
More Effective Protection Supports Male Better Than Female Siblings over Water Deficit in Artificially Bred Poplar Hybrids
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Water Deficit Treatment
2.2. Morphology and Photosynthesis Assay
2.3. Chlorophyll Pigment Content Assay
2.4. Relative Electrolyte Leakage Assay
2.5. ROS Scavenging Enzyme Activity Assay
2.6. Malondialdehyde Content Assay
2.7. RNA Extraction and qRT-PCR Assay
2.8. Statistical Analysis
3. Results
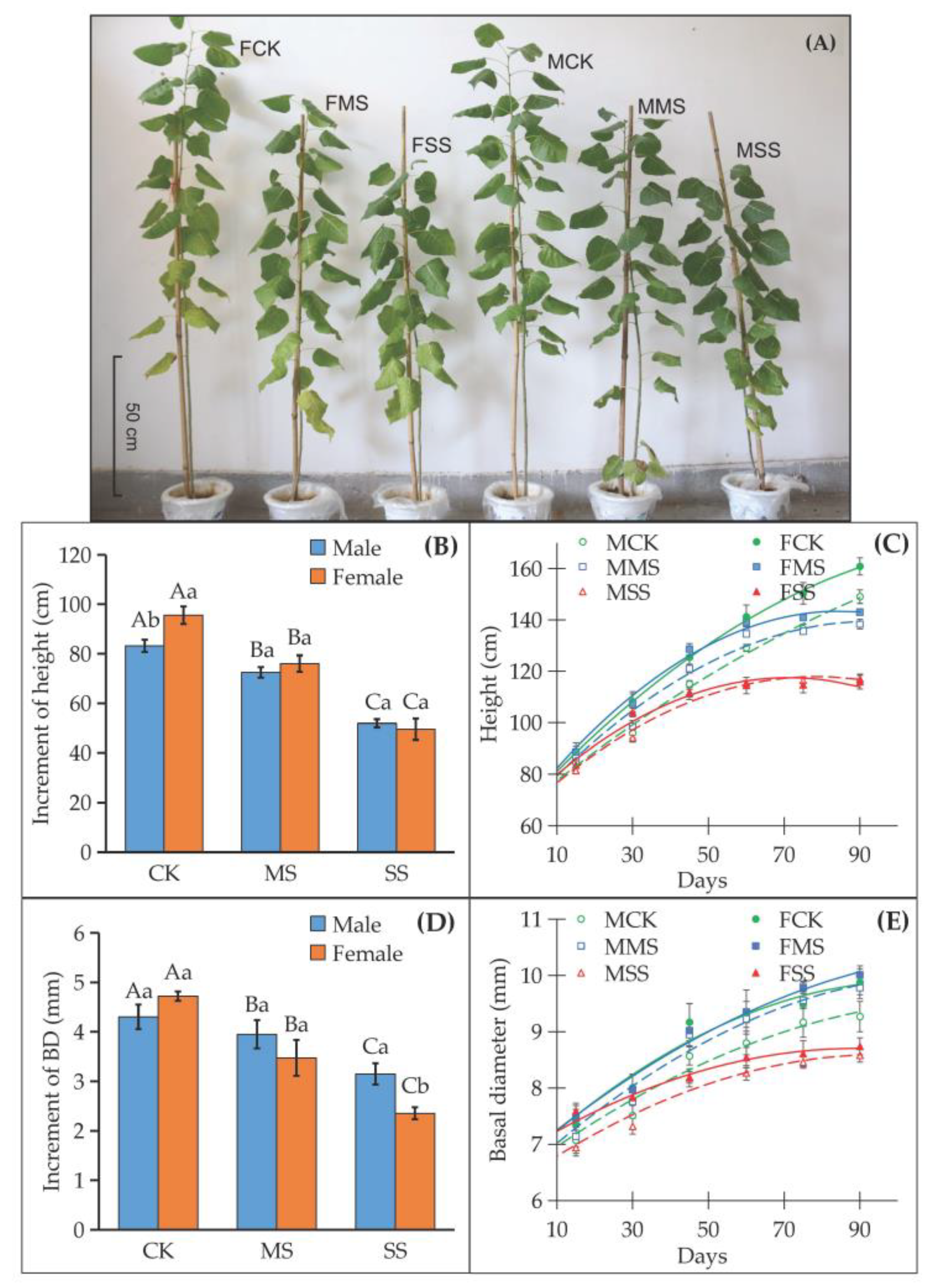
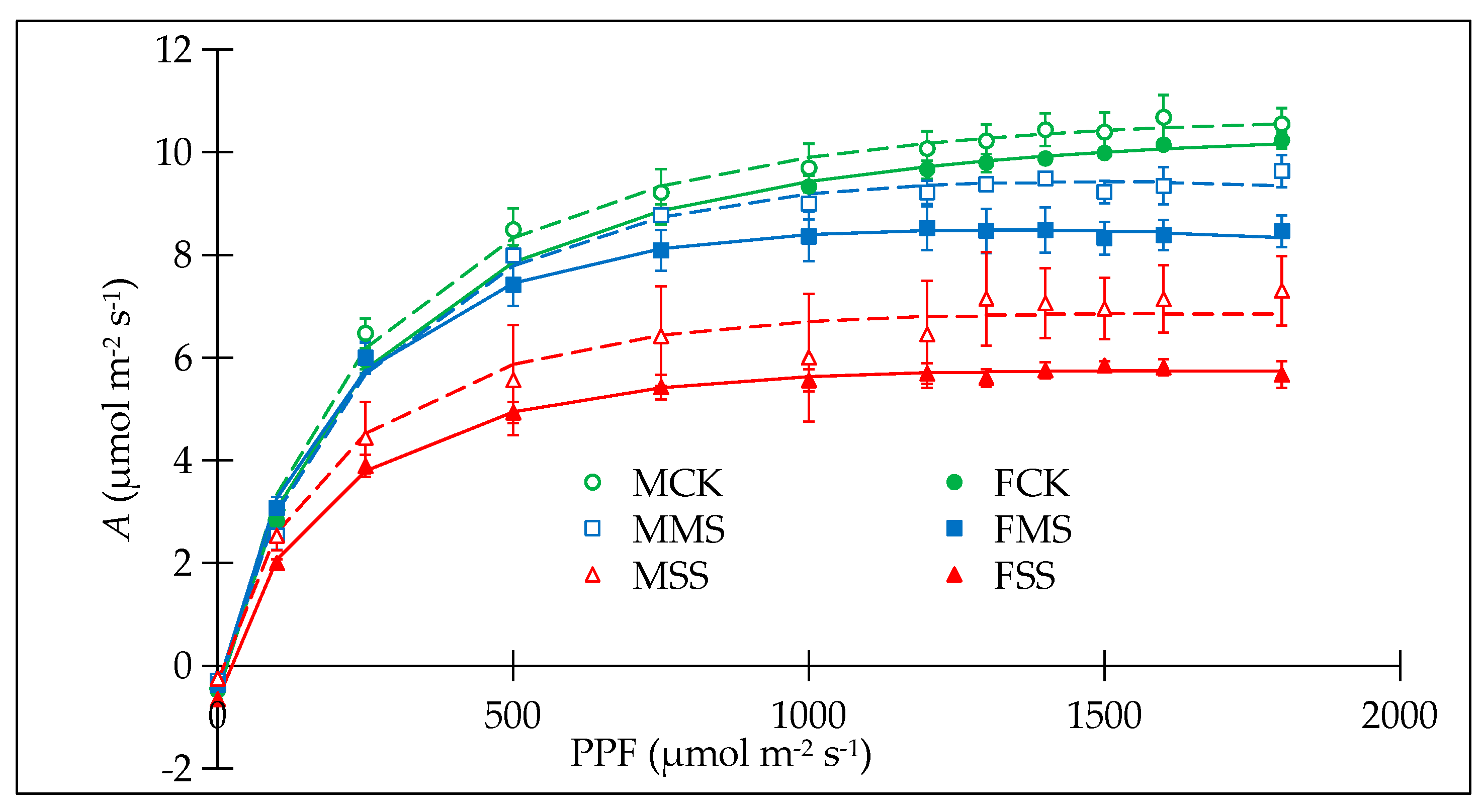
3.1. Water Deficit Had Different Effects on Growth, Net Photosynthesis, and Chlorophyll Fluorescence in Female and Male Poplar Sibs
3.2. Female and Male Poplar Sibs Responded Differently to Light and the Chlorophyll Content Was Reduced Differently by Water Deficit
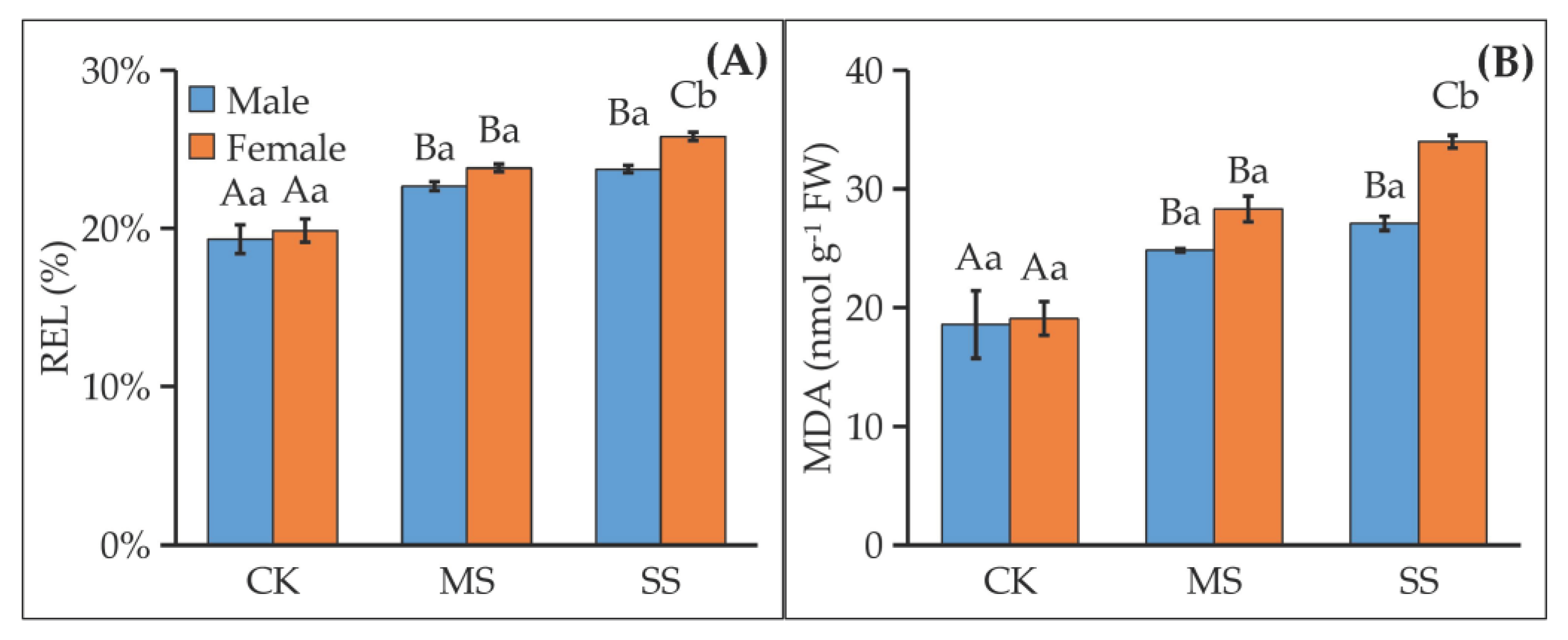
3.3. Water Deficit Damaged Cell Membrane Differently and Induced MDA Contents Differently between Female and Male Sibs
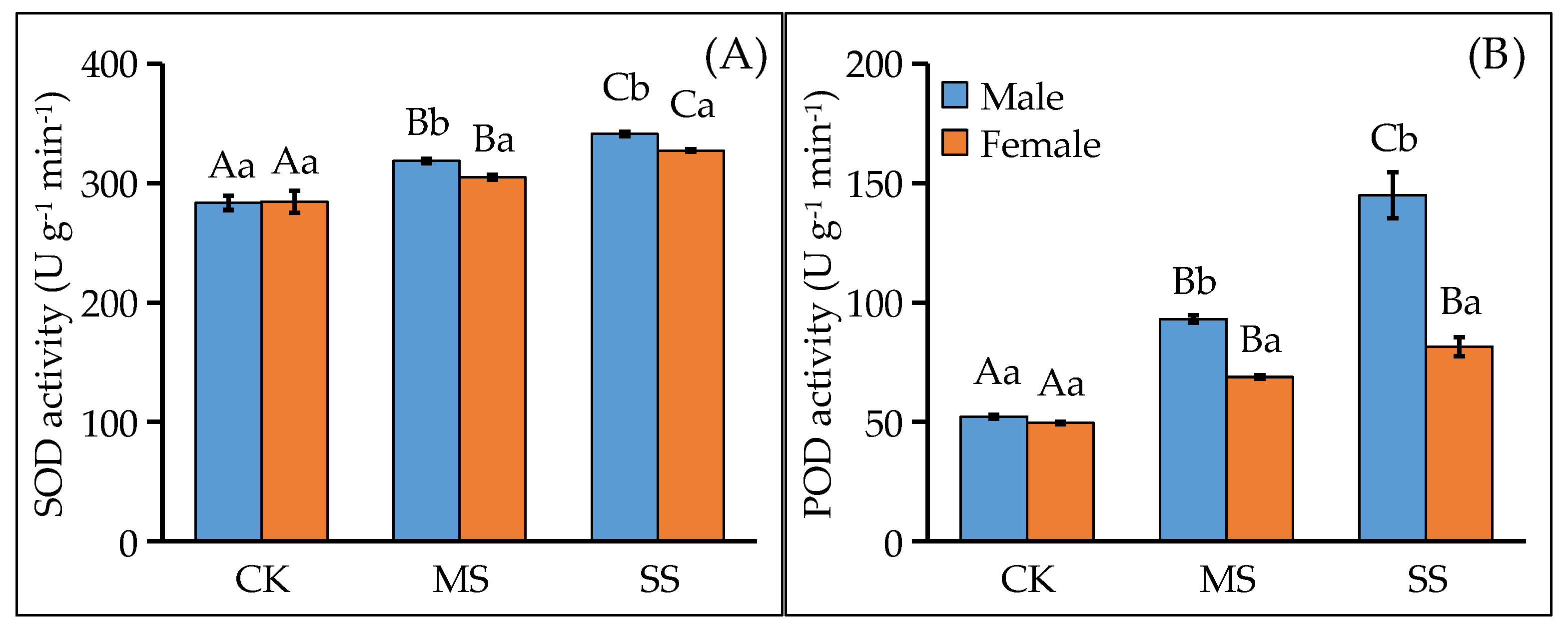
3.4. Water Deficit Induced Different Activities of ROS Scavenging Enzymes
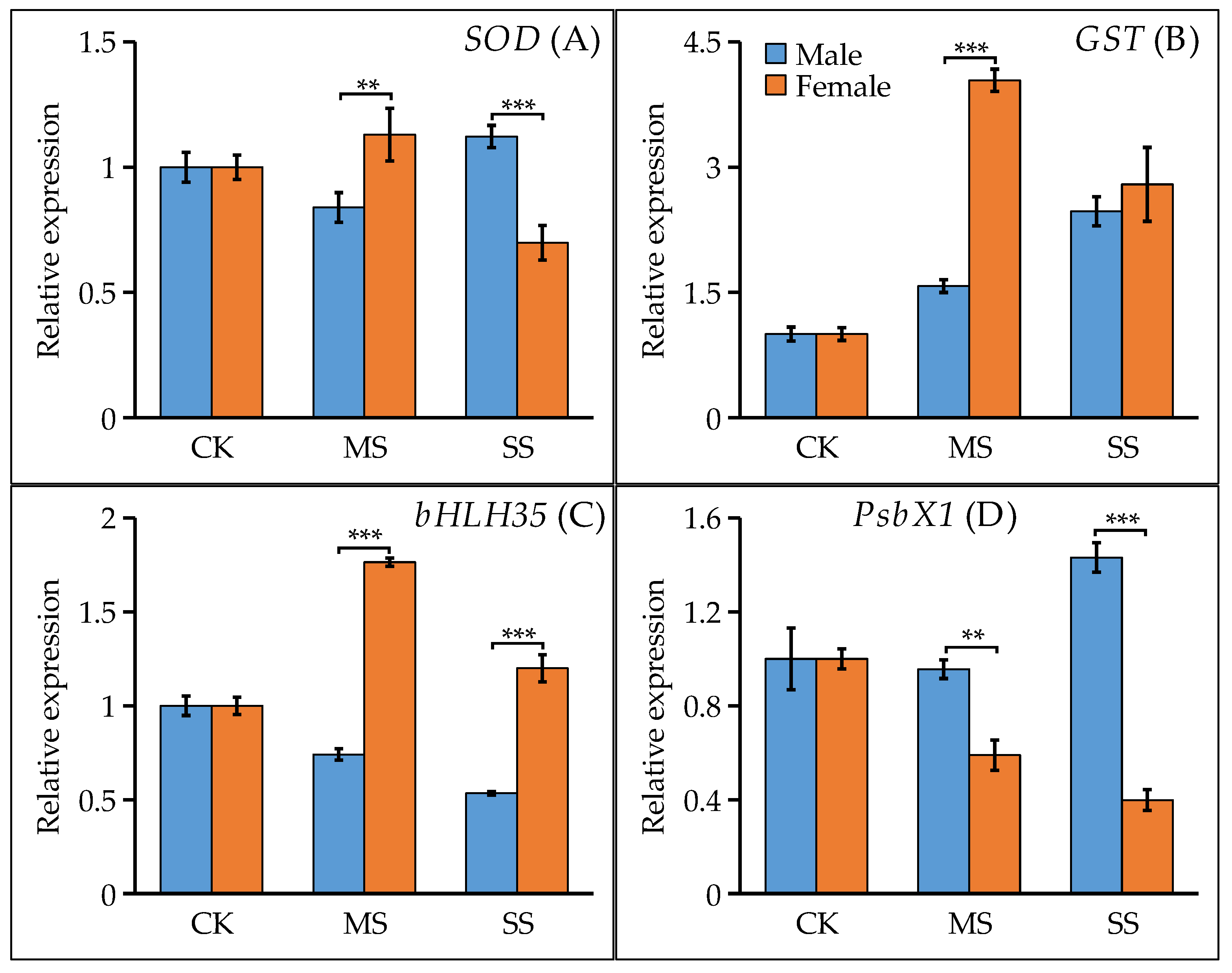
3.5. Water Deficit Regulated Stress-Related Gene Expression Differently in Male and Female Poplar Hybrids
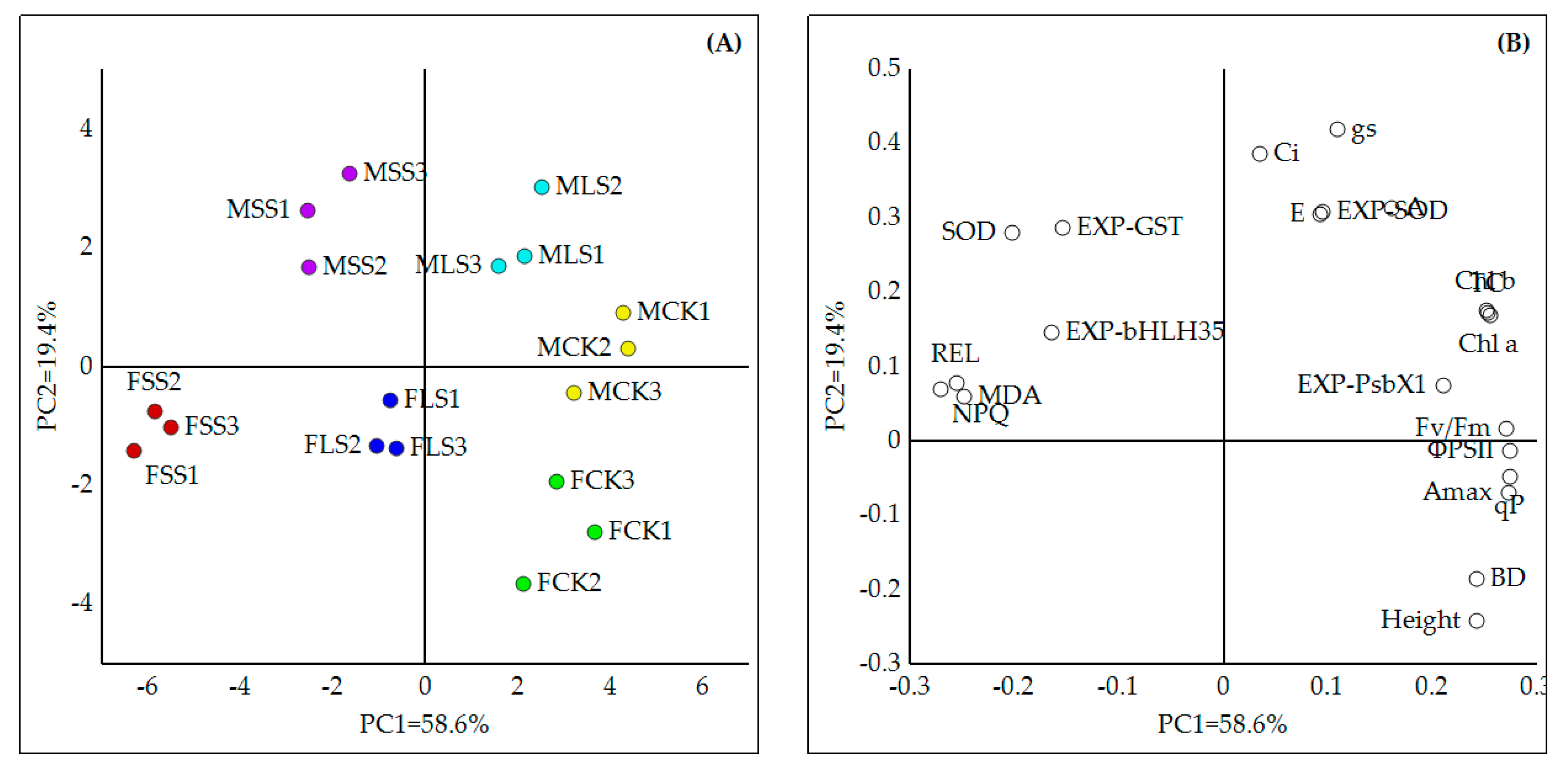
3.6. PCA Displayed Comprehensively Different Responses between Male and Female Sibs to Water Deficit
4. Discussion
4.1. Females Were More Vulnerable to Water Deficit in Growth and Photosynthesis Than Males
4.2. Female Was Damaged More Severely by Water Deficit Due to Its Weaker ROS Scavenging Enzyme Activities
4.3. Water Deficit Regulated Genes Encoding Stress-Related Proteins Differently in Female and Male Lines of P. × euramericana
4.4. PCA Showed an Overall Difference in Water Deficit Response between Female and Male Poplar Sibs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Renner, S.S. The relative and absolute frequencies of angiosperm sexual systems: Dioecy, monoecy, gynodioecy, and an updated online database. Am. J. Bot. 2014, 101, 1588–1596. [Google Scholar] [CrossRef] [PubMed]
- Bachtrog, D.; Mank, J.E.; Peichel, C.L.; Kirkpatrick, M.; Otto, S.P.; Ashman, T.-L.; Hahn, M.W.; Kitano, J.; Mayrose, I.; Ming, R.; et al. Sex Determination: Why So Many Ways of Doing It? PLoS Biol. 2014, 12, e1001899. [Google Scholar] [CrossRef] [PubMed]
- Zemp, N.; Tavares, R.; Muyle, A.; Charlesworth, D.; Marais, G.A.B.; Widmer, A. Evolution of sex-biased gene expression in a dioecious plant. Nat. Plants 2016, 2, 16168. [Google Scholar] [CrossRef] [PubMed]
- Henry, I.M.; Akagi, T.; Tao, R.; Comai, L. One hundred ways to invent the sexes: Theoretical and observed paths to dioecy in plants. Annu. Rev. Plant Biol. 2018, 69, 553–575. [Google Scholar] [CrossRef]
