; Xu, X.; Qiu, X.; Zhang, J. Biochar influences the succession of microbial communities and the metabolic functions during rice straw composting with pig manure. Bioresour. Technol. 2019, 272, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Bayer, E.A.; Shoham, Y.; Lamed, R. Lignocellulose-Decomposing Bacteria and Their Enzyme Systems. In The Prokaryotes; Springer: Berlin/Heidelberg, Germany, 2013; pp. 215–266. [Google Scholar] [CrossRef]
- Wilhelm, R.C.; Singh, R.; Eltis, L.D.; Mohn, W.W. Bacterial contributions to delignification and lignocellulose degradation in forest soils with metagenomic and quantitative stable isotope probing. ISME J. 2018, 13, 413–429. [Google Scholar] [CrossRef] [PubMed]
- Bolobova, A.; Simankova, M.; Markovich, N. Cellulase complex of a new halophilic bacterium halocella-cellulolytica. Microbiology 1992, 61, 557–562. [Google Scholar]
- Akhtar, N.; Aanchal; Goyal, D.; Goyal, A. Biodiversity of cellulase producing bacteria and their applications. Cellul. Chem. Technol. 2016, 50, 983–995. [Google Scholar]
- Wang, Z.; Zhang, H.; Liu, L.; Li, S.; Xie, J.; Xue, X.; Jiang, Y. Screening of phosphate-solubilizing bacteria and their abilities of phosphorus solubilization and wheat growth promotion. Bmc Microbiol. 2022, 22, 296. [Google Scholar] [CrossRef]
- Bernacki, E. Koncepcje hodowli lasu na gruntach porolnych. Sylwan 1990, 134, 3–12. [Google Scholar]
- Szujecki, A. Ekologiczne aspekty odtwarzania ekosystemów leśnych na gruntach porolnych. Sylwan 1990, 134, 3–12. [Google Scholar]
- Hytönen, J.; Ekola, E. Maan ja puuston ravinnetila Keski-Pohjanmaan metsitetyillä pelloilla. Summary: Soil nutrient regime and tree nutrition on afforested fields in central Ostrobothnia, western Finland. Folia For. 1993, 822, 32. [Google Scholar]
- Mazur, S. Znaczenie zabiegów fito-i zoomelioracyjnych w biologicznym uodparnianiu drzewostanów na gruntach porolnych. Biblioteczka Leśniczego 2003, 176, 1–9. [Google Scholar]
- Olejarski, I.; Walendzik, R. Wspieranie upraw w warunkach zagrożenia. Głos Lasu 2001, 7, 10–11. [Google Scholar]
- Oszako, T.; Olejarski, I. Inicjowanie procesów przekształcenia gleb porolnych w gleby leśne poprzez wykorzystanie pozostałości zrębowych, kompostów i trocin. Pr. Inst. Badaw. Leśnictwa. Ser. A 2003, 1 [948–951], 76–79. [Google Scholar]
- Małecka, M.; Hilszczańska, D. Influence of resting and pine sawdust application on chemical changes in post-agricultural soil and the ectomycorrhizal community of growing Scots pine saplings. Forest Res. Papers 2015, 76, 15272. [Google Scholar] [CrossRef]
- Małecka, M.; Sierota, Z. Threat and risk assessment of root and butt rot disease in a stand on post-agricultural land. Sylwan 2003, 147, 12–25. [Google Scholar]
- Hilszczańska, D.; Sierota, Z. The role of Thelephora terrestris fungus in mycorrhization on Scots pine (Pinus sylvestris L.) seedlings. II. Field study. Sylwan 2006, 150, 20–28. [Google Scholar]
- Hilszczańska, D. Wpływ podłoży szkólkarskich na rozwój mikoryz sosny Pinus sylvestris L. Sylwan 2000, 144, 93–97. [Google Scholar]
- Boerner, R.E.J.; DeMars, B.G.; Leicht, P.N. Spatial patterns of mycorrhizal infectiveness of soils long a successional chronosequence. Mycorrhiza 1996, 6, 79–90. [Google Scholar] [CrossRef]
- Aleksandrowicz-Trzcińska, M.; Sikora, K.; Drozdowski, S.; Nowakowska, J. Ectomycorrhizal fungal assemblages in 35-years-old Scots pine stand growing on former agricultural land. Sylwan 2021, 165, 50–60. [Google Scholar] [CrossRef]
- Hilszczańska, D.; Małecka, M.; Sierota, Z. Changes in nitrogen level and mycorrhizal structure of Scots pine seedlings inoculated with Thelephora terrestris. Ann. For. Sci. 2008, 65, 1. [Google Scholar] [CrossRef]
- Hilszczańska, D. Influence of watering of Pinus sylvestris L. seedlings on mycorrhisal and soil fungi. For. Res. Pap. 2005, 4, 103–113. [Google Scholar]
- Menkis, A.; Vasiliauskas, R.; Taylor, A.F.S.; Stenlid, J.; Finlay, R. Fungal communities in mycorrhizal roots of conifer seedlings in forest nurseries under different cultivation systems, assessed by morphotyping, direct sequencing and mycelial isolation. Mycorrhiza 2005, 16, 33–41. [Google Scholar] [CrossRef]
- Sierota, Z.; Hilszczańska, D. Ectomycorrhizal structure and biometric parameters of pine after planting on post-agricultural land. Sylwan 2009, 153, 108–116. [Google Scholar]
- Olszowska, G.; Zwoliński, J.; Matuszczyk, I.; Syrek, D.; Zwolińska, B.; Pawlak, U.; Kwapis, Z.; Dudzińska, M. The use of biological activity studies to determine a soil fertility indicator in pine stands on fresh coniferous an on mixed fresh coniferous forest sites. For. Res. Pap. 2005, 3, 17–37. [Google Scholar]
- Olszowska, G.; Zwoliński, J.; Matuszczyk, I.; Syrek, D. Application of biochemical soil parameters in typological diagnosis of forest sites. For. Res. Pap. 2007, 4, 83–105. [Google Scholar]
Figure 1.
Map of Poland with marked locations of the experimental plots in FD Bielsk and Czarne Człuchowskie.
Figure 1.
Map of Poland with marked locations of the experimental plots in FD Bielsk and Czarne Człuchowskie.

