
A New Species of Ampelovirus Detected in Persea lingue (Ruiz & Pav.) Nees ex Kopp, a Common Tree of the Threatened Chilean Sclerophyll Forest
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. RNA Extraction and Genome Sequencing Analyses
2.3. RT-PCR Detection
3. Results
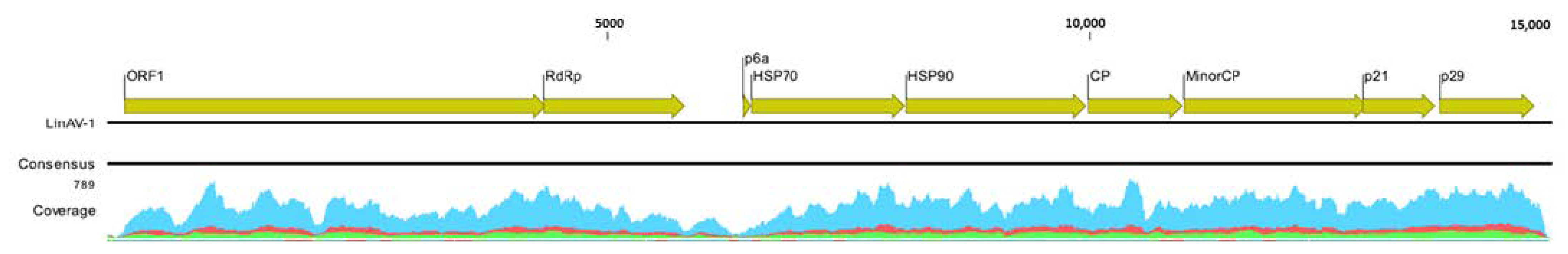
3.1. Identification and Molecular Characterization of the New Virus in Lingue
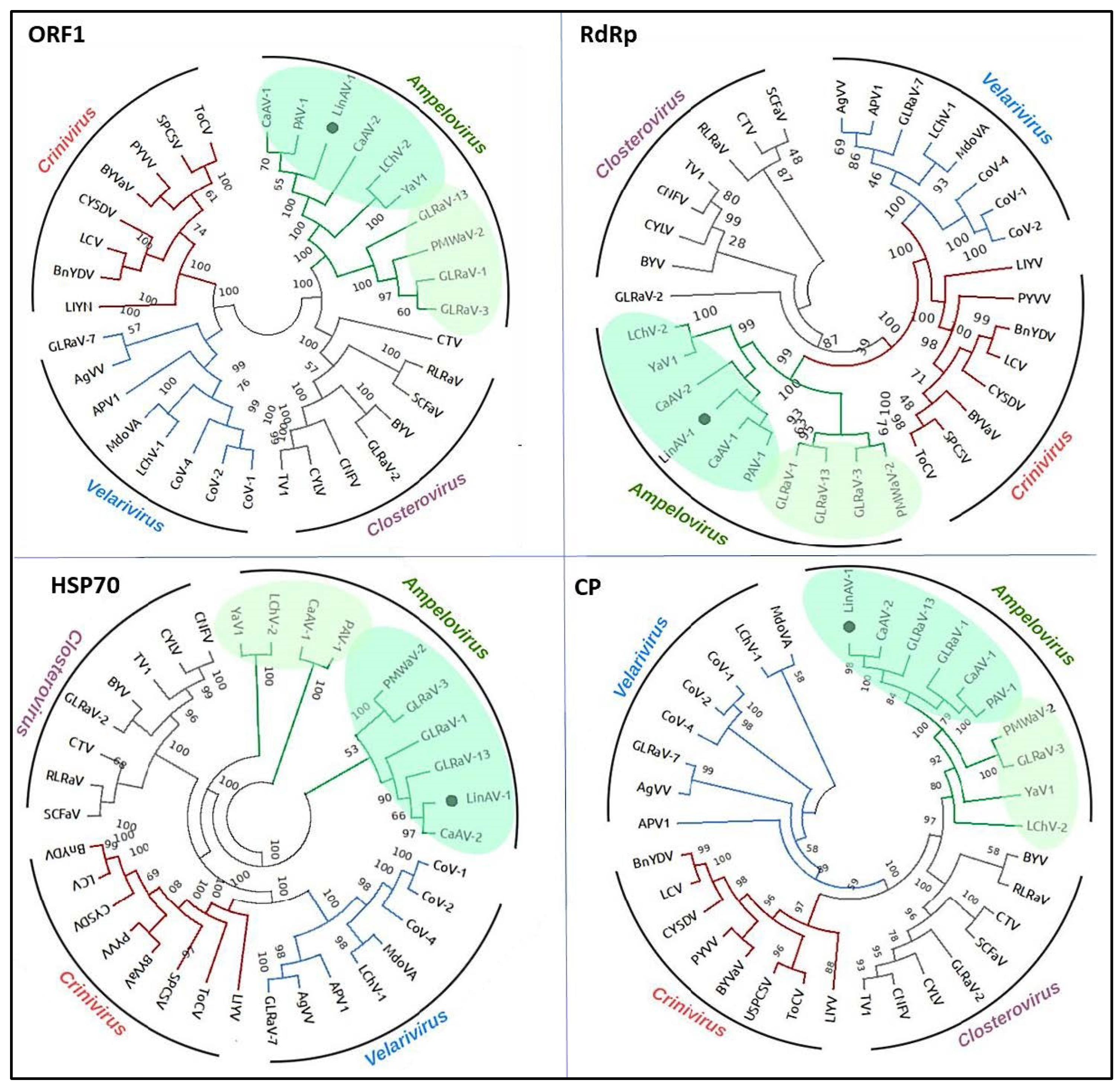
3.2. Phylogenetic Relationships of the New Virus in the Closteroviridae Family
3.3. Survey of LinAV
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fuchs, M.; Bar-Joseph, M.; Candresse, T.; Maree, H.J.; Martelli, G.P.; Melzer, M.J.; Menzel, W.; Minafra, A.; Sabanadzovic, S.; ICTV Report Consortium. ICTV Virus Taxonomy Profile: Closteroviridae. J. Gen. Virol. 2020, 101, 364–365. [Google Scholar] [CrossRef]
- Dolja, V.V.; Kreuze, J.F.; Valkonen, J.P.T. Comparative and functional genomics of closteroviruses. Virus Res. 2006, 117, 38–51. [Google Scholar] [CrossRef]
- Liu, Q.; Zhang, S.; Mei, S.; Zhou, Y.; Wang, J.; Han, G.-Z.; Chen, L.; Zhou, C.; Cao, M. Viromics unveils extraordinary genetic diversity of the family Closteroviridae in wild citrus. PLOS Pathog. 2021, 17, e1009751. [Google Scholar] [CrossRef]
- Arroyo MT, K.; Marquet, P.A.; Marticorena, C.; Cavieres, L.A.; Squeo, F.A.; Simonetti Zambelli, J.A.; Rozzi, R.; Massardo, F. El hotspot chileno, prioridad mundial para la conservación. In Diversidad de Chile: Patrimonios y Desafíos; Comisión Nacional del Medio Ambiente (Chile): Santiago, Chile, 2006; pp. 94–97. [Google Scholar]
- Echeverria, C.; Coomes, D.; Salas, J.; Rey-Benayas, J.M.; Lara, A.; Newton, A. Rapid deforestation and fragmentation of Chilean Temperate Forests. Biol. Conserv. 2006, 130, 481–494. [Google Scholar] [CrossRef]
- Luebert, F.; Pliscoff, P. Sinopsis Bioclimática y Vegetacional de Chile; Editorial Universitaria: Santiago, Chile, 2006. [Google Scholar]
- Miranda, A.; Altamirano, A.; Cayuela, L.; Lara, A.; González, M. Native Forest loss in the Chilean biodiversity hotspot: Revealing the evidence. Reg. Environ. Chang. 2017, 17, 285–297. [Google Scholar] [CrossRef]
