
Litter Inputs Control the Pattern of Soil Aggregate-Associated Organic Carbon and Enzyme Activities in Three Typical Subtropical Forests

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design and Soil Sampling
2.3. Soil Aggregate Separation
2.4. Soil Sample Analyses
2.5. Statistical Analyses
3. Results
3.1. Basic Physical and Chemical Properties of Soils
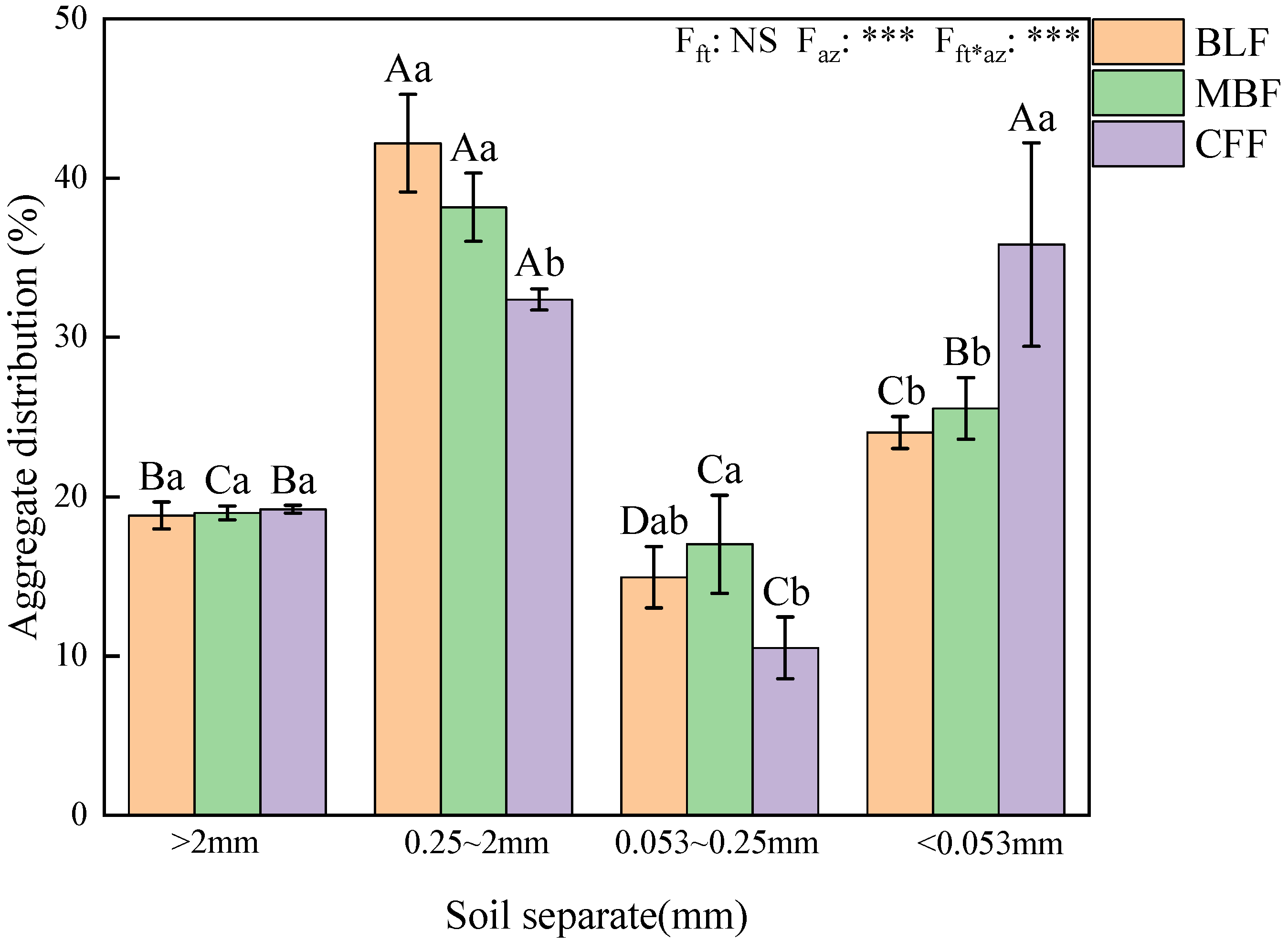
3.2. Distribution Characteristics of Soil Aggregates
3.3. Stability Characteristics of Soil Aggregates
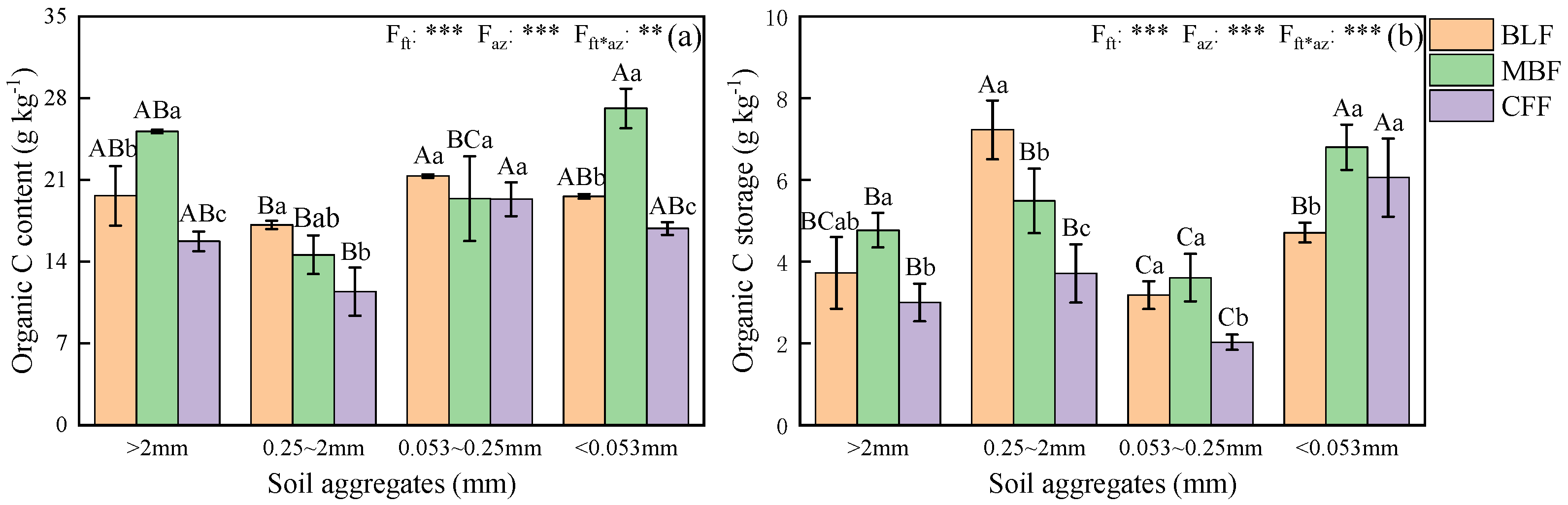
3.4. Aggregate-Associated SOC Concentrations and Storage
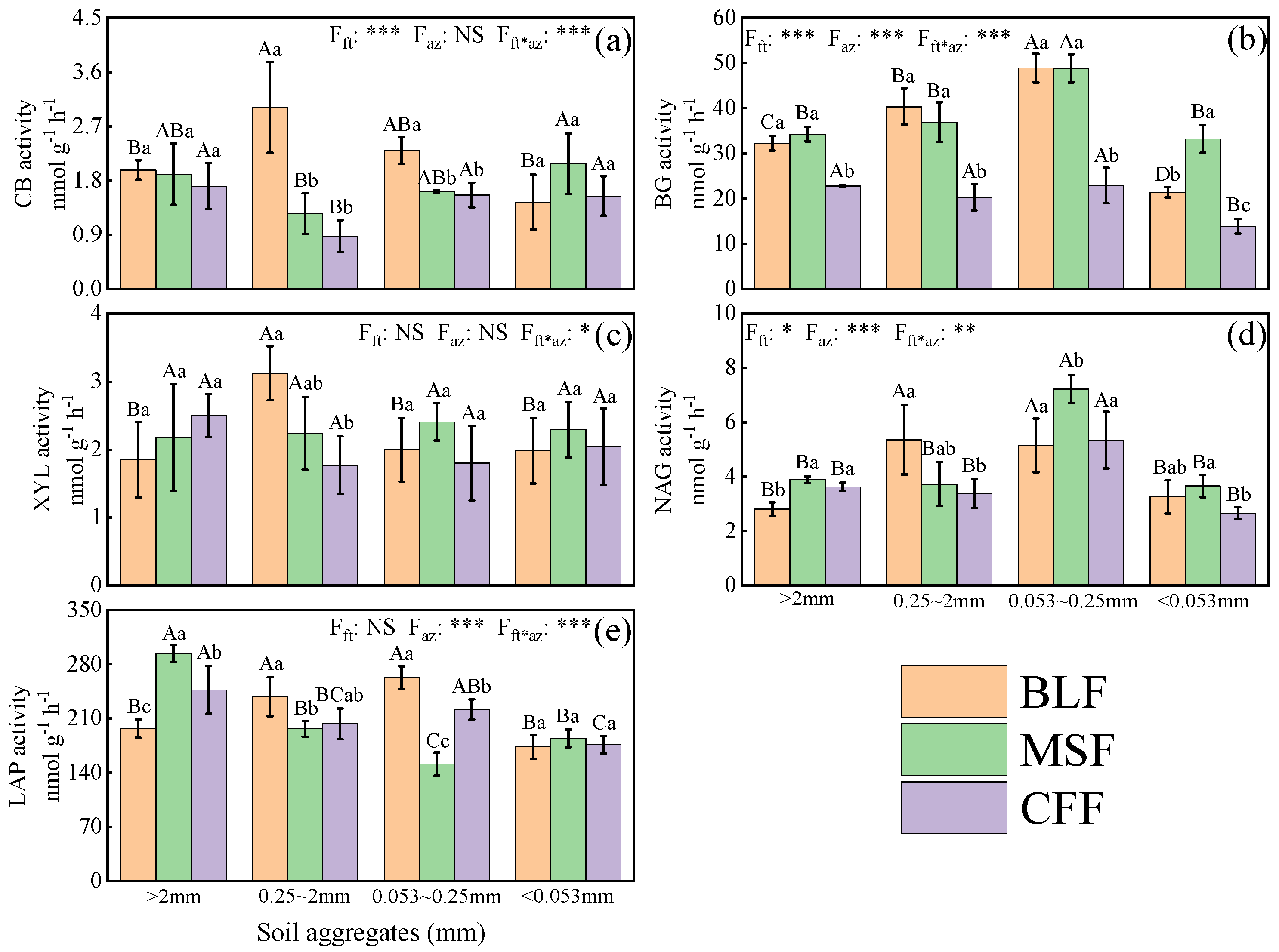
3.5. Soil Enzyme Activities
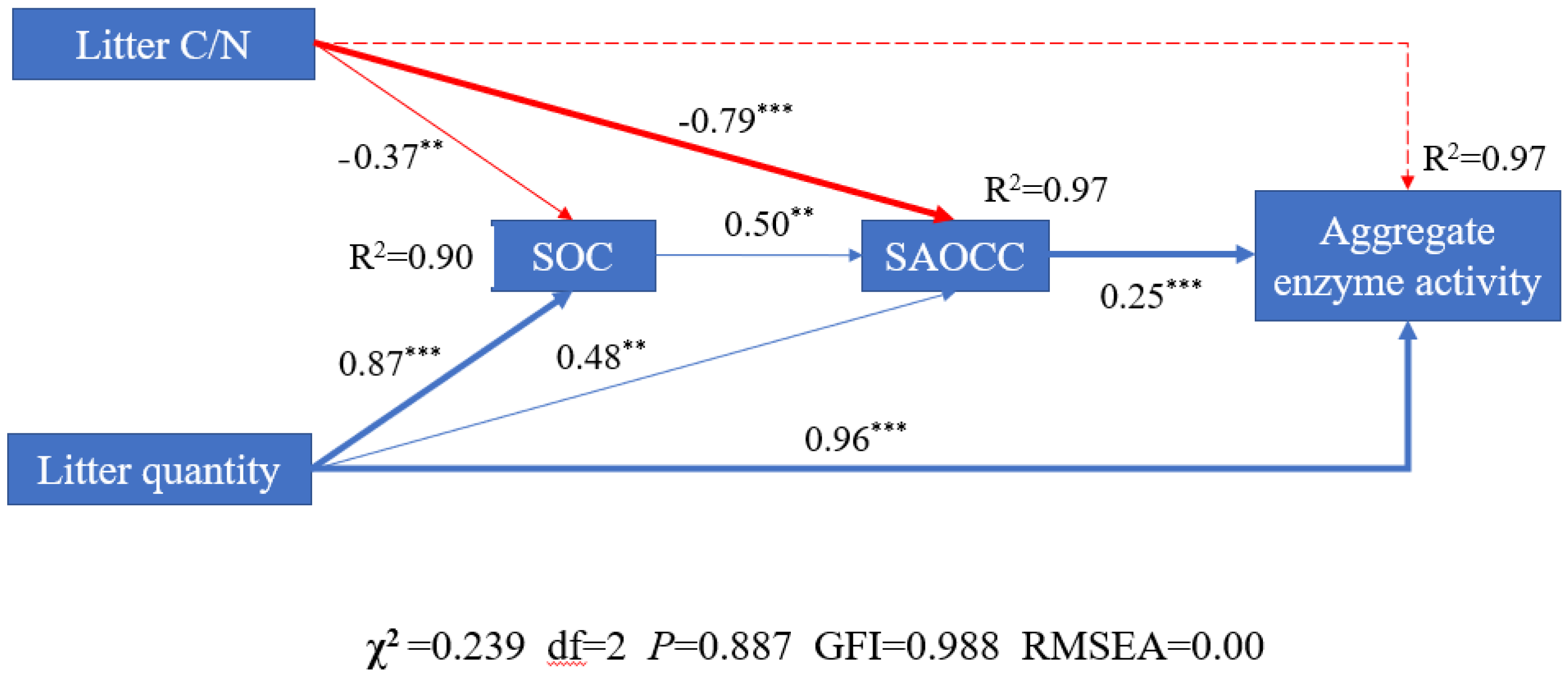
3.6. Relationship between Litter C/N, Litter Quantity, Soil Aggregate Organic C Contents and Aggregate Enzyme Activity
4. Discussion
4.1. Effect of Forest Types on the Distribution and Stability of Soil Aggregates
4.2. Effect of Forest Type on Soil Aggregate-Associated Carbon Content and Storage
4.3. Activities of Soil Aggregate-Related C and N-Cycling Enzymes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Khan, K.S.; Mack, R.; Castillo, X.; Kaiser, M.; Joergensen, R.G. Microbial biomass, fungal and bacterial residues, and their rela tionships to the soil organic matter C/N/P/S ratios. Geoderma 2016, 271, 115–123. [Google Scholar] [CrossRef]
