
Shrub Diversity and Niche Characteristics in the Initial Stage of Reconstruction of Low-Efficiency Cupressus funebris Stands

Abstract
:1. Introduction
2. Materials and Methods
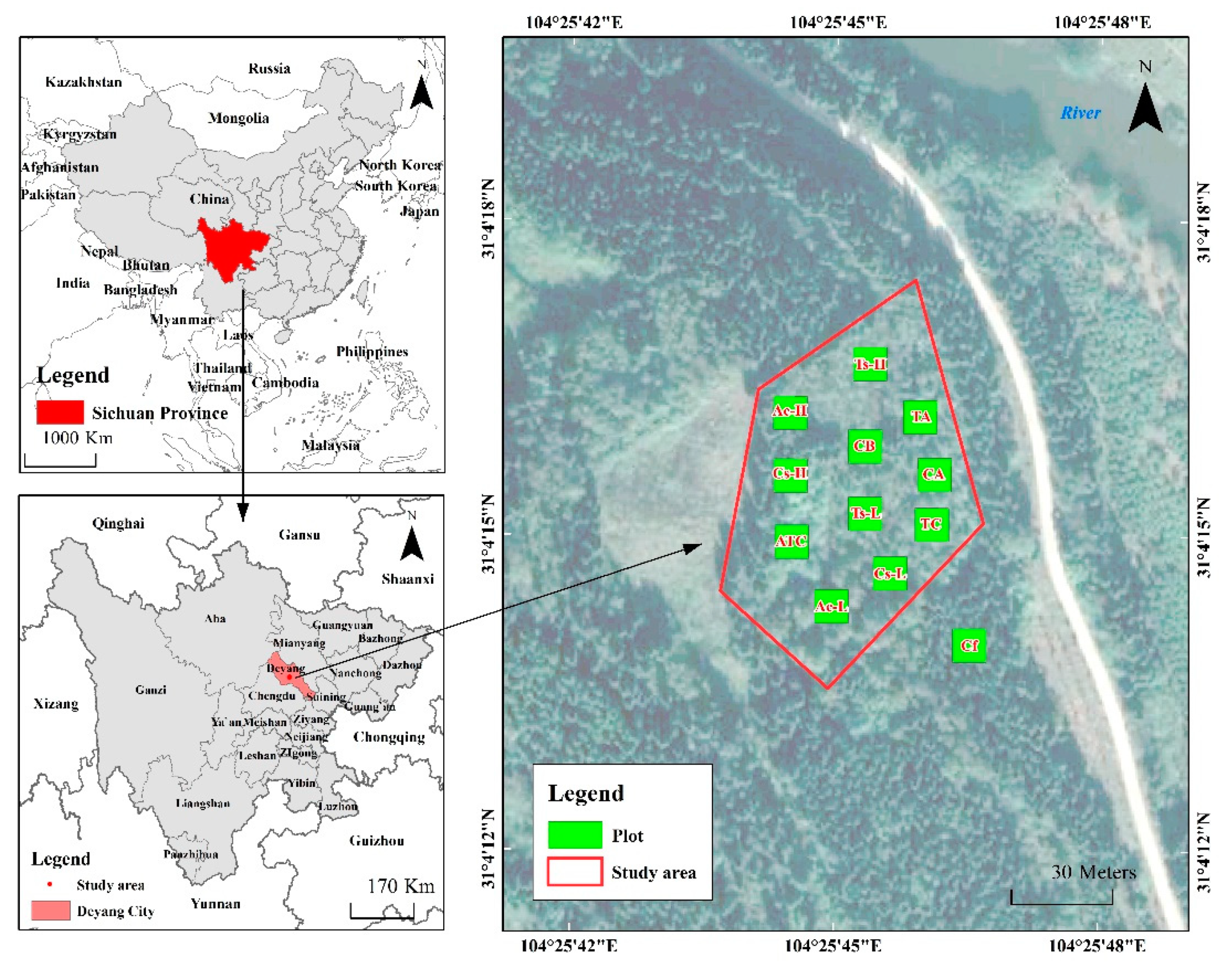
2.1. Study Area
2.2. Survey Methods
2.3. Measurement Methods
- (1)
- Diversity
- (a)
- The Shannon–Wiener (H) and Simpson (H′) indexes were used for the calculation of the diversity index:
- (b)
- The Pielou index was used as the evenness index:
- (c)
- Margalef richness index:where S is the number of species in the sample; N is the total quantity of species; ; and ni is the number of species i.