- Darwin, C. The Different Forms of Flowers on Plants of the Same Species; Murray: London, UK, 1877. [Google Scholar] [CrossRef]
- Álvarez-Cansino, L.; Diaz Barradas, M.C.; Zunzunegui, M.; Paz Esquivias, M.; Dawson, T.E. Gender-specific variation in physiology in the dioecious shrub Corema album throughout its distributional range. Funct. Plant Biol. 2012, 39, 968–978. [Google Scholar] [CrossRef]
- Liu, X.; Dong, T.; Zhang, S. Adaptation Responses of Dioecious and Hermaphroditic Tree Species to Abiotic Stress. Forests 2023, 14, 383. [Google Scholar] [CrossRef]
- Barrett, S.C.; Hough, J. Sexual dimorphism in flowering plants. J. Exp. Bot. 2013, 64, 67–82. [Google Scholar] [CrossRef]
- Sun, Z.; Shen, Y.; Niinemets, Ü. Responses of isoprene emission and photochemical efficiency to severe drought combined with prolonged hot weather in hybrid Populus. J. Exp. Bot. 2020, 71, 7364–7381. [Google Scholar] [CrossRef]
- Lin, T.; Tang, J.; He, F.; Chen, G.; Shi, Y.; Wang, X.; Han, S.; Li, S.; Zhu, T.; Chen, L. Sexual differences in above- and belowground herbivore resistance between male and female poplars as affected by soil cadmium stress. Sci. Total Environ. 2022, 803, 150081. [Google Scholar] [CrossRef] [PubMed]
- Choat, B.; Brodribb, T.J.; Brodersen, C.R.; Duursma, R.A.; López, R.; Medlyn, B.E. Triggers of tree mortality under drought. Nature 2018, 558, 531–539. [Google Scholar] [CrossRef]
- Basu, S.; Ramegowda, V.; Kumar, A.; Pereira, A. Plant adaptation to drought stress. F1000Research 2016, 5, 1554. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhu, J.; Gong, Z.; Zhu, J. Abiotic stress responses in plants. Nat. Rev. Genet. 2022, 23, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Devireddy, A.R.; Zandalinas, S.I.; Fichman, Y.; Mittler, R. Integration of reactive oxygen species and hormone signaling during abiotic stress. Plant J. 2021, 105, 459–476. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Sachdev, S.; Ansari, S.A.; Ansari, M.I.; Fujita, M.; Hasanuzzaman, M. Abiotic Stress and Reactive Oxygen Species: Generation, Signaling, and Defense Mechanisms. Antioxidants 2021, 10, 277. [Google Scholar] [CrossRef]
- Mukarram, M.; Choudhary, S.; Kurjak, D.; Petek, A.; Khan, M.M.A. Drought: Sensing, signalling, effects and tolerance in higher plants. Physiol. Plant. 2021, 172, 1291–1300. [Google Scholar] [CrossRef]
- Wahab, A.; Abdi, G.; Saleem, M.H.; Ali, B.; Ullah, S.; Shah, W.; Mumtaz, S.; Yasin, G.; Muresan, C.C.; Marc, R.A. Plants’ Physio-Biochemical and Phyto-Hormonal Responses to Alleviate the Adverse Effects of Drought Stress: A Comprehensive Review. Plants 2022, 11, 1620. [Google Scholar] [CrossRef]
- Wang, X.; Curtis, P.S. Gender-specific responses of Populus tremuloides to atmospheric CO2 enrichment. New Phytol. 2001, 150, 675–684. [Google Scholar] [CrossRef]