- Jing, X.; Chen, X.; Xiao, W.; Lin, L.; Wang, C.; He, J.S.; Zhu, B. Soil enzymatic responses to multiple environmental drivers in the Tibetan grasslands. Insights from two manipulative field experiments and a meta-analysis. Pedobiologia 2018, 71, 50–58. [Google Scholar] [CrossRef]
- Bach, E.M.; Hofmockel, K.S. Soil aggregate isolation method affects measures of intra-aggregate extracellular enzyme activity. Soil Biol. Biochem. 2014, 69, 54–62. [Google Scholar] [CrossRef]
- Qi, R.; Li, J.; Lin, Z.; Li, Z.; Li, Y.; Yang, X.; Zhang, J.; Zhao, B. Temperature effects on soil organic carbon, soil labile organic carbon fractions, and soil enzyme activities under long-term fertilization regimes. Appl. Soil Ecol. 2016, 102, 36–45. [Google Scholar] [CrossRef]
- Mori, T.; Imai, N.; Yokoyama, D.; Kitayama, K. Effects of nitrogen and phosphorus fertilization on the ratio of activities of carbon-acquiring to nitrogen-acquiring enzymes in a primary lowland tropical rainforest in Borneo, Malaysia. Soil Sci. Plant. Nutr. 2018, 64, 554–557. [Google Scholar] [CrossRef]
- Zheng, T.T.; Liang, C.; Xie, H.T.; Zhao, J.S.; Yan, E.S.; Zhou, X.H.; Bao, X.L. Rhizosphere effects on soil microbial community structure and enzyme activity in a successional subtropical forest. FEMS Microbiol Ecol. 2019, 95, 43. [Google Scholar] [CrossRef]