- (2)
- Importance values = (relative density + relative frequency + relative coverage)/3
- (3)
- Niche breadth
- (4)
- Niche overlap
- (5)
- Canopy closure
2.4. Data Processing and Statistical Analysis
3. Results
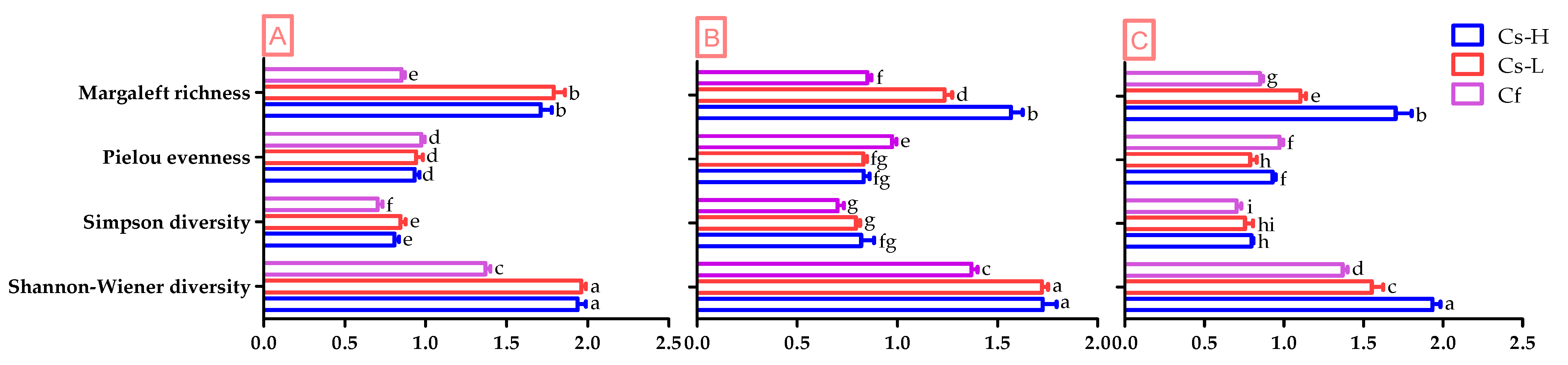
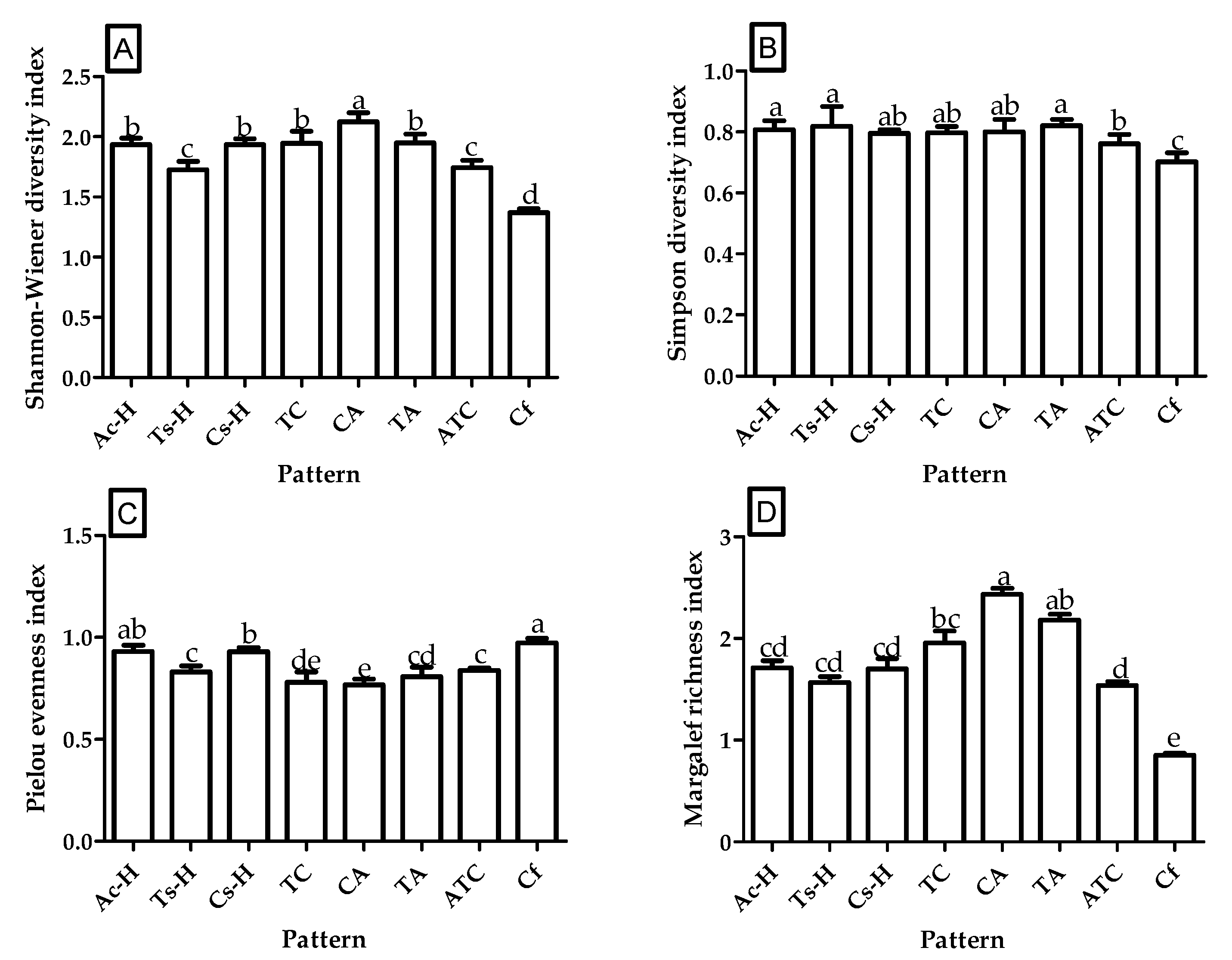
3.1. Shrub Diversity among Different Overstory Densities
3.2. Shrub Niche Breadth
3.3. Shrub Importance Values and Niche Overlap
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tiscar-Oliver, P.A. Patterns of shrub diversity and tree regeneration across topographic and stand-structural gradients in a Mediterranean forest. For. Syst. 2015, 24, 11. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Bu, Y.; Li, Y.; Li, W.; Zhao, P.; Yang, Y.; Qi, N.; Gou, R. Quantifying the relationship among impact factors of shrub layer diversity in Chinese pine plantation forest ecosystems. Forests 2019, 10, 781. [Google Scholar] [CrossRef] [Green Version]
- Foronda, A.; Pueyo, Y.; Arroyo, A.I.; Saiz, H.; de la Luz Giner, M.; Alados, C.L. The role of nurse shrubs on the spatial patterning of plant establishment in semi-arid gypsum plant communities. J. Arid Environ. 2019, 160, 82–90. [Google Scholar] [CrossRef]
- Laanisto, L.; Tamme, R.; Hiiesalu, I.; Szava-Kovats, R.; Gazol, A.; Pärtel, M. Microfragmentation concept explains non-positive environmental heterogeneity-diversity relationships. Oecologia 2013, 171, 217–226. [Google Scholar] [CrossRef]
- Costa-Pereira, R.; Araújo, M.S.; Souza, F.L.; Ingram, T. Competition and resource breadth shape niche variation and overlap in multiple trophic dimensions. Proc. R. Soc. B Biol. Sci. 2019, 286. [Google Scholar] [CrossRef] [Green Version]
- Bar-Massada, A. Complex relationships between species niches and environmental heterogeneity affect species co-occurrence patterns in modelled and real communities. Proc. R. Soc. B Biol. Sci. 2015, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pistón, N.; Schöb, C.; Armas, C.; Prieto, I.; Pugnaire, F.I. Contribution of co-occurring shrub species to community richness and phylogenetic diversity along an environmental gradient. Perspect. Plant Ecol. Evol. Syst. 2016, 19, 30–39. [Google Scholar] [CrossRef] [Green Version]
- Markgraf, R.; Doyon, F.; Kneeshaw, D. Forest landscape heterogeneity increases shrub diversity at the expense of tree seedling diversity in temperate mixedwood forests. Forests 2020, 11, 160. [Google Scholar] [CrossRef] [Green Version]