- Schulz, J.J.; Cayuela, L.; Echeverria, C.; Salas, J.; Rey Benayas, J.M. Monitoring land cover change of the dryland forest landscape of Central Chile (1975–2008). Appl. Geogr. 2010, 30, 436–447. [Google Scholar] [CrossRef] [Green Version]
- Garreaud, R.D.; Alvarez-Garreton, C.; Barichivich, J.; Boisier, J.P.; Christie, D.; Galleguillos, M.; LeQuesne, C.; McPhee, J.; Zambrano-Bigiarini, M. The 2010–2015 megadrought in central Chile: Impacts on regional hydroclimate and vegetation. Hydrol. Earth Syst. Sci. 2017, 21, 6307–6327. [Google Scholar] [CrossRef] [Green Version]
- Garreaud, R.D.; Boisier, J.P.; Rondanelli, R.; Montecinos, A.; Sepúlveda, H.H.; Veloso-Aguila, D. The Central Chile Mega Drought (2010–2018): A climate dynamics perspective. Int. J. Climatol. 2020, 40, 421–439. [Google Scholar] [CrossRef]
- Magni, C.R.; Saavedra, N.; Espinoza, S.E.; Yáñez, M.A.; Quiroz, I.; Faúndez, Á.; Martinez-Herrera, E. The recruitment of the recalcitrant-seeded Cryptocarya alba (mol.) looser, established via direct seeding is mainly affected by the seed source and forest cover. Plants 2022, 11, 2918. [Google Scholar] [CrossRef]
- Bandte, M.; Rehanek, M.; Leder, B.; von Bargen, S.; Büttner, C. Identification of an Emaravirus in a Common Oak (Quercus robur L.) Conservation Seed Orchard in Germany: Implications for Oak Health. Forests 2020, 11, 1174. [Google Scholar] [CrossRef]
- Büttner, C.; von Bargen, S.; Bandte, M.; Mühlbach, H.-P. Forest Diseases Caused by Viruses. In Infectious Forest Diseases; Gonthier, P., Nicolotti, G., Eds.; CABI Digital Library: Wallingford, UK, 2013. [Google Scholar]
- Donoso, C. Bosques Templados de Chile y Argentina: Variación, Estructura y Dinámica; Editorial Universitaria: Santiago, Chile, 1993; pp. 151–175. [Google Scholar]
- de Kok, R. Persea lingue (The IUCN Red List of Threatened Species) 2021, e.T32036A68077320. Available online: https://www.iucnredlist.org/species/32036/68077320 (accessed on 3 May 2023).
