
An Integrative Taxonomic Study of Parasola (Psathyrellaceae, Fungi) Reveals a New Saprotrophic Species from European Temperate Deciduous Forests
 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fieldwork, Sampling, and Morphological Study
2.2. DNA Extraction, PCR Amplification, and Sequencing
2.3. Sequence Alignment and Phylogenetic Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Country | Voucher | ITS | LSU | tef-1α | β-tub | Ref. |
|---|---|---|---|---|---|---|---|
| Coprinopsis lagopus | Hungary | NL-2143 | FM163179 | FM160730 | –– | –– | [33] |
| Coprinopsis marcescibilis | Hungary | NL-2140 | FM878020 | FM876277 | –– | –– | [34] |
| Coprinopsis picacea | Hungary | NL-0174 | FN396115 | FN396166 | –– | –– | [26] |
| Coprinopsis pseudonivea | Hungary | NL-2340 | FM163181 | FM160728 | –– | –– | [33] |
| Parasola aporos | France | RC-F92.191 holotype | MK397584 | MK397604 | –– | –– | [17] |
| Parasola aporos | France | CL-F09.005 | MK397586 | MK397606 | –– | –– | [17] |
| Parasola auricoma | Hungary | NL-0087 | JN943107 | JQ045871 | FM897236 | FN396252 | [35] |
| Parasola auricoma | Croatia | CNF 1/4718 | OQ842767 | OQ842768 | OQ850152 | OQ850168 | This study |
| Parasola auricoma | Croatia | CNF 1/4618 | OQ845889 | OQ845835 | OQ850153 | OQ850169 | This study |
| Parasola conopilea | The Netherlands | CBS 325.39 | MH856033 | MH867531 | –– | –– | [36] |
| Parasola conopilea | Croatia | CNF 1/5310 | OQ845887 | OQ845888 | OQ850154 | OQ850170 | This study |
| Parasola conopilea | Croatia | CNF 1/5735 | OQ845890 | OQ843455 | OQ850155 | OQ850171 | This study |
| Parasola crataegi | Germany | SSt08-154 holotype | KY928605 | –– | –– | –– | [10] |
| Parasola crataegi | Hungary | NL-4175 paratype | KY928603 | KY928631 | –– | –– | [10] |
| Parasola crataegi | Croatia | CNF 1/8905 | OQ852892 | –– | –– | –– | This study |
| Parasola cuniculorum | United Kingdom | K(M)191984 holotype | OL630105 | –– | –– | –– | [16] |
| Parasola cuniculorum | Croatia | CNF 1/5143 | OQ848756 | OQ848757 | OQ850156 | OQ850172 | This study |