- Velázquez, E.; Paine, C.E.T.; May, F.; Wiegand, T. Linking trait similarity to interspecific spatial associations in a moist tropical forest. J. Veg. Sci. 2015, 26, 1068–1079. [Google Scholar] [CrossRef]
- D’Andrea, R.; Ostling, A. Biodiversity maintenance may be lower under partial niche differentiation than under neutrality. Ecology 2017, 98, 3211–3218. [Google Scholar] [CrossRef] [PubMed]
- Elliott, T.L.; Davies, T.J. Jointly modeling niche width and phylogenetic distance to explain species co-occurrence. Ecosphere 2017, 8, e01891. [Google Scholar] [CrossRef]
- Nilsson, M.C.; Wardle, D.A. Understory Vegetation as a Forest Ecosystem Driver: Evidence from the Northern Swedish Boreal Forest. Ecol. Soc. Am. 2005, 3, 421–428. [Google Scholar] [CrossRef]
- Gómez-Aparicio, L.; Zavala, M.A.; Bonet, F.J.; Zamora, R. Are pine plantations valid tools for restoring Mediterranean forests? An assessment along abiotic and biotic gradients. Ecol. Appl. 2009, 19, 2124–2141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, M.; Ma, L.; Jia, Y.; Liu, M. Integrating the effects of latitude and altitude on the spatial differentiation of plant community diversity in a mountainous ecosystem in China. PLoS ONE 2017, 12, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Légaré, S.; Bergeron, Y.; Paré, D. Influence of forest composition on understory cover in boreal mixed-wood forests of western Quebec. Silva Fenn. 2002, 36, 353–366. [Google Scholar] [CrossRef] [Green Version]
- Bergstedt, J.; Milberg, P. The impact of logging intensity on field-layer vegetation in swedish boreal forests. For. Ecol. Manag. 2001, 154, 105–115. [Google Scholar] [CrossRef]
- Li, Y.; He, J.; Lu, L.; Xu, J.; Wang, H.; Ye, S. The long-term effects of thinning and mixing on species and structural diversity of Chinese fir plantations. NEW For. 2020, 52, 285–302. [Google Scholar] [CrossRef]
- Ewald, J. The partial influence of Norway spruce stands on understorey vegetation in Montane forests of the Bavarian Alps. Mt. Res. Dev. 2000, 20, 364–371. [Google Scholar] [CrossRef] [Green Version]
- Thomsen, R.P.; Svenning, J.C.; Balslev, H. Overstorey control of understorey species composition in a near-natural temperate broadleaved forest in Denmark. Plant Ecol. 2005, 181, 113–126. [Google Scholar] [CrossRef]