- Xu, X.; Yang, F.; Xiao, X.; Zhang, S.; Korpelainen, H.; Li, C. Sex-specific responses of Populus cathayana to drought and elevated temperatures. Plant Cell Environ. 2008, 31, 850–860. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, S.; Zhao, H.; Korpelainen, H.; Li, C. Sex-related adaptive responses to interaction of drought and salinity in Populus yunnanensis. Plant Cell Environ. 2010, 33, 1767–1778. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.B.; Jiang, Y.L.; Chen, K.; Duan, B.; Zhang, S.; Korpelainen, H.; Niinemets, Ü.; Li, C. Reproductive investments driven by sex and altitude in sympatric Populus and Salix trees. Tree Physiol. 2017, 37, 1503–1514. [Google Scholar] [CrossRef]
- Melnikova, N.V.; Borkhert, E.V.; Snezhkina, A.V.; Kudryavtseva, A.V.; Dmitriev, A.A. Sex-Specific Response to Stress in Populus. Front. Plant Sci. 2017, 8, 1827. [Google Scholar] [CrossRef]
- Xia, Z.; He, Y.; Yu, L.; Lv, R.; Korpelainen, H.; Li, C. Sex-specific strategies of phosphorus (P) acquisition in Populus cathayana as affected by soil P availability and distribution. New Phytol. 2020, 225, 782–792. [Google Scholar] [CrossRef]
- Yu, L.; Huang, Z.; Tang, S.; Korpelainen, H.; Li, C.J.E.; Botany, E. Populus euphratica males exhibit stronger drought and salt stress resistance than females. Environ. Exp. Bot. 2023, 205, 105114. [Google Scholar] [CrossRef]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Annu. Rev. Plant Biol. 2006, 57, 781–803. [Google Scholar] [CrossRef]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Gene networks involved in drought stress response and tolerance. J. Exp. Bot. 2006, 58, 221–227. [Google Scholar] [CrossRef]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef]
- Wei, T.; Wang, Y.; Xie, Z.; Guo, D.; Chen, C.; Fan, Q.; Deng, X.; Liu, J. Enhanced ROS scavenging and sugar accumulation contribute to drought tolerance of naturally occurring autotetraploids in Poncirus trifoliata. Plant Biotechnol. J. 2019, 17, 1394–1407. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, Z.; Li, X.; Gao, X.; Dai, Z.; Cui, Y.; Zhi, Y.; Liu, Q.; Zhai, H.; Gao, S.; et al. The IbBBX24–IbTOE3–IbPRX17 module enhances abiotic stress tolerance by scavenging reactive oxygen species in sweet potato. New Phytol. 2022, 233, 1133–1152. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Movahedi, A.; Xu, C.; Sun, W.; Li, L.; Wang, P.; Li, D.; Zhuge, Q. Overexpression of PtHMGR enhances drought and salt tolerance of poplar. Ann. Bot. 2019, 125, 785–803. [Google Scholar] [CrossRef]
- Edwards, R.; Dixon, D.P. Plant Glutathione Transferases. In Methods Enzymology; Sies, H., Packer, L., Eds.; Academic Press: Cambridge, MA, USA, 2005; Volume 401, pp. 169–186. [Google Scholar]