- Six, J.; Bossuyt, H.; Degryze, S.; Denef, K. A history of research on the link between (micro)aggregates, soil biota, and soil organic matter dynamics. Soil Tillage Res. 2004, 79, 7–31. [Google Scholar] [CrossRef]
- Tisdall, J.; Oades, J.M. Organic matter and water- stable aggregates in soils. Eur. J. Soil Sci. 1982, 33, 141–163. [Google Scholar] [CrossRef]
- Mummey, D.L.; Stahl, P.D. Analysis of soil whole- and inner-microaggregate bacterial communities. Microb. Ecol. 2004, 48, 41–50. [Google Scholar] [CrossRef]
- Tisdall, J.M.; Oades, J.M. Landmark Papers: No. 1. Organic matter and water-stable aggregates in soils. Eur. J. Soil Sci. 2012, 63, 8–21. [Google Scholar]
- Oades, J.M.; Waters, A.G. Aggregate hierarchy in soils. Soil Res. 1991, 29, 815–828. [Google Scholar] [CrossRef]
- Almajmaie, A.; Hardie, M.; Doyle, R.; Birch, C.; Acuna, T. Influence of soil properties on the aggregate stability of cultivated sandy clay loams. J. Soils Sediments 2017, 17, 800–809. [Google Scholar] [CrossRef]
- Allison, S.D.; Jastrow, J.D. Activities of extracellular enzymes in physically isolated fractions of restored grassland soils. Soil Biol. Biochem. 2006, 38, 3245–3256. [Google Scholar] [CrossRef]