- Huo, H.; Feng, Q.; Su, Y.H. The influences of canopy species and topographic variables on understory species diversity and composition in coniferous forests. Sci. World J. 2014, 2014, 252489. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Calcerrada, J.; Nanos, N.; del Rey, M.C.; de Heredia, U.L.; Escribano, R.; Gil, L. Small-scale variation of vegetation in a mixed forest understorey is partly controlled by the effect of overstory composition on litter accumulation. J. For. Res. 2011, 16, 473–483. [Google Scholar] [CrossRef] [Green Version]
- Yılmaz, O.Y.; Yılmaz, H.; Akyüz, Y.F. Effects of the overstory on the diversity of the herb and shrub layers of Anatolian black pine forests. Eur. J. For. Res. 2018, 137, 433–445. [Google Scholar] [CrossRef]
- Ma, K. Measurement of biotic community diversity: I .α diversity (Part 1). Biodivers. Sci. 1994, 2, 162–168. [Google Scholar] [CrossRef] [Green Version]
- Petraitis, P.S.; Latham, R.E.; Niesenbaum, R.A. The Maintenance of Species Diversity by Disturbance. Rev. Lit. Arts Am. 2012, 64, 393–418. [Google Scholar] [CrossRef]
- Chen, S.L.; Zhao, M.; Li, X.W.; Fan, C.; Xiao, B.R. Study on plant diversity and niche characteristics of dominant herbaceous populations under different reconstruction patterns in low efficiency stands of Cupressus Funebris. Shengtai Xuebao/Acta Ecol. Sin. 2018, 38, 143–155. [Google Scholar] [CrossRef]
- Liu, J.; Hong, W. A Study on the Community Ecology of Castanopsis kawakamii—Study on the Niche of the Main Tree Population in Castanopsis kawakamii Community. Acta Ecol. Sin. 1999, 19, 347–352. [Google Scholar]
- Huston, M.; The, S.; Naturalist, A.; Jan, N. The University of Chicago A General Hypothesis of Species Diversity. Am. Nat. 1979, 113, 81–101. [Google Scholar] [CrossRef]
- Strathmann, R.R. Environmental Heterogeneity and Plant Species Diversity: A Hypothesis. Am. Nat. 2012, 111, 376–381. [Google Scholar]
- Chen, H.Y.H.; Légaré, S.; Bergeron, Y. Variation of the understory composition and diversity along a gradient of productivity in Populus tremuloides stands of northern British Columbia, Canada. Can. J. Bot. 2004, 82, 1314–1323. [Google Scholar] [CrossRef]
- Stevens, M.H.H.; Carson, W.P. Resource quantity, not resource heterogeneity, maintains plant diversity. Ecol. Lett. 2002, 5, 420–426. [Google Scholar] [CrossRef]
- Srivastava, D.S.; Lawton, J.H. Why more productive sites have more species: An experimental test of theory using tree-hole communities. Am. Nat. 1998, 152, 510–529. [Google Scholar] [CrossRef]
