
Diurnal and Sex Ratio Flight Activity of Rare Cavity-Dweller Eucnemis capucina Ahrens, 1812 (Coleoptera: Eucnemidae) in Lowland Deciduous Forest: Case Study from Czech Republic

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Data Collection
2.3. Data Analysis
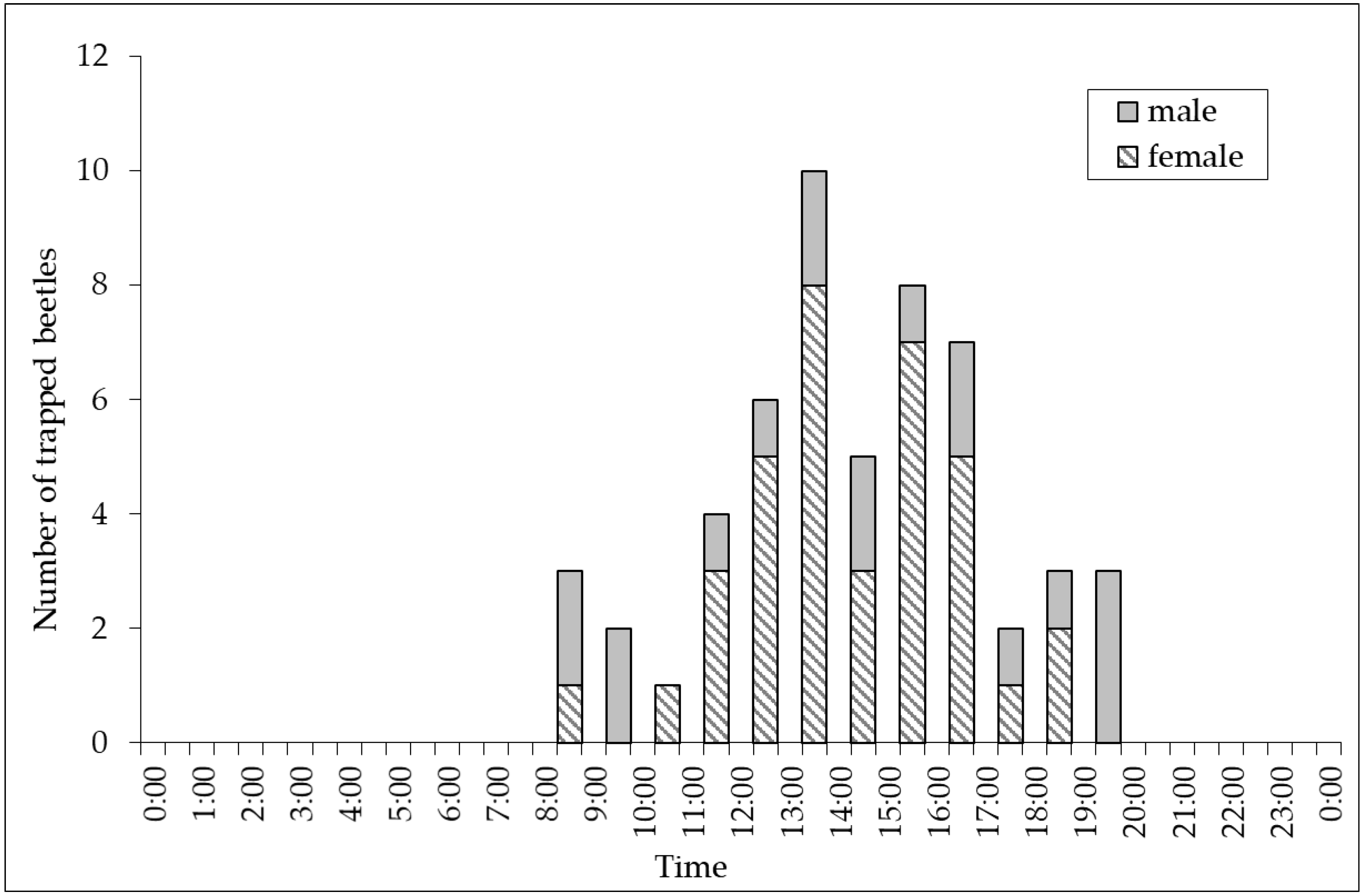
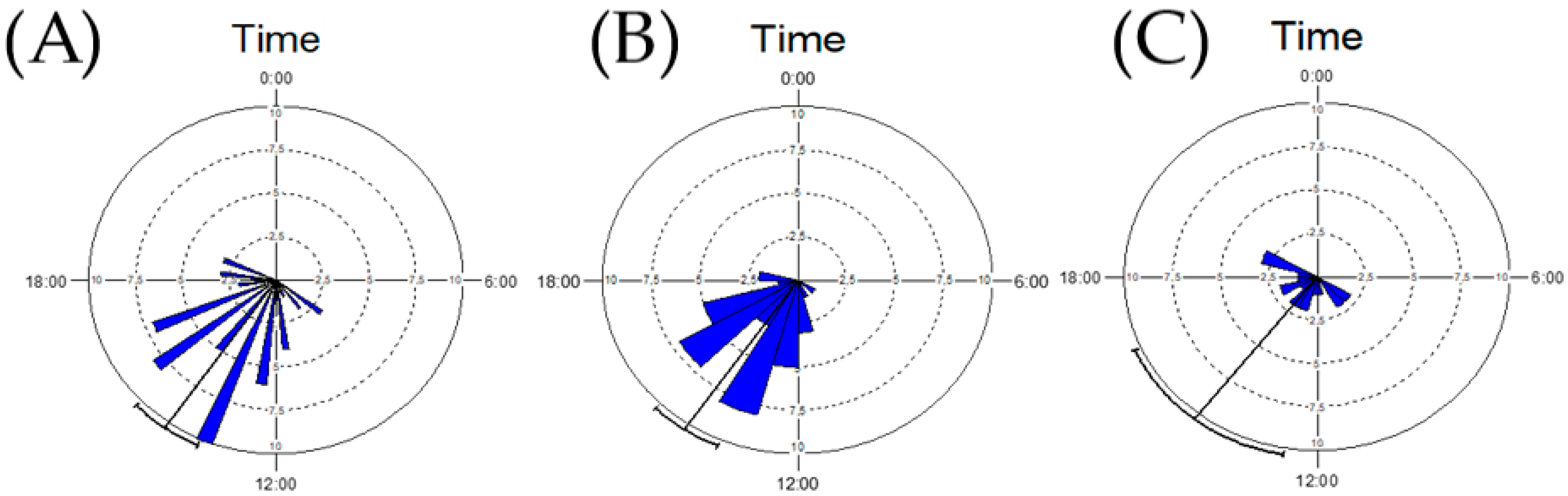
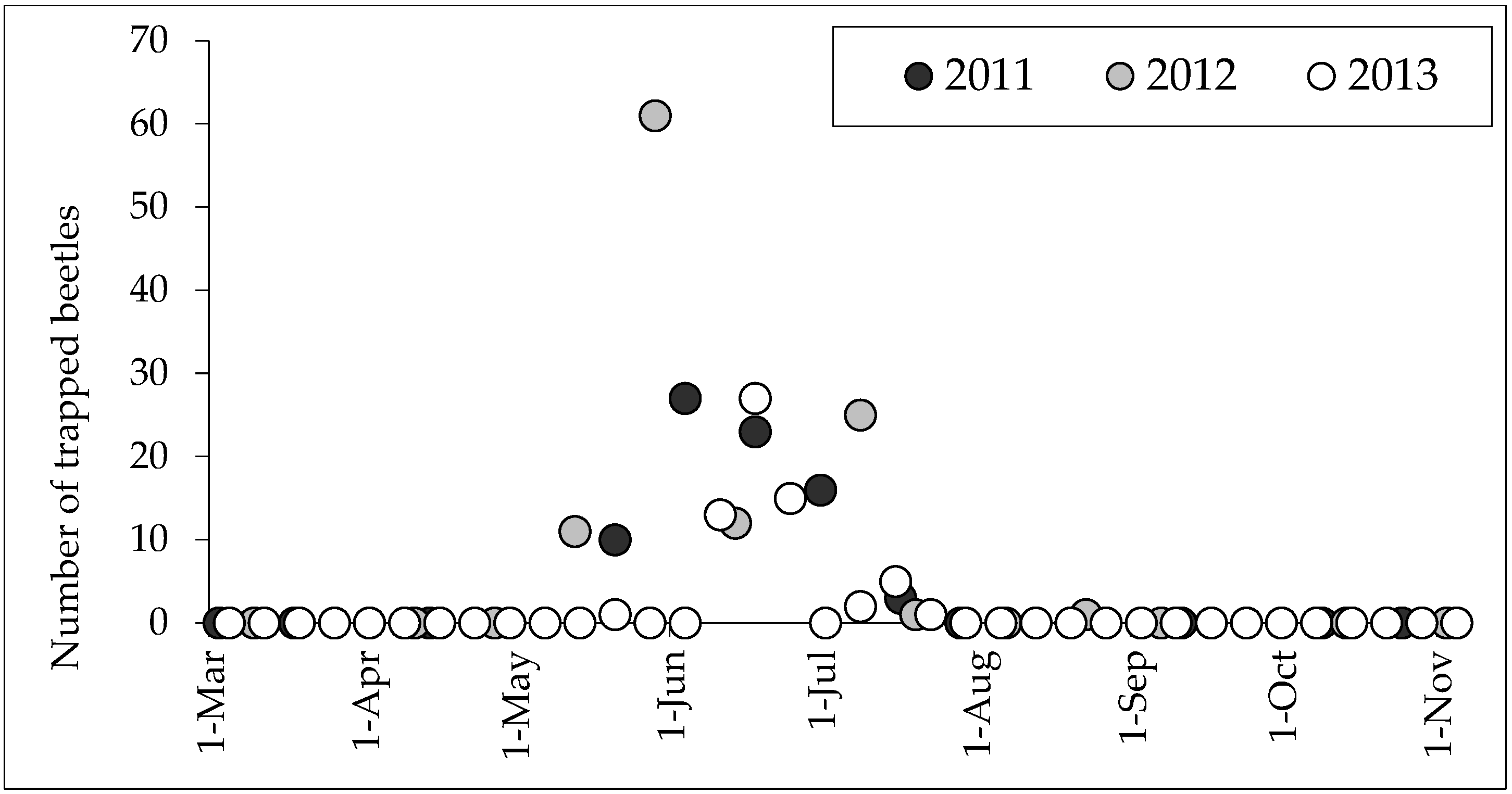
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seibold, S.; Bässler, C.; Brandl, R.; Gossner, M.M.; Thorn, S.; Ulyshen, M.D.; Müller, J. Experimental studies of dead-wood biodiversity—A review identifying global gaps in knowledge. Biol. Conserv. 2015, 191, 139–149. [Google Scholar] [CrossRef]
- Graf, M.; Seibold, S.; Gossner, M.M.; Hagge, J.; Weiß, I.; Bässler, C.; Müller, J. Coverage based diversity estimates of facultative saproxylic species highlight the importance of deadwood for biodiversity. For. Ecol. Manag. 2022, 517, 120275. [Google Scholar] [CrossRef]
- Zumr, V.; Remeš, J.; Nakládal, O. Small-scale spontaneous dynamics in temperate beech stands as an importance driver for beetle species richness. Sci. Rep. 2022, 12, 11974. [Google Scholar] [CrossRef] [PubMed]
- Seibold, S.; Brandl, R.; Buse, J.; Hothorn, T.; Schmidl, J.; Thorn, S.; Müller, J. Association of extinction risk of saproxylic beetles with ecological degradation of forests in Europe. Conserv. Biol. 2015, 29, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Procházka, J.; Schlaghamerský, J. Does dead wood volume affect saproxylic beetles in montane beech-fir forests of Central Europe? J. Insect Conserv. 2019, 23, 157–173. [Google Scholar] [CrossRef]
