
Effects of Pine Plantation on Native Ecuadorian Páramo Vegetation

1
Facultad de Ciencias Biológicas, Universidad Central del Ecuador, Quito 170403, Ecuador
2
Earth and Life Institute, Université Catholique de Louvain, Croix-du-Sud 2 Box L7.05.14–B, 1348 Louvain-la-Neuve, Belgium
*
Author to whom correspondence should be addressed.
Forests 2022, 13(9), 1499; https://doi.org/10.3390/f13091499
Submission received: 8 August 2022
/
Revised: 25 August 2022
/
Accepted: 6 September 2022
/
Published: 16 September 2022
Abstract
:Exotic pine plantations, installed mainly for timber production and carbon sequestration purposes, cover an ever-expanding area of equatorial and tropical regions; however, their economic and environmental benefits are under debate due to their potential negative effects on native plant species conservation. The native understory plant species richness and cover under pine (Pinus radiata D. Don) plantations were compared with natural grasslands in the Ecuadorian Páramo. We analyzed the vegetation in four zones: Antisana (8-year-old pines), Cajas (16-year-old pines), Tisaleo (41-year-old pines) and Cotopaxi (53-year-old pines). The total understory plant cover decreased between 29% and 90% under pine plantations in all zones. The mean species richness in the pine plantations decreased by 44% in Antisana, Cajas and Tisaleo but not in Cotopaxi. Pine plantations strongly reduced the abundance of herbaceous light-demanding species (59%), except small herbs whose cover increased under pine (17.6%). Shrub cover was also negatively affected in Tisaleo and Cotopaxi (7.4%). Pine afforestation effects on Páramo vegetation depend mainly on canopy cover which changes with age and growing conditions (altitude) and secondarily on the vegetation state at the time of planting, resulting from land use history and ecological conditions. The pine effect was therefore more pronounced in the young and dense plantations of Antisana and Cajas than in the old and open stand of Cotopaxi, whose understory vegetation was more similar to that of grasslands.
1. Introduction
Afforested zones are still expanding and cover ca. 290 million hectares worldwide, which represents around 7% of the total forest cover [1]. The main benefits of these plantations are wood production for the construction and paper industries, as well as energy sources [2]. Approximately 24% of these plantations have an environmental goal, such as protection against erosion, rehabilitation of degraded soils and carbon sequestration [3,4,5,6,7]. Plantations of exotic species have facilitated forest succession on sites where high disturbances prevent natural regeneration [8,9]. Plantations typically utilize exotic fast-growing species such as pines and eucalypts [10,11], which are adapted to a wide range of ecological conditions [12,13].
In South America, more than 5 million hectares have been planted with Pinus species [14,15], including 1.64 million hectares in Brazil [16], 1.27 million hectares in Chile [17] and 0.7 million hectares in Argentina [18]. Pine plantations were also established in the Andean mountains of Perú and Ecuador [4,19].
Most of the pine plantations, mainly Pinus radiata and P. patula Schltdl. & Cham., in the Andean mountains were established in zones degraded by overgrazing or burning to prevent further land degradation. Such plantations might constitute a rehabilitation technique in highly degraded grasslands, because they are expected to favor the seedling survival of typical native species by improving the microclimate conditions below the canopy [20,21,22]. They are also considered to be an opportunity to enhance ecosystem services, such as carbon sequestration, and to provide additional outcomes to local peasant communities through the production of firewood and timber [23,24,25]. Furthermore, pine plantations in degraded highlands contribute to ecological functions such as soil stability and minimize the risk of landslides in steep slope conditions [21,26]. Some pine plantations have shown higher species richness than adjacent high-altitude grasslands [27]. Moreover, exotic Pinus radiata plantations increased species richness and floristic composition in restoration experiences at forest altitudes in Tenerife, Canary Island [28].
The high-altitude tropical grasslands in the Andean mountains, ranging from 3200 m to 4700 m a.s.l., constitute a highly diverse and unique ecosystem called the “Páramo” [29,30]. In the Ecuadorian Páramo, 1524 species of vascular plants, including 628 endemic species, have been registered [31]. Páramo grasslands provide several ecosystem services for more than 100 million people, including food production, water regulation and supply, carbon storage and biodiversity conservation [32,33,34]. Extensive cattle grazing and timber plantations among other land uses transformed about 40% of the original Páramo into agroecosystems [31]. About 80% (213 species) of the endemic vascular plants in the Ecuadorian Páramo are threatened according to the International Union for Conservation of Nature (IUCN) [35]. The main threat is the agricultural land degradation caused by burning, overgrazing and plantations of exotic species.
In this context, the impact of pine plantations in Páramo regions is controversial [36,37,38]. Introducing pine trees could have negative effects on the plant community by affecting the diversity and the abundance of native species [39,40,41]. Decreases in Páramo grassland main species cover have been reported under pine plantations [42,43,44]. Afforestation also affects floristic composition by modifying microclimate conditions and resource availability [27,45,46]. The microclimate changes in the understory depend on tree species and age, stand density and, eventually, intensity of pruning [4,34]. Generally, in Páramo, pine afforestation favors shade-tolerant species to the detriment of light-demanding species with a marked influence on understory life forms [30,47,48]. In some rare cases, a higher plant species richness was observed under plantations compared with natural grasslands but with changes in dominant life forms [27,46].
Our research aims to determine the changes in understory native vegetation caused by Pinus radiata D. Don plantations in four Ecuadorian Páramo grassland zones, Antisana, Cajas, Tisaleo and Cotopaxi. We hypothesized that (1) exotic Pinus radiata plantations reduce the plant cover and species richness of understory species, and (2) that this change is dependent on the zone and/or age of the pine plantations.
2. Materials and Methods
2.1. Study Zones
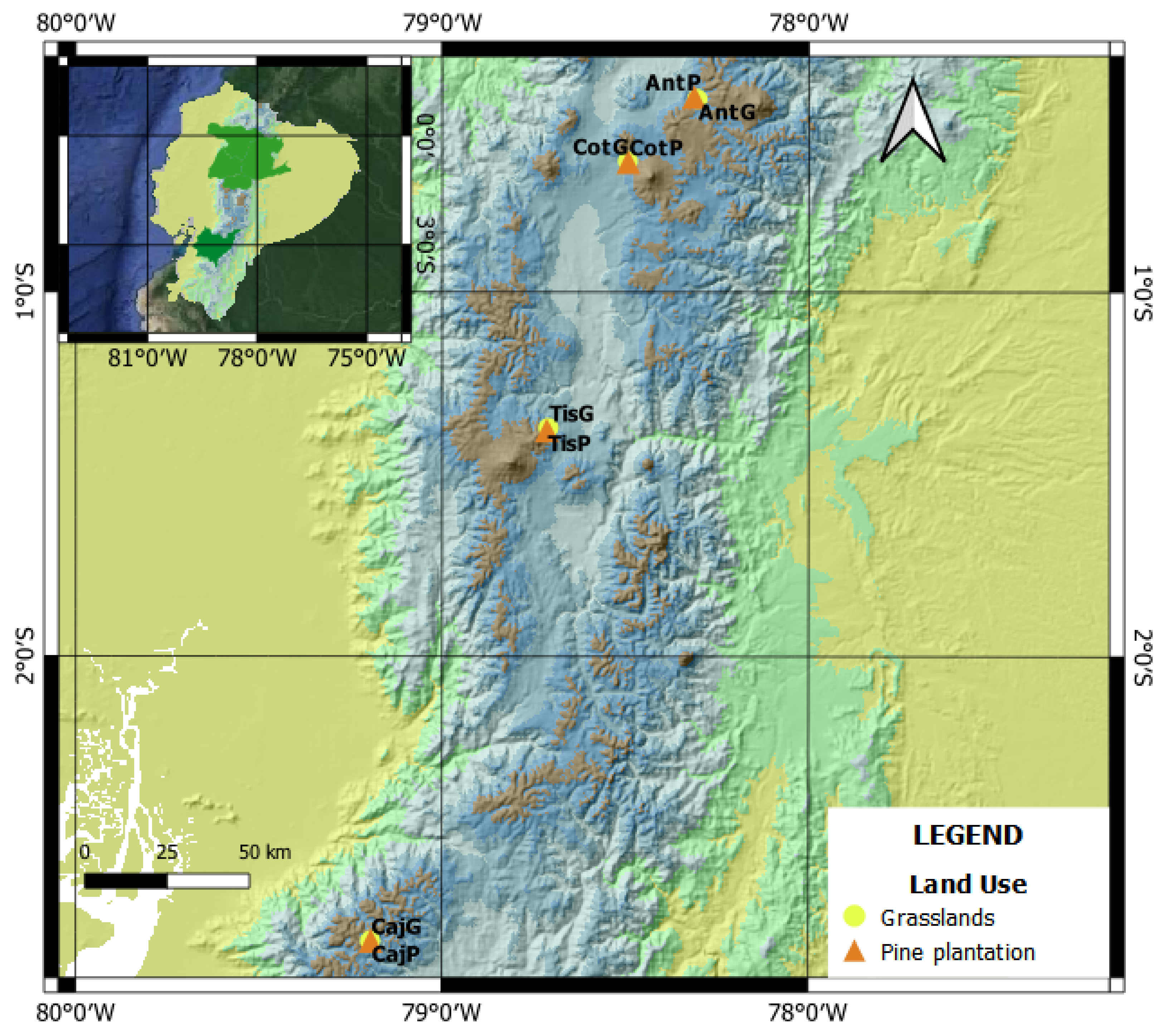
The study was carried out in four zones of Páramo situated over 3500 m a.s.l., in the Ecuadorian Andes (Figure 1; Table 1). The “grass Páramo”, or “Páramo grassland”, is found above the high-altitude woody vegetation, and is dominated by tussock grasses belonging mainly to the genera Calamagrostis, Agrostis and Festuca [49]. The main Angiosperm families in this region include the Asteraceae (850 species), Poaceae (230 species) and Orchidaceae (150 species) [29]. In these zones, Monterrey pine (Pinus radiata) was planted for timber production, without any prior clearing or burning of the existing vegetation. All plantations are currently abandoned without pruning or thinning. The slope of the four pine plantations were locally irregular, ranging from 10 to 50 degrees. Trees were planted with spacing of about 3 m in all directions.
We have chosen four zones in very similar ecological conditions, the main difference between them being the age of the pine plantation. Each pine plantation is located less than 1 km from their correspondent grassland. The youngest plantation is located in Antisana (8 years old), while the oldest is in Cotopaxi (53 years old); the plantations in Cajas (16 years old) and Tisaleo (41 years old) have intermediate ages. In these plantations, basal area increased with age to reach a maximum around 30 m2/ha, while the canopy cover strongly increased during the first 15–20 years before progressively declining due to self-thinning and reduction of the crown-to-stem diameter ratio (Supplementary Figure S1, Table 1). Land use history is similar in the four study zones, always linked to livestock activities with small differences in intensity and frequency. Currently, our study zones are protected by the government or landowners.
2.2. Experimental Design and Data Collection
Vegetation surveys were carried out from July 2018 to December 2020. We established paired sampling areas to compare the vegetation in native Páramo grasslands and under pine plantations. Nine plots of 10 × 10 m were established at least 50 m apart for each land use (pine plantation vs. grassland) in the four study zones. The plots were divided into 2 × 2 m quadrats, among which six were randomly selected to perform the vegetation surveys [54,55]. In each quadrat, the cover estimation was made using a 10 by 10 cm grid of 1 m2. One sample of each species was taken for identification in the herbaria. Most vascular plants were identified to the species level, except for some young or grazed non-flowering individuals that were identified at the genus or family level. The average cover of mosses, fungi and lichens was also assessed [56].
