
The Effects of Thinning on Tree Growth and Stand Biomass in a Chronosequence of Pinus tabulaeformis Plantations in the Loess Plateau of China

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Field Investigation
2.3. Predictors of Stand Biomass
2.4. Statistical Analyses
3. Results
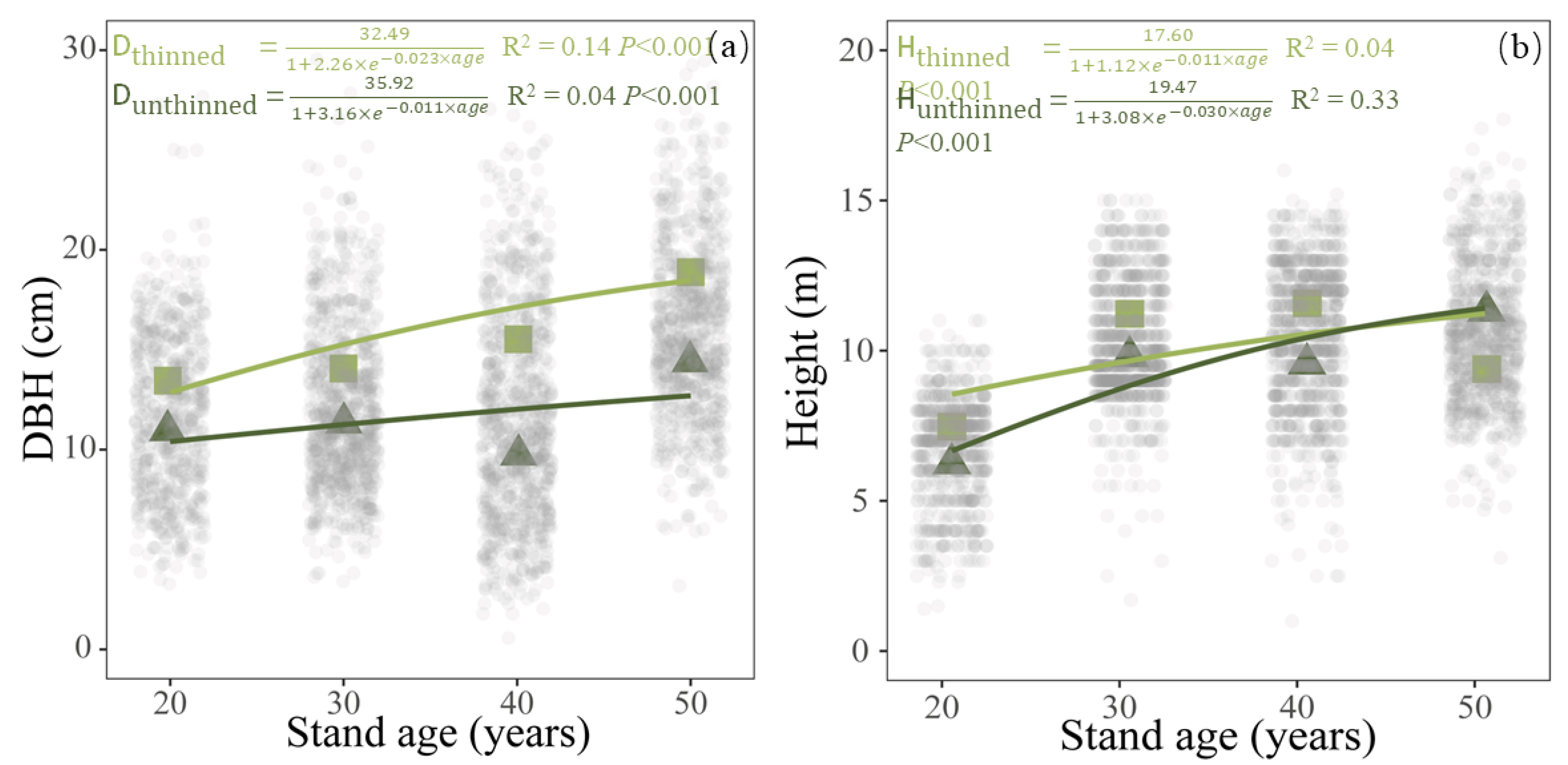
3.1. Individual Tree Growth under Different Thinning Managements
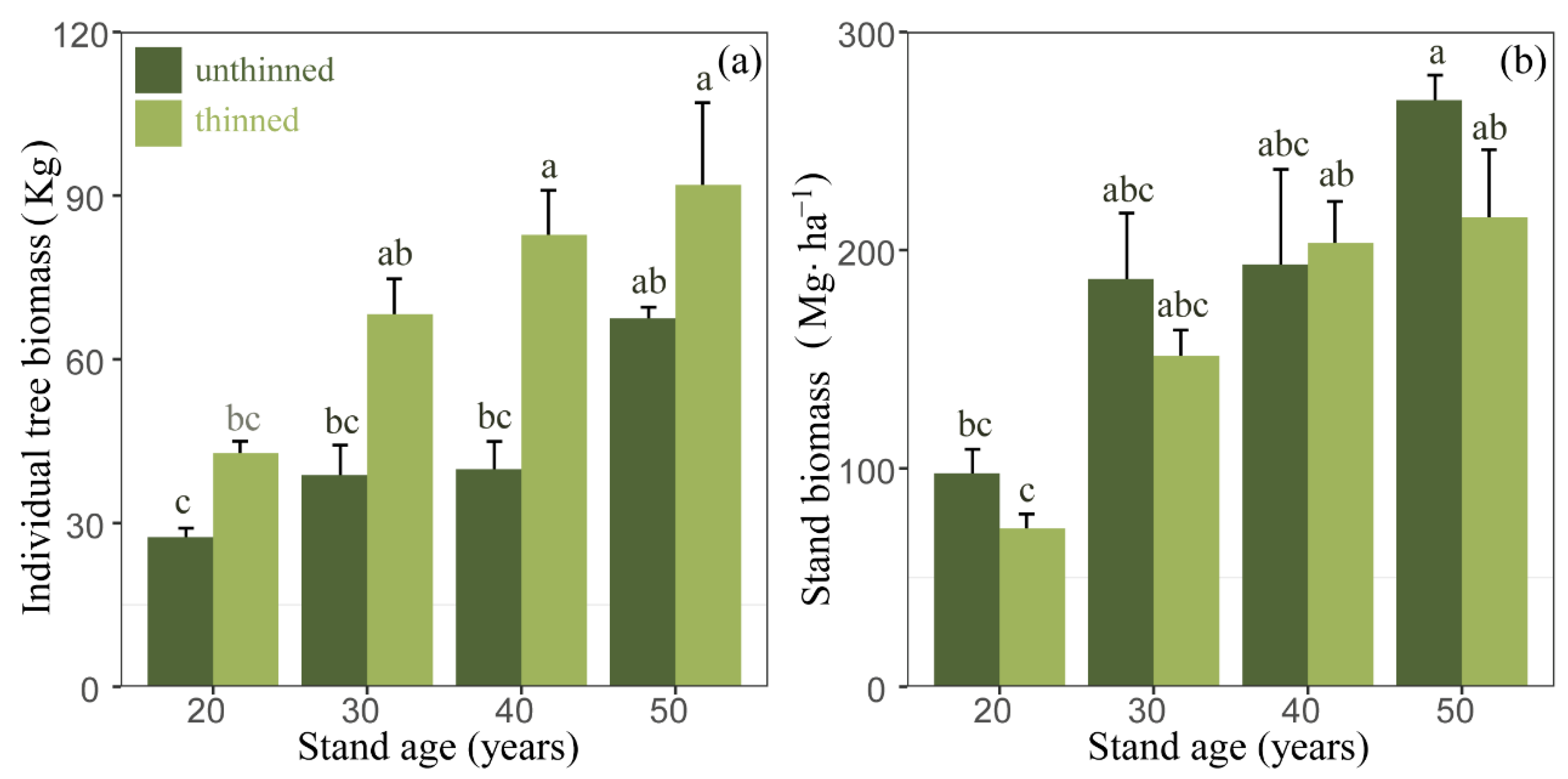
3.2. Individual Tree and Stand Biomass under Different Thinning Managements
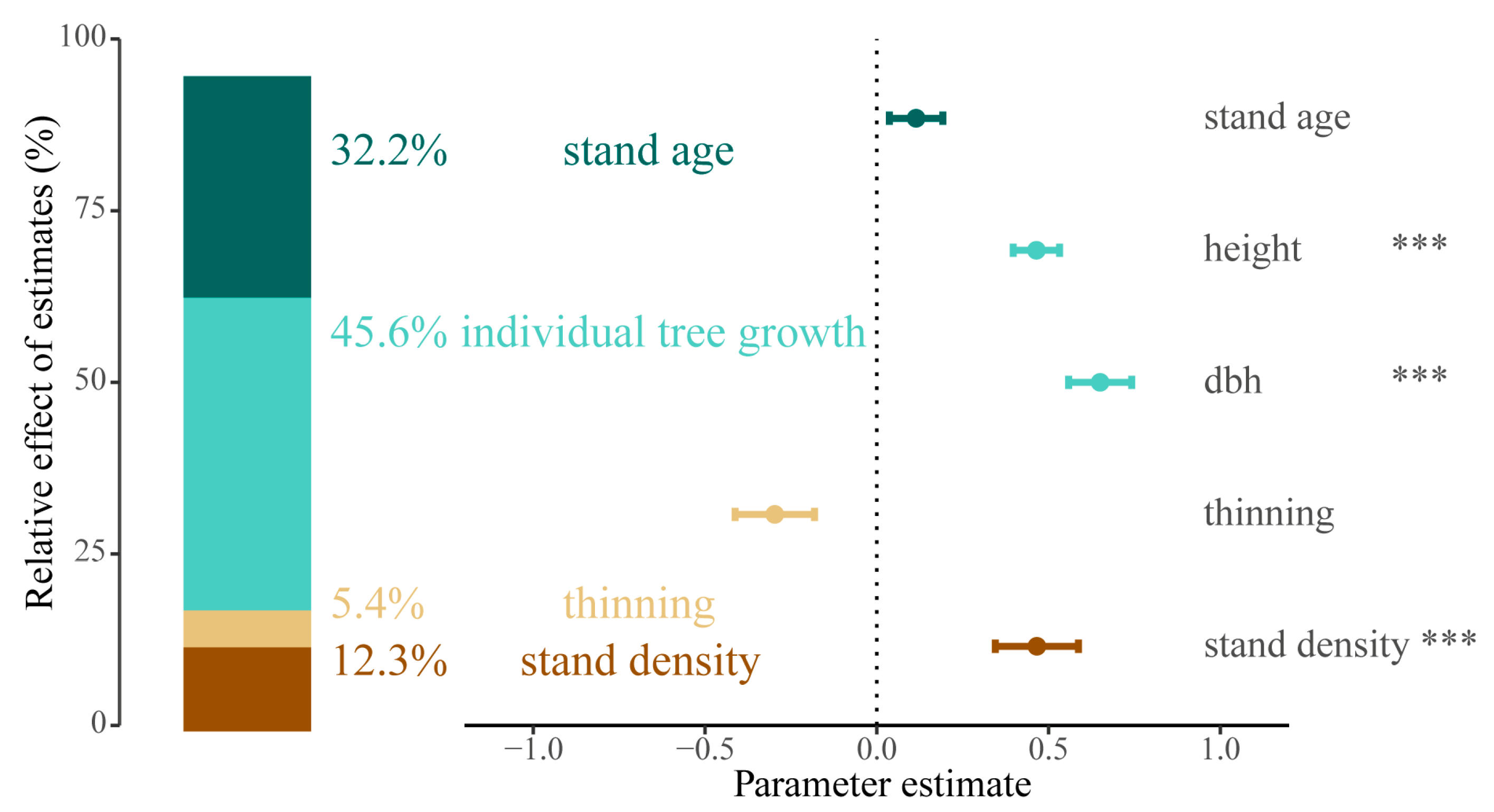
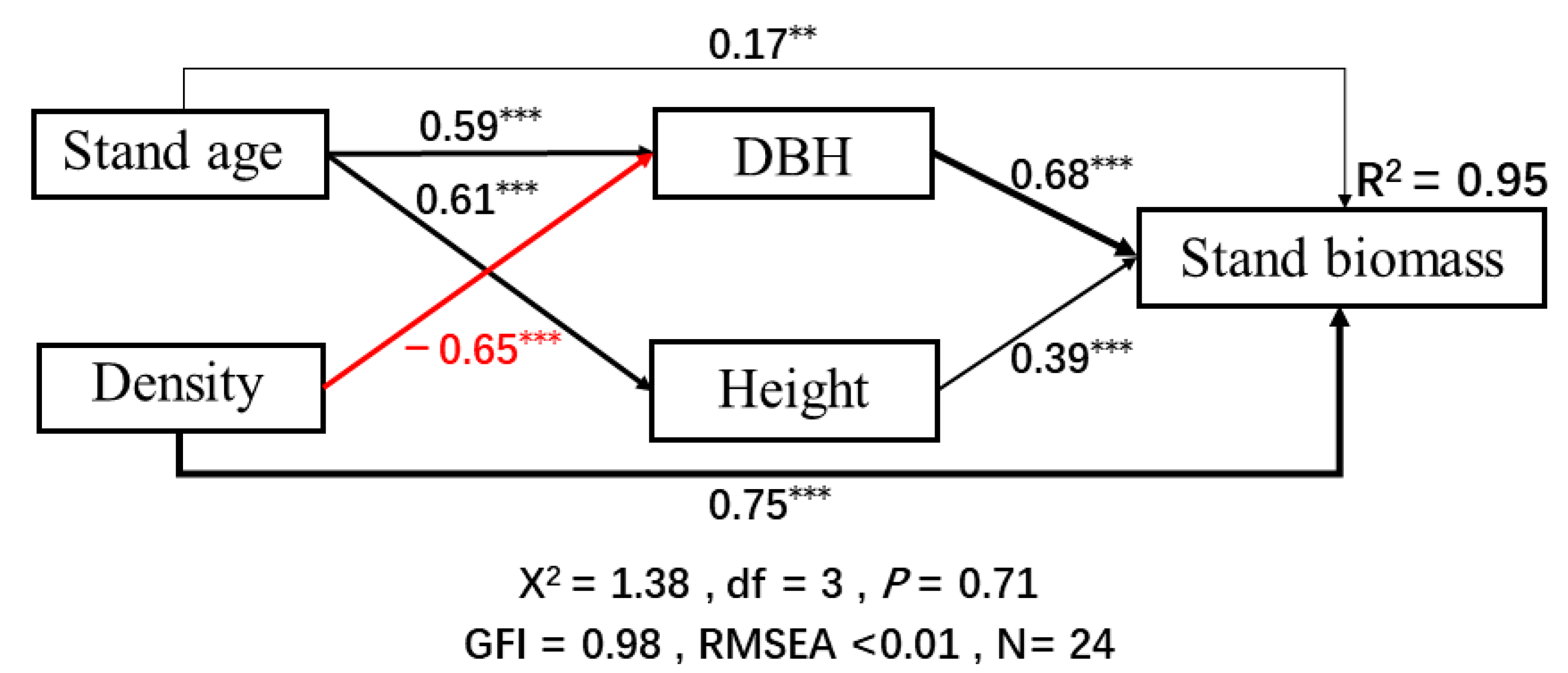
3.3. Potential Predictors of Stand Biomass
4. Discussion
4.1. Thinning Promoted Individual Tree Radial Growth
4.2. Thinning Effects on Stand Biomass
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Li, Z.; Xiao, J.; Lu, G.; Sun, W.; Ma, C.; Jin, Y. Productivity and Profitability of Larix principis-rupprechtii and Pinus tabuliformis Plantation Forests in Northeast China. For. Policy Econ. 2020, 121, 102314. [Google Scholar] [CrossRef]
- Feng, Y.; Schmid, B.; Loreau, M.; Forrester, D.I.; Fei, S.; Zhu, J.; Tang, Z.; Zhu, J.; Hong, P.; Ji, C.; et al. Multispecies Forest Plantations Outyield Monocultures across a Broad Range of Conditions. Science 2022, 376, 865–868. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Chen, A.; Peng, C.; Zhao, S.; Ci, L. Changes in Forest Biomass Carbon Storage in China between 1949 and 1998. Science 2001, 292, 2320–2322. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Peng, S.; Li, X.; Guo, Z.; Piao, S. Changes in Forest Biomass over China during the 2000s and Implications for Management. For. Ecol. Manag. 2015, 357, 76–83. [Google Scholar] [CrossRef]
- Loewe-Muñoz, V.; Balzarini, M.; Ortega González, M. Pure and Mixed Plantations of Persian Walnut (Juglans regia L.) for High Quality Timber Production in Chile, South America. J. Plant Ecol. 2020, 13, 12–19. [Google Scholar] [CrossRef]
- FAO. Global Forest Resources Assessment 2020; FAO: Rome, Italy, 2020.