- Henneberg, B.; Bauer, S.; Birkenbach, M.; Mertl, V.; Steinbauer, M.J.; Feldhaar, H.; Obermaier, E. Influence of tree hollow characteristics and forest structure on saproxylic beetle diversity in tree hollows in managed forests in a regional comparison. Ecol. Evol. 2021, 11, 17973–17999. [Google Scholar] [CrossRef]
- Fridman, J.; Walheim, M. Amount, structure, and dynamics of dead wood on managed forestland in Sweden. For. Ecol. Manag. 2000, 131, 23–36. [Google Scholar] [CrossRef]
- Kapusta, P.; Kurek, P.; Piechnik, Ł.; Szarek-Łukaszewska, G.; Zielonka, T.; Żywiec, M.; Holeksa, J. Natural and human-related determinants of dead wood quantity and quality in a managed European lowland temperate forest. For. Ecol. Manag. 2020, 459, 117845. [Google Scholar] [CrossRef]
- Cálix, M.; Alexander, K.N.A.; Nieto, A.; Dodelin, B. European Red List of Saproxylic Beetles; IUCN: Brussels, Belgium, 2018; p. 19. Available online: http://www.iucnredlist.org/initiatives/europe/publications (accessed on 1 January 2023).
- Seibold, S.; Gossner, M.M.; Simons, N.K.; Blüthgen, N.; Müller, J.; Ambarlı, D.; Ammer, C.; Bauhus, J.; Fischer, M.; Habel, J.C.; et al. Arthropod decline in grasslands and forests is associated with landscape-level drivers. Nature 2019, 574, 671–674. [Google Scholar] [CrossRef]
- Müller, J.; Bütler, R. A review of habitat thresholds for dead wood: A baseline for management recommendations in European forests. Eur. J. For. Res. 2010, 129, 981–992. [Google Scholar] [CrossRef]