- Gao, S.; Li, C.; Chen, X.; Li, S.; Liang, N.; Wang, H.; Zhan, Y.; Zeng, F. Basic helix-loop-helix transcription factor PxbHLH02 enhances drought tolerance in Populus (Populus simonii × P. nigra). Tree Physiol. 2023, 43, 185–202. [Google Scholar] [CrossRef]
- Liang, B.; Wan, S.; Ma, Q.; Yang, L.; Hu, W.; Kuang, L.; Xie, J.; Huang, Y.; Liu, D.; Liu, Y. A Novel bHLH Transcription Factor PtrbHLH66 from Trifoliate Orange Positively Regulates Plant Drought Tolerance by Mediating Root Growth and ROS Scavenging. Int. J. Mol. Sci. 2022, 23, 15053. [Google Scholar] [CrossRef] [PubMed]
- Abe, H.; Urao, T.; Ito, T.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcriptional activators in abscisic acid signaling. Plant Cell 2003, 15, 63–78. [Google Scholar] [CrossRef] [PubMed]
- García-Cerdán, J.G.; Sveshnikov, D.; Dewez, D.; Jansson, S.; Funk, C.; Schröder, W.P. Antisense Inhibition of the PsbX Protein Affects PSII Integrity in the Higher Plant Arabidopsis thaliana. Plant Cell Physiol. 2008, 50, 191–202. [Google Scholar] [CrossRef]
- Xiao, L.; Liu, X.; Lu, W.; Chen, P.; Quan, M.; Si, J.; Du, Q.; Zhang, D. Genetic dissection of the gene coexpression network underlying photosynthesis in Populus. Plant Biotechnol. J. 2020, 18, 1015–1026. [Google Scholar] [CrossRef]
- Biswas, S.; Eaton-Rye, J.J. PsbX maintains efficient electron transport in Photosystem II and reduces susceptibility to high light in Synechocystis sp. PCC 6803. Biochim. Biophys. Acta (BBA)—Bioenerg. 2022, 1863, 148519. [Google Scholar] [CrossRef]
- Xiao, Y.; Huang, G.; You, X.; Zhu, Q.; Wang, W.; Kuang, T.; Han, G.; Sui, S.-F.; Shen, J.-R. Structural insights into cyanobacterial photosystem II intermediates associated with Psb28 and Tsl0063. Nat. Plants 2021, 7, 1132–1142. [Google Scholar] [CrossRef]
- Jansson, S.; Douglas, C.J. Populus: A model system for plant biology. Annu. Rev. Plant Biol. 2007, 58, 435–458. [Google Scholar] [CrossRef]
- Wullschleger, S.; Weston, D.; Davis, J. Populus responses to edaphic and climatic cues: Emerging evidence from systems biology research. Crit. Rev. Plant Sci. 2009, 28, 368–374. [Google Scholar] [CrossRef]
- Wullschleger, S.D.; Jansson, S.; Taylor, G. Genomics and Forest Biology: Populus Emerges as the Perennial Favorite. Plant Cell 2002, 14, 2651–2655. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Shen, J.; Min, D.; Ke, L.; Tian, X.; Korpelainen, H.; Li, C. Male Populus cathayana than female shows higher photosynthesis and less cellular injury through ABA-induced manganese transporting inhibition under high manganese condition. Trees 2018, 32, 255–263. [Google Scholar] [CrossRef]