| Parasola galericuliformis | Hungary | NL-6601 | FM163187 | FM160722 | –– | –– | [33] |
| Parasola galericuliformis | Sweden | NL-0095 | FM163188 | FM160721 | –– | –– | [33] |
| Parasola glabra | Pakistan | LAH-SHP-5 holotype | KY461717 | KY621806 | KY461735 | –– | [12] |
| Parasola glabra | Pakistan | HUP-SHP-23 paratype | KY461718 | KY621805 | –– | –– | [12] |
| Parasola hercules | The Netherlands | Uljé 10.8.1984 (L146) holotype | HQ847027 | HQ847112 | –– | –– | [37] |
| Parasola kuehneri | The Netherlands | Uljé 31.5.1987 holotype | KY928608 | KY928633 | –– | –– | [10] |
| Parasola kuehneri | The Netherlands | Uljé 1241 (L133) | HQ847026 | HQ847111 | –– | –– | [37] |
| Parasola kuehneri | Croatia | CNF 1/4334 | OQ849153 | OQ849154 | OQ850157 | OQ850173 | This study |
| Parasola lactea | Hungary | NL-0283 | JN943113 | JQ045887 | FM897239 | FN396248 | [35] |
| Parasola lactea | Hungary | NL-0288 | JN943106 | JQ045872 | FM897233 | FN396250 | [35] |
| Parasola lilatincta | Hungary | NL-0468a | FM163200 | FM160709 | –– | –– | [33] |
| Parasola lilatincta | Hungary | NL-0281 | FM163197 | FM160712 | –– | –– | [33] |
| Parasola lilatincta | Hungary | NL-0296 | FM163196 | FM160713 | –– | –– | [33] |
| Parasola lilatinctoides | Pakistan | LAH-SHP-8 holotype | KY461722 | KY461725 | KY461731 | –– | [12] |
| Parasola litoralis | Cyprus | K(M)264814 holotype | OL630108 | –– | –– | –– | [16] |
| Parasola litoralis | Cyprus | DJS20130125001 paratype | OL630107 | –– | –– | –– | [16] |
| Parasola malakandensis | Pakistan | HUP 17501 holotype | KP738713 | KU599829 | KU599831 | –– | [9] |
| Parasola malakandensis | Pakistan | LAH-SHP-17 paratype | KU599827 | KU599830 | KU599832 | –– | [9] |
| Parasola malakandensis | Croatia | CNF 1/8698 | OQ849158 | OQ849167 | OQ850158 | OQ850174 | This study |
| Parasola megasperma | United Kingdom | E:Orton 4132 holotype | OL630101 | –– | OL630935 | –– | [16] |
| Parasola megasperma | The Netherlands | Ulje 1275 | KY928618 | KY928637 | –– | –– | [10] |
| Parasola megasperma | Croatia | CNF 1/5704 | OQ849166 | OQ849224 | OQ850159 | OQ850175 | This study |