- Marhoul, P. Význam červených seznamů a červených knih pro ochranu ohrožených druhů. In Brouci Vázaní na Dřeviny = Beetles Associated with Trees: Sborník Referátů; Horák, J., Ed.; Lesnická práce: Pardubice, Czech Republic, 2008; pp. 58–62. [Google Scholar]
- Nieto, A.; Alexander, K.N.A. European Red List of Saproxylic Beetles; Publications Office of the European Union: Luxembourg, 2010; 45p. [Google Scholar]
- Paillet, Y.; Bergès, L.; Hjältén, J.; Ódor, P.; Avon, C.; Bernhardt-Römermann, M.; Bijlsma, R.-J.; De Bruyn, L.; Fuhr, M.; Grandin, U.; et al. Biodiversity Differences between Managed and Unmanaged Forests: Meta-Analysis of Species Richness in Europe. Conserv. Biol. 2010, 24, 101–112. [Google Scholar] [CrossRef]
- Gao, T.; Nielsen, A.B.; Hedblom, M. Reviewing the strength of evidence of biodiversity indicators for forest ecosystems in Europe. Ecol. Indic. 2015, 57, 420–434. [Google Scholar] [CrossRef]
- Oettel, J.; Lapin, K. Linking forest management and biodiversity indicators to strengthen sustainable forest management in Europe. Ecol. Indic. 2021, 122, 107275. [Google Scholar] [CrossRef]
- Lettenmaier, L.; Seibold, S.; Bässler, C.; Brandl, R.; Gruppe, A.; Müller, J.; Hagge, J. Beetle diversity is higher in sunny forests due to higher microclimatic heterogeneity in deadwood. Oecologia 2022, 198, 825–834. [Google Scholar] [CrossRef]
- Foit, J.; Kašák, J.; Nevoral, J. Habitat requirements of the endangered longhorn beetle Aegosoma scabricorne (Coleoptera: Cerambycidae): A possible umbrella species for saproxylic beetles in European lowland forests. J. Insect Conserv. 2016, 20, 837–844. [Google Scholar] [CrossRef]