- Ministerio del Medio Ambiente. Decreto Supremo N° 42/2011. Aprueba y Oficializa Clasificación de Especies Según su Estado de Conservación, Sexto Proceso; Gobierno de Chile: Santiago, Chile; Diario oficial de la República de Chile: Santiago, Chile, 2012.
- Rodríguez, R.; Matthei, S.; Quezada, M. Flora arbórea de Chile; Editorial de la Universidad de Concepción: Santiago, Chile, 1993; p. 407. [Google Scholar]
- Amigo, F.; Flores, L. A new contribution to the syntaxonomy of the sclerophyllous forests and pre-forests of Central Chile: The Lithraeion causticae alliance. Int. J. Geobot. Res. 2013, 3, 47–67. [Google Scholar] [CrossRef]
- Cob, J.; Sabja, A.M.; Ríos, D.; Lara, A.; Donoso, P.J.; Arias, L.; Escobar, B. Potencial de la organogenesis como estrategia para la masificación in vitro de Persea lingue en la zona centro-sur de Chile. Bosque 2010, 31, 202–208. [Google Scholar] [CrossRef] [Green Version]
- Holler, J.G.; Søndergaard, K.; Slotved, H.-C.; Gúzman, A.; Mølgaard, P. Evaluation of the antibacterial activity of Chilean plants traditionally used for wound healing therapy against multidrug-resistant Staphylococcus aureus. Planta Med. 2012, 78, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Lillo, A.; Carvajal-Caiconte, F.; Vital, W.; Silva Junior, P.I.; Paredes-Gamero, E.J.; Alvear, M.; Miranda, A. Bioactive properties of Persea lingue Ness (Lauraceae) fruit and leaf extracts. Braz. J. Biol. 2021, 83, e248063. [Google Scholar] [CrossRef]
- Ibarra, J.T.; Novoa, F.J.; Jaillard, H.; Altamirano, T.A. Large trees and decay: Suppliers of a keystone resource for cavity-using wildlife in old-growth and secondary Andean temperate forests. Austral Ecol. 2020, 45, 1135–1144. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Martelli, G.P.; Agranovsky, A.A.; Bar-Joseph, M.; Boscia, D.; Candresse, T.; Coutts, R.H.A.; Dolja, V.V.; Hu, J.S.; Jelkmann, W.; Karasev, A.V.; et al. Family Closteroviridae. In Virus Taxonomy. Ninth Report of the International Committee on Taxonomy of Viruses; King, A., Adams, M.J., Carstens, E.B., Lefkowitz, E., Eds.; Elsevier-Academic Press: Amsterdam, The Netherlands, 2012; pp. 987–1001. [Google Scholar]
- Abu-Ghanem-Sabanadzovic, N.A.; Sabanadzovic, S.; Gugerli, P.; Rowhani, A. Genome organization, serology and phylogeny of Grapevine leafroll-associated viruses 4 and 6: Taxonomic implications. Virus Res. 2012, 63, 120–128. [Google Scholar] [CrossRef]
- Miranda, A.; Lara, A.; Altamirano, A.; Di Bella, C.; González, M.E.; Camarero, J.J. Forest browning trends in response to drought in a highly threatened mediterranean landscape of South America. Ecol. Indic. 2020, 115, 106401. [Google Scholar] [CrossRef]
- Castillo, M.E.; Plaza, A.J. Proposals of rehabilitation and management actions for the protection of sclerophyllous forests affected by forest fires. Forestist 2021, 72, 112–119. [Google Scholar]
- Miles, L.; Newton, A.C.; Alvarez-Aquino, C.; Armesto, J.J.; del Castillo, R.F.; Cayuela, L.; Echeverría, C.; González-Espinosa, M.; Lara, A.; López-Barrera, F.; et al. Future scenarios for tropical montane and south temperate forest biodiversity in Latin America. In Biodiversity Loss and Conservation in Fragmented Forest Landscapes: The Forests of Montane Mexico and Temperate South America; CABI Digital Library: Wallingford, UK, 2007. [Google Scholar]