| Parasola misera | Hungary | NL-0280 neotype | FM163210 | FM160699 | –– | –– | [33] |
| Parasola misera | Hungary | NL-0677 | FM163211 | FM160698 | FM897240 | FN396249 | [26,33] |
| Parasola nudiceps | United Kingdom | E:Orton 4133 holotype | OL630102 | –– | –– | –– | [16] |
| Parasola nudiceps | Germany | HB19870911A | MK063783 | –– | –– | –– | [16] |
| Parasola nudiceps | Croatia | CNF 1/4804 | OQ849230 | OQ849229 | OQ850160 | OQ850176 | This study |
| Parasola nudiceps (as P. ochracea) | Norway | NL-3621, holotype of P. ochracea | JN943134 | JQ045875 | –– | –– | [38] |
| Parasola nudiceps (as P. ochracea) | Sweden | NL-3167, paratype of P. ochracea | JN943136 | JQ045865 | –– | –– | [38] |
| Parasola papillatospora sp. nov. | Croatia | CNF 1/3473 | OQ862758 | OQ862756 | OQ850161 | –– | This study |
| Parasola papillatospora sp. nov. | Croatia | CNF 1/5428 | OQ862789 | OQ862577 | OQ850162 | OQ850177 | This study |
| Parasola papillatospora sp. nov. | Croatia | CNF 1/7600 | OQ862790 | OQ862578 | OQ850163 | OQ850178 | This study |
| Parasola papillatospora sp. nov. | Croatia | CNF 1/7858 holotype | OQ862770 | OQ862755 | OQ850164 | OQ850179 | This study |
| Parasola papillatospora sp. nov. | Croatia | CNF 1/7861 | OQ862757 | OQ862771 | OQ850165 | OQ850182 | This study |
| Parasola papillatospora sp. nov. | Croatia | CNF 1/7902 | OQ862788 | OQ862772 | OQ850167 | OQ850181 | This study |
| Parasola papillatospora sp. nov. | Hungary | SZMC-NL-2952 | HQ847028 | HQ847113 | –– | –– | [10] |
| Parasola parvula | India | CAL 1667 holotype | MH379796 | MH393599 | –– | –– | [11] |
| Parasola plicatilis | Hungary | NL-0075 epitype | FM163214 | FM160695 | –– | –– | [33] |
| Parasola plicatilis | Hungary | NL-0284 | FM163189 | FM160720 | FM897235 | FN396251 | [26,33] |
| Parasola plicatilis aff. | China | HMJAU46405 | OL355167 | OL376339 | –– | –– | [39] |
| Parasola plicatilis-similis | Sweden | NL-2125 holotype | KY928620 | –– | –– | –– | [10] |