In the pine plantations, pine tree cover was assessed at the four corners and in the center of each 10 × 10 m plot, using a convex spherical densiometer (Forestry Suppliers, Model A, Rockledge, FL, USA) [57], and their average was used for the analyses. The basal area was obtained based on the diameter at breast height of all trees in the plot.
2.3. Data Analysis
Plant life forms were classified into five classes, adapted from the classification of Ramsay and Oxley [58]. Tussock or bunch grasses have erected tillers produced from dense culms at the soil surface. Cushions include soft mat cushions and hard, compact bolster plants, all of them profusely branched with short internodes. The acaulescent or basal rosettes have leaves attached to the same level, at or below ground level. Creeping and prostrate herbaceous species lack erect, leafy stems and possess stolon or rhizomes for vegetative reproduction. Upright and dwarf shrubs, or woody species, present thin and distinctly woody branches with thin bark. Species were also categorized as herbaceous or shrub species and according to their shade tolerance [59,60,61,62].
To evaluate the extent to which the two land uses (grassland vs. pine plantation) were characterized by distinct plant communities while taking the variations among study zones into account, canonical correspondence analysis (CCA), linear models (LM), generalized nonlinear models (GNLM) and a phytosociological analysis (IndVal) were performed. All statistical analyses were performed at a significance level of 95%, Bonferroni corrections were applied when multiple tests had been performed.
For the CCA, the analysis was based on the cover of each plant species (active variables), while four environmental and stand properties known to influence plant community were retained as illustrative variables: altitude, stand age, basal area and canopy cover. LMs were applied to the cover and richness (dependent variables) of the various plant types (mosses, herbs, shrubs, total) and of the different life forms of the herbaceous species (tussocks, cushions, rosettes, small herbs) to highlight the effects of the study zone, the land use and their interaction (independent variables); no variable was used as covariable:
plant type cover or richness = zone + land use + zone × Land use
BoxCox transformations were used for all cover variables since they did not follow a normal distribution. Contrast analyses were then applied for evaluating differences between land uses within study zones. As pine cover present a major role in explaining the effects of pine plantations on moss, herbaceous and shrub cover and richness, the quantitative relationships between their cover/richness and pine cover was analyzed by means of GNLMs (k represents the parameters of the exponential function):
plant type cover or richness ~ k1 + k2 exp (k3 pine cover)
From a taxonomic perspective, a statistical phytosociological analysis was performed based on life forms and shade tolerance of plant species. For this statistical analysis, plant cover values were transformed in numeric coefficients using the van der Maarel transformation [63]. To characterize the plant species community in each zone and land use, indicator species were identified according to the method IndVal [64]. An indicator species is mostly found in a single group of the typology (specificity) and is present in the majority of the sites belonging to this group (fidelity).
Data treatment was achieved using R program (version 4.0.3, R Core Team, Vienna, Austria) and several of its packages: “vegan” for the CCA, “stats” for the LMs, “MASS” for BoxCox transformation when the variables did not show normality, “contrast” to detect differences between land uses within zones, “nlme” and “nlraa” for GNLM and figures and “indicspecies” to apply IndVal and identify the indicator species.
3. Results
A total of 168 species belonging to 95 genera and 41 families were found in our four study zones on grasslands and under pine plantations, 14 families were represented by only one species and 14 species were found only in one quadrat. Asteraceae (42 species) and Poaceae (10 species) were the most represented families, followed by Rosaceae (9 species) and Apiaceae and Cyperaceae, both with 8 species. A total of 118 species were found under the four pine plantations and 48 were exclusive of them. In grasslands, a total of 119 species were found with 49 species specific to this land use.
3.1. Multivariate Analysis (CCA) Illustrating Pine Plantation Effects on Plant Community
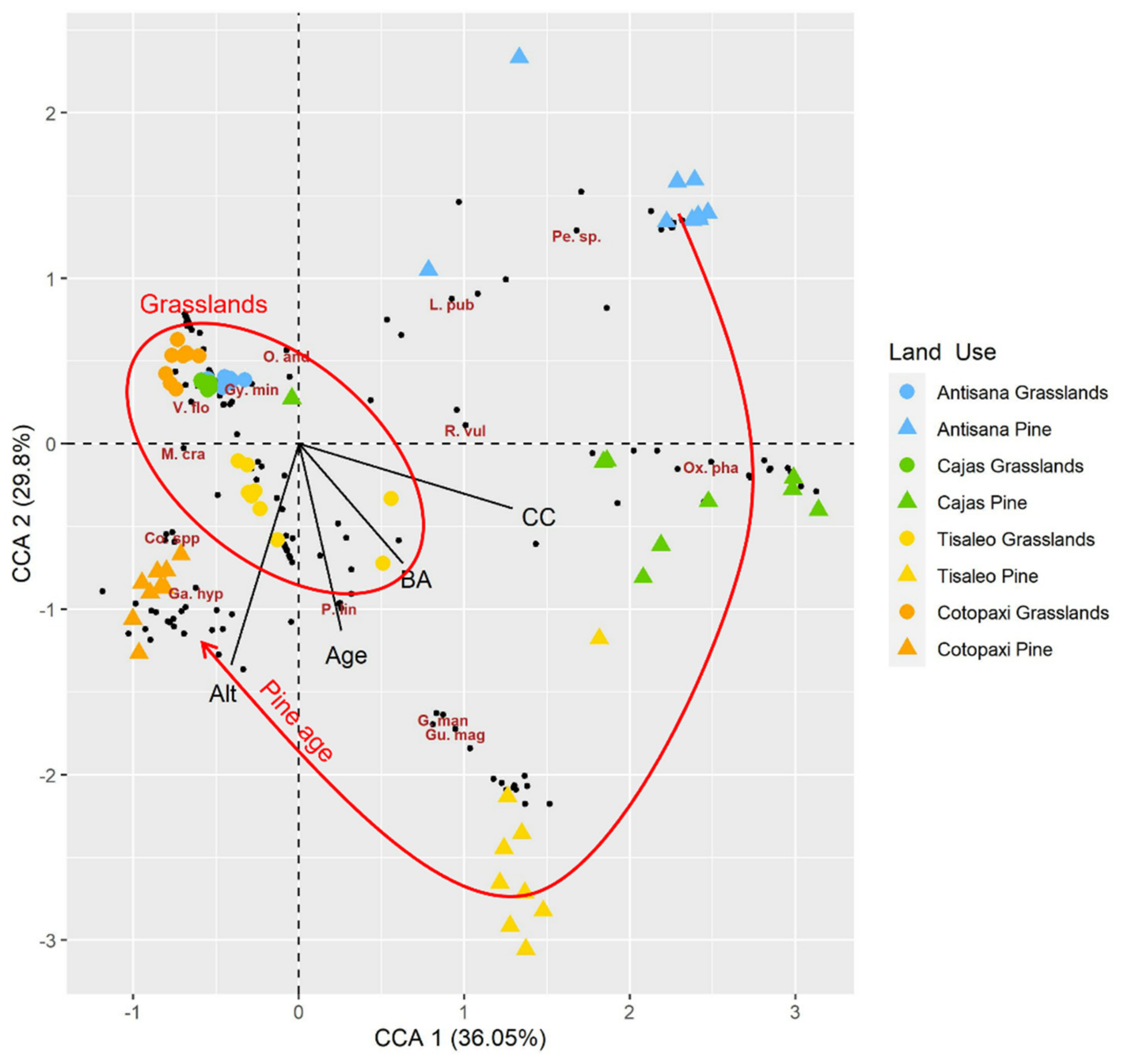
In the plane formed by the first two axes of the CCA explaining 66% (total inertia: 9.18, constrained inertia: 2.07) of the variability, grassland plots were grouped together close to the origin of the axes, while pine plantation plots were well discriminated by zone. Plots in Antisana and Cajas grasslands were placed together due to their similar species composition dominated by Calamagrostis intermedia (J. Presl) Steud. Tisaleo and Cotopaxi grassland plots were slightly apart mainly due to lower cover of C. intermedia (37% and 28%, respectively) and presence of shrubs in Tisaleo and Puya hamata L.B. Sm. and Cortaderia jubata (Lem.) Stapf. in Cotopaxi. Pine plantation plots were grouped by zone and the zones were placed relative to each other in such a way that a trajectory reflecting the age of the pine plantation appeared. The plots in the oldest plantation located in Cotopaxi were the closest to the grassland plots (Figure 2).
3.2. Land Use and Zone Effects on Plant Cover and Species Richness
The linear model including land use (grassland vs. pine plantation) and zone effects as well as their interaction explained 73% of the total plant cover variability. The pine plantation effect was significant (p < 0.001) and contributed the most to the variability (46%). Pine plantations reduced the total plant cover by 29.5% in Tisaleo and by 90.6% in Cajas, with an average reduction of 47.5% (Table 2). Moss cover varied mainly among study zones (26% of the variability) and its change under pine was on average not significant (Table 2). Only in Cotopaxi, moss cover was significantly reduced under pine (from 17.3 to 8.6%). The woody species cover varied strongly among study zones (48% of the variability), while the effect of pine plantation was much more limited and restricted to Tisaleo and Cotopaxi. In these zones, woody species cover decreased by 10.3% and 18.8%, respectively (Table 2). For the herbaceous species, cover change under pine was significant (p < 0.001) and explained most of the variability (39% for the land use effect + 21% for its interaction with the zone effect). A 40.1% decrease of herbaceous cover occurred under pine plantations compared to grasslands and this reduction was more pronounced in Cajas (89.7%) than in the other study zones (16%–25.6%) (Table 2).
Among the life forms of the herbaceous species, the tussock cover (33.6%–86.3%) was strongly affected by pine plantation, with a reduction amounting to 50.8% on average (p < 0.001, R2 = 0.57). The largest cover reduction was registered in Cajas (83%), while no change in tussock cover was detected in Cotopaxi (Table 2). Cushion cover ranged between 1.4% in Cotopaxi grasslands and 8.6% in Tisaleo grasslands and was reduced to 0% and 0.4%, respectively, under the pine plantations in the same zones (p < 0.001). The linear model explained 62% of cushion cover variability, with the largest contribution of the land-use effect (24%) followed by the zone effect (21%) and their interaction (17%). Rosette cover variability was well explained (R2 = 0.71) by the linear model. Rosette cover declined under pine in all zones except Tisaleo, where it was already quite low in the grasslands (0.8%). Compared to the other plant species, small herbs cover increased under pine, especially in Tisaleo (Table 2). The cover of light-demanding species was drastically reduced under pine in all zones (−59% on average, p < 0.001) but this effect was less pronounced in Cotopaxi (Table 2). Shade-tolerant species cover varied mainly among study zones (34% of the variability) and its increase under pine was significant only in Antisana (from 11.8 to 53.2%). Total species richness was reduced significantly in all zones except in Cotopaxi. The loss of species (on average, 8 species) occurred in herbaceous species, while richness of woody species was not affected by pine plantations except in Cotopaxi where it increased (Table 2).
In Ecuadorian Páramo vegetation (grasslands), the dominant species was Calamagrostis intermedia, even if this dominance was less pronounced in Tisaleo and Cotopaxi (Appendix A). In these areas, some shrub species were also quite abundant: Disterigma empetrifolium (Kunth) Nied. and Baccharis caespitosa (Ruiz and Pav.) Pers. in Tisaleo and Puya hamata, Cortaderia jubata, Loricaria ilinissae (Benth.) Cuatrec and Diplostephium ericoides (Lam.) Cabrera in Cotopaxi. Pine plantations reduced Calamagrostis intermedia cover by 21% to 84% in Cotopaxi and Antisana, respectively. In Antisana, it was mainly replaced by the small herbs plant species Lachemilla orbiculata (Ruiz and Pav.) Rydb and Agrostis breviculmis Hitchc. In Cajas, no species was clearly more abundant than any other under pines. In Tisaleo, the most abundant species under pines were Gunnera magellanica Lam., Sibthorpia repens (Mutis ex L.) Kuntze, Alchemilla hirta Rothm. and Aristeguietia sp. Finally, in Cotopaxi, Cortaderia jubata became the most abundant species under pines. Disterigma empetrifolium cover and Puya hamata cover were reduced almost completely under pines in the Tisaleo and Cotopaxi zone (Appendix A).