- Busse, A.; Cizek, L.; Čížková, P.; Drag, L.; Dvorak, V.; Foit, J.; Heurich, M.; Hubený, P.; Kašák, J.; Kittler, F.; et al. Forest dieback in a protected area triggers the return of the primeval forest specialist Peltis grossa (Coleoptera, Trogossitidae). Conserv. Sci. Pract. 2022, 4, e612. [Google Scholar] [CrossRef]
- Pezzi, M.; Carlomagno, F.; Mendicino, F.; Bonelli, D.; Pelle, R.; Leis, M.; Chicca, M.; Bonacci, T. Pycnomerus italicus (Coleoptera: Zopheridae), an Endemic Endangered Species: A New Report on Its Presence in Southern Italy. Forests 2022, 13, 1838. [Google Scholar] [CrossRef]
- Traylor, C.R.; Caterino, M.S.; Ulyshen, M.D.; Ferro, M.L.; McHugh, J.V. Assessing the Old-Growth Dependency of Two Saproxylic Beetle Species in the Southern Appalachian Mountains. Insect Syst. Divers. 2022, 6, 1–13. [Google Scholar] [CrossRef]
- Buse, J.; Schröder, B.; Assmann, T. Modelling habitat and spatial distribution of an endangered longhorn beetle—A case study for saproxylic insect conservation. Biol. Conserv. 2007, 137, 372–381. [Google Scholar] [CrossRef]
- Kostanjsek, F.; Sebek, P.; Baranova, B.; Jelaska, L.S.; Riedl, V.; Cizek, L. Size matters! Habitat preferences of the wrinkled bark beetle, Rhysodes sulcatus, the relict species of European primeval forests. Insect Conserv. Divers. 2018, 11, 545–553. [Google Scholar] [CrossRef]
- Cizek, L.; Hauck, D.; Miklin, J.; Platek, M.; Kocarek, P.; Olsovsky, T.; Sebek, P. Relict of primeval forests in an intensively farmed landscape: What affects the survival of the hermit beetle (Osmoderma barnabita) (Coleoptera: Scarabaeidae) in pollard willows? J. Insect Conserv. 2021, 25, 407–415. [Google Scholar] [CrossRef]