Figure 2.
Photos of the trial plots with treatments applied to the soil surface. On the left—Bielsk, and on the right—Czarne Człuchowskie. (a,b) wood waste; (c,d) sawdust; (e,f) bark compost.
Figure 2.
Photos of the trial plots with treatments applied to the soil surface. On the left—Bielsk, and on the right—Czarne Człuchowskie. (a,b) wood waste; (c,d) sawdust; (e,f) bark compost.

Figure 3.
The number of bacterial genera detected in relation to the studied soil variants: control (C), bark compost under the roots (BCR), bark compost on the surface (BCS), wood waste (WW), and sawdust (S).
Figure 3.
The number of bacterial genera detected in relation to the studied soil variants: control (C), bark compost under the roots (BCR), bark compost on the surface (BCS), wood waste (WW), and sawdust (S).

Figure 4.
The number of bacterial genera in relation to soil management: control (C), bark compost under the roots (BCR), bark compost on the surface (BCS), wood waste (WW), and sawdust (S).
Figure 4.
The number of bacterial genera in relation to soil management: control (C), bark compost under the roots (BCR), bark compost on the surface (BCS), wood waste (WW), and sawdust (S).

Figure 5.
Venn diagrams showing the number of common bacterial genera between the studied soil variants at the Bielsk and Czarne sites: control (C), bark compost under the roots (BCR), bark compost on the surface (BCS), wood waste (WW), and sawdust (S).
Figure 5.
Venn diagrams showing the number of common bacterial genera between the studied soil variants at the Bielsk and Czarne sites: control (C), bark compost under the roots (BCR), bark compost on the surface (BCS), wood waste (WW), and sawdust (S).