| Parasola plicatilis-similis | Sweden | NL-0287 paratype | FM163218 | FM160691 | –– | FN396245 | [26,33,35] |
| Parasola plicatilis-similis | Croatia | CNF 1/5484 | OQ850018 | OQ850017 | OQ850166 | OQ850180 | This study |
| Parasola psathyrelloides | India | CAL 1753 holotype | MK682756 | MK682754 | –– | –– | [13] |
| Parasola psathyrelloides | India | AMH 10119 paratype | MK682752 | MK682759 | –– | –– | [13] |
| Parasola pseudolactea | Pakistan | HUP-SU-412 holotype | KY461719 | KY621799 | KY461733 | –– | [12] |
| Parasola pseudolactea | Pakistan | HUP-SU-413 paratype | KY461720 | KY621800 | KY461734 | –– | [12] |
| Parasola schroeteri | Germany | Dähncke 1502 | KY928616 | KY928635 | –– | –– | [10] |
| Parasola schroeteri | The Netherlands | Brier 10.5.1999 | FM163219 | FM160690 | –– | –– | [33] |
| Parasola schroeteri | The Netherlands | Uljé 1067 | KY928627 | –– | –– | –– | [10] |
| Parasola schroeteri | The Netherlands | Vellinga 1140 | KY928629 | KY928645 | –– | –– | [10] |
| Parasola setulosa | Hungary | Maruyama 14.7.1999/ L32 | HQ847030 | HQ847115 | –– | –– | [37] |
| Parasola setulosa | China | HMJAU46367 | MW822929 | OL376319 | –– | –– | [39] |
3. Results
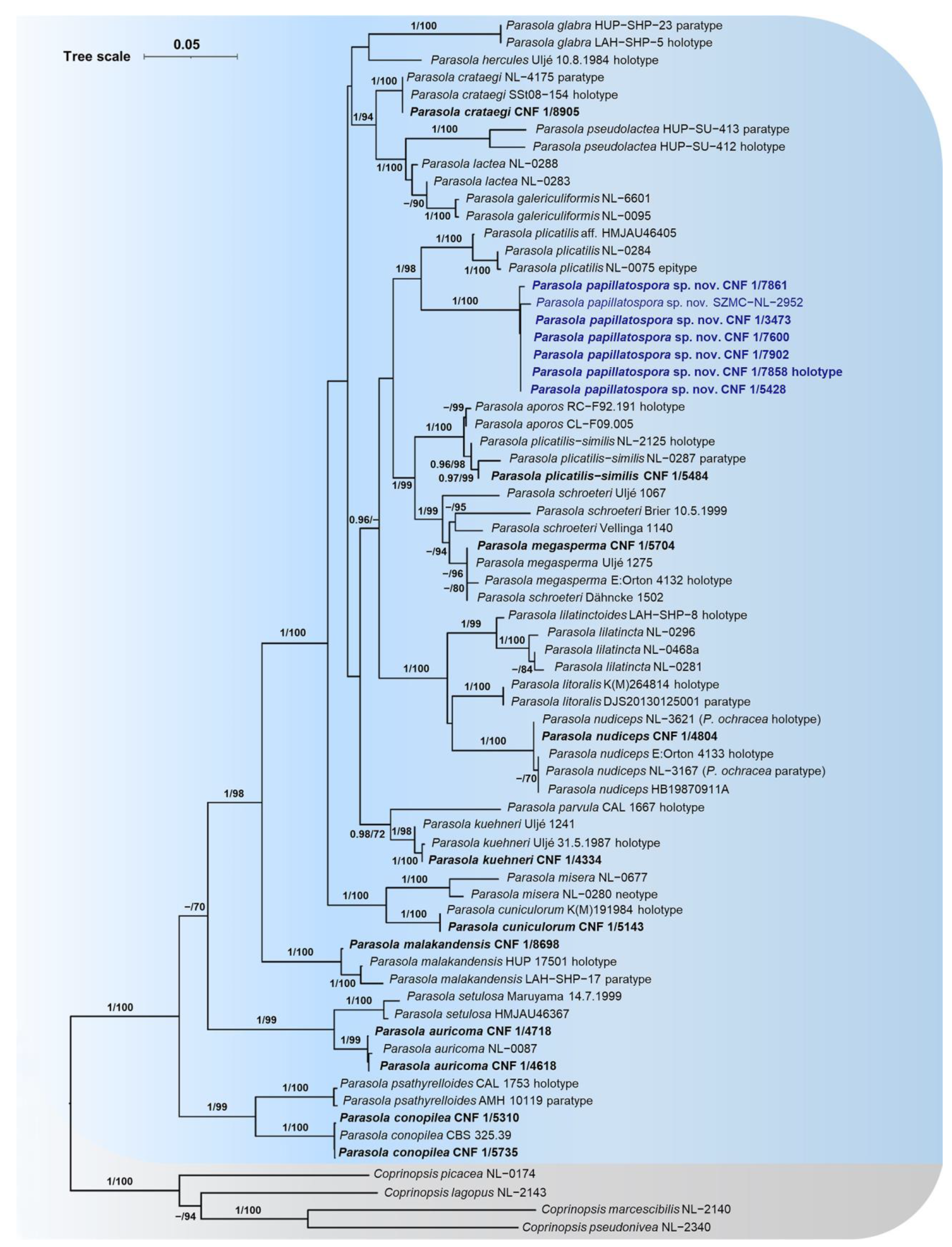
3.1. Molecular Phylogenetic Analyses
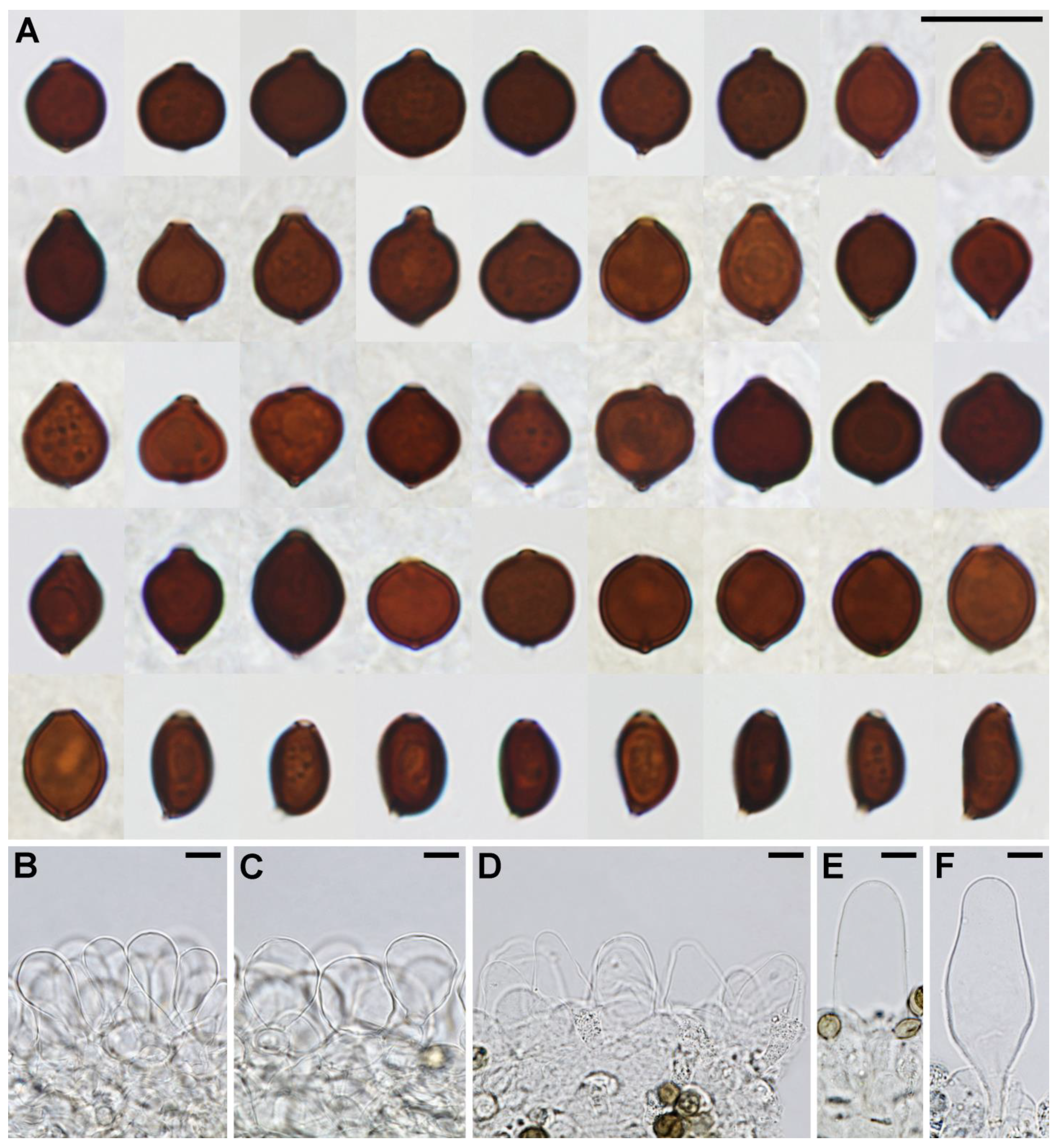
3.2. Taxonomy
3.3. Additional Data on Parasola from Croatia
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Redhead, S.A.; Vilgalys, R.; Moncalvo, J.-M.; Johnson, J.; Hopple, J.S. Coprinus Pers. and the Disposition of Coprinus Species Sensu Lato. Taxon 2001, 50, 203–241. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hopple, J.S.; Hibbett, D.S. Phylogenetic Implications of Generic Concepts in Fungal Taxonomy: The Impact of Molecular Systematic Studies. Mycol. Helv. 1994, 6, 73–91. [Google Scholar]