- Santelices Moya, R.; Drake, F.; Mena, C.; Ordenes, R.; Navarro-Cerrillo, R. Current and potential distribution areas for Nothofagus alessandrii, an endangered tree species from central Chile. Cienc. Investig. Agrar. 2012, 39, 521–531. [Google Scholar] [CrossRef] [Green Version]
- Alabi, O.J.; Casassa, L.F.; Gutha, L.R.; Larsen, R.C.; Henick-Kling, T.; Harbertson, J.F.; Naidu, R.A. Impacts of Grapevine Leafroll Disease on Fruit Yield and Grape and Wine Chemistry in a Wine Grape (Vitis vinifera L.) Cultivar. PLoS ONE 2016, 11, e0149666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jelkmann, W.; Eastwell, K.C. Little Cherry virus 1 and 2. In Virus and Virus-Like Diseases of Pome and Stone Fruits; Hadidi, A., Barba, M., Candresse, T., Jelkmann, W., Eds.; APS Press: St. Paul, MN, USA, 2011; p. 153. [Google Scholar]
- Wallis, C.M. Potential effects of Grapevine leafroll-associated virus 3 (genus Ampelovirus; family Closteroviridae) or Grapevine red blotch virus (genus Grablovirus; family Geminiviridae) infection on foliar phenolic and amino acid levels. BMC Res. Notes 2022, 15, 213. [Google Scholar] [CrossRef] [PubMed]
- Lemaguet, J.; Beuve, M.; Herrbach, E.; Lemaire, O. Transmission of Six Ampeloviruses and Two Vitiviruses to Grapevine by Phenacoccus aceris. Phytopathology 2011, 102, 717–723. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Virus | ORF1 | RdRp | HSP70 | CP |
|---|---|---|---|---|
| CaAV-1 | 42.30% | 60.80% | 39.50% | 24.10% |
| CaAV-2 | 39.00% | 59.30% | 49.90% | 47.70% |
| GLRaV-1 | 14.90% | 39.80% | 43.30% | 26.40% |
| GLRaV-13 | 14.90% | 36.10% | 40.50% | 24.40% |
| GLRaV-3 | 15.30% | 37.00% | 42.70% | 39.50% |
| LChV-2 | 19.00% | 42.90% | 30.40% | 10.60% |
| PAV-1 | 45.10% | 59.60% | 38.00% | 22.50% |
| PMWaV-2 | 16.10% | 33.40% | 39.50% | 24.90% |
| YaV1 | 25.40% | 44.20% | 30.90% | 14.60% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zamorano, A.; Gamboa, C.; Camilla, C.; Beltrán, F.; Magni, C.; Vaswani, S.; Martínez-Herrera, E.; Fiore, N. A New Species of Ampelovirus Detected in Persea lingue (Ruiz & Pav.) Nees ex Kopp, a Common Tree of the Threatened Chilean Sclerophyll Forest. Forests 2023, 14, 1257. https://doi.org/10.3390/f14061257
Zamorano A, Gamboa C, Camilla C, Beltrán F, Magni C, Vaswani S, Martínez-Herrera E, Fiore N. A New Species of Ampelovirus Detected in Persea lingue (Ruiz & Pav.) Nees ex Kopp, a Common Tree of the Threatened Chilean Sclerophyll Forest. Forests. 2023; 14(6):1257. https://doi.org/10.3390/f14061257
Chicago/Turabian StyleZamorano, Alan, Camila Gamboa, Colombina Camilla, Francisca Beltrán, Carlos Magni, Suraj Vaswani, Eduardo Martínez-Herrera, and Nicola Fiore. 2023. "A New Species of Ampelovirus Detected in Persea lingue (Ruiz & Pav.) Nees ex Kopp, a Common Tree of the Threatened Chilean Sclerophyll Forest" Forests 14, no. 6: 1257. https://doi.org/10.3390/f14061257