Some species were considered as indicators of pine plantations in a given study area due to their exclusive presence there (specificity). Taraxacum campylodes G.E. Haglund, Margyricarpus pinnatus (Lam.) Kuntze, Oxalis lotoides Kunth, Arcytophyllum nitidum (Kunth) Schltdl., Calceolaria crenata Lam. and Calamagrostis ligulata (Kunth) Hitchc. were only recorded under the pine plantation in Antisana. Miconia salicifolia (Bonpl. ex Naudin) Naudin, Asplenium undetermined (sp.2), Asplenium fragans Sw. , Polypodium thyssanolepis A. Braun ex Klotzsch, Arcytophyllum filiforme (Ruiz and Pav.) Standl. and Cystopteris fragilis (L.) Bernh. were found only in the pine plantation of Cajas. Likewise, in Tisaleo, the species Pteridium arachnoideum (Kaulf.) Maxon and Aristeguietia sp. were exclusively recorded in the pine plantation. Finally, Rubus nubigenus Kunth, Bomarea multiflora (L.f.) Mirb., Cyathea xenoxyla Lehnert, Ribes ecuadorense Jancz., Senecio tephrosioides Turcz., Solanum stenophyllum Dunal, Aristeguietia lamiifolia (Kunth) R.M.King and H.Rob. and Asplenium monanthes L. were exclusive species of the Cotopaxi pine plantation.
A total of 16 endemic species were registered in this study, 10 in pine plantations and 9 in grasslands, 3 species shared between both land uses. Among them, Ageratina dendroides (Spreng.) R.M.King and H.Rob., Brachyotum jamesonii Triana. and Gentianella hyssopifolia (Kunth) Fabris are considered vulnerable. However, Halenia serpyllifolia J.S. Pringle, Loricaria azuayensis Cuatrec. and Ribes lehmannii Jancz. are endangered, and Geranium antisanae R. Knuth is critically endangered according to the International Union for Conservation of Nature and natural resources (IUCN) [30]. The total cover of endemic species did not change significantly under pines.
3.3. Relationships between Pine Cover and Plant Cover/Richness
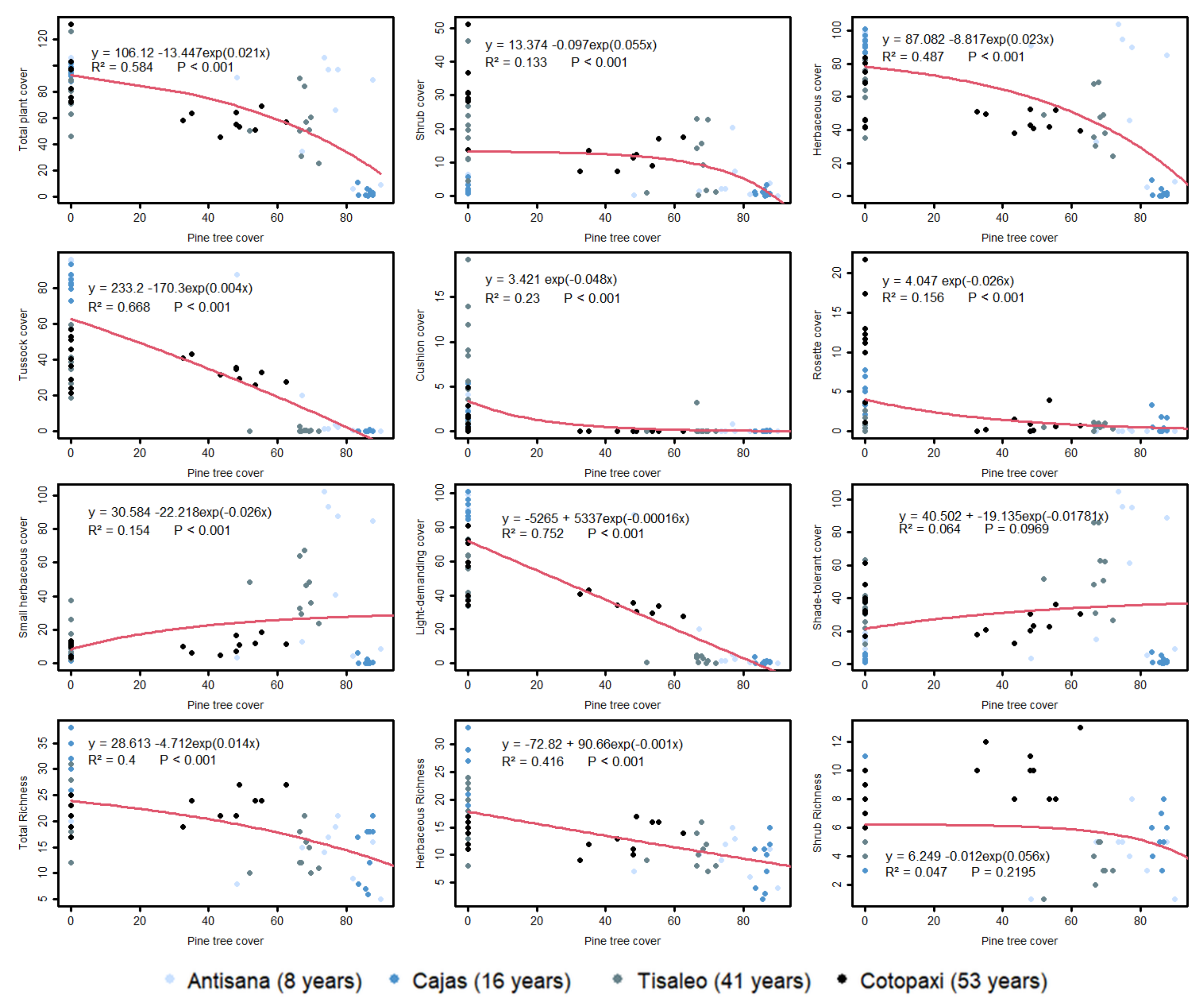
Total plant cover first decreased slowly with pine cover (until 50%) and then dropped more sharply. The same pattern was observed for shrub and herbaceous species but the relationship between moss and pine cover was not significant, as well as that for shade-tolerant species cover (Figure 3). Tussock cover (as well as that of light-demanding species cover) declined linearly with pine cover, while cushion and rosette species cover decreased exponentially with pine cover. The cover of small herbs increased with pine cover but progressively leveled off beyond 35% of pine cover. Total species richness first declined slowly and progressively more sharply. The decline in herbaceous species richness was linear and no significant relationship was found for shrub species richness (Figure 3).
3.4. Indicator Species of the Study Zone and of the Pine Effect
Zone effects—The indicator species determined by indicator value according IndVal in both land uses of Antisana were Geranium antisanae, Oreomyrrhis andicola (Kunth) Endl. ex Hook. f., Peperomia sp. and Lupinus pubescens Benth. In Cajas, Rhynchospora vulcani Boeckeler, Gynoxys miniphylla Cuatrec. and Oxalis phaeotricha Diels. were indicator species. Gunnera magellanica, Geranium maniculatum H.E. Moore and Plantago linearis Kunth. were indicator species of Tisaleo. In Cotopaxi, Cortaderia jubata, Cortaderia nitida Pilg., Vaccinium floribundum Kunth, Monnina crassifolia (Bonpl.) Kunth, Galium hypocarpium (L.) Endl. ex Griseb. and Carex polystachya Sw. ex Wahlenb. were identified as indicators and were present in both land uses (Figure 1, Appendix A, Supplementary Table S1).
Pine effects—The indicator species in the understory of the pine plantation depended on stand age. Indicator species under pines in Antisana (youngest plantation) were Lachemilla orbiculata, Margyricarpus pinnatus, Agrostis breviculmis and Taraxacum campylodes, all of which are shade-tolerant species. In Cajas (higher canopy), the indicator species under pines were two Melastomataceae shrub species, Brachyotum jamesonii and Miconia salicifolia, species common in subPáramo. Indicator species under pine plantations in Tisaleo were Sibthorpia repens (Mutis ex L.) Kuntze, Pteridium arachnoideum and Asplenium cuspidatum Lam., shade-tolerant herbaceous species. In the older Cotopaxi plantation, the indicator species comprised Rubus coriaceus Poir. and Rubus nubigenus, Ribes ecuadorense and Bomarea multiflora, naturally abundant in subPáramo and high montane forests, also shade-tolerant species (Appendix A, Supplementary Table S1).
4. Discussion
4.1. What Is the Impact of Pine Plantations on the Cover and Diversity of Understory Species?
Our results indicate that pine plantations affect the Páramo vegetation by strongly reducing the cover and diversity of herbaceous species, mainly light-demanding and tussock species such as Calamagrostis intermedia. Compared to tussocks, cushions and rosettes have a much more limited cover in the Páramo grasslands (0.5%–11.3%). These two herbaceous life forms are, however, also negatively impacted by the presence of pine trees. In contrast, the abundance of small herbs substantially increases under pine. Shrub abundance declines in the understory of pine plantations while their species richness remains similar compared to the grasslands. Regarding moss cover, no significant effect of pine was detected.
Results reported by several authors [29,43,45] coincide with our study regarding the reduction of tussock and light-demanding species, as well as the increase in the cover of small herbs species, which are normally scarce in the natural grassland. Lower cover values of Calamagrostis intermedia are associated with forested areas with intermediate or dense canopy that partly prevents light transmission to the understory. Furthermore, Gloser et al. [65] and Gloser and Gloser [44] found that Calamagrostis epigejos (L.) Roth shows lower growth in controlled shaded environments.
When the canopy closes, light conditions in the understory are reduced, which explains the decline in light-demanding species, as reported for New Zealand Pinus radiata plantations by Brockerhoff et al. [46], who found a reduction in adventive and indigenous (herbaceous and shrubs) plant cover under pine. Likewise, Quiroz-Dahik et al. [45] aimed at assessing differences in natural succession between pine plantations and grasslands in the Ecuadorian southern Páramo and found a lower cover of light-demanding species in the understory of pine plantations.
Trees provide protection from high solar radiation, strong winds and night frosts, which can facilitate the colonization of species from subPáramo [27,66]. This was obvious in our results, where cover of common upper montane forest and subPáramo species (mainly shrubs) as Aristeguietia sp., Brachyotum ledifolium (Desr.) Triana, Rubus coriaceus and Rubus nubigenus were higher under pine than in grasslands. These species have wide altitudinal distribution [67], and were registered in other studies as being common in upper montane forest or subPáramo but rarer in natural Páramo grasslands [56,68,69]. In the same way, Van Wesenbeeck et al. [42] also found small differences between plots with low canopy of pine trees and subPáramo plots. Thus, tree protection against climate constraints can explain the presence of these species in our study plantations [30,48,70].
Pine plantations showed no clear effects on endemic or threatened species, due to the scarcity of such species. Given the random distribution and scarcity of these species, it is impossible to say whether these differences between land uses were obtained by chance or resulted from real effects of pines. For this reason, we cannot provide a clear response regarding the influence of pine plantations on endemic and endangered species.