- Gouix, N.; Sebek, P.; Valladares, L.; Brustel, H.; Brin, A. Habitat requirements of the violet click beetle (Limoniscus violaceus), an endangered umbrella species of basal hollow trees. Insect Conserv. Divers. 2015, 8, 418–427. [Google Scholar] [CrossRef]
- Rukavina, I.; Kostanjšek, F.; Jelaska, S.; Pirnat, A.; Šerić Jelaska, L. Distribution and habitat suitability of two rare saproxylic beetles in Croatia—A piece of puzzle missing for South-Eastern Europe. iForest-Biogeosci. For. 2018, 11, 765–774. [Google Scholar] [CrossRef]
- Mertlik, J. The species of the family Melasidae (Coleoptera: Elateroidea) Czech and Slovak Republics. Elateridarium 2008, 2, 69–137. [Google Scholar]
- Muona, J. Family Eucnemidae. In Catalogue of Palaearctic Coleoptera. Volume 4, Elateroidea Derodontoidea Bostrichoidea Lymexyloidea Cleroidea Cucujoidea; Löbl, I., Smetana, A., Eds.; Apollo Books: Stenstrup, Denmark, 2007; 935p, ISBN 978-87-88757-67-5. [Google Scholar]
- Burakowski, B. Klucze do oznaczania owadów Polski Coleoptera, Cerophytidae, Eucnemidae, Throscidae, Lissomidae. Pol. Tow. Entomol. 1991, 1, 1–91. [Google Scholar]
- Carpaneto, G.M.; Baviera, C.; Biscaccianti, A.B.; Brandmayr, P.; Mazzei, A.; Mason, F.; Battistoni, A.; Teofili, C.; Rondinini, C.; Fattorini, S.; et al. A Red List of Italian Saproxylic Beetles: Taxonomic overview, ecological features and conservation issues (Coleoptera). Fragm. Èntomol. 2015, 47, 53–126. [Google Scholar] [CrossRef]
- Hejda, R.; Farkač, J.; Chobot, K. Red List of Threatened Species of the Czech Republic; Agentura Ochrany Přírody a Krajiny České Republiky: Praha, Czech Republic, 2017; Volume 36, pp. 1–612. ISBN 978-80-88076-53-7.