- Zeng, W.; Tomppo, E.; Healey, S.P.; Gadow, K.V. The National Forest Inventory in China: History-Results-International Context. For. Ecosyst. 2015, 2, 23. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Wei, Y.; Fu, B.; Wang, S.; Zhao, Y.; Moran, E.F. Evolution and Effects of the Social-Ecological System over a Millennium in China’s Loess Plateau. Sci. Adv. 2020, 6, eabc0276. [Google Scholar] [CrossRef]
- Wang, T.; Wu, J.; Kou, X.; Oliver, C.; Mou, P.; Ge, J. Ecologically Asynchronous Agricultural Practice Erodes Sustainability of the Loess Plateau of China. Ecol. Appl. 2010, 20, 1126–1135. [Google Scholar] [CrossRef]
- Zhao, G.; Mu, X.; Wen, Z.; Wang, F.; Gao, P. Soil Erosion and Eco-environment Changes in the Loess Plateau. Land Degrad. Dev. 2013, 24, 499–510. [Google Scholar] [CrossRef]
- Li, Y.; Piao, S.; Li, L.Z.X.; Chen, A.; Wang, X.; Ciais, P.; Huang, L.; Lian, X.; Peng, S.; Zeng, Z.; et al. Divergent Hydrological Response to Large-Scale Afforestation and Vegetation Greening in China. Sci. Adv. 2018, 4, eaar4182. [Google Scholar] [CrossRef] [Green Version]
- Fu, B.; Wang, S.; Liu, Y.; Liu, J.; Liang, W.; Miao, C. Hydrogeomorphic Ecosystem Responses to Natural and Anthropogenic Changes in the Loess Plateau of China. Annu. Rev. Earth Planet. Sci. 2017, 45, 223–243. [Google Scholar] [CrossRef]
- Augusto, L.; Boča, A. Tree Functional Traits, Forest Biomass, and Tree Species Diversity Interact with Site Properties to Drive Forest Soil Carbon. Nat. Commun. 2022, 13, 1097. [Google Scholar] [CrossRef]
- Li, Y.; Bao, W.; Bongers, F.; Chen, B.; Chen, G.; Guo, K.; Jiang, M.; Lai, J.; Lin, D.; Liu, C.; et al. Drivers of Tree Carbon Storage in Subtropical Forests. Sci. Total Environ. 2019, 654, 684–693. [Google Scholar] [CrossRef]
- Jucker, T.; Bongalov, B.; Burslem, D.F.R.P.; Nilus, R.; Dalponte, M.; Lewis, S.L.; Phillips, O.L.; Qie, L.; Coomes, D.A. Topography Shapes the Structure, Composition and Function of Tropical Forest Landscapes. Ecol. Lett. 2018, 21, 989–1000. [Google Scholar] [CrossRef] [Green Version]
- Hui, D.; Wang, J.; Le, X.; Shen, W.; Ren, H. Influences of Biotic and Abiotic Factors on the Relationship between Tree Productivity and Biomass in China. For. Ecol. Manag. 2012, 264, 72–80. [Google Scholar] [CrossRef]