4.2. Does the Zone/Age Influence the Pine Plantation Impact on Páramo Vegetation?
The pine plantation effects on Páramo vegetation differed from one study zone to another, which was reflected by the interaction term of the linear model (land use x study zone, Table 2). The main factor explaining the differences between zones was canopy cover (Figure 3), depending on stand age, but also on climate conditions changing with altitude. At the plantation time, the canopy is quite open but closes as pines grow. In these pine plantations, basal area was lower than expected, and tree mortality was due to not only self-thinning but also the climate conditions, which are not suitable for tree growth above 3600 m. The canopy cover of pine plantations tended to decrease with age but was also influenced by altitude. Growing conditions were better in the Antisana plantation located at 3556 m than in the other zones, which ranged from 3699 to 3874 m.
The pine effect was therefore more pronounced in the young and dense plantations of Antisana and Cajas than in the old and open pine stand in Cotopaxi, whose understory vegetation was more similar to that of grasslands (Table 2, Figure 2). In the dense young plantations, the cover of the light-demanding and tussock species largely decreased, favoring an increase in shade-tolerant species, except in Cajas. Under the canopy of the oldest pine plantation in Cotopaxi, shade-tolerant species were interspersed with light-demanding species, and this mixture resulted in a high number of native species, as reported in other studies [27,45,71,72,73]. Open canopy allowed cohabitation of light-demanding species and shade-tolerant species, this increased the species richness and total plant cover.
A decrease in the cover of the most dominant species, Calamagrostis intermedia, was registered in our four zones under pines (Appendix A). In Cajas, where the pine cover is the highest, Calamagrostis intermedia disappeared without being replaced by other species, while it was replaced by more shade-tolerant species of other life forms, Lachemilla orbiculata and Agrostis breviculmis in Antisana and Gunnera magellanica and Sibthorpia repens in Tisaleo, and by other tussock species less light-demanding, Cortaderia spp. and Carex polystachya in Cotopaxi. Similarly, Matson and Bart [74] also observed an increased abundance of Cortaderia nitida in response to expanding shrub cover in a protected area of the Zuleta Páramo. Such substitutions were also noticed by Grubbs et al. [75], who described changes in the structure and life forms of the vegetation in a higher zone of the Páramo in Antisana as a result of human activities.
The contrasting vegetation composition between our study zones reflected differences due to land-use history and induces variations in the pine effects on Páramo vegetation. Grazing intensity and grassland degradation probably were higher in Antisana than in other zones, because sheep grazing occurred there and has severe impacts on vegetation since sheep consumed both the above- and below-ground parts of the herbaceous species growing between the less palatable Calamagrostis tussocks [76]. Due to grazing, in Antisana and Tisaleo grasslands, Calamagrostis intermedia was partly replaced by stoloniferous ground-covering herbs, such as Lachemilla orbiculata or Gunnera magellanica and by short grasses, such as Agrostis breviculmis (only in Antisana), which are typical species of grazed Páramo [40,76,77]. These changes in Páramo vegetation due to land-use history were still exacerbated under pines.
5. Conclusions
Our results showed that pine afforestation effects on Páramo vegetation depend mainly on the canopy cover and secondarily on the vegetation composition at the time of planting. After plantation, pine growth induces a closing of the canopy in 15 years which strongly reduces light reaching the understory and affects light-demanding herbaceous species. Then, canopy cover is progressively reduced due to tree mortality and to the fact that the crown-to-stem diameter decreases with age. This high mortality rate is probably due to the harsh climate conditions at altitudes above 3600 m. In the oldest and more open pine plantation (more light availability), the abundance and composition of the Páramo vegetation is more similar to the grassland vegetation than in the pine plantations of the other study zones. We recommend heavy thinning operations in existing pine plantations to maintain good light conditions in the understory and favor its development. Where native species have disappeared, an active restoration technique can be used to re-introduce these species after canopy opening by tree removal. As the light conditions improve with stand age, long rotation and/or continuous forestry should be preferred, which will also limit soil and vegetation disturbances. As pioneer tree species, pines create microclimate conditions that could be favorable to the growth of native woody species present in the native mountain forests below 3600 m.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/f13091499/s1. Figure S1: Relationship between pine canopy cover and basal area. Table S1. Phytosociological table.
Author Contributions
A.-L.J. and M.J. initiated the study, obtained funding and supervised the project. B.M.-T. and M.R. performed the field and herbarium work and the statistical analyses. All authors discussed the results and contributed to writing the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by l’Académie de recherche et d’enseignement supérieur (ARES from Belgium)—Universidad Central del Ecuador (UCE) cooperation through a doctoral fellowship awarded to Byron Medina-Torres (Funding: P340900031).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are openly available in Mendeley Data at doi: 10.17632/dm7pf3zr6w.2.
Acknowledgments
The authors thank the Ministry of Environment of Ecuador, the staff of the Direction of Environment; Marco Vallejo and Gilda Cepeda, owners of “La Guaytara”, Alonso Córdova, owner of “Casa Vieja”, for the permission to carry out field measurements in the Cotopaxi, Antisana and Cajas zones, and to QCA and QCNE Herbaria for their advice concerning the sampling and taxonomic identification of plant species. For their collaboration during fieldwork, we also thank Edwin Llumitasig, Francisco Núñez, Alexandra García, Daysi Gualavisi and Carolina Cachiguango in Cotopaxi, as well as Gabriela Quinto, Estefania Aguagüiña, Bécquer Súarez and Estefania Pillajo in Antisana, and Mikaela Morales in Cajas and Tisaleo. Many thanks to Arthur Guignabert and Léna Jeannerod for their help in statistical analysis and to Jennifer Mach (Plant Editors) for language improvement and paper editing. Byron Medina-Torres is particularly grateful to the ARES-UCE Belgium-Ecuador cooperation for awarding him a doctoral fellowship. He is also very grateful to Veerle Vanacker and Pierre Delmelle of his doctoral support committee and to Gabriela Bidegain for all her help with the administrative procedures.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study, in the collection, analyses, or interpretation of data, in the writing of the manuscript, or in the decision to publish the results.
Appendix A

{kind=link}
{kind=link}
{kind=link}
Table A1.
Plant cover (%) of species in Páramo grasslands and under pine plantations in Antisana, Cajas, Tisaleo and Cotopaxi.
Table A1.
Plant cover (%) of species in Páramo grasslands and under pine plantations in Antisana, Cajas, Tisaleo and Cotopaxi.
| Antisana | Cajas | Tisaleo | Cotopaxi | Shade | Life | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Family | Species | Grasslands | Pine | Grasslands | Pine | Grasslands | Pine | Grasslands | Pine | Tolerance | Form |
| Poaceae | Calamagrostis intermedia | 86.3 | 1.9 | 83.7 | 0.1 | 37.3 | 0.2 | 28.4 | 6.7 | LD | T |
| Rosaceae | Lachemilla orbiculata | 1.9 | 42.9 | <0.1 | 0.2 | 3.4 | 5 | 0.9 | 0.5 | ST | H |
| Poaceae | Cortaderia jubata | 7.8 | 18.3 | LD | T | ||||||
| Gunneraceae | Gunnera magellanica | 4.9 | 14.4 | ST | H | ||||||
| Plantaginaceae | Sibthorpia repens | <0.1 | <0.1 | 12 | ST | H | |||||
| Poaceae | Agrostis breviculmis | <0.1 | 11 | LD | T | ||||||
| Bromeliaceae | Puya hamata | <0.1 | 10.7 | 0.2 | LD | R | |||||
| Ericaceae | Disterigma empetrifolium | 0.3 | 0.6 | 0.1 | 8.9 | 0.1 | ST | S | |||
| Rosaceae | Alchemilla hirta | 0.2 | 0.1 | 2.9 | 8.8 | ST | H | ||||
| Asteraceae | Baccharis caespitosa | <0.1 | 7.8 | LD | C | ||||||
| Asteraceae | Loricaria ilinissae | 0.4 | 7.5 | ST | S | ||||||
| Asteraceae | Diplostephium ericoidese | 6.3 | ST | S | |||||||
| Ericaceae | Gaultheria myrsinoides | 0.7 | <0.1 | <0.1 | 4.3 | 1.4 | 5.4 | 0.3 | ST | S | |
| Poaceae | Cortaderia nitida | 2.2 | 5.3 | LD | T | ||||||
| Hypericaceae | Hypericum laricifolium | 0.1 | 0.1 | 2.5 | 4.3 | <0.1 | ST | S | |||
| Asteraceae | Aristeguietia sp. | 4.1 | ST | S | |||||||
| Caprifoliaceae | Valeriana microphylla | 2.7 | 0.5 | 3.7 | 2.4 | 0.1 | 0.1 | ST | S | ||
| Ericaceae | Vaccinium floribundum | <0.1 | 0.1 | 3.6 | 1.3 | ST | S | ||||
| Poaceae | Paspalum bonplandianum | 3.4 | 0.1 | LD | T | ||||||
| Cyperaceae | Carex polystachya | 1.5 | 3.3 | LD | T | ||||||
| Geraniaceae | Geranium reptans | 2.9 | 3 | 0.1 | <0.1 | 0.4 | 0.6 | ST | H | ||
| Asteraceae | Werneria nubigena | 0.2 | 2.8 | 0.1 | 0.2 | LD | R | ||||
| Poaceae | Cortaderia sericantha | 2.8 | LD | T | |||||||