- Hopple, J.S.; Vilgalys, R. Phylogenetic Relationships in the Mushroom Genus Coprinus and Dark-Spored Allies Based on Sequence Data from the Nuclear Gene Coding for the Large Ribosomal Subunit RNA: Divergent Domains, Outgroups, and Monophyly. Mol. Phylogenet. Evol. 1999, 13, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Moncalvo, J.M.; Lutzoni, F.M.; Rehner, S.A.; Johnson, J.; Vilgalys, R. Phylogenetic Relationships of Agaric Fungi Based on Nuclear Large Subunit Ribosomal DNA Sequences. Syst. Biol. 2000, 49, 278–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uljé, C.B. Coprinus Pers. In Flora Agaricina Neerlandica, Critical Monographs on Families of Agarics and Boleti Occurring in the Netherlands; Noordeloos, M.E., Kuyper, T.W., Veliinga, E.C., Eds.; Taylor & Francis: London, UK, 2005; pp. 22–109. [Google Scholar]
- Nagy, L.G.; Vágvölgyi, C.; Papp, T. Type Studies and Nomenclatural Revisions in Parasola (Psathyrellaceae) and Related Taxa. Mycotaxon 2010, 112, 103–141. [Google Scholar] [CrossRef]
- Larsson, E.; Örstadius, L. Fourteen Coprophilous Species of Psathyrella Identified in the Nordic Countries Using Morphology and Nuclear RDNA Sequence Data. Mycol. Res. 2008, 112, 1165–1185. [Google Scholar] [CrossRef]
- Schafer, D.J. The Genus Parasola in Britain Including Parasola cuniculorum sp. nov. Field Mycol. 2014, 15, 77–99. [Google Scholar] [CrossRef]
- Hussain, S.; Afshan, N.u.S.; Ahmad, H.; Khalid, A.N.; Niazi, A.R. Parasola malakandensis sp. nov. (Psathyrellaceae; Basidiomycota) from Malakand, Pakistan. Mycoscience 2017, 58, 69–76. [Google Scholar] [CrossRef]
- Szarkandi, J.G.; Schmidt-Stohn, G.; Dima, B.; Hussain, S.; Kocsube, S.; Papp, T.; Vagvolgyi, C.; Nagy, L.G. The Genus Parasola: Phylogeny and the Description of Three New Species. Mycologia 2017, 109, 620–629. [Google Scholar] [CrossRef] [Green Version]
- Ganga, K.G.G.; Manimohan, P. A New Species and a New Record of Parasola from Kerala State, India. Phytotaxa 2018, 369, 260–268. [Google Scholar] [CrossRef]
- Hussain, S.; Ahmad, H.; Ullah, S.; Afshan, N.U.S.; Pfister, D.H.; Sher, H.; Ali, H.; Khalid, A.N. The Genus Parasola in Pakistan with the Description of Two New Species. MycoKeys 2018, 30, 41–60. [Google Scholar] [CrossRef] [PubMed]
- Greeshma Ganga, K.G.; Manimohan, P. Parasola psathyrelloides (Psathyrellaceae), a New Species from Kerala State, India. Phytotaxa 2019, 405, 255–262. [Google Scholar] [CrossRef]