- FAO. Global Forest Resources Assessment 2020; FAO: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- Bai, K.D.; He, C.X.; Wan, X.C.; Jiang, D.B. Leaf economics of evergreen and deciduous tree species along an elevational gradient in a subtropical mountain. AoB Plants 2015, 7, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.L.; Shalom, A.D.D.; Wu, P.F.; Li, S.B.; Jia, Y.Y.; Ma, X.Q. Litterfall production and nutrient return in different-aged Chinese fir(Cunninghamia lanceolata) plantations in South China. J. For. Res. 2015, 26, 79–89. [Google Scholar] [CrossRef]
- Xu, Q.F.; Liang, C.F.; Chen, J.H.; Li, Y.C.; Qin, H.; Fuhrmann, J.J. Rapid bamboo invasion (expansion) and its effects on biodiversity and soil processes +. Glob. Ecol. Conserv. 2020, 21, e00787. [Google Scholar] [CrossRef]
- Van Bavel, C.H.M. Mean Weight-Diameter of Soil Aggregates as a Statistical Index of Aggregation 1. Soil Sci. Soc. Am. J. 1950, 14, 20–23. [Google Scholar] [CrossRef] [Green Version]
- Duan, L.X.; Sheng, H.; Yuan, H.; Zhou, Q.; Li, Z.W. Land use conversion and lithology impacts soil aggregate stability in subtropical China. Geoderma 2021, 389, 114953. [Google Scholar] [CrossRef]
- Castrignanò, A.; Stelluti, M. Fractal Geometry and Geostatistics for describing the Field Variability of Soil Aggregation. J. Agric. Eng. Res. 1998, 73, 13–18. [Google Scholar] [CrossRef]
- Lu, R.K. Methods of Soil and Agro-Chemical Analysis; China Agricultural Science and Technology Press: Beijing, China, 2000. [Google Scholar]