- Liu, L.; Zeng, F.; Song, T.; Wang, K.; Du, H. Stand Structure and Abiotic Factors Modulate Karst Forest Biomass in Southwest China. Forests 2020, 11, 443. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.; Yan, E.-R.; Chen, H.Y.H.; Chang, S.X.; Zhao, Y.-T.; Yang, X.-D.; Xu, M.-S. Stand Structural Diversity Rather than Species Diversity Enhances Aboveground Carbon Storage in Secondary Subtropical Forests in Eastern China. Biogeosciences 2016, 13, 4627–4635. [Google Scholar] [CrossRef] [Green Version]
- Dănescu, A.; Albrecht, A.T.; Bauhus, J. Structural Diversity Promotes Productivity of Mixed, Uneven-Aged Forests in Southwestern Germany. Oecologia 2016, 182, 319–333. [Google Scholar] [CrossRef]
- Ullah, F.; Gilani, H.; Sanaei, A.; Hussain, K.; Ali, A. Stand Structure Determines Aboveground Biomass across Temperate Forest Types and Species Mixture along a Local-Scale Elevational Gradient. For. Ecol. Manag. 2021, 486, 118984. [Google Scholar] [CrossRef]
- Noh, N.J.; Kim, C.; Bae, S.W.; Lee, W.K.; Yoon, T.K.; Muraoka, H.; Son, Y. Carbon and Nitrogen Dynamics in a Pinus Densiflora Forest with Low and High Stand Densities. J. Plant Ecol. 2013, 6, 368–379. [Google Scholar] [CrossRef] [Green Version]
- Ali, A. Forest Stand Structure and Functioning: Current Knowledge and Future Challenges. Ecol. Indic. 2019, 98, 665–677. [Google Scholar] [CrossRef]
- Yachi, S.; Loreau, M. Does Complementary Resource Use Enhance Ecosystem Functioning? A Model of Light Competition in Plant Communities. Ecol. Lett. 2007, 10, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Waters, C.M.; Gonsalves, L.; Law, B.; Melville, G.; Toole, I.; Brassil, T.; Tap, P. The Effect of Thinning on Structural Attributes of a Low Rainfall Forest in Eastern Australia. For. Ecol. Manag. 2018, 409, 571–583. [Google Scholar] [CrossRef]
- Hu, J.; Herbohn, J.; Chazdon, R.L.; Baynes, J.; Vanclay, J.K. Above-Ground Biomass Recovery Following Logging and Thinning over 46 Years in an Australian Tropical Forest. Sci. Total Environ. 2020, 734, 139098. [Google Scholar] [CrossRef] [PubMed]