- Storch, D.; Bohdalková, E.; Okie, J. The more-individuals hypothesis revisited: The role of community abundance in species richness regulation and the productivity–diversity relationship. Ecol. Lett. 2018, 21, 920–937. [Google Scholar] [CrossRef] [PubMed]
- Sercu, B.K.; Baeten, L.; van Coillie, F.; Martel, A.; Lens, L.; Verheyen, K.; Bonte, D. How tree species identity and diversity affect light transmittance to the understory in mature temperate forests. Ecol. Evol. 2017, 7, 10861–10870. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Chen, H.Y.H.; Thomas, S.C.; Shahi, C. Linking resource availability and heterogeneity to understorey species diversity through succession in boreal forest of Canada. J. Ecol. 2018, 106, 1266–1276. [Google Scholar] [CrossRef]
- Jordan, S.E.; Palmquist, K.A.; Bradford, J.B.; Lauenroth, W.K. Soil water availability shapes species richness in mid-latitude shrub steppe plant communities. J. Veg. Sci. 2020, 31, 646–657. [Google Scholar] [CrossRef]
- Van der Putten, W.H.; Bardgett, R.D.; Bever, J.D.; Bezemer, T.M.; Casper, B.B.; Fukami, T.; Kardol, P.; Klironomos, J.N.; Kulmatiski, A.; Schweitzer, J.A.; et al. Plant-soil feedbacks: The past, the present and future challenges. J. Ecol. 2013, 101, 265–276. [Google Scholar] [CrossRef]
- Tinya, F.; Ódor, P. Congruence of the spatial pattern of light and understory vegetation in an old-growth, temperate mixed forest. For. Ecol. Manag. 2016, 381, 84–92. [Google Scholar] [CrossRef] [Green Version]
- Royer-Tardif, S.; Bradley, R.L.; Parsons, W.F.J. Evidence that plant diversity and site productivity confer stability to forest floor microbial biomass. Soil Biol. Biochem. 2010, 42, 813–821. [Google Scholar] [CrossRef]
- Williams, L.J.; Paquette, A.; Cavender-Bares, J.; Messier, C.; Reich, P.B. Spatial complementarity in tree crowns explains overyielding in species mixtures. Nat. Ecol. Evol. 2017, 1, 1–7. [Google Scholar] [CrossRef]
- Ampoorter, E.; Baeten, L.; Koricheva, J.; Vanhellemont, M.; Verheyen, K. Do diverse overstoreys induce diverse understoreys? Lessons learnt from an experimental-observational platform in Finland. For. Ecol. Manag. 2014, 318, 206–215. [Google Scholar] [CrossRef]
- Fort, H. Community diversity and total abundance: Quantitative predictions from competition niche theory. Ecol. Complex. 2015, 21, 120–127. [Google Scholar] [CrossRef]
- Gauze, G.F.; Georgi, F. The Struggle for Existence; Williams & Wilkins: Baltimore, MD, USA, 1934. [Google Scholar]