- Saiya-Cork, K.R.; Sinsabaugh, R.L.; Zak, D.R. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil. Soil Biol. Biochem. 2002, 34, 1309–1315. [Google Scholar] [CrossRef]
- Aye, Y.Y.; Pampasit, S.; Umponstira, C.; Thanacharoenchanaphas, K.; Sasaki, N. Estimation of Carbon Emission Reductions by Managing Dry Mixed Deciduous Forest: Case Study in Popa Mountain Park. Low Carbon Econ. 2014, 5, 80–93. [Google Scholar] [CrossRef] [Green Version]
- Liang, C.; Schimel, J.P.; Jastrow, J.D. The importance of anabolism in microbial control over soil carbon storage. Nat. Microbiol. 2017, 2, 17105. [Google Scholar] [CrossRef] [PubMed]
- Cotrufo, M.F.; Wallenstein, M.D.; Boot, C.M.; Denef, K.; Paul, E. The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter? Glob. Chang. Biol. 2013, 19, 988–995. [Google Scholar] [CrossRef] [Green Version]
- Kögel-Knabner, I. The macromolecular organic composition of plant and microbial residues as inputs to soil organic matter. Soil Biol. Biochem. 2002, 34, 139–162. [Google Scholar] [CrossRef]
- Dornbush, M.E. Grasses, litter, and their interaction affect microbial biomass and soil enzyme activity. Soil Biol. Biochem. 2007, 39, 2241–2249. [Google Scholar] [CrossRef]