- Harrod, R.J.; Peterson, D.W.; Povak, N.A.; Dodson, E.K. Thinning and Prescribed Fire Effects on Overstory Tree and Snag Structure in Dry Coniferous Forests of the Interior Pacific Northwest. For. Ecol. Manag. 2009, 258, 712–721. [Google Scholar] [CrossRef]
- Duque Lazo, J.; Navarro-Cerrillo, R.; Sanchez-Salguero, R.; Rodriguez Vallejo, C. Is Thinning an Alternative When Trees Could Die in Response to Drought? The Case of Planted Pinus nigra and P. sylvestris Stands in Southern Spain. For. Ecol. Manag. 2019, 433, 313–324. [Google Scholar]
- Wertz, B.; Bembenek, M.; Karaszewski, Z.; Ochał, W.; Skorupski, M.; Strzeliński, P.; Węgiel, A.; Mederski, P.S. Impact of Stand Density and Tree Social Status on Aboveground Biomass Allocation of Scots Pine Pinus sylvestris L. Forests 2020, 11, 765. [Google Scholar] [CrossRef]
- McMahon, T. Size and Shape in Biology. Science 1973, 179, 1201–1204. [Google Scholar] [CrossRef]
- Bullock, S.H. Developmental Patterns of Tree Dimensions in a Neotropical Deciduous Forest. Biotropica 2000, 32, 42–52. [Google Scholar] [CrossRef]
- Deng, C.; Zhang, S.; Lu, Y.; Froese, R.E.; Ming, A.; Li, Q. Thinning Effects on the Tree Height-Diameter Allometry of Masson Pine (Pinus massoniana Lamb.). Forests 2019, 10, 1129. [Google Scholar] [CrossRef] [Green Version]
- Fedorová, B.; Kadavý, J.; Adamec, Z.; Kneifl, M.; Knott, R. Response of Diameter and Height Increment to Thinning in Oak–Hornbeam Coppice in the Southeastern Part of the Czech Republic. J. For. Sci. 2016, 62, 229–235. [Google Scholar] [CrossRef] [Green Version]
- Dwyer, J.M.; Fensham, R.; Buckley, Y.M. Restoration Thinning Accelerates Structural Development and Carbon Sequestration in an Endangered Australian Ecosystem. J. Appl. Ecol. 2010, 47, 681–691. [Google Scholar] [CrossRef]
- Moreno-Fernández, D.; Cañellas, I.; Calama, R.; Gordo, J.; Sánchez-González, M. Thinning Increases Cone Production of Stone Pine (Pinus pinea L.) Stands in the Northern Plateau (Spain). Ann. For. Sci. 2013, 70, 761–768. [Google Scholar] [CrossRef] [Green Version]