- Wittman, S.E.; Sanders, N.J.; Ellison, A.M.; Jules, E.S.; Ratchford, J.S.; Gotelli, N.J. Species interactions and thermal constraints on ant community structure. Oikos 2010, 119, 551–559. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Pattern | Main Arbor Species | Average Height/m | Average DBH/cm | Canopy Density | Slope/° | Aspect |
|---|---|---|---|---|---|---|
| Ac-H | A. cremastogyne | 5.3 | 3.5 | 0.7 | 25° | NE68° |
| Ts-H | T. sinensis | 4.1 | 4.3 | 0.6 | 26° | NE68° |
| Cs-H | C. septentrionale | 4.2 | 3.4 | 0.6 | 27° | NE68° |
| TC | T. sinensis | 4.4 | 3.7 | 0.7 | 27° | NE68° |
| C. septentrionale | 5.3 | 4.5 | ||||
| CA | C. septentrionale | 5.7 | 4.7 | 0.7 | 26° | NE68° |
| A. cremastogyne | 5.4 | 5.0 | ||||
| TA | A. cremastogyne | 6.1 | 4.9 | 0.7 | 24° | NE68° |
| T. sinensis | 3.8 | 3.6 | ||||
| ATC | A. cremastogyne | 5.3 | 4.5 | 0.7 | 23° | SE15° |
| T. sinensis | 4.8 | 4 | ||||
| C. septentrionale | 4.7 | 3.8 | ||||
| Ac-L | A. cremastogyne | 6 | 5.3 | 0.6 | 21° | SE24° |
| Ts-L | T. sinensis | 4.5 | 3.7 | 0.5 | 24° | SE22° |
| Cs-L | C. septentrionale | 4.4 | 4.2 | 0.5 | 22° | SE28° |
| CB | cutting-blank | 4.9 | 4.2 | 0.5 | 25° | NE68° |
| Cf | Cupressus funebris | 9.2 | 8.8 | 0.8 | 24° | E |
| Species | Niche Breadth | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Total | Pattern | ||||||||||||
| Ac-H | Ts-H | Cs-H | TC | CA | TA | ATC | Ac-L | Ts-L | Cs-L | CB | Cf | ||
| Alangium chinense | 2.057 | 1.075 | 1.087 | 0.693 | 0.691 | 0.690 | |||||||
| Cupressus funebris | 1.839 | 1.087 | 1.090 | 1.048 | 0.614 | 0.671 | |||||||
| Rubus coreanus | 1.886 | 0.681 | 1.074 | 0.645 | 0.927 | 0.958 | 0.693 | 0.690 | |||||
| Broussonetia papyrifera | 2.371 | 1.337 | 1.373 | 1.373 | 1.090 | 1.030 | 1.035 | 1.351 | 1.336 | 1.383 | 1.369 | 1.371 | |
| Toona ciliata | 1.075 | ||||||||||||
| Elaeagnus pungens | 1.805 | 1.371 | 1.051 | 1.024 | 0.660 | ||||||||
| Zanthoxylum bungeanum | 2.092 | 1.098 | 0.583 | 0.693 | 0.691 | 1.088 | 1.349 | 1.330 | 1.085 | ||||
| Vitex negundo | 2.191 | 0.647 | 1.065 | 1.365 | 1.003 | 1.070 | 1.340 | 1.374 | 1.306 | 1.092 | 1.084 | ||
| Pyracantha fortuneana | 2.027 | 1.045 | 0.581 | 0.621 | 1.350 | 0.675 | 0.631 | 0.688 | 1.098 | ||||
| Coriaria nepalensis | 1.537 | 0.686 | |||||||||||
| Ligustrum lucidum | 1.751 | 1.368 | 0.636 | 1.080 | 0.675 | ||||||||
| Rhamnus davurica | 1.591 | ||||||||||||
| Myrsine africana | 2.028 | 0.665 | 1.343 | 1.095 | 0.660 | 0.692 | 0.655 | 0.681 | 0.693 | 1.357 | |||
| Sapium sebiferum | 1.560 | 0.689 | 0.685 | ||||||||||