- Hu, Y.L.; Wang, S.L.; Zeng, D.H. Effects of Single Chinese Fir and Mixed Leaf Litters on Soil Chemical, Microbial Properties and Soil Enzyme Activities. Plant Soil 2006, 282, 379–386. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Lauber, C.L.; Weintraub, M.N.; Ahmed, B.; Allison, S.D.; Crenshaw, C.; Contosta, A.R.; Cusack, D.; Frey, S.; Gallo, M.E.; et al. Stoichiometry of soil enzyme activity at global scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
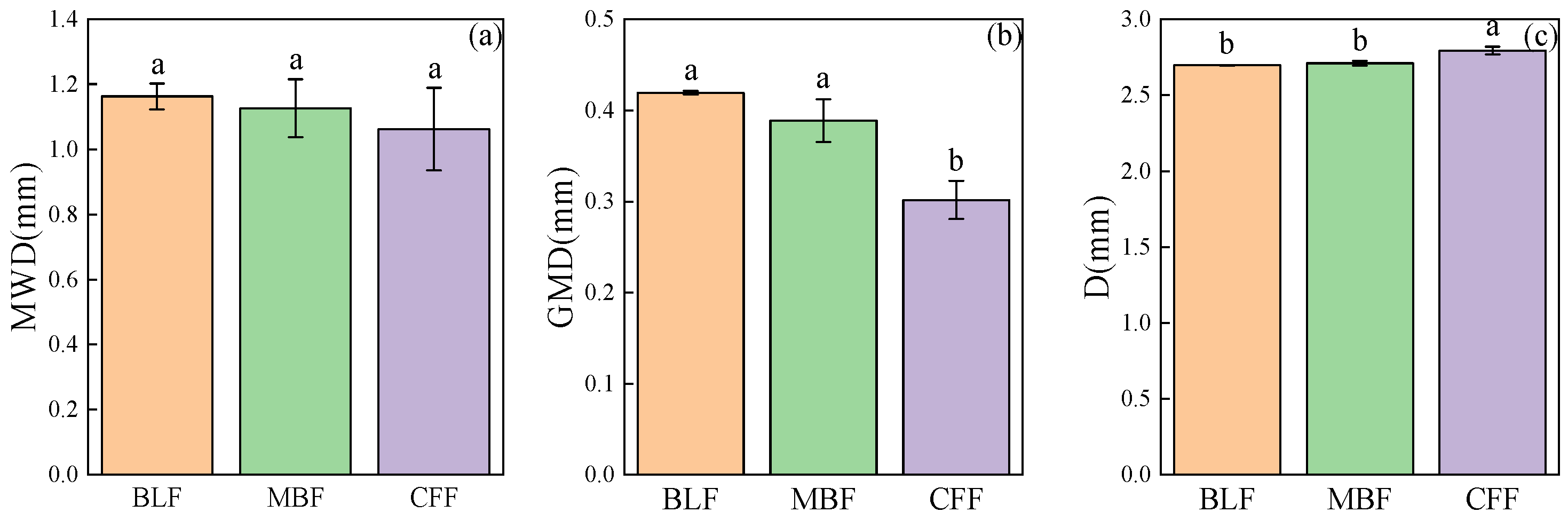{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites | Longitude (°E) | Latitude (°Nbr) | MAT (°C) | MAP (mm) | Altitude (m) | Parent Material | Soil Type | Canopy Density |
|---|---|---|---|---|---|---|---|---|
| LH | 121°4′52″ | 28°50′44″ | 17.1 | 1550 | 202 | Granite | Ferralsol | 0.92 |
| LQ | 118°45′55″ | 28°00′10″ | 17.6 | 1699 | 193 | Granite | Ferralsol | 0.75 |
| HZ | 119°40′41″ | 30°13′9″ | 16.0 | 1613 | 210 | Granite | Ferralsol | 0.87 |
| Forest Types | BLF | MBF | CFF |
|---|---|---|---|
| Canopy density | 0.92 ± 0.02 a | 0.63 ± 0.02 b | 0.77 ± 0.03 c |
| Annual litter production (t hm−2) | 11.11 ± 1.63 a | 5.71 ± 0.37 b | 7.25 ± 0.31 b |
| Litter C/N | 35.33 ± 2.28 b | 31.3 ± 1.96 b | 49.76 ± 4.62 a |
| Soil organic carbon (g kg−1) | 18.47 ± 1.11 a | 13.08 ± 0.61 b | 10.82 ± 1.44 c |
| Total N (g kg−1) | 1.61 ± 0.32 a | 1.66 ± 0.12 a | 0.9 ± 0.07 b |
| C/N | 11.83 ± 0.93 a | 7.91 ± 0.69 b | 12.19 ± 0.86 a |
| Alkaline N (mg kg−1) | 47.83 ± 7.29 a | 35.93 ± 2.38 b | 28.93 ± 2.25 b |