- Dobner, M.; Nicoletti, M.F.; Arce, J.E. Influence of Crown Thinning on Radial Growth Pattern of Pinus taeda in Southern Brazil. New For. 2019, 50, 437–454. [Google Scholar] [CrossRef]
- Valinger, E. Effects of Thinning and Nitrogen Fertilization on Stem Growth and Stem Form of Pinus sylvestris Trees. Scand. J. For. Res. 1992, 7, 219–228. [Google Scholar] [CrossRef]
- Pape, R. Influence of Thinning and Tree Diameter Class on the Development of Basic Density and Annual Ring Width in Picea abies. Scand. J. For. Res. 1999, 14, 27–37. [Google Scholar] [CrossRef]
- Duan, J.; Abduwali, D. Basic Theory and Methods of Afforestation. In Silviculture; Cristina Gonçalves, A., Ed.; IntechOpen: London, UK, 2021. [Google Scholar]
- Ara, M.; Barbeito, I.; Elfving, B.; Johansson, U.; Nilsson, U. Varying Rectangular Spacing Yields No Difference in Forest Growth and External Wood Quality in Coniferous Forest Plantations. For. Ecol. Manag. 2021, 489, 119040. [Google Scholar] [CrossRef]
- Zhou, G.; Yin, G.; Tang, X. Carbon Stocks of Forest Ecosystems in China: Biomass Equation; Science Press: Beijing, China, 2018. (In Chinese) [Google Scholar]
- Lai, J.; Zou, Y.; Zhang, J.; Peres-Neto, P.R. Generalizing Hierarchical and Variation Partitioning in Multiple Regression and Canonical Analyses Using the rdacca. hp R Package. Methods Ecol. Evol. 2022, 13, 782–788. [Google Scholar] [CrossRef]
- Niklas, K.J. The Scaling of Plant Height: A Comparison Among Major Plant Clades and Anatomical Grades. Ann. Bot. 1993, 72, 165–172. [Google Scholar] [CrossRef]
- Martínez-Vilalta, J.; Vanderklein, D.; Mencuccini, M. Tree Height and Age–Related Decline in Growth in Scots Pine (Pinus sylvestris L.). Oecologia 2006, 150, 529–544. [Google Scholar] [CrossRef]
- Kerr, G. The Effect of Heavy or “Free Growth” Thinning on Oak (Quercus petraea and Q. robur). Forestry 1996, 69, 303–317. [Google Scholar] [CrossRef] [Green Version]
- Makinen, H. Thinning Intensity and Growth of Norway Spruce Stands in Finland. Forestry 2004, 77, 349–364. [Google Scholar] [CrossRef] [Green Version]