- Song, H.; Cai, Z.; Liao, J.; Zhang, S. Phosphoproteomic and Metabolomic Analyses Reveal Sexually Differential Regulatory Mechanisms in Poplar to Nitrogen Deficiency. J. Proteome Res. 2020, 19, 1073–1084. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.F.; Yang, F.; Han, C.Y.; Pu, Y.J.; Ding, Y.; Zhang, J. Sex-specific responses to winter flooding, spring waterlogging and post-flooding recovery in Populus deltoides. Sci. Rep. 2017, 7, 2534. [Google Scholar] [CrossRef]
- Zhang, R.; Liu, J.Y.; Liu, Q.S.; He, H.G.; Xu, X.; Dong, T.F. Sexual differences in growth and defence of Populus yunnanensis under drought stress. Can. J. For. Res. 2019, 49, 491–499. [Google Scholar] [CrossRef]
- Zhang, Y.D.; Virjamo, V.; Sobuj, N.; Du, W.; Yin, Y.; Nybakken, L.; Guo, H.; Julkunen-Tiitto, R. Sex-related responses of European aspen (Populus tremula L.) to combined stress: TiO2 nanoparticles, elevated temperature and CO2 concentration. J. Hazard. Mater. 2018, 352, 130–138. [Google Scholar] [CrossRef]
- Stromme, C.B.; Julkunen-Tiitto, R.; Olsen, J.E.; Nybakken, L. The dioecious Populus tremula displays interactive effects of temperature and ultraviolet-B along a natural gradient. Environ. Exp. Bot. 2018, 146, 13–26. [Google Scholar] [CrossRef]
- Li, C.; Ren, J.; Luo, J.; Lu, R. Sex-specific physiological and growth responses to water stress in Hippophae rhamnoides L. populations. Acta Physiol. Plant. 2004, 26, 123. [Google Scholar] [CrossRef]
- Yang, Z.; Ma, L.; Jia, Z.; Sun, Y. Application on Spss Software in Photosynthetic Light-Response Curve of Quercus Variabilis Leaf. In Recent Advances in Computer Science and Information Engineering; Qian, Z., Cao, L., Su, W., Wang, T., Yang, H., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; Volume 3, pp. 567–573. [Google Scholar]
- Ye, Z.; Suggett, D.J.; Robakowski, P.; Kang, H. A mechanistic model for the photosynthesis–light response based on the photosynthetic electron transport of photosystem II in C3 and C4 species. New Phytol. 2013, 199, 110–120. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. In Methods Enzymology; Academic Press: Cambridge, MA, USA, 1987; Volume 148, pp. 350–382. [Google Scholar]
- Liao, J.; Song, H.F.; Tang, D.T.; Zhang, S. Sexually differential tolerance to water deficiency of Salix paraplesia—A female-biased alpine willow. Ecol. Evol. 2019, 9, 8450–8464. [Google Scholar] [CrossRef]
- Chen, F.; Zhang, S.; Zhu, G.; Korpelainen, H.; Li, C. Populus cathayana males are less affected than females by excess manganese: Comparative proteomic and physiological analyses. Proteomics 2013, 13, 2424–2437. [Google Scholar] [CrossRef]
- Maehly, A.C. The Assay of Catalases and Peroxidases. Methods Biochem. Anal. 1954, 1, 357–424. [Google Scholar]
- Giannopolitis, C.N.; Ries, S.K. Superoxide Dismutases: I. Occurrence in Higher Plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Canning, C.M.; Mood, B.J.; Bonsal, B.; Howat, B.; Laroque, C.P. Comparison of tree-growth drought legacies of three shelterbelt species in the Canadian prairies. Agric. For. Meteorol. 2023, 330, 109317. [Google Scholar] [CrossRef]