| Lycopodiaceae | Lycopodium magellanicum | 2.8 | ST | H | |||||||
| Asteraceae | Senecio tephrosioides | 2.5 | ST | H | |||||||
| Rubiaceae | Galium hypocarpium | <0.1 | <0.1 | 0.3 | 2.4 | ST | H | ||||
| Asteraceae | Baccharis teindalensis | 2.4 | <0.1 | ST | S | ||||||
| Melastomataceae | Brachyotum ledifolium | 0.1 | 2.2 | ST | S | ||||||
| Rubiaceae | Arcytophyllum nitidum | 2.1 | ST | S | |||||||
| Geraniaceae | Geranium maniculatum | <0.1 | 0.9 | 2.1 | ST | H | |||||
| Rosaceae | Rubus coriaceus | 0.2 | 2 | ST | S | ||||||
| Rosaceae | Rubus nubigenus | 1.8 | ST | S | |||||||
| Rubiaceae | Nertera granadensis | 0.4 | 0.1 | 0.2 | 1.8 | ST | H | ||||
| Rosaceae | Margyricarpus pinnatus | 1.8 | ST | S | |||||||
| Geraniaceae | Geranium antisanaee | 1.4 | 1.7 | ST | H | ||||||
| Apiaceae | Azorella biloba | 1.5 | LD | C | |||||||
| Apiaceae | Azorella pedunculata | 1.4 | 0.1 | 0.2 | 0.6 | 1.4 | LD | C | |||
| Cyatheaceae | Cyathea xenoxyla | 1.4 | ST | S | |||||||
| Rosaceae | Polylepis sp. | <0.1 | 1.3 | ST | S | ||||||
| Lycopodiaceae | Lycopodium clavatum | 0.1 | 1.2 | ST | H | ||||||
| Gentianaceae | Gentianella cerastioides | 0.1 | <0.1 | 1.2 | LD | H | |||||
| Solanaceae | Solanum stenophyllum | 1 | ST | S | |||||||
| Cyperaceae | Carex sp.1 | 1 | 0.3 | LD | T | ||||||
| Lycopodiaceae | Phlegmariurus crassus | 0.3 | <0.1 | 1 | ST | H | |||||
| Alstroemeriaceae | Bomarea multiflora | 0.9 | ST | H | |||||||
| Asteraceae | Aristeguietia lamiifolia | 0.8 | ST | S | |||||||
| Caprifoliaceae | Aretiastrum aretioidese | 0.8 | LD | C | |||||||
| Poaceae | Bromus lanatus | 0.7 | <0.1 | 0.4 | LD | H | |||||
| Dryopteridaceae | Elaphoglossum engelii | 0.7 | 0.2 | ST | H | ||||||
| Araliaceae | Hydrocotyle bonplandii | 0.6 | 0.2 | <0.1 | 0.7 | 0.1 | ST | H | |||
| Rosaceae | Acaena ovalifolia | 0.2 | 0.7 | ST | H | ||||||
| Rosaceae | Lachemilla hispidula | <0.1 | 0.6 | ST | H | ||||||
| Grossulariaceae | Ribes lehmanniie | <0.1 | 0.6 | ST | S | ||||||
| Cyperaceae | Rhynchospora vulcani | 0.6 | 0.5 | <0.1 | LD | R | |||||
| Cyperaceae | Carex sp. 2 | 0.5 | LD | T | |||||||
| Asteraceae | Senecio sp. | 0.5 | ST | H | |||||||
| Asteraceae | Galinsoga parviflora | <0.1 | 0.5 | ST | H | ||||||
| Apiaceae | Eryngium humile | <0.1 | 0.5 | 0.1 | LD | R | |||||
| Caryophyllaceae | Arenaria serpyllifolia | 0.1 | 0.1 | <0.1 | 0.5 | ST | H | ||||
| Lamiaceae | Clinopodium nubigenum | 0.5 | 0.3 | 0.2 | LD | H | |||||
| Asteraceae | Hieracium frigidum | <0.1 | <0.1 | 0.5 | <0.1 | ST | R | ||||
| Piperaceae | Peperomia sp. | 0.1 | 0.5 | ST | H | ||||||
| Lycopodiaceae | Austrolycopodium magellanicum | 0.4 | <0.1 | 0.4 | LD | H | |||||
| Berberidaceae | Berberis rigida | 0.1 | 0.1 | 0.4 | 0.2 | ST | S | ||||
| Geraniaceae | Geranium sibbaldioides | 0.4 | <0.1 | ST | H | ||||||
| Cyperaceae | Uncinia hamata | 0.4 | 0.3 | LD | R | ||||||
| Plantaginaceae | Plantago rigida | 0.4 | LD | C | |||||||
| Melastomataceae | Brachyotum jamesoniie | 0.1 | 0.4 | ST | S | ||||||
| Grossulariaceae | Ribes ecuadorense | 0.3 | ST | S | |||||||
| Cyperaceae | Uncinia tenuis | 0.3 | <0.1 | 0.3 | <0.1 | LD | R | ||||
| Gentianaceae | Halenia weddelliana | 0.3 | <0.1 | 0.1 | LD | H | |||||
| Asteraceae | Taraxacum campylodes | 0.3 | ST | H | |||||||
| Caryophyllaceae | Cerastium mollissimum | 0.3 | ST | H | |||||||
| Dennstaedtiaceae | Pteridium arachnoideum | 0.3 | ST | R | |||||||
| Asteraceae | Gynoxys buxifolia | 0.3 | LD | S | |||||||
| Polygalaceae | Monnina crassifolia | 0.3 | 0.2 | ST | S | ||||||
| Asteraceae | Gamochaeta cf. humilis | 0.3 | LD | H | |||||||
| Asteraceae | Bidens andicola | 0.2 | LD | H | |||||||
| Hypericaceae | Hypericum lancioides | 0.2 | ST | S | |||||||
| Cyperaceae | Rhynchospora sp. | 0.2 | LD | R | |||||||
| Unidentified | Unknown 3 | <0.1 | 0.2 | ST | R | ||||||
| Apiaceae | Azorella aretioides | <0.1 | 0.2 | LD | C | ||||||
| Asteraceae | Bidens pilosa | 0.2 | LD | H | |||||||
| Lamiaceae | Salvia pichinchensis | 0.2 | LD | S | |||||||
| Asteraceae | Gynoxys miniphyllae | 0.2 | <0.1 | LD | S | ||||||
| Aspleniaceae | Asplenium monanthes | 0.2 | ST | R | |||||||
| Melastomataceae | Miconia chionophila | 0.2 | ST | S | |||||||
| Blechnaceae | Blechnum loxense | 0.1 | 0.2 | ST | R | ||||||
| Polypodiaceae | Polypodium mindensee | 0.2 | ST | H | |||||||
| Asteraceae | Diplostephium sp. | 0.2 | 0.1 | ST | S | ||||||
| Poaceae | Calamagrostis ligulata | 0.2 | LD | T | |||||||
| Calceolariaceae | Calceolaria crenata | 0.2 | ST | S | |||||||
| Plantaginaceae | Plantago linearis | 0.2 | <0.1 | LD | T | ||||||
| Asteraceae | Ageratina dendroidese | 0.2 | ST | S | |||||||
| Hypericaceae | Hypericum decandrum | 0.2 | <0.1 | ST | S | ||||||
| Iridaceae | Orthrosanthus chimboracensis | 0.1 | 0.1 | LD | H | ||||||
| Ranunculaceae | Ranunculus praemorsus | 0.1 | <0.1 | LD | H | ||||||
| Asteraceae | Pentacalia peruviana | 0.1 | LD | S | |||||||
| Aspleniaceae | Unknown 2 | <0.1 | 0.1 | ST | H | ||||||
| Apiaceae | Oreomyrrhis andicola | 0.1 | <0.1 | <0.1 | ST | H | |||||
| Asteraceae | Dorobaea pimpinellifolia | <0.1 | 0.1 | <0.1 | LD | H | |||||
| Geraniaceae | Geranium multipartitum | 0.1 | ST | H | |||||||
| Poaceae | Agrostis sp. 1 | 0.1 | LD | H | |||||||
| Rubiaceae | Arcytophyllum filiforme | 0.1 | ST | S | |||||||
| Aspleniaceae | Asplenium cuspidatum | 0.1 | ST | R | |||||||
| Asteraceae | Gynoxys sp. | 0.1 | <0.1 | LD | S | ||||||
| Blechnaceae | Blechnum auratum | 0.1 | ST | R | |||||||
| Cyatheaceae | Cyathea caracasana | 0.1 | ST | S | |||||||
| Asteraceae | Gynoxys halliie | 0.1 | LD | S | |||||||
| Escalloniaceae | Escallonia myrtilloides | 0.1 | <0.1 | LD | S | ||||||
| Pteridaceae | Jamesonia alstonii | 0.1 | ST | H | |||||||
| Hypericaceae | Hypericum aciculare | 0.1 | ST | S | |||||||
| Asteraceae | Hypochaeris sessiliflora | 0.1 | 0.1 | ST | R | ||||||
| Rosaceae | Lachemilla nivalis | 0.1 | LD | H | |||||||
| Bromeliaceae | Puya clava-herculis | <0.1 | 0.1 | LD | R | ||||||
| Asteraceae | Monticalia myrsinitese | 0.1 | <0.1 | LD | S | ||||||
| Asteraceae | Loricaria azuayensise | 0.1 | ST | S | |||||||
| Asteraceae | Erigeron ecuadoriensis | 0.1 | <0.1 | LD | H | ||||||
| Dryopteridaceae | Polystichum orbiculatum | 0.1 | ST | H | |||||||
| Melastomataceae | Brachyotum sp. | 0.1 | <0.1 | ST | S | ||||||
| Melastomataceae | Miconia salicifolia | 0.1 | ST | S | |||||||
| Asteraceae | Gamochaeta americana | <0.1 | <0.1 | 0.1 | LD | H | |||||
| Ophioglossaceae | Ophioglossum crotalophoroides | 0.1 | <0.1 | LD | H | ||||||
| Poaceae | Festuca andicola | 0.1 | LD | T | |||||||
| Rubiaceae | Galium corymbosum | 0.1 | ST | H | |||||||
| Asteraceae | Aphanactis ollgaardiie | <0.1 | LD | H | |||||||
| Gentianaceae | Halenia serpyllifoliae | <0.1 | LD | H | |||||||
| Oxalidaceae | Oxalis phaeotricha | <0.1 | <0.1 | LD | H | ||||||
| Dryopteridaceae | Polystichum sp. | <0.1 | ST | H | |||||||
| Fabaceae | Trifolium pratense | <0.1 | <0.1 | ST | H | ||||||
| Gentianaceae | Gentianella hyssopifoliae | <0.1 | ST | H | |||||||
| Polypodiaceae | Polypodium thyssanolepis | <0.1 | ST | H | |||||||
| Asteraceae | Cronquistianthus pseudoriganoidese | <0.1 | ST | S | |||||||
| Unidentified | Unknown 5 | <0.1 | ST | R | |||||||
| Plantaginaceae | Plantago australis | <0.1 | <0.1 | LD | H | ||||||
| Asteraceae | Monticalia arbutifolia | <0.1 | LD | S | |||||||
| Asteraceae | Baccharis genistelloides | <0.1 | ST | H | |||||||
| Cystopteridaceae | Cystopteris fragilis | <0.1 | <0.1 | ST | H | ||||||
| Dryopteridaceae | Elaphoglossum mathewsii | <0.1 | ST | H | |||||||
| Oxalidaceae | Oxalis lotoides | <0.1 | ST | H | |||||||
| Unidentified | Unknown 4 | <0.1 | ST | H | |||||||
| Aspleniaceae | Asplenium fragans | <0.1 | <0.1 | ST | R | ||||||
| Asteraceae | Baccharis tricuneata | <0.1 | <0.1 | LD | S | ||||||
| Apiaceae | Oreomyrrhis sp. | <0.1 | ST | H | |||||||
| Dryopteridaceae | Polystichum polyphyllum | <0.1 | ST | H | |||||||
| Fabaceae | Lupinus microphyllus | <0.1 | ST | H | |||||||
| Polygonaceae | Muehlenbeckia tamnifolia | <0.1 | ST | S | |||||||
| Asteraceae | Cotula mexicana | <0.1 | LD | H | |||||||
| Asteraceae | Gnaphalium dombeyanum | <0.1 | LD | H | |||||||
| Gentianaceae | Gentiana sedifolia | <0.1 | <0.1 | LD | H | ||||||
| Ranunculaceae | Ranunculus limoselloides | <0.1 | LD | H | |||||||
| Cyperaceae | Rhynchospora ruiziana | <0.1 | LD | R | |||||||
| Aspleniaceae | Asplenium sp. 2 | <0.1 | ST | H | |||||||
| Asteraceae | Achyrocline alata | <0.1 | ST | H | |||||||
| Asteraceae | Pluchea biformis | <0.1 | ST | H | |||||||
| Unidentified | Unknown 6 | <0.1 | ST | H | |||||||
| Ericaceae | Disterigma sp. 1 | <0.1 | ST | S | |||||||
| Fabaceae | Lupinus pubescens | <0.1 | <0.1 | ST | S | ||||||
| Aspleniaceae | Asplenium polyphyllum | <0.1 | ST | R | |||||||
| Asteraceae | Unknown 7 | <0.1 | ST | R | |||||||
| Apiaceae | Daucus montanus | LD | H | ||||||||
| Apiaceae | Niphogeton dissecta | LD | H | ||||||||
| Asteraceae | Taraxacum sp. 1 | ST | H | ||||||||
| Unidentified | Unknown 1 | ST | H | ||||||||
| Asteraceae | Aristeguietia cf. glutinosa | ST | S | ||||||||
| Asteraceae | Hypochaeris sonchoidese | ST | R | ||||||||
| Asteraceae | Ageratina sp. | ST | S | ||||||||
| Total cover (%) | 101.5 | 66.3 | 99 | 3.4 | 85 | 56.5 | 92.2 | 57.4 | |||
| Total species richness | 29 | 25 | 52 | 42 | 45 | 35 | 36 | 48 | |||
LD: light-demanding, ST: shade-tolerant, S: shrub, T: tussock, C: cushion, R: rosette, H: small herbs, e = endemic species.