- Vávra, J.; Škorpík, M. False click beetles (Coleoptera: Eucnemidae) in the Podyjí National Park and surrounding area, with notes to their bionomics. Thayensia 2013, 10, 53–90. [Google Scholar]
- Muona, J. Review of the phylogeny, classification and biology of the family Eucnemidae (Coleoptera). Entomol. Scand. Suppl. 1993, 44, 1–133. [Google Scholar]
- Schmidl, J.; Wurst, C.; Bussler, H. Rote Liste und Gesamtartenliste der “Diversicornia” (Coleop-tera) Deutschlands.(Jürgen Schmidl, Claus Wurst und Heinz Bussler). In Rote Liste gefährdeter Tiere, Pflanzen und Pilze Deutschlands, Band 5: Wirbellose Tiere (Teil 3); Ries, M., Balzer, S., Gruttke, H., Haupt, H., Hofbauer, N., Ludwig, G., Matzke-Hajek, G., Eds.; Münster (Landwirtschaftsverlag): Münster, Germany, 2021; Volume 70, pp. 99–124. [Google Scholar]
- Freude, H.; Harde, K.W.; Lohse, G.A. Die Kafer Mitteleuropas. Band 6. Diversicornia (Lycidae—Byrrhidae); Goecke & Evers Verlag: Kresfeld, Germany, 1979; 367p. [Google Scholar]
- Lepš, J.; Šmilauer, P. Biostatistika; Nakladatelství Jihočeské Univerzity v Českých Budějovicích: České Budějovice, Czech Republic, 2016. [Google Scholar]
- Horak, J.; Vodka, S.; Kout, J.; Halda, J.P.; Bogusch, P.; Pech, P. Biodiversity of most dead wood-dependent organisms in thermophilic temperate oak woodlands thrives on diversity of open landscape structures. For. Ecol. Manag. 2014, 315, 80–85. [Google Scholar] [CrossRef]
- Müller, J.; Brunet, J.; Brin, A.; Bouget, C.; Brustel, H.; Bussler, H.; Förster, B.; Isacsson, G.; Köhler, F.; Lachat, T.; et al. Implications from large-scale spatial diversity patterns of saproxylic beetles for the conservation of European Beech forests. Insect Conserv. Divers. 2013, 6, 162–169. [Google Scholar] [CrossRef]
- Mertlik, J. Faunistics of Crepidophorus mutilatus (Coleoptera: Elateridae) in the Czech Republic and Slovakia. Elateridarium 2014, 8, 36–56. [Google Scholar]
- Brunet, J.; Isacsson, G. Restoration of beech forest for saproxylic beetles—Effects of habitat fragmentation and substrate density on species diversity and distribution. Biodivers. Conserv. 2009, 18, 2387–2404. [Google Scholar] [CrossRef]
- Müller, J.; Jarzabek-Müller, A.; Bussler, H.; Gossner, M.M. Hollow beech trees identified as keystone structures for saproxylic beetles by analyses of functional and phylogenetic diversity. Anim. Conserv. 2014, 17, 154–162. [Google Scholar] [CrossRef]
- Ranius, T.; Hedin, J. The dispersal rate of a beetle, Osmoderma eremita, living in tree hollows. Oecologia 2001, 126, 363–370. [Google Scholar] [CrossRef] [Green Version]
- Graf, M.; Lettenmaier, L.; Müller, J.; Hagge, J. Saproxylic beetles trace deadwood and differentiate between deadwood niches before their arrival on potential hosts. Insect Conserv. Divers. 2022, 15, 48–60. [Google Scholar] [CrossRef]
- Martínez-Pérez, S.; Galante, E.; Micó, E. Sex specificity of dispersal behaviour and flight morphology varies among tree hollow beetle species. Mov. Ecol. 2022, 10, 41. [Google Scholar] [CrossRef]