- Missanjo, E.; Kamanga-Thole, G. Effect of First Thinning and Pruning on the Individual Growth of Pinus patula Tree Species. J. For. Res. 2015, 26, 827–831. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, H.; Dong, B.; Zhou, M.; Ma, L.; Jia, Z.; Duan, J. Regeneration Response to Canopy Gap Size in a Chinese Pine Plantation: Species Diversity Patterns, Size Structures and Spatial Distributions. For. Ecol. Manag. 2017, 397, 97–107. [Google Scholar] [CrossRef]
- Moles, A.T.; Warton, D.I.; Warman, L.; Swenson, N.G.; Laffan, S.W.; Zanne, A.E.; Pitman, A.; Hemmings, F.A.; Leishman, M.R. Global Patterns in Plant Height. J. Ecol. 2009, 97, 923–932. [Google Scholar] [CrossRef]
- Botero, C.A.; Weissing, F.J.; Wright, J.; Rubenstein, D.R. Evolutionary Tipping Points in the Capacity to Adapt to Environmental Change. Proc. Natl. Acad. Sci. USA 2015, 112, 184–189. [Google Scholar] [CrossRef]
- Bosela, M.; Štefančík, I.; Marčiš, P.; Rubio-Cuadrado, Á.; Lukac, M. Thinning Decreases Above-Ground Biomass Increment in Central European Beech Forests but Does Not Change Individual Tree Resistance to Climate Events. Agric. For. Meteorol. 2021, 306, 108441. [Google Scholar] [CrossRef]
- Bouriaud, O.; Don, A.; Janssens, I.A.; Marin, G.; Schulze, E.D. Effects of forest management on biomass stocks in Romanian beech forests. For. Ecosyst. 2019, 6, 19. [Google Scholar] [CrossRef] [Green Version]
- Coletta, V.; Menguzzato, G.; Pellicone, G.; Veltri, A.; Marziliano, P.A. Effect of Thinning on Above-Ground Biomass Accumulation in a Douglas-Fir Plantation in Southern Italy. J. For. Res. 2016, 27, 1313–1320. [Google Scholar] [CrossRef]
- Eriksson, E. Thinning Operations and Their Impact on Biomass Production in Stands of Norway Spruce and Scots Pine. Biomass Bioenerg. 2006, 30, 848–854. [Google Scholar] [CrossRef]
- Węgiel, A.; Bembenek, M.; Łacka, A.; Mederski, P.S. Relationship between stand density and value of timber assortments: A case study for Scots pine stands in north-western Poland. N. Z. J. For. Sci. 2018, 48, 12. [Google Scholar] [CrossRef]