- Brodribb, T.J.; Powers, J.; Cochard, H.; Choat, B. Hanging by a thread? Forests and drought. Science 2020, 368, 261–266. [Google Scholar] [CrossRef]
- Hultine, K.R.; Bush, S.E.; Ward, J.K.; Dawson, T.E. Does sexual dimorphism predispose dioecious riparian trees to sex ratio imbalances under climate change? Oecologia 2018, 187, 921–931. [Google Scholar] [CrossRef]
- Morales, M.; Pinto-Marijuan, M.; Munne-Bosch, S. Seasonal, sex- and plant size-related effects on photoinhibition and photoprotection in the dioecious Mediterranean dwarf palm, Chamaerops humilis. Front. Plant Sci. 2016, 7, 1116. [Google Scholar] [CrossRef]
- Korgiopoulou, C.; Bresta, P.; Nikolopoulos, D.; Karabourniotis, G. Sex-specific structural and functional leaf traits and sun-shade acclimation in the dioecious tree Pistacia vera (Anacardiaceae). Funct. Plant Biol. 2019, 46, 649–659. [Google Scholar] [CrossRef]
- Hu, H.; He, B.; Ma, L.; Chen, X.; Han, P.; Luo, Y.; Liu, Y.; Fei, X.; Wei, A. Physiological and transcriptome analyses reveal the photosynthetic response to drought stress in drought-sensitive (Fengjiao) and drought-tolerant (Hanjiao) Zanthoxylum bungeanum cultivars. Front. Plant Sci. 2022, 13, 968714. [Google Scholar] [CrossRef] [PubMed]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [PubMed]
- Helm, L.T.; Shi, H.; Lerdau, M.T.; Yang, X. Solar-induced chlorophyll fluorescence and short-term photosynthetic response to drought. Ecol. Appl. 2020, 30, e02101. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, L. Autofluorescence in Plants. Molecules. 2020, 25, 2393. [Google Scholar] [CrossRef]
- Sherstneva, O.; Khlopkov, A.; Gromova, E.; Yudina, L.; Vetrova, Y.; Pecherina, A.; Kuznetsova, D.; Krutova, E.; Sukhov, V.; Vodeneev, V. Analysis of chlorophyll fluorescence parameters as predictors of biomass accumulation and tolerance to heat and drought stress of wheat Triticum aestivum. Funct. Plant Biol. 2022, 49, 155–169. [Google Scholar] [CrossRef]
- He, F.; Li, H.; Wang, J.; Su, Y.; Wang, H.; Feng, C.; Yang, Y.; Niu, M.; Liu, C.; Yin, W.; et al. PeSTZ1, a C2H2-type zinc finger transcription factor from Populus euphratica, enhances freezing tolerance through modulation of ROS scavenging by directly regulating PeAPX2. Plant Biotechnol. J. 2019, 17, 2169–2183. [Google Scholar] [CrossRef]
- Zhang, S.; Chen, L.; Duan, B.; Korpelainen, H.; Li, C. Populus cathayana males exhibit more efficient protective mechanisms than females under drought stress. For. Ecol. Manag. 2012, 275, 68–78. [Google Scholar] [CrossRef]
- Bowler, C.; Van Montagu, M.; Inzé, D. Superoxide dismutase and stress tolerance. Annu. Rev. Plant Biol. 1992, 43, 83–116. [Google Scholar] [CrossRef]