- Kehat, M.; Wyndham, M. Differences in Flight Behaviour Of Male And Female Nysius Vinitor Bergroth (Hemiptera: Lygaeidae). Aust. J. Èntomol. 1974, 13, 27–29. [Google Scholar] [CrossRef]
- Snäll, N.; Tammaru, T.; Wahlberg, N.; Viidalepp, J.; Ruohomäki, K.; Savontaus, M.-L.; Huoponen, K. Phylogenetic relationships of the tribe Operophterini (Lepidoptera, Geometridae): A case study of the evolution of female flightlessness. Biol. J. Linn. Soc. 2007, 92, 241–252. [Google Scholar] [CrossRef]
- Macek, J.; Dvořák, J.; Traxler, L.; Červenka, V. Motýli a housenky střední Evropy Noční motýli I; Academia: Praha, Czech Republic, 2007. [Google Scholar]
- Griffin, M.J.; Holwell, G.I.; Symonds, M.R.E. Sex ratio and female allocation to harems in a polygynous bark beetle. Austral Èntomol. 2019, 59, 149–155. [Google Scholar] [CrossRef]
- van Tol, R.; Helsen, H.; Griepink, F.; de Kogel, W. Female-induced increase of host-plant volatiles enhance specific attraction of aphid male Dysaphis plantaginea (Homoptera: Aphididae) to the sex pheromone. Bull. Èntomol. Res. 2009, 99, 593–602. [Google Scholar] [CrossRef] [Green Version]
- Schebeck, M.; Schopf, A.; Ragland, G.J.; Stauffer, C.; Biedermann, P.H.W. Evolutionary ecology of the bark beetles Ips typographus and Pityogenes chalcographus. Bull. Èntomol. Res. 2023, 113, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Torres-Vila, L. Reproductive biology of the great capricorn beetle, Cerambyx cerdo(Coleoptera: Cerambycidae): A protected but occasionally harmful species. Bull. Èntomol. Res. 2017, 107, 799–811. [Google Scholar] [CrossRef] [PubMed]
- Cuff, J.P.; Windsor, F.M.; Gilmartin, E.C.; Boddy, L.; Jones, T.H. Influence of European Beech (Fagales: Fagaceae) Rot Hole Habitat Characteristics on Invertebrate Community Structure and Diversity. J. Insect Sci. 2021, 21, 7. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Statistical Characteristics | Both Sexes | Males | Females |
|---|---|---|---|
| Top of flight activity (time) | 14:20 | 14:31 | 14:16 |
| Circular Standard Deviation | 2:48 | 3:53 | 2:11 |
| N | 54 | 18 | 36 |
| Circular Variance | 2.478 | 0.405 | 0.152 |
| Length of Mean Vector (r) | 0.763 | 0.595 | 0.848 |
| Rayleigh Test (Z) | 31.462 | 6.366 | 25.893 |
| Rayleigh Test (p-value) | <10−12 | 0.001 | 2.41 × 10−11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakládal, O.; Synek, J.; Zumr, V. Diurnal and Sex Ratio Flight Activity of Rare Cavity-Dweller Eucnemis capucina Ahrens, 1812 (Coleoptera: Eucnemidae) in Lowland Deciduous Forest: Case Study from Czech Republic. Forests 2023, 14, 720. https://doi.org/10.3390/f14040720
Nakládal O, Synek J, Zumr V. Diurnal and Sex Ratio Flight Activity of Rare Cavity-Dweller Eucnemis capucina Ahrens, 1812 (Coleoptera: Eucnemidae) in Lowland Deciduous Forest: Case Study from Czech Republic. Forests. 2023; 14(4):720. https://doi.org/10.3390/f14040720
Chicago/Turabian StyleNakládal, Oto, Jiří Synek, and Václav Zumr. 2023. "Diurnal and Sex Ratio Flight Activity of Rare Cavity-Dweller Eucnemis capucina Ahrens, 1812 (Coleoptera: Eucnemidae) in Lowland Deciduous Forest: Case Study from Czech Republic" Forests 14, no. 4: 720. https://doi.org/10.3390/f14040720