- Voto, P. Novelties in the Family Psathyrellaceae. Part II. Boll. Am. 2019, 108, 127–133. [Google Scholar]
- Voto, P. Novelties in the Family Psathyrellaceae. Part V. Micol. E Veg. Mediterr. 2021, 35, 149–168. [Google Scholar]
- Schafer, D.; Alvarado, P.; Smith, L.; Liimatainen, K.; Loizides, M. Coprinoid Psathyrellaceae Species from Cyprus: Three New Sabulicolous Taxa from Sand Dunes and a Four-Spored Form of the Fimicolous Species Parasola cuniculorum. Mycol. Prog. 2022, 21, 52. [Google Scholar] [CrossRef]
- Malysheva, E.; Moreno, G.; Villarreal, M.; Malysheva, V.; Svetasheva, T. The Secotioid Genus Galeropsis (Agaricomycetes, Basidiomycota): A Real Taxonomic Unit or Ecological Phenomenon? Mycol. Prog. 2019, 18, 805–831. [Google Scholar] [CrossRef]
- Tibpromma, S.; Hyde, K.D.; Jeewon, R.; Maharachchikumbura, S.S.N.; Liu, J.K.; Bhat, D.J.; Jones, E.B.G.; McKenzie, E.H.C.; Camporesi, E.; Bulgakov, T.S.; et al. Fungal Diversity Notes 491–602: Taxonomic and Phylogenetic Contributions to Fungal Taxa. Fungal Divers. 2017, 83, 1–261. [Google Scholar] [CrossRef]
- Mešić, A.; Tkalčec, Z. Preliminary Checklist of Agaricales from Croatia IV: Families Bolbitiaceae, Coprinaceae, Entolomataceae and Pluteaceae. Mycotaxon 2003, 87, 283–309. [Google Scholar]
- Vrščaj, D. Popis Gljiva Otoka Krka—1. Dio. Gljiv. Glas. 2002, 15, 21–25. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. PCR Protoc. 1990, 315–322. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS Primers with Enhanced Specificity for Basidiomycetes—Application to the Identification of Mycorrhizae and Rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Vilgalys, R.; Hester, M. Rapid Genetic Identification and Mapping of Enzymatically Amplified Ribosomal DNA from Several Cryptococcus Species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehner, S. Primers for Elongation Factor 1-α (EF1-α). 2001. Available online: http://ocid.NACSE.ORG/research/deephyphae/EF1primer.pdf (accessed on 11 February 2022).