References
- FAO. Global Forest Resources Assessment 2015. Desk Reference; 1st ed.; Food and Agriculture Organization of the United Nations: Rome, Italy, 2015; pp. 34–69. [Google Scholar]
- FAO. Global Forest Resources Assessment 2010. Desk Reference; Food and Agriculture Organization of the United Nations: Rome, Italy, 2010; pp. 9–48. [Google Scholar]
- FAO. Global Planted Forests Thematic Study: Results and Analysis; Planted Forests and Trees Working Paper 38; Food and Agriculture Organization of the United Nations: Rome, Italy, 2006; pp. 13–28. [Google Scholar]
- Farley, K.A. Grasslands to Tree Plantations: Forest Transition in the Andes of Ecuador. Ann. Assoc. Am. Geogr. 2007, 97, 755–771. [Google Scholar] [CrossRef]
- Vanacker, V.; von Blanckenburg, F.; Govers, G.; Molina, A.; Poesen, J.; Deckers, J.; Kubik, P. Restoring Dense Vegetation Can Slow Mountain Erosion to near Natural Benchmark Levels. Geology 2007, 35, 303–306. [Google Scholar] [CrossRef]
- Geldenhuys, C. Native Forest Regeneration in Pine and Eucalypt Plantations in Northern Province, South Africa. For. Ecol. Manag. 1997, 99, 101–115. [Google Scholar] [CrossRef]
- Parrotta, J.A.; Turnbull, J.W.; Jones, N. Catalyzing Native Forest Regeneration on Degraded Tropical Lands. For. Ecol. Manag. 1997, 99, 1–7. [Google Scholar] [CrossRef]
- Parrotta, J.A. Influence of Overstory Composition on Understory Colonization by Native Species in Plantations on a Degraded Tropical Site. J. Veg. Sci. 1995, 6, 627–636. [Google Scholar] [CrossRef]
- Fimbel, R.A.; Fimbel, C.C. The Role of Exotic Conifer Plantations in Rehabilitating Degraded Tropical Forest Lands: A Case Study from the Kibale Forest in Uganda. For. Ecol. Manag. 1996, 81, 215–226. [Google Scholar] [CrossRef]
- Richardson, D.M.; Van Wilgen, B.W.; Nuñez, M.A. Alien Conifer Invasions in South America: Short Fuse Burning? Biol. Invasions 2008, 10, 573–577. [Google Scholar] [CrossRef]
- Simberloff, D.; Nuñez, M.A.; Ledgard, N.J.; Pauchard, A.; Richardson, D.M.; Sarasola, M.; Van Wilgen, B.W.; Zalba, S.M.; Zenni, R.D.; Bustamante, R.; et al. Spread and Impact of Introduced Conifers in South America: Lessons from Other Southern Hemisphere Regions. Austral Ecol. 2010, 35, 489–504. [Google Scholar] [CrossRef]
- Freeman, J.; Marques, C.M.P.; Carocha, V.; Borralho, N.; Potts, B.M.; Vaillancourt, R. Origins and Diversity of the Portuguese Landrace of Eucalyptus globulus. Ann. For. Sci. 2007, 64, 639–647. [Google Scholar] [CrossRef]
- Estades, C.F.; Escobar, M.A. los Ecosistemas de las Plantaciones de Pino de la Cordillera de la Costa. In Historia, Biodiversidad y Ecología de los Bosques Costeros de Chile; Smith-Ramírez, C., Armesto, J., Valdovinos, C., Eds.; Editorial Universitaria: Santiago de Chile, Chile, 2015; pp. 600–616. [Google Scholar]
- Carle, J.B.; Holmgren, L.P.B. Wood from Planted Forests: Global Outlook to 2030. Plant. For. Uses Impacts Sustain. 2009, 58, 47–59. [Google Scholar] [CrossRef]
- Payn, T.; Carnus, J.M.; Freer-Smith, P.; Kimberley, M.; Kollert, W.; Liu, S.; Orazio, C.; Rodriguez, L.; Silva, L.N.; Wingfield, M.J. Changes in Planted Forests and Future Global Implications. For. Ecol. Manag. 2015, 352, 57–67. [Google Scholar] [CrossRef]
- Indústria Brasileira de Árvores. Report 2019; Brasilan Tree Industry: Brasilia, Brasil, 2019; pp. 32–41. [Google Scholar]
- Instituto Forestal. Anuario Forestal 2019; Área de información y economía del Instituto Forestal: Santiago de Chile, Chile, 2019; pp. 19–72. [Google Scholar]
- Ministerio de Agricultura Ganadería y Pesca de Argentina. Mapa de Plantaciones Forestales Promocionadas (MPF) de la República Argentina; Ministerio de Agricultura Ganadería y Pesca de Argentina: Buenos Aires, Argentina, 2018. [Google Scholar]
- Jonard, M.; Colmant, R.; Heylen, C.; Ysebaert, C.; Carton, C.; Picard, L.; Cassart, B.; Hounzandji, A.P.I.; Ponette, Q. Impact de Boisements Résineux Sur la Séquestration du Carbone dans les Andes Péruviennes: Cas des Plantations de Pinus patula dans la Coopérative de Granja Porcon (Cajamarca), Pérou. Bois. For. Trop. 2014, 68, 17–27. [Google Scholar] [CrossRef]
- Chávez, J.; Sabogal, C. Restoring Degraded Forest Land with Native Tree Species: The Experience of “Bosques Amazónicos” in Ucayali, Peru. Forests 2019, 10, 851. [Google Scholar] [CrossRef]
- Aguirre, N. Silvicultural Contributions to the Reforestation with Native Species in the Tropical Mountain Rainforest Region of South Ecuador; Technischen Universität München: Munchen, Germany, 2007. [Google Scholar]
- Avendaño-Yáñez, M.; Sánchez-Velásquez, L.; Meave, J.A.; Pineda-López, M. Can Pinus Plantations Facilitate Reintroduction of Endangered Cloud Forest Species? Landsc. Ecol. Eng. 2015, 12, 99–104. [Google Scholar] [CrossRef]
- Bremer, L.L.; Farley, K.A.; Chadwick, O.A.; Harden, C.P. Changes in Carbon Storage with Land Management Promoted by Payment for Ecosystem Services. Environ. Conserv. 2016, 43, 397–406. [Google Scholar] [CrossRef]
- Cargua, F.E.; Rodríguez, M.V.; Recalde, C.G.; Vinueza, L.M. Cuantificación del Contenido de Carbono En una Plantación de Pino Insigne (Pinus radiata) y En Estrato de Páramo de Ozogoche Bajo, Parque Nacional Sangay, Ecuador. Inf. Tecnol. 2014, 25, 83–92. [Google Scholar] [CrossRef]
- Farley, K.A.; Bremer, L.L.; Harden, C.P.; Hartsig, J. Changes in Carbon Storage under Alternative Land Uses in Biodiverse Andean Grasslands: Implications for Payment for Ecosystem Services. Consservation Lett. 2013, 6, 21–27. [Google Scholar] [CrossRef]
- Feyera, S.; Beck, E.; Lüttge, U. Exotic Trees as Nurse-Trees for the Regeneration of Natural Tropical Forests. Trees Struct. Funct. 2002, 16, 245–249. [Google Scholar] [CrossRef]
- Hofstede, R.G.; Groenendijk, J.P.; Coppus, R.; Fehse, J.C.; Sevink, J. Impact of Pine Plantations on Soils and Vegetation in the Ecuadorian High Andes. Mt. Res. Dev. 2002, 22, 159–167. [Google Scholar] [CrossRef]
- Arévalo, J.R.; Fernández-Palacios, J.M. From Pine Plantations to Natural Stands. Ecological Restoration of a Pinus Canariensis Sweet, Ex Spreng Forest. Plant Ecol. 2005, 181, 217–226. [Google Scholar] [CrossRef]
- Luteyn, J.L.; Churchill, S.P. Paramos: A Checklist of Plant Diversity, Geographical Distribution and Botanical Literature; Luteyn, J.L., Ed.; New York Botanical Garden: New York, NY, USA, 1999; ISBN 0893274275. [Google Scholar]
- Ramsay, P.M. The Páramo Vegetation of Ecuador: The Community Ecology, Dynamics and Productivity of Tropical Grasslands in the Andes; University of Wales, Bangor: Bangor, Wales, 1992. [Google Scholar]
- Hofstede, R.G.; Calles, J.; López, V.; Polanco, R.; Torres, F.; Ulloa, J.; Vásquez, A.; Cerra, M. Los Páramos Andinos ¿Qué Sabemos? Estado de Conocimiento Sobre El Impacto del Cambio Climático En El Ecosistema Páramo, 1st ed.; International Union for Conservation of Nature: Quito, Ecuador, 2014. [Google Scholar]
- Vásconez, P.M.; Hofstede, R.G. Los Páramos Ecuatorianos. In Botánica Económica de los Andes Centrales; Moraes, R.M., Øllgaard, B., Kvist, L.P., Borchsenius, F., Balslev, H., Eds.; Universidad Mayor de San Andrés: La Paz, Bolivia, 2006; pp. 91–109. ISBN 9995401215. [Google Scholar]
- Albán, M. Mecanismos Relacionados Con Servicios Ambientales Como una Herramienta para Conservación de los Páramos. In Páramo. Paisaje Estudiado, Habitado, Manejado e Institucionalizado; Mena Vásconez, P., Campaña, J., Castillo, A., Flores, S., Hofstede, R.G., Josse, C., Lasso, S., Medina, G., Ochoa, N., Ortiz, D., Eds.; Abya-Yala: Quito, Ecuador, 2011; pp. 231–246. [Google Scholar]
- Quiroz-Dahik, C.; Crespo, P.; Stimm, B.; Murtinho, F.; Weber, M.; Hildebrandt, P. Contrasting Stakeholders’ Perceptions of Pine Plantations in the Páramo Ecosystem of Ecuador. Sustainability 2018, 10, 1707. [Google Scholar] [CrossRef]
- León-Yánez, S.; Valencia, R.; Pitman, N.; Endara, L.; Ulloa-Ulloa, C.; Navarrete, H. Libro Rojo de las Plantas Endémicas del Ecuador, Segunda Edición, 2nd ed.; Publicaciones del Herbario QCA; León-Yánez, S., Valencia, R., Pitman, N., Endara, L., Ulloa Ulloa, C., Navarrete, H., Eds.; Pontificia Universidad Católica del Ecuador: Quito, Ecuador, 2012; ISBN 9789942033932. [Google Scholar]
- Froude, V. Wilding Conifers in New Zealand: Status Report; Pacific Eco-Logic Ltd.: Northland, New Zealand, 2011. [Google Scholar]
- Williams, C.M.; Wardle, M.G. Pinus radiata Invasion in New South Wales: The Extent of Spread. Plant Prot. Q. 2009, 24, 146–156. [Google Scholar]
- Palmberg-Lerche, C.; Ball, J.B. Present Status of Forest Plantations in Latin America and the Caribbean and Review of Related Activities in Tree Improvement. Proceedings of El Manejo Sustentable de los Recursos Forestales, Desafío del Siglo XXI, Valdivia, Chile, 22 November 1998. [Google Scholar]
- Vásquez, D.L.A.; Balslev, H.; Sklenář, P. Human Impact on Tropical-Alpine Plant Diversity in the Northern Andes. Biodivers. Conserv. 2015, 24, 2673–2683. [Google Scholar] [CrossRef]