- Ma, S.-H.; Eziz, A.; Tian, D.; Yan, Z.-B.; Cai, Q.; Jiang, M.-W.; Ji, C.-J.; Fang, J.-Y. Size- and Age-Dependent Increases in Tree Stem Carbon Concentration: Implications for Forest Carbon Stock Estimations. J. Plant Ecol. 2020, 13, 233–240. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Management | Thinned | Unthinned | ||||||
|---|---|---|---|---|---|---|---|---|
| Stand (years) | 20 | 30 | 40 | 50 | 20 | 30 | 40 | 50 |
| Location | 36.08° N | 36.06° N | 36.11° N | 35.93° N | 36.08° N | 36.25° N | 36.11° N | 35.87° N |
| 108.57° E | 108.10° E | 108.67° E | 108.29° E | 108.56° E | 108.57° E | 108.34° E | 108.30° E | |
| Elevation (m) | 1225 | 1337 | 1258 | 1426 | 1329 | 1422 | 1598 | 1352 |
| Slope (°, aspect) | 25 SE | 15 W | 20 NW | 10 N | 8 NW | 5 N | 5 NE | 20 SE |
| Mean DBH (cm) | 7.0 | 9.6 | 7.8 | 7.0 | 5.5 | 5.7 | 4.9 | 7.2 |
| Max DBH (cm) | 11.6 | 13.8 | 13.3 | 13.2 | 11.5 | 10.6 | 10.6 | 11.4 |
| Mean Height (m) | 7.6 | 9.6 | 11.6 | 11.3 | 6.3 | 9.8 | 9.6 | 11.3 |
| Max Height (m) | 10.2 | 11.5 | 14.8 | 15.0 | 8.2 | 12.2 | 12.9 | 16.9 |
| TBA (m2 ha−1) | 3.8 | 5.6 | 6.1 | 5.5 | 8.3 | 11.8 | 10.5 | 9.9 |
| Stand Density | 1717 | 2258 | 2483 | 2383 | 3575 | 4816 | 4850 | 4008 |
| (stems/ha−1) | ||||||||
| Site index (m) | 9.27 | 9.75 | 10.22 | 10.69 | 7.24 | 8.64 | 10.1 | 11.53 |
| Stand area (ha) | 200 | 200 | 200 | 200 | 200 | 200 | 200 | 200 |
| Species | AGB (kg) | BGB (kg) | Species in This Study |
|---|---|---|---|
| Pinus tabulaeformis | Pinus tabulaeformis | ||
| Quercus mongolica | Quercus mongolica | ||
| Broad-leaved trees | Acer pilosum, A. tataricum, Malus baccata |
| Forest Types | Variables | Model Name | df | AIC | R2 | p Value |
|---|---|---|---|---|---|---|
| Thinned | DBH | linear | 1051 | 6189 | 0.155 | <0.001 |
| exponential | 1051 | 2778 | 0.121 | <0.001 | ||
| logistic | 1051 | 2034 | 0.143 | <0.001 | ||
| Height | linear | 1051 | 4811 | 0.042 | <0.001 | |
| exponential | 1051 | 1851 | 0.028 | <0.001 | ||
| logistic | 1051 | 3445 | 0.039 | <0.001 | ||
| Unthinned | DBH | linear | 2060 | 1156 | 0.048 | <0.001 |
| exponential | 2060 | 3114 | 0.008 | 0.004 | ||
| logistic | 2060 | 1901 | 0.036 | <0.001 | ||
| Height | linear | 2060 | 9071 | 0.334 | <0.001 | |
| exponential | 2060 | 4019 | 0.256 | <0.001 | ||