- Rehner, S.A.; Buckley, E. A Beauveria Phylogeny Inferred from Nuclear ITS and EF1-Alpha Sequences: Evidence for Cryptic Diversification and Links to Cordyceps Teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Nagy, L.G.; Walther, G.; Házi, J.; Vágvölgyi, C.; Papp, T. Understanding the Evolutionary Processes of Fungal Fruiting Bodies: Correlated Evolution and Divergence Times in the Psathyrellaceae. Syst. Biol. 2011, 60, 303–317. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Misawa, K.; Kuma, K.I.; Miyata, T. MAFFT: A Novel Method for Rapid Multiple Sequence Alignment Based on Fast Fourier Transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A Fast Online Phylogenetic Tool for Maximum Likelihood Analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian Inference of Phylogenetic Trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree of Life (ITOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Nagy, L.G.; Kocsubé, S.; Papp, T.; Vágvölgyi, C. Phylogeny and Character Evolution of the Coprinoid Mushroom Genus Parasola as Inferred from LSU and ITS nrDNA Sequence Data. Persoonia Mol. Phylogeny Evol. Fungi 2009, 22, 28–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, L.G.; Urban, A.; Örstadius, L.; Papp, T.; Larsson, E.; Vágvölgyi, C. The Evolution of Autodigestion in the Mushroom Family Psathyrellaceae (Agaricales) Inferred from Maximum Likelihood and Bayesian Methods. Mol. Phylogenet. Evol. 2010, 57, 1037–1048. [Google Scholar] [CrossRef] [PubMed]
- Wächter, D.; Melzer, A. Proposal for a Subdivision of the Family Psathyrellaceae Based on a Taxon-Rich Phylogenetic Analysis with Iterative Multigene Guide Tree. Mycol. Prog. 2020, 19, 1151–1265. [Google Scholar] [CrossRef]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-Scale Generation and Analysis of Filamentous Fungal DNA Barcodes Boosts Coverage for Kingdom Fungi and Reveals Thresholds for Fungal Species and Higher Taxon Delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Nagy, L.G.; Házi, J.; Szappanos, B.; Kocsubé, S.; Bálint, B.; Rákhely, G.; Vágvölgyi, C.; Papp, T. The Evolution of Defense Mechanisms Correlate with the Explosive Diversification of Autodigesting Coprinellus Mushrooms (Agaricales, Fungi). Syst. Biol. 2012, 61, 595–607. [Google Scholar] [CrossRef] [Green Version]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Bolchacova, E.; Voigt, K.; Crous, P.W.; et al. Nuclear Ribosomal Internal Transcribed Spacer (ITS) Region as a Universal DNA Barcode Marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef]
- Zhu, L.; Huang, M.; Bau, T. Taxonomy of Coprinoid Fungi in China. Mycosystema 2022, 41, 878–898. [Google Scholar] [CrossRef]
- Kits van Waveren, E. The Dutch, French and British Species of Psathyrella. Persoonia-Supplement 1985, 2, 3–300. [Google Scholar]
- Ludwig, E. Pilzkompendium. Band 2. Die Größeren Gattungen der Agaricales mit Farbigem Sporenpulver (Ausgenommen Cortinariaceae); FUNGICON Verlag: Berlin, Germany, 2007; ISBN 9783940316004. [Google Scholar]
- Orton, P.D. Notes on British Agarics IV. Notes R. Bot. Gard. Edinb. 1972, 32, 135–150. [Google Scholar]
- Uljé, C.B.; Bender, H. Additional Studies in Coprinus Subsection Glabri. Persoonia 1997, 16, 373–381. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions, and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions, or products referred to in the content. |




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pošta, A.; Tkalčec, Z.; Kušan, I.; Matočec, N.; Pole, L.; Čerkez, M.; Mešić, A. An Integrative Taxonomic Study of Parasola (Psathyrellaceae, Fungi) Reveals a New Saprotrophic Species from European Temperate Deciduous Forests. Forests 2023, 14, 1387. https://doi.org/10.3390/f14071387
Pošta A, Tkalčec Z, Kušan I, Matočec N, Pole L, Čerkez M, Mešić A. An Integrative Taxonomic Study of Parasola (Psathyrellaceae, Fungi) Reveals a New Saprotrophic Species from European Temperate Deciduous Forests. Forests. 2023; 14(7):1387. https://doi.org/10.3390/f14071387
Chicago/Turabian StylePošta, Ana, Zdenko Tkalčec, Ivana Kušan, Neven Matočec, Lucia Pole, Milan Čerkez, and Armin Mešić. 2023. "An Integrative Taxonomic Study of Parasola (Psathyrellaceae, Fungi) Reveals a New Saprotrophic Species from European Temperate Deciduous Forests" Forests 14, no. 7: 1387. https://doi.org/10.3390/f14071387