| pH | 4.37 ± 0.07 b | 4.83 ± 0.13 a | 4.65 ± 0.15 a |
| Available K (mg kg−1) | 89.33 ± 8.62 a | 64.67 ± 7.37 b | 73.67 ± 3.21 b |
| Sand (2~0.02 mm)% | 33.13 ± 5.23 a | 26.86 ± 8.54 a | 35.71 ± 7.09 a |
| Silt (0.02~0.002 mm)% | 40.57 ± 8.21 a | 40.63 ± 11.42 a | 35.29 ± 5.16 a |
| Clay (<0.002 mm)% | 26.30 ± 9.36 a | 32.51 ± 10.26 a | 29.00 ± 6.42 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Wang, Z.; Fan, B.; Mao, X.; Luo, H.; Jiang, F.; Liang, C.; Chen, J.; Qin, H.; Xu, Q.; et al. Litter Inputs Control the Pattern of Soil Aggregate-Associated Organic Carbon and Enzyme Activities in Three Typical Subtropical Forests. Forests 2022, 13, 1210. https://doi.org/10.3390/f13081210
Wang S, Wang Z, Fan B, Mao X, Luo H, Jiang F, Liang C, Chen J, Qin H, Xu Q, et al. Litter Inputs Control the Pattern of Soil Aggregate-Associated Organic Carbon and Enzyme Activities in Three Typical Subtropical Forests. Forests. 2022; 13(8):1210. https://doi.org/10.3390/f13081210
Chicago/Turabian StyleWang, Shanshan, Zhongqian Wang, Bo Fan, Xiahua Mao, Heng Luo, Feiyan Jiang, Chenfei Liang, Junhui Chen, Hua Qin, Qiufang Xu, and et al. 2022. "Litter Inputs Control the Pattern of Soil Aggregate-Associated Organic Carbon and Enzyme Activities in Three Typical Subtropical Forests" Forests 13, no. 8: 1210. https://doi.org/10.3390/f13081210