- Verweij, P.A.; Budde, P.E. Burning and Grazing Gradients in Páramo Vegetation: Initial Ordination Analyses. In Páramo. An Andean Ecoystem under Human Influence; Balslev, H., Luteyn, J.L., Eds.; Academic Press Limited: London, UK, 1992; pp. 177–195. [Google Scholar]
- Cavelier, J.; Santos, C. Efectos de Plantaciones Abandonadas de Especies Exóticas y Nativas Sobre la Regeneración Natural de Un Bosque Montano En Colombia. Rev. De Biol. Trop. 1999, 47, 775–784. [Google Scholar] [CrossRef]
- Van Wesenbeeck, B.K.; Van Mourik, T.; Duivenvoorden, J.F.; Cleef, A.M. Strong Effects of a Plantation with Pinus patula on Andean Subpáramo Vegetation: A Case Study from Colombia. Biol. Conserv. 2003, 114, 207–218. [Google Scholar] [CrossRef]
- Bremer, L.L. Land-Use Change, Ecosystem Services, and Local Livelihoods: Ecological and Socioeconomic Outcomes of Payment for Ecosystem Services in Ecuadorian Páramo Grasslands. Ph.D Thesis, San Diego State University, University of California, Santa Barbara, CA, USA, 2012. [Google Scholar]
- Gloser, V.; Gloser, J. Acclimation Capability of Calamagrostis epigejos and C. arundinacea to Changes in Radiation Environment. Photosynthetica 1996, 32, 203–212. [Google Scholar]
- Quiroz-Dahik, C.; Marín, F.; Arias, R.; Crespo, P.; Weber, M.; Palomeque, X. Comparison of Natural Regeneration in Natural Grassland and Pine Plantations across an Elevational Gradient in the Páramo Ecosystem of Southern Ecuador. Forests 2019, 10, 745. [Google Scholar] [CrossRef]
- Brockerhoff, E.G.; Ecroyd, C.E.; Leckie, A.C.; Kimberley, M.O. Diversity and Succession of Adventive and Indigenous Vascular Understorey Plants in Pinus radiata Plantation Forests in New Zealand. For. Ecol. Manag. 2003, 185, 307–326. [Google Scholar] [CrossRef]
- Hall, J.M.; Daniels, A.E.; Lambin, E.F.; Van Holt, T.; Balthazar, V. Trade-Offs between Tree Cover, Carbon Storage and Floristic Biodiversity in Reforesting Landscapes. Landsc. Ecol. Eng. 2012, 27, 1135–1147. [Google Scholar] [CrossRef]
- Sarmiento, L.; Llambí, L.D.; Escalona, A.; Marquez, N. Vegetation Patterns, Regeneration Rates and Divergence in an Old-Field Succession of the High Tropical Andes. Plant Ecol. 2003, 166, 63–74. [Google Scholar] [CrossRef]
- Van der Hammen, T.; Cleef, A. Development of the High Andean Paramo Flora and Vegetation. In High Altitude Tropical Biogeography; Vuilleumier, F., Monasterio, M., Eds.; Oxford University Press: New York, NY, USA, 1986; pp. 153–201. [Google Scholar]
- Secretaria General de la Comunidad Andina; Ministerio del Ambiente del Ecuador; Proyecto de Adaptación al Impacto del Retroceso Acelerado de Glaciares en los Andes Tropicales; Fondo para la Protección del Agua. Diseño de la Red de Monitoreo Hidrológico en las Microcuencas de los Ríos Pita, San Pedro, Papallacta y Antisana; Ministerio del Ambiente del Ecuador: Quito, Ecuador, 2011; pp. 60–104. [Google Scholar]
- FONAG; EPMAPS. Anuario Hidrometeorológico 2019; Fondo para la Protección del Agua: Quito, Ecuador, 2019; Volume 13. [Google Scholar]
- FONAG; EPMAPS. Anuario Hidrometeorológico 2020; Fondo para la Protección del Agua: Quito, Ecuador, 2020; Volume 14. [Google Scholar]
- INAMHI. Anuario Meteorológico; Instituto Nacional de Meteorología e Hidrología del Ecuador: Quito, Ecuador, 2013. [Google Scholar]
- Aguirre, N.; Torres, J.; Velasco-Linares, P. Guía para la Restauración Ecológica En los Páramos del Antisana; FONAG, Ed.; Fondo para la Protección del Agua-FONAG: Quito, Ecuador, 2014; ISBN 9942-9983-3. [Google Scholar]
- Sklenár, P.; Ramsay, P.M. Diversity of Zonal Páramo Plant Communities in Ecuador. Divers. Distrib. 2001, 7, 113–124. [Google Scholar] [CrossRef]
- Beltrán, K.; Salgado, S.; Cuesta, F.; León-Yánez, S.; Romoleroux, K.; Ortiz, E.; Cárdenas, A.; Velástegui, A. Distribución Espacial Sistemas Ecológicos y Caracterización Florística de los Páramos En El Ecuador; EcoCiencia, Proyecto Páramo Andino y Herbario QCA, Ed.; EcoCiencia: Quito, Ecuador, 2009. [Google Scholar]
- Lemmon, P.E. A New Instrument for Measuring Forest Overstory Density. J. For. 1957, 55, 667–668. [Google Scholar]
- Ramsay, P.M.; Oxley, E.R.B. The Growth Form Composition of Plant Communities in the Ecuadorian Páramos. Plant Ecol. 1997, 131, 173–192. [Google Scholar] [CrossRef]
- Romoleroux, K.; Cárate Tandalla, D.; Erler, R.; Navarrete, H. Plantas Vasculares de los Bosques de Polylepis En los Páramos de Oyacachi; Centro de Publicaciones de la Pontificia Universidad Católica del Ecuador: Quito, Ecuador, 2016; ISBN 9978772731. [Google Scholar]
- MECN-INB. Plantas de los Páramos del Distrito Metropolitano de Quito, Ecuador; Ulloa-Ulloa, C., Fernández, D., Eds.; Publicacio.: Quito, Ecuador, 2015; ISBN 9789942998842. [Google Scholar]
- Aguilar, Z.; Hidalgo, P.; Ulloa Ulloa, C. Plantas Útiles de los Páramos de Zuleta, Ecuador; Proyecto d.; PPA-Ecociencia: Quito, Ecuador, 2009; ISBN 9789978994023. [Google Scholar]
- Chimbolema, S.; Suárez-Duque, D.; Peñafiel Cevallos, M.; Acurio, C.; Paredes, T. Guía de Plantas de la Reserva Ecológica El Ángel; Peñafiel Cevallos, M., Ed.; Corporación Grupo Randi Randi: Quito, Ecuador, 2014; ISBN 978-9942-13-550-6. [Google Scholar]
- Van der Maarel, E. Relations between Sociological-Ecological Species Groups and Ellenberg Indicator Values. Phytocoenologia 1993, 23, 343–362. [Google Scholar] [CrossRef]
- Dufrêne, M.; Legendre, P. Species Assemblages and Indicator Species: The Need for a Flexible Asymmetrical Approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Gloser, V.; Scheurwater, I.; Lambers, H. The Interactive Effect of Irradiance and Source of Nitrogen on Growth and Root Respiration of Calamagrostis Epigejos. New Phytol. 1996, 134, 407–412. [Google Scholar] [CrossRef]
- Bader, M.Y.; Van Geloof, I.; Rietkerk, M. High Solar Radiation Hinders Tree Regeneration above the Alpine Treeline in Northern Ecuador. Plant Ecol. 2007, 191, 33–45. [Google Scholar] [CrossRef]
- Jørgensen, P.M.; León-Yánez, S. (Eds.) Catalogue of the Vascular Plants of Ecuador; Missouri Botanical Garden Press: St Louis, MO, USA, 1999. [Google Scholar]
- Ministerio del Ambiente del Ecuador. Sistema de Clasificación de Ecosistemas del Ecuador Continental, 1st ed; Galeas, R., Guevara, J.E., Medina-Torres, B., Chinchero, M.Á., Herrera, X., Eds.; Subsecretaria de Patrimonio Natural: Quito, Ecuador, 2013. [Google Scholar]
- Subsecretaría de Patrimonio Natural; Acosta-Buenaño, N.; Beltrán, L.; Chinchero, M.Á.; Galeas, R.; Herrera, X.; Iglesias, J.; Medina-Torres, B.; Morales, C.; Rivas-Torres, G.; et al. Información florística del Proyecto Mapa de Vegetación para la Clasificación de Ecosistemas del Ecuador Continental. Available online: https://www.gbif.org/dataset/9bbdb0ea-b792-4047-be06-c4285ad47f3f (accessed on 14 December 2020).
- Bremer, L.L.; Farley, K.A. Does Plantation Forestry Restore Biodiversity or Create Green Deserts? A Synthesis of the Effects of Land-Use Transitions on Plant Species Richness. Biodivers. Conserv. 2010, 19, 3893–3915. [Google Scholar] [CrossRef]
- Mena Vasconez, P.; Medina, G.; Hofstede, R.G.M. los Páramos del Ecuador. Particularidades, Problemas y Perspectivas; Mena Vásconez, P., Medina, G., Hofstede, R.G., Eds.; Abya Yala/Proyecto Páramo: Quito, Ecuador, 2001; ISBN 9978047271. [Google Scholar]
- Lemenih, M.; Gidyelew, T.; Teketay, D. Effects of Canopy Cover and Understory Environment of Tree Plantations on Richness, Density and Size of Colonizing Woody Species in Southern Ethiopia. For. Ecol. Manag. 2004, 194, 1–10. [Google Scholar] [CrossRef]
- Corredor-Velandia, S.; Vargas Ríos, O. Efectos de la Creación de Claros Experimentales Con Diferentes Densidades, Sobre los Patrones Iniciales de Sucesión Vegetal En Plantaciones de Pinus patula. In Restauración Ecológica del Bosque Altoandino. Estudios Diagnósticos y Experimentales en los Alrededores del Embalse de Chisacá; Universidad Nacional de Colombia: Bogotá, Colombia, 2007; pp. 336–352. [Google Scholar]
- Matson, E.C.; Bart, D.J. Plant—Community Responses to Shrub Cover in a Páramo Grassland Released from Grazing and Burning. Austral Ecol. 2014, 39, 918–928. [Google Scholar] [CrossRef]
- Grubb, P.J.; Lloyd, J.R.; Pennington, T.D.; Páez-Bimos, S. A Historical Baseline Study of the Páramo of Antisana in the Ecuadorian Andes Including the Impacts of Burning, Grazing and Trampling. Plant Ecol. Divers. 2020, 13, 225–256. [Google Scholar] [CrossRef]
- Podwojewski, P.; Poulenard, J.; Zambrana, T.; Hofstede, R.G. Overgrazing Effects on Vegetation Cover and Properties of Volcanic Ash Soil in the Páramo of Llangahua and la Esperanza (Tungurahua, Ecuador). Soil Use Manag. 2002, 18, 45–55. [Google Scholar] [CrossRef]
- Hofstede, R.G.; Rossenaar, A.J. Biomass of Grazed, Burned, and Undisturbed Paramo Grasslands, Colombia. II. Root Mass and Aboveground: Belowground Ratio. Arct. Alp. Res. 1995, 27, 13–18. [Google Scholar] [CrossRef]
Figure 1.
Location of the study zones with both land uses, pine plantations and grasslands. Coordinates system: World geodetic system 84 (degrees and minutes). (Ant = Antisana, Caj = Cajas, Tis = Tisaleo, Cot = Cotopaxi, G = Grassland and P = pine plantation).