| Rhus chinensis | 2.296 | 0.676 | 1.286 | 1.359 | 1.008 | 1.381 | 1.101 | 1.024 | 1.093 | 1.379 | 1.336 | 1.098 | |
| Vernicia fordii | 1.771 | 0.652 | 1.351 | 0.658 | 0.684 | 0.684 | |||||||
| Species | Importance Values | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Total | Pattern | ||||||||||||
| Ac-H | Ts-H | Cs-H | TC | CA | TA | ATC | AcⅡ | TsⅡ | CsⅡ | CB | Cf | ||
| Alangium chinense | 4.856 | 1.512 | 1.497 | 2.954 | 5.712 | 1.582 | 1.888 | 11.490 | 6.478 | 2.762 | 10.388 | 17.633 | |
| Cupressus funebris | 3.264 | 7.823 | 3.263 | 1.989 | 1.308 | 7.819 | 9.591 | 3.377 | 15.016 | ||||
| Rubus coreanus | 5.151 | 6.324 | 8.499 | 6.602 | 1.578 | 21.699 | 14.215 | 2.766 | 2.960 | 2.515 | |||
| Broussonetia papyrifera | 18.160 | 23.396 | 10.650 | 20.970 | 11.953 | 10.909 | 9.422 | 22.803 | 14.668 | 26.653 | 32.873 | 31.286 | 5.729 |
| Toona ciliata | 0.827 | 1.497 | 2.491 | 1.768 | |||||||||
| Elaeagnus pungens | 2.891 | 8.100 | 8.494 | 1.798 | 2.727 | 5.012 | 4.117 | 2.462 | |||||
| Zanthoxylum bungeanum | 7.397 | 8.196 | 4.961 | 3.333 | 3.948 | 1.582 | 7.543 | 12.928 | 20.633 | 13.823 | 6.514 | ||
| Vitex negundo | 11.878 | 4.749 | 7.394 | 28.201 | 13.467 | 13.774 | 21.755 | 14.121 | 20.297 | 11.352 | 27.113 | ||
| Pyracantha fortuneana | 5.730 | 3.110 | 6.478 | 5.533 | 6.840 | 20.598 | 3.525 | 6.140 | 7.555 | 12.201 | |||
| Coriaria nepalensis | 1.507 | 1.370 | 3.489 | 3.529 | 1.517 | 2.484 | |||||||
| Ligustrum lucidum | 2.522 | 8.668 | 3.151 | 8.223 | 1.494 | 4.446 | 2.718 | 1.651 | |||||
| Rhamnus davurica | 1.400 | 1.646 | 1.795 | 2.409 | 2.491 | 1.610 | |||||||
| Myrsine africana | 5.460 | 7.813 | 16.688 | 8.242 | 3.793 | 4.293 | 2.567 | 4.429 | 5.734 | 7.388 | 27.993 | ||
| Sapium sebiferum | 1.320 | 3.138 | 1.757 | 2.703 | 1.200 | 2.411 | |||||||
| Rhus chinensis | 17.036 | 4.728 | 19.000 | 28.872 | 14.052 | 11.543 | 8.248 | 20.247 | 16.611 | 28.000 | 27.833 | 16.092 | |
| Vernicia fordii | 2.736 | 7.890 | 6.941 | 3.973 | 1.865 | 1.410 | 2.933 | 8.776 | |||||
| Patterns | Dominant Shrub | Importance Value | Proportion of Total Importance Value (%) | Niche Overlap | Average Niche Overlap |
|---|---|---|---|---|---|
| Cf | Myrsine africana | 27.993 | 72.739 | 0.592 | 0.7103 ± 0.1066 |
| Vitex negundo | 27.113 | 0.799 | |||
| Alangium chinense | 17.633 | 0.74 | |||
| Ts-H | Myrsine africana | 16.688 | 35.837 | 0.469 | 0.6293 ± 0.2505 |
| Broussonetia papyrifera | 10.65 | 0.918 | |||
| Rubus coreanus | 8.499 | 0.501 | |||