- Bajji, M.; Kinet, J.-M.; Lutts, S. The use of the electrolyte leakage method for assessing cell membrane stability as a water stress tolerance test in durum wheat. Plant Growth Regul. 2002, 36, 61–70. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Whitlow, T.H.; Bassuk, N.L.; Ranney, T.G.; Reichert, D.L. An improved method for using electrolyte leakage to assess membrane competence in plant tissues. Plant Physiol. 1992, 98, 198–205. [Google Scholar] [CrossRef]
- Qin, F.; Liu, G.; Huang, G.Q.; Dong, T.F.; Liao, Y.M.; Xu, X. Zinc application alleviates the adverse effects of lead stress more in female Morus alba than in males. Environ. Exp. Bot. 2018, 146, 68–76. [Google Scholar] [CrossRef]
- Kumar, V.; Khare, T.; Sharma, M.; Wani, S.H. ROS-Induced Signaling and Gene Expression in Crops Under Salinity Stress. In Reactive Oxygen Species and Antioxidant Systems in Plants: Role and Regulation under Abiotic Stress; Khan, M.I.R., Khan, N.A., Eds.; Springer: Singapore, 2017; pp. 159–184. [Google Scholar]
- Melandri, G.; Abd Elgawad, H.; Floková, K.; Jamar, D.C.; Asard, H.; Beemster, G.T.S.; Ruyter-Spira, C.; Bouwmeester, H.J. Drought tolerance in selected aerobic and upland rice varieties is driven by different metabolic and antioxidative responses. Planta 2021, 254, 13. [Google Scholar] [CrossRef]
- Li, C.; Wan, Y.; Shang, X.; Fang, S. Responses of Microstructure, Ultrastructure and Antioxidant Enzyme Activity to PEG-Induced Drought Stress in Cyclocarya paliurus Seedlings. Forests 2022, 13, 836. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Anee, T.I.; Fujita, M. Glutathione in plants: Biosynthesis and physiological role in environmental stress tolerance. Physiol. Mol. Biol. Plants 2017, 23, 249–268. [Google Scholar] [CrossRef]
- Nianiou-Obeidat, I.; Madesis, P.; Kissoudis, C.; Voulgari, G.; Chronopoulou, E.; Tsaftaris, A.; Labrou, N.E. Plant glutathione transferase-mediated stress tolerance: Functions and biotechnological applications. Plant Cell Rep. 2017, 36, 791–805. [Google Scholar] [CrossRef]
- Yang, G.; Wang, Y.; Xia, D.; Gao, C.; Wang, C.; Yang, C. Overexpression of a GST gene (ThGSTZ1) from Tamarix hispida improves drought and salinity tolerance by enhancing the ability to scavenge reactive oxygen species. Plant Cell Tissue Org. 2014, 117, 99–112. [Google Scholar] [CrossRef]
- Toledo-Ortiz, G.; Huq, E.; Quail, P.H. The Arabidopsis basic/helix-loop-helix transcription factor family. Plant Cell 2003, 15, 1749–1770. [Google Scholar] [CrossRef]
- Dong, Y.; Wang, C.; Han, X.; Tang, S.; Liu, S.; Xia, X.; Yin, W. A novel bHLH transcription factor PebHLH35 from Populus euphratica confers drought tolerance through regulating stomatal development, photosynthesis and growth in Arabidopsis. Biochem. Biophys. Res. Commun. 2014, 450, 453–458. [Google Scholar] [CrossRef]