Figure 1.
Location of the study zones with both land uses, pine plantations and grasslands. Coordinates system: World geodetic system 84 (degrees and minutes). (Ant = Antisana, Caj = Cajas, Tis = Tisaleo, Cot = Cotopaxi, G = Grassland and P = pine plantation).

Figure 2.
Canonical correspondence analysis of grasslands and pine plantations in Ecuadorian Páramo. CCA to discriminate the study zones and the land-use types based on plant species abundance. The red ellipse encompasses the grassland plots. The red line ending with an arrow shows the increasing age of pine stands. Ant = Antisana, Caj = Cajas, Tis = Tisaleo, Cot = Cotopaxi, BA = basal zone, CC = pine canopy cover, Alt = altitude and Age = stand age. Each black dot represents one plant species. Indicator species (Supplementary Table S1): Pe. sp.: Peperomia sp., L. pub = Lupinus pubescens, R. vul = Rhynchospora vulcanica, Ox. pha = Oxalis phaeotricha, G. man = Geranium maniculatum, Gu. mag = Gunnera magellanica, P. lin = Plantago linearis, Ga. hyp = Galium hypocarpium, Co. spp. = Cortaderia nitida and C. jubata, M. cra = Monnina crassifolia, V. flo = Vaccinum floribundum, Gyn. min = Gynoxys miniphylla, O. and = Oreomyrrhis andicola.
Figure 2.
Canonical correspondence analysis of grasslands and pine plantations in Ecuadorian Páramo. CCA to discriminate the study zones and the land-use types based on plant species abundance. The red ellipse encompasses the grassland plots. The red line ending with an arrow shows the increasing age of pine stands. Ant = Antisana, Caj = Cajas, Tis = Tisaleo, Cot = Cotopaxi, BA = basal zone, CC = pine canopy cover, Alt = altitude and Age = stand age. Each black dot represents one plant species. Indicator species (Supplementary Table S1): Pe. sp.: Peperomia sp., L. pub = Lupinus pubescens, R. vul = Rhynchospora vulcanica, Ox. pha = Oxalis phaeotricha, G. man = Geranium maniculatum, Gu. mag = Gunnera magellanica, P. lin = Plantago linearis, Ga. hyp = Galium hypocarpium, Co. spp. = Cortaderia nitida and C. jubata, M. cra = Monnina crassifolia, V. flo = Vaccinum floribundum, Gyn. min = Gynoxys miniphylla, O. and = Oreomyrrhis andicola.

Figure 3.
Pine effects on plant cover (total, types of vegetation and life forms) and species richness. Formula for exponential or linear trend with their respective R2 and model p value are presented in each scatter plot. Color varies among study areas and therefore also reflects pine age.
Figure 3.
Pine effects on plant cover (total, types of vegetation and life forms) and species richness. Formula for exponential or linear trend with their respective R2 and model p value are presented in each scatter plot. Color varies among study areas and therefore also reflects pine age.

Table 1.
Climate, soil and stand characteristics of the four study zones.
| Environmental Variables | Antisana | Cajas | Tisaleo | Cotopaxi |
|---|---|---|---|---|
| Mean annual temperature (°C) [50,51,52,53] | 8.3 | 10 * | 10 * | 11 * |
| Mean annual rainfall (mm) [52,53] | 900 | 1200 * | 900 * | 1110 * |
| Mean altitude in grasslands (m a.s.l.) | 3739 | 3734 | 3858 | 3695 |
| Mean altitude in pine plantations (m a.s.l.) | 3556 | 3699 | 3874 | 3740 |
| Age of pine plantation | 8 | 16 | 41 | 53 |
| Pine tree cover (%) | 75 | 86 | 66 | 47 |
| Basal area of pine trees (m2/ha) | 17 | 10.5 | 26 | 29.5 |
| Mean height of pine trees (m) | 8 | 15 | 15 | 18 |
| pH H2O under grassland and pine plantation (10–30 cm depth) + | 5.8–6.2 | 6.2–6.4 | 6.1–5.7 | 6.2–5.6 |
| Mean slope of grasslands and pine plantation plots + | 15.8–19.1 | 26.1–30.1 | 14.4–14.7 | 27.7–27.6 |
| Slope of grasslands and pine plantation plots | S–SW | NE–SE | SE–SE | N–N |
* These climate data come from the closest meteorological station, which is located at about 2 km from the study zone and is lower in altitude (about 400 m less). + No statistical differences were found between grasslands and pine plantations.
Table 2.
Comparison of the plant composition and diversity among the four study zones (Antisana, Cajas, Tisaleo and Cotopaxi) and the land-use types (grasslands vs. pine plantations).
Table 2.
Comparison of the plant composition and diversity among the four study zones (Antisana, Cajas, Tisaleo and Cotopaxi) and the land-use types (grasslands vs. pine plantations).
| Antisana | Cajas | Tisaleo | Cotopaxi | Contribution to R2 | Change under Pine (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Grassland | Pine Plantation | Grassland | Pine Plantation | Grassland | Pine Plantation | Grassland | Pine Plantation | Zone | Land Use | Z × LU | ||
| Location WGS 84 | 0°00′13″ S 78°05′11″ W | 0°07′1.2 S 78°05′36″ W | 2°01′00″ S 79°03′0.4″ W | 2°01′00″ S 79°03′36″ W | 1°06′00″ S 78°01′31″ W | 1°06′53″ S 78°11′0.1″ W | 0°01′11″ S 78°08′36″ W | 0°10′12″ S 78°08′00″ W | ||||
| Altitude (m a.s.l.) | 3740 | 3560 | 3735 | 3698 | 3858 | 3871 | 3695 | 3742 | ||||
| Herbaceous height (cm) | 80 | 15 | 60 | 20 | 60 | 25 | 100 | 60 | ||||
| Shrub height (cm) | 50 | 20 | 140 | 40 | 100 | 50 | 120 | 40 | ||||
| Plant cover (%) | ||||||||||||
| Total cover | 101.5 ± 3.3 | 66.3 ± 39.72 | 94.0 ± 4.7 | 3.4 ± 3.4 *** | 85.0 ± 23.5 | 55.5 ± 21.3 ** | 92.2 ± 19.1 | 57.4 ± 7.3 *** | 0.14 | 0.46 | 0.13 | −47.5 *** |
| Moss cover | 2.5 ± 1.2 | 1.7 ± 2.1 | 2.5 ± 1.7 | 10.5 ± 9.4 | 23.6 ± 14 | 9.9 ± 5.4 | 17.3 ± 15.2 | 8.6 ± 10.6 ** | 0.26 | 0.03 | 0.13 | −3.8 |
| Woody species | 3.9 ± 2.1 | 4.3 ± 6.5 | 2.0 ± 1.7 | 1.1 ± 1.0 | 20.2 ± 12.0 | 9.9 ± 9.3 * | 30.8 ± 9.7 | 12.0 ± 3.7 ** | 0.48 | 0.10 | 0.11 | −7.4 * |
| Herbaceous | 97.6 ± 2.6 | 62.0 ± 38.9 * | 92.0 ± 5.0 | 2.3 ± 3.0 *** | 64.8 ± 15.8 | 45.6 ± 15.4 * | 61.4 ± 17.3 | 45.4 ± 5.9 | 0.15 | 0.39 | 0.21 | −40.1 *** |
| Tussocks | 86.3 ± 5.4 | 13.1 ± 28.6 *** | 83.1 ± 5.7 | 0.1 ± 0.3 *** | 41.3 ± 13.7 | 0.5 ± 0.9 *** | 39.9 ± 13 | 33.6 ± 5.8 | 0.10 | 0.57 | 0.20 | −50.8 *** |
| Cushions | 2.2 ± 1.0 | 0.1 ± 0.3 * | 1.6 ± 1.5 | 0.03 ± 0.05 | 8.6 ± 5.7 | 0.4 ± 1.1 *** | 1.4 ± 1.6 | 0.0 | 0.21 | 0.24 | 0.17 | −3.4 *** |
| Rosettes | 0.5 ± 0.3 | 0.04 ± 0.1 * | 4.3 ± 2.2 | 0.9 ± 1.2 *** | 0.8 ± 0.8 | 0.7 ± 0.4 | 11.3 ± 6.3 | 0.9 ± 1.2 *** | 0.29 | 0.18 | 0.24 | −3.6 *** |
| Small herbs | 8.6 ± 3.9 | 48.8 ± 42.8 | 3.0 ± 1.1 | 1.2 ± 2.0 | 14.1 ± 11.2 | 44.1 ± 14.9 *** | 8.8 ± 3.8 | 10.9 ± 4.5 | 0.26 | 0.14 | 0.15 | 17.6 ** |
| Light-demanding species cover | 89.7 ± 5.0 | 13.3 ± 28.5 *** | 91.2 ± 5.3 | 1.1 ± 1.2 *** | 51.9 ± 14.9 | 2.0 ± 1.9 *** | 54 ± 18.2 | 34.5 ± 5.1 ** | 0.07 | 0.67 | 0.14 | −59.0 *** |
| Shade-tolerant species cover | 11.8 ± 5.5 | 53.2 ± 44.4 * | 2.8 ± 1.7 | 2.4 ± 2.4 | 33.2 ± 14.3 | 56.1 ± 20.8 | 38.2 ± 12.1 | 23.8 ± 7.2 | 0.34 | 0.05 | 0.16 | 12.4 |
| Species richness | ||||||||||||
| Total | 21.8 ± 1.6 | 13.8 ± 5.4 ** | 29.7 ± 4.9 | 13.9 ± 5.7 *** | 22.2 ± 6.3 | 13.9 ± 3.9 *** | 21.9 ± 2.8 | 23.1 ± 2.8 | 0.10 | 0.32 | 0.20 | −7.7 *** |
| Herbaceous | 17.7 ± 1.5 | 9.7 ± 3.5 *** | 23.4 ± 5.8 | 8.3 ± 4.5 *** | 17.2 ± 5.5 | 10.6 ± 3 ** | 13.4 ± 2.3 | 13 ± 2.8 | 0.03 | 0.40 | 0.19 | −7.5 *** |
| Woody species | 4.1 ± 0.6 | 4.1 ± 2.2 | 6.2 ± 2.3 | 5.6 ± 1.5 | 5.0 ± 1.0 | 3.3 ± 1.3 | 8.4 ± 1.5 | 10.0 ± 1.8 | 0.62 | 0.00 | 0.05 | −0.2 |
Z × LU: Interaction between zone and land use. Coordinates datum: World geodetic system (WGS). Change under pine trees was taken from raw model and their significance was measured after BoxCox transformation for cover variables. Contrast analyses significance are presented in pine plantation columns. (Superscripts show significance p-value: * ≤0.05, ** ≤0.01, *** ≤0.001, the Bonferroni correction was applied to account for the fact that multiple tests were performed).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Medina-Torres, B.; Jonard, M.; Rendón, M.; Jacquemart, A.-L. Effects of Pine Plantation on Native Ecuadorian Páramo Vegetation. Forests 2022, 13, 1499. https://doi.org/10.3390/f13091499
AMA Style
Medina-Torres B, Jonard M, Rendón M, Jacquemart A-L. Effects of Pine Plantation on Native Ecuadorian Páramo Vegetation. Forests. 2022; 13(9):1499. https://doi.org/10.3390/f13091499
Chicago/Turabian StyleMedina-Torres, Byron, Mathieu Jonard, Melina Rendón, and Anne-Laure Jacquemart. 2022. "Effects of Pine Plantation on Native Ecuadorian Páramo Vegetation" Forests 13, no. 9: 1499. https://doi.org/10.3390/f13091499
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.