Effects of Twenty Years of Ungulate Browsing on Forest Regeneration at Paneveggio Reserve, Italy

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Characteristics
2.2. Experimental Design and Data Collection
2.3. Statistical Analyses
3. Results
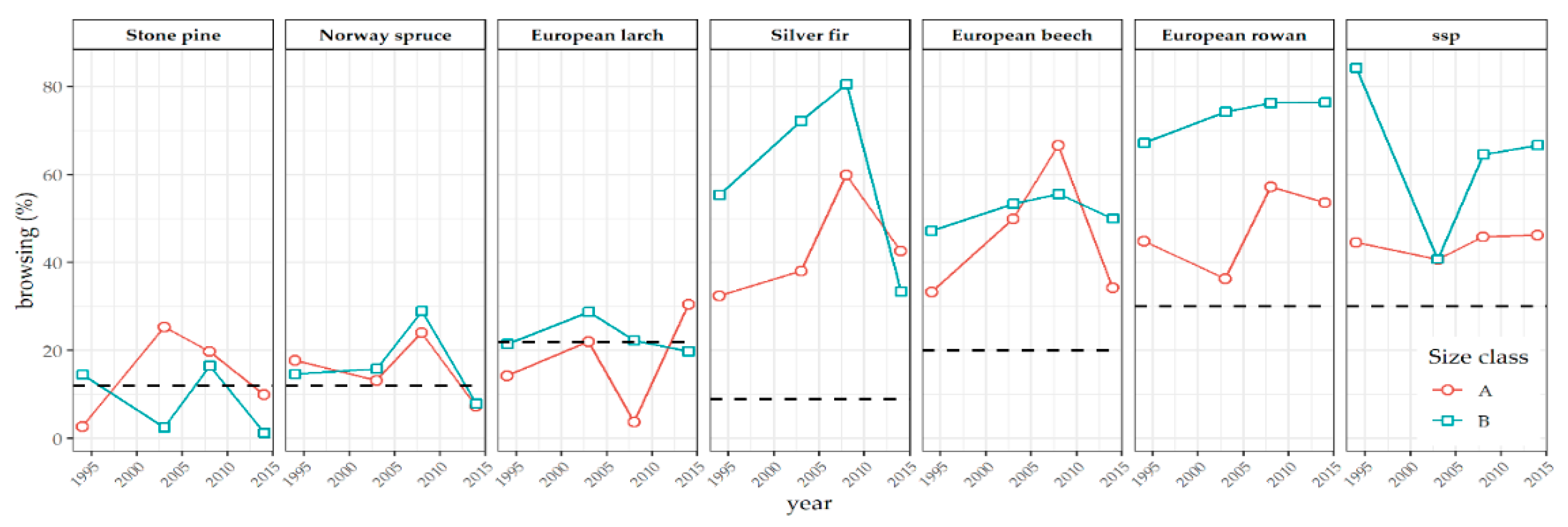
3.1. Browsing Damage
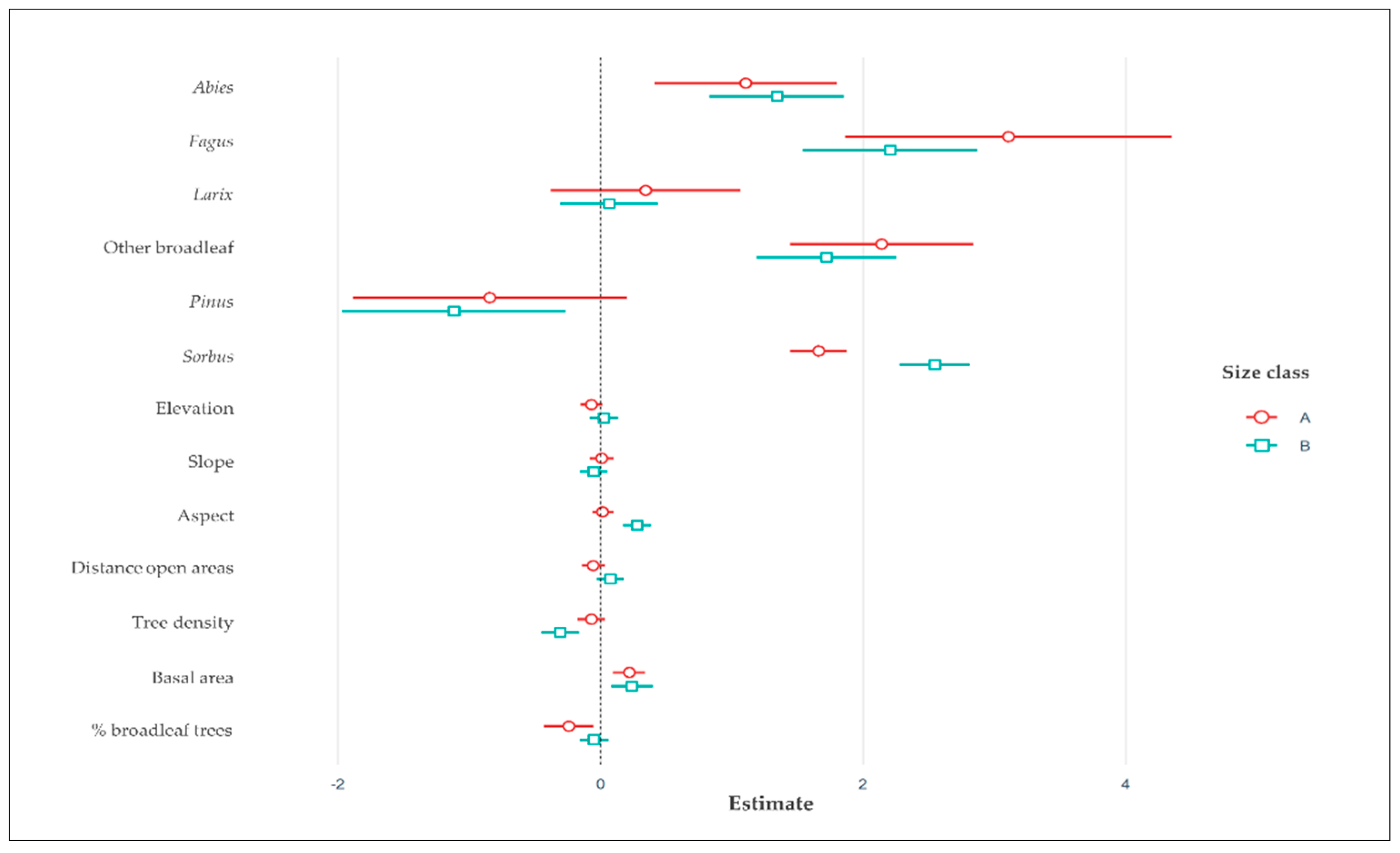
3.2. Drivers of Browsing Activity
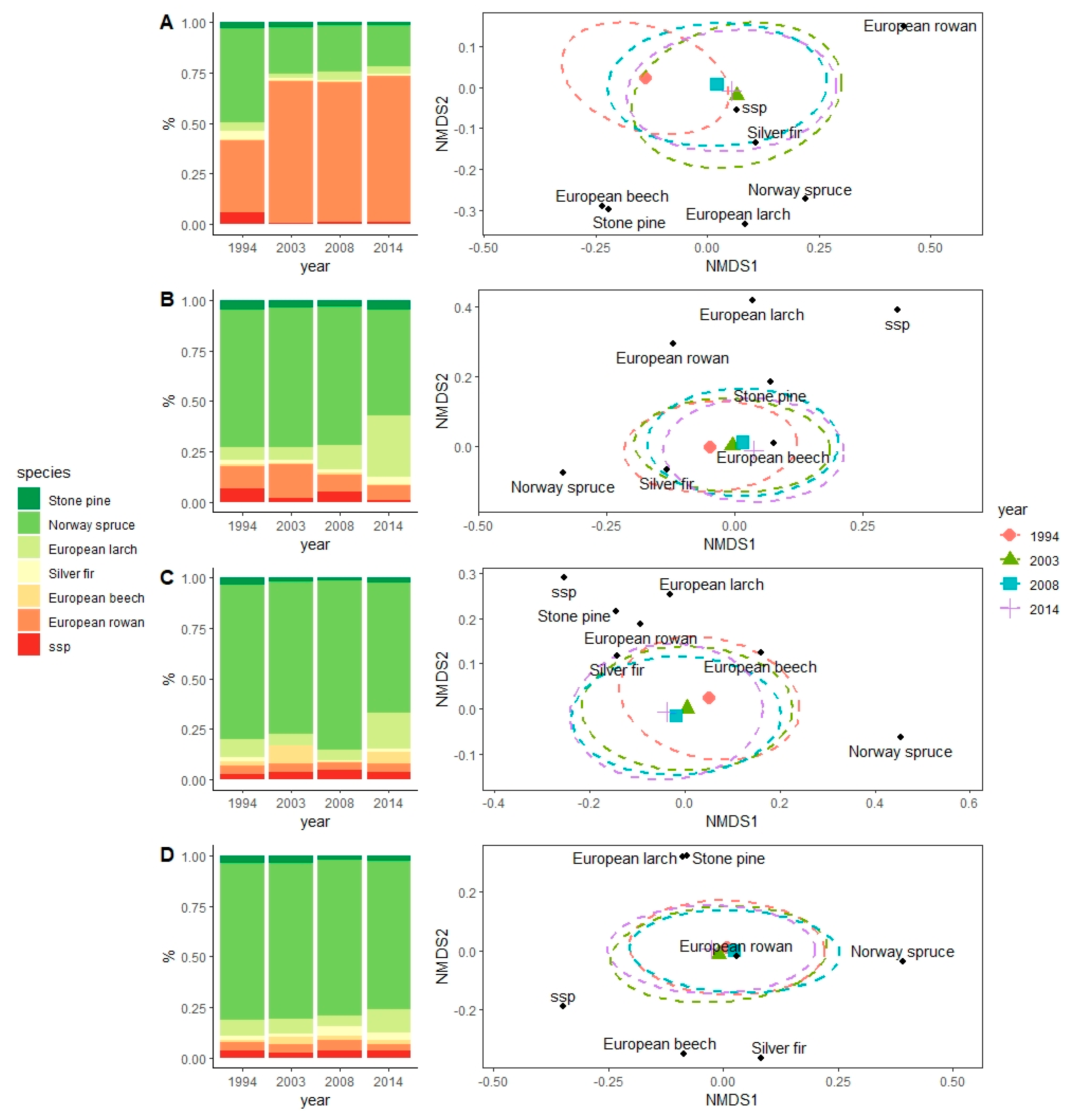
3.3. Effects on Specific Composition
4. Discussion
4.1. Damage Analysis
4.2. Drivers of Browsing Activity
4.3. Effects on Specific Composition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Côté, S.D.; Rooney, T.P.; Tremblay, J.P.; Dussault, C.; Waller, D.M. Ecological Impacts of Deer Overabundance. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 113–147. [Google Scholar] [CrossRef] [Green Version]
- Horsley Stephen, B.; Stout Susan, L.; De Calesta, D.S. White-tailed deer impact on the vegetation dynamics of a Northern Hardwood forest. Ecol. Appl. 2003, 13, 98–118. [Google Scholar] [CrossRef]
- Ripple, W.J.; Beschta, R.L. Trophic cascades involving cougar, mule deer, and black oaks in Yosemite National Park. Biol. Conserv. 2008, 141, 1249–1256. [Google Scholar] [CrossRef]
- Hansen, B.B.; Henriksen, S.; Aanes, R.; Sæther, B.E. Ungulate impact on vegetation in a two-level trophic system. Polar Biol. 2007, 30, 549–558. [Google Scholar] [CrossRef]
- Takatsuki, S. Effects of sika deer on vegetation in Japan: A review. Biol Conserv. 2009, 142, 1922–1929. [Google Scholar] [CrossRef]