| Cs-H | Rhus chinensis | 28.872 | 58.084 | 0.918 | 0.6293 ± 0.2505 |
| Broussonetia papyrifera | 20.97 | 0.469 | |||
| Myrsine africana | 8.242 | 0.501 | |||
| Ac-H | Broussonetia papyrifera | 23.396 | 40.26 | 0.561 | 0.5443 ± 0.2444 |
| Ligustrum lucidum | 8.668 | 0.78 | |||
| Zanthoxylum bungeanum | 8.196 | 0.292 | |||
| Ts-L | Rhus chinensis | 28 | 75.286 | 0.918 | 0.8230 ± 0.0824 |
| Broussonetia papyrifera | 26.653 | 0.771 | |||
| Zanthoxylum bungeanum | 20.633 | 0.78 | |||
| Cs-L | Broussonetia papyrifera | 32.873 | 81.003 | 0.981 | 0.7713 ± 0.1834 |
| Rhus chinensis | 27.833 | 0.692 | |||
| Vitex negundo | 20.297 | 0.641 | |||
| Ac-L | Rhus chinensis | 16.611 | 45.4 | 0.918 | 0.7797 ± 0.1385 |
| Broussonetia papyrifera | 14.668 | 0.641 | |||
| Vitex negundo | 14.121 | 0.78 | |||
| TA | Pyracantha fortuneana | 20.598 | 48.587 | 0.702 | 0.5597 ± 0.1294 |
| Rubus coreanus | 14.215 | 0.528 | |||
| Vitex negundo | 13.774 | 0.449 | |||
| TC | Vitex negundo | 28.201 | 54.206 | 0.641 | 0.7503 ± 0.1474 |
| Rhus chinensis | 14.052 | 0.692 | |||
| Broussonetia papyrifera | 11.953 | 0.918 | |||
| CA | Rubus coreanus | 21.699 | 46.709 | 0.449 | 0.5263 ± 0.1013 |
| Vitex negundo | 13.467 | 0.489 | |||
| Rhus chinensis | 11.543 | 0.641 | |||
| ATC | Broussonetia papyrifera | 22.803 | 64.805 | 0.692 | 0.7503 ± 0.1474 |
| Vitex negundo | 21.755 | 0.918 | |||
| Rhus chinensis | 20.247 | 0.641 | |||
| CB | Broussonetia papyrifera | 31.286 | 59.579 | 0.918 | 0.7010 ± 0.1889 |
| Rhus chinensis | 16.092 | 0.612 | |||
| Pyracantha fortuneana | 12.201 | 0.573 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Chen, S.; He, W.; Ren, J.; Wen, X.; Wang, Y.; Li, X.; Chen, G.; Feng, M.; Fan, C. Shrub Diversity and Niche Characteristics in the Initial Stage of Reconstruction of Low-Efficiency Cupressus funebris Stands. Forests 2021, 12, 1492. https://doi.org/10.3390/f12111492
Wang Y, Chen S, He W, Ren J, Wen X, Wang Y, Li X, Chen G, Feng M, Fan C. Shrub Diversity and Niche Characteristics in the Initial Stage of Reconstruction of Low-Efficiency Cupressus funebris Stands. Forests. 2021; 12(11):1492. https://doi.org/10.3390/f12111492
Chicago/Turabian StyleWang, Yang, Silu Chen, Wenchun He, Jingjing Ren, Xiaochen Wen, Yu Wang, Xianwei Li, Gang Chen, Maosong Feng, and Chuan Fan. 2021. "Shrub Diversity and Niche Characteristics in the Initial Stage of Reconstruction of Low-Efficiency Cupressus funebris Stands" Forests 12, no. 11: 1492. https://doi.org/10.3390/f12111492