Figure 6.
Similarity index according to the Jaccard formula for bacterial genera, comparison of similarities between different studied soil variants: control (C), bark compost under the roots (BCR), bark compost on the surface (BCS), wood waste (WW), and sawdust (S). Lower figures—hierarchical grouping of the studied plots according to the similarity index using the inverse of Jaccard’s formula for bacterial genera.
Figure 6.
Similarity index according to the Jaccard formula for bacterial genera, comparison of similarities between different studied soil variants: control (C), bark compost under the roots (BCR), bark compost on the surface (BCS), wood waste (WW), and sawdust (S). Lower figures—hierarchical grouping of the studied plots according to the similarity index using the inverse of Jaccard’s formula for bacterial genera.

Figure 7.
Soil carbon content at the organic level and in mineral layers (0-20 cm and 20-40 cm) compared to different studied soil variants: control (C), bark compost under the roots (BCR), bark compost on the surface (BCS), wood waste (WW), and sawdust (S). The geographical locations of the plots are indicated above the sub-figures.
Figure 7.
Soil carbon content at the organic level and in mineral layers (0-20 cm and 20-40 cm) compared to different studied soil variants: control (C), bark compost under the roots (BCR), bark compost on the surface (BCS), wood waste (WW), and sawdust (S). The geographical locations of the plots are indicated above the sub-figures.

Figure 8.
Nitrogen content in soil at the organic level and in the mineral layers (0–20 cm and 20–40 cm) compared to different studied soil variants: control (C), bark compost under the roots (BCR), bark compost on the surface (BCS), wood waste (WW), and sawdust (S). The geographical locations of the plots are indicated above sub-figures.
Figure 8.
Nitrogen content in soil at the organic level and in the mineral layers (0–20 cm and 20–40 cm) compared to different studied soil variants: control (C), bark compost under the roots (BCR), bark compost on the surface (BCS), wood waste (WW), and sawdust (S). The geographical locations of the plots are indicated above sub-figures.

Figure 9.
P2O5 content in soil at the organic level and in mineral layers (0–20 cm and 20–40 cm) compared to different studied soil variants: control (C), bark compost under the roots (BCR), bark compost on the surface (BCS), wood waste (WW), and sawdust (S). The geographical locations of the plots are indicated above sub-figures.
Figure 9.
P2O5 content in soil at the organic level and in mineral layers (0–20 cm and 20–40 cm) compared to different studied soil variants: control (C), bark compost under the roots (BCR), bark compost on the surface (BCS), wood waste (WW), and sawdust (S). The geographical locations of the plots are indicated above sub-figures.

Figure 10.
The sum of alkalis [cmol(+)/kg] in the soil at the organic level and in the mineral layers (0–20 cm and 20–40 cm) compared to the different studied soil variants: control (C), bark compost under the roots (BCR), bark compost on the surface (BCS), wood waste (WW), and sawdust (S). The geographical locations of the plots are indicated in the sub-figures.
Figure 10.
The sum of alkalis [cmol(+)/kg] in the soil at the organic level and in the mineral layers (0–20 cm and 20–40 cm) compared to the different studied soil variants: control (C), bark compost under the roots (BCR), bark compost on the surface (BCS), wood waste (WW), and sawdust (S). The geographical locations of the plots are indicated in the sub-figures.

Figure 11.
Sorption capacity [cmol(+)/kg] in soil at the organic level and in the mineral layers (0–20 and 20–40 cm) compared to different studied soil variants: control (C), bark compost under the roots (BCR), bark compost on the surface (BCS), wood waste (WW), and sawdust (S). The geographical locations of the plots are indicated above sub-figures.
Figure 11.
Sorption capacity [cmol(+)/kg] in soil at the organic level and in the mineral layers (0–20 and 20–40 cm) compared to different studied soil variants: control (C), bark compost under the roots (BCR), bark compost on the surface (BCS), wood waste (WW), and sawdust (S). The geographical locations of the plots are indicated above sub-figures.

Figure 12.
Soil moisture and density in mineral layers (0–20 and 20–40 cm) compared to different studied soil variants: control (C), bark compost under the roots (BCR), bark compost on the surface (BCS), wood waste (WW), and sawdust (S). The geographical locations of the plots are indicated above the sub-figures.
Figure 12.
Soil moisture and density in mineral layers (0–20 and 20–40 cm) compared to different studied soil variants: control (C), bark compost under the roots (BCR), bark compost on the surface (BCS), wood waste (WW), and sawdust (S). The geographical locations of the plots are indicated above the sub-figures.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Measured carbon and nitrogen content in soil at organic and mineral levels (0–20 cm and 20–40 cm), compared with different studied soil variants: control (C), bark compost under the roots (BCR), bark compost on the surface (BCS), wood waste (WW), and sawdust (S). The geographical locations of the plots are indicated above the sub-tables.
Table 1.
Measured carbon and nitrogen content in soil at organic and mineral levels (0–20 cm and 20–40 cm), compared with different studied soil variants: control (C), bark compost under the roots (BCR), bark compost on the surface (BCS), wood waste (WW), and sawdust (S). The geographical locations of the plots are indicated above the sub-tables.
| Bielsk | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Carbon | Nitrogen | C/N | |||||||
| Org. Lev. | 0–20 cm | 20–40 cm | Org. Lev. | 0–20 cm | 20–40 cm | Org. Lev. | 0–20 cm | 20–40 cm | |
| C | 9.6 | 0.21 | 0.06 | 0.42 | 0.03 | 0.01 | 22.9 | 7.0 | 6.0 |
| BCS | 14.5 | 0.61 | 0.35 | 0.73 | 0.02 | 0.03 | 19.7 | 30.5 | 11.6 |
| BCR | 7.2 | 0.68 | 0.23 | 0.28 | 0.04 | 0.02 | 25.7 | 17,0 | 11.5 |
| S | 15.6 | 0.30 | 0.08 | 1.35 | 0.02 | 0.01 | 11.6 | 15.0 | 8.0 |
| WW | 31.6 | 0.56 | 0.19 | 0.59 | 0.03 | 0.02 | 53.6 | 18.7 | 9.5 |
| Czarne | |||||||||
| Carbon | Nitrogen | C/N | |||||||
| Org. Lev. | 0–20 cm | 20–40 cm | Org. Lev. | 0–20 cm | 20–40 cm | Org. Lev. | 0–20 cm | 20–40 cm | |
| C | 34.9 | 1.06 | 0.77 | 1.20 | 0.06 | 0.04 | 29.1 | 17.7 | 19.3 |
| BCS | 18.0 | 0.69 | 0.97 | 0.61 | 0.04 | 0.05 | 29.5 | 17.3 | 19.4 |
| BCR | 8.2 | 1.03 | 1.04 | 0.30 | 0.06 | 0.06 | 27.3 | 17.2 | 17.3 |
| S | 28.9 | 1.24 | 0.91 | 0.94 | 0.07 | 0.05 | 30.7 | 17.7 | 18.2 |
| WW | 19.7 | 0.74 | 0.72 | 0.69 | 0.04 | 0.04 | 28.6 | 18.5 | 18.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Malewski, T.; Borowik, P.; Golińska, P.; Okorski, A.; Olejarski, I.; Oszako, T. Organic Inputs Positively Alter the Bacteriome of Post-Agricultural Soils. Forests 2023, 14, 1711. https://doi.org/10.3390/f14091711
AMA Style
Malewski T, Borowik P, Golińska P, Okorski A, Olejarski I, Oszako T. Organic Inputs Positively Alter the Bacteriome of Post-Agricultural Soils. Forests. 2023; 14(9):1711. https://doi.org/10.3390/f14091711
Chicago/Turabian StyleMalewski, Tadeusz, Piotr Borowik, Patrycja Golińska, Adam Okorski, Ireneusz Olejarski, and Tomasz Oszako. 2023. "Organic Inputs Positively Alter the Bacteriome of Post-Agricultural Soils" Forests 14, no. 9: 1711. https://doi.org/10.3390/f14091711
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.