- Husheer, S.W.; Coomes, D.A.; Robertson, A.W. Long-term influences of introduced deer on the composition and structure of New Zealand Nothofagus forests. For. Ecol. Manag. 2003, 181, 99–117. [Google Scholar] [CrossRef]
- Vacek, Z. Structure and dynamics of spruce-beech-fir forests in Nature Reserves of the Orlické hory Mts. in relation to ungulate game. Cent. Eur. For. J. 2017, 63, 22–34. [Google Scholar] [CrossRef] [Green Version]
- Motta, R. Impact of wild ungulates on forest regeneration and tree composition of mountain forests in the Western Italian Alps. For. Ecol. Manag. 1996, 88, 93–98. [Google Scholar] [CrossRef]
- Witt, J.C.; Webster, C.R. Regeneration dynamics in remnant Tsuga canadensis stands in the northern Lake States: Potential direct and indirect effects of herbivory. For. Ecol. Manag. 2010, 260, 519–525. [Google Scholar] [CrossRef]
- Hidding, B.; Tremblay, J.P.; Côté, S.D. Survival and growth of balsam fir seedlings and saplings under multiple controlled ungulate densities. For. Ecol. Manag. 2012, 276, 96–103. [Google Scholar] [CrossRef]
- Rooney, T.P.; Waller, D.M. Direct and indirect effects of white-tailed deer in forest ecosystems. For. Ecol. Manag. 2003, 181, 165–176. [Google Scholar] [CrossRef]
- Bernard, M.; Boulanger, V.; Dupouey, J.L.; Laurent, L.; Montpied, P.; Morin, X.; Picard, J.F.; Saïd, S. Deer browsing promotes Norway spruce at the expense of silver fir in the forest regeneration phase. For. Ecol. Manag. 2017, 400, 269–277. [Google Scholar] [CrossRef]