| logistic | 2060 | 2978 | 0.328 | <0.001 |
| Variables | Forest Management | Stand Age (Years) | |||
|---|---|---|---|---|---|
| 20 | 30 | 40 | 50 | ||
| DBH (cm) | unthinned | 10.97 ± 0.32 b | 11.34 ± 0.88 b | 9.73 ± 1.12 b | 14.42 ± 0.38 a |
| thinned | 13.45 ± 0.51 b * | 14.06 ± 1.03 b | 15.53 ± 1.06 ab ** | 18.88 ± 2.15 a | |
| Height (m) | unthinned | 6.25 ± 0.41 b | 9.81 ± 0.90 a | 9.76 ± 1.81 a | 11.32 ± 0.56 a |
| thinned | 7.47 ± 0.48 b * | 11.22 ± 0.65 a | 11.58 ± 0.92 a | 9.39 ± 0.95 ab * | |
| Individual tree biomass (kg) | unthinned | 27.38 ± 2.94 b | 38.78 ± 9.48 b | 39.87 ± 8.73 b | 67.47 ± 3.58 a |
| thinned | 42.75 ± 3.88 b * | 68.25 ± 11.23 ab | 82.77 ± 14.16 ab * | 91.96 ± 26.12 a | |
| Stand biomass (Mg ha−1) | unthinned | 97.69 ± 19.2 b | 186.76 ± 52.32 ab | 193.34 ± 75.56 ab | 268.70 ± 19.82 b |
| thinned | 72.52 ± 11.37 b | 151.63 ± 20.21 ab | 203.17 ± 33.15 a | 215.05 ± 53.55 a | |
| Predictor | Direct Effects | Indirect Effects | Total Effects |
|---|---|---|---|
| Stand biomass | |||
| Thinning | −0.36 | 0.11 | −0.25 |
| Stand age | 0.00 | 0.75 | 0.75 |
| Density | 0.54 | −0.09 | 0.45 |
| DBH | 0.85 | 0.00 | 0.85 |
| Height | 0.50 | 0.00 | 0.50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Li, H.; Zhang, W.; Chen, B.; Yang, L.; Li, M.; Zhu, J.; Cai, Q. The Effects of Thinning on Tree Growth and Stand Biomass in a Chronosequence of Pinus tabulaeformis Plantations in the Loess Plateau of China. Forests 2023, 14, 1620. https://doi.org/10.3390/f14081620
Li Y, Li H, Zhang W, Chen B, Yang L, Li M, Zhu J, Cai Q. The Effects of Thinning on Tree Growth and Stand Biomass in a Chronosequence of Pinus tabulaeformis Plantations in the Loess Plateau of China. Forests. 2023; 14(8):1620. https://doi.org/10.3390/f14081620
Chicago/Turabian StyleLi, Yuanchun, Huipeng Li, Wei Zhang, Baolin Chen, Lei Yang, Mengfan Li, Jianxiao Zhu, and Qiong Cai. 2023. "The Effects of Thinning on Tree Growth and Stand Biomass in a Chronosequence of Pinus tabulaeformis Plantations in the Loess Plateau of China" Forests 14, no. 8: 1620. https://doi.org/10.3390/f14081620