- Ringnér, M. What is principal component analysis? Nat. Biotechnol. 2008, 26, 303–304. [Google Scholar] [CrossRef] [PubMed]
- Rozas, V.; Le Quesne, C.; Rojas-Badilla, M.; Gonzalez-Reyes, A.; Donoso, S.; Olano, J.M. Climatic cues for secondary growth and cone production are sex-dependent in the long-lived dioecious conifer Araucaria araucana. Agric. For. Meteorol. 2019, 274, 132–143. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | A (μmol m−2 s−1) | Fv/Fm | ΦPSII | qP | |
|---|---|---|---|---|---|
| Male | CK | 9.976 ± 0.208 Aa | 0.824 ± 0.002 Aa | 0.057 ± 0.003 Aa | 0.116 ± 0.005 Aa |
| MS | 9.135 ± 0.391 ABa | 0.823 ± 0.003 Aa | 0.047 ± 0.001 Ba | 0.096 ± 0.001 Ba | |
| SS | 8.008 ± 0.752 Ba | 0.808 ± 0.003 Ba | 0.036 ± 0.001 Ca | 0.075 ± 0.002 Ca | |
| Female | CK | 8.367 ± 0.562 Ab | 0.825 ± 0.002 Aa | 0.053 ± 0.001 Aa | 0.116 ± 0.004 Aa |
| MS | 6.945 ± 0.494 ABb | 0.817 ± 0.002 Ba | 0.039 ± 0.001 Bb | 0.083 ± 0.003 Bb | |
| SS | 5.897 ± 0.676 Bb | 0.799 ± 0.002 Cb | 0.026 ± 0.001 Cb | 0.050 ± 0.004 Cb | |
| P-level | Pstress | 0.001 | 0.000 | 0.000 | 0.000 |
| Psex | 0.000 | 0.018 | 0.000 | 0.000 | |
| Parameters | Amax (μmol m−2 s−1) | Chl a (mg mL−1) | Chl b (mg mL−1) | TC (mg mL−1) | |
|---|---|---|---|---|---|
| Male | CK | 11.045 ± 0.501 Aa | 26.086 ± 0.194 Aa | 47.599 ± 0.358 Aa | 73.686 ± 0.552 Aa |
| MS | 9.473 ± 0.195 Ba | 24.721 ± 0.176 Ba | 45.086 ± 0.328 Ba | 69.806 ± 0.504 Ba | |
| SS | 7.444 ± 0.635 Ca | 21.290 ± 0.139 Ca | 38.787 ± 0.255 Ca | 60.077 ± 0.394 Ca | |
| Female | CK | 10.684 ± 0.400 Aa | 23.134 ± 0.261 Ab | 42.155 ± 0.478 Ab | 65.289 ± 0.738 Ab |
| MS | 8.522 ± 0.371 Ba | 18.008 ± 0.601 Bb | 32.802 ± 1.096 Bb | 50.810 ± 1.697 Bb | |
| SS | 6.059 ± 0.084 Cb | 12.382 ± 0.399 Cb | 22.569 ± 0.706 Cb | 34.951 ± 1.105 Cb | |
| P-level | Pstress | 0.019 | 0.000 | 0.000 | 0.000 |
| Psex | 0.000 | 0.000 | 0.000 | 0.000 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, F.; Gong, Y.; Liu, S.; Wang, Y.; Luo, L.; Zhu, G.; Zhao, H. More Effective Protection Supports Male Better Than Female Siblings over Water Deficit in Artificially Bred Poplar Hybrids. Forests 2023, 14, 995. https://doi.org/10.3390/f14050995
Chen F, Gong Y, Liu S, Wang Y, Luo L, Zhu G, Zhao H. More Effective Protection Supports Male Better Than Female Siblings over Water Deficit in Artificially Bred Poplar Hybrids. Forests. 2023; 14(5):995. https://doi.org/10.3390/f14050995
Chicago/Turabian StyleChen, Fugui, Yu Gong, Shuangyan Liu, Yiyun Wang, Linjie Luo, Guoping Zhu, and Han Zhao. 2023. "More Effective Protection Supports Male Better Than Female Siblings over Water Deficit in Artificially Bred Poplar Hybrids" Forests 14, no. 5: 995. https://doi.org/10.3390/f14050995