- Churski, M.; Bubnicki, J.W.; Jędrzejewska, B.; Kuijper, D.P.J.; Cromsigt, J.P.G.M. Brown world forests: Increased ungulate browsing keeps temperate trees in recruitment bottlenecks in resource hotspots. New Phytol. 2017, 214, 158–168. [Google Scholar] [CrossRef] [Green Version]
- Motta, R. Ungulate impact on rowan (Sorbus aucuparia L.) and Norway spruce (Picea abies (L.) Karst.) height structure in mountain forests in the eastern Italian Alps. For. Ecol. Manag. 2003, 181, 139–150. [Google Scholar] [CrossRef]
- Didion, M.; Kupferschmid, A.D.; Bugmann, H. Long-term effects of ungulate browsing on forest composition and structure. For. Ecol. Manag. 2009, 258, 44–55. [Google Scholar] [CrossRef]
- Eiberle, K.; Nigg, H. Grundlagen zur Beurteilung des Wildverbisses im Gebirgswald. Schweiz. Z. Forstwes. 1987, 138, 747–785. [Google Scholar]
- Mencuccini, M.; Piussi, P.; Zanzi Sulli, A. Thirty years of seed production in a subalpine Norway spruce forest: Patterns of temporal and spatial variation. For. Ecol. Manag. 1995, 76, 109–125. [Google Scholar] [CrossRef]
- Brüllhardt, M.; Risch, A.C.; Filli, F.; Haller, R.M.; Schütz, M. Spatiotemporal dynamics of natural tree regeneration in unmanaged subalpine conifer forests with high wild ungulate densities. Can. J. For. Res. 2015, 45, 607–614. [Google Scholar] [CrossRef]
- Loison, A.; Toïgo, C.; Gaillard, J.M. Large Herbivores in European Alpine Ecosystems: Current Status and Challenges for the Future. In Alpine Biodiversity in Europe. Ecological Studies (Analysis and Synthesis); Nagy, L., Grabherr, G., Körner, C., Thompson, D.B.A., Eds.; Springer: Berlin, Germany, 2003; Volume 167. [Google Scholar]
- Motta, R.; Franzoi, M. Foreste di montagna ed ungulati selvatici nel Parco naturale di Paneveggio–Pale di S. Martino (TN). Dendronatura 1997, 18, 15–32. [Google Scholar]
- Apollonio, M.; Chirichella, R.; De Marinis, A.M.; Bazzanella, G.; Brugnoli, A.; Ferraro, E.; Giovannini, R.; Luchesa, L.; Rocca, M.; Zambotti, S. Camoscio, cervo e capriolo in Trentino. Rapporto su status e gestione. Quad. Assoc. Cacciatori Trentini 2019, 1, 248. [Google Scholar]
- Associazione Cacciatori Trentini. Relazioni Annuali Cervo (Cervus Elaphus Hippelaphus, L. 1758), Distretti Faunistici di Fassa, Fiemme e Primiero; Relazioni interne delle Consulte di Fassa: Fiemme e Primiero, Italy, 2019. [Google Scholar]
- Turchetto, S.; Obber, F.; Permunian, R.; Vendrami, S.; Lorenzetto, M.; Ferré, N.; Stancampiano, L.; Rossi, L.; Citterio, C.V. Spatial and temporal explorative analysis of sarcoptic mange in Alpine chamois (Rupicapra rupicapra). Hystrix, It. J. Mamm 2014, 25. [Google Scholar] [CrossRef]
- Lovari, S.; Ferretti, F.; Corazza, M.; Minder, I.; Troiani, N.; Ferrari, C.; Saddi, A. Unexpected consequences of reintroductions: Competition between reintroduced red deer and Apennine chamois. Anim. Conserv. 2014, 17, 359–370. [Google Scholar] [CrossRef]
- Richard, E.; Gaillard, J.M.; Saïd, S.; Hamann, J.L.; Klein, F. High red deer density depresses body mass of roe deer fawns. Oecologia 2010, 163, 91–97. [Google Scholar] [CrossRef]
- Hair Jr, J.F.; Anderson, R.E.; Tatham, R.L.; Black, W.C. Multivariate Data Analysis, 5th ed.; Prentice-Hall: Upper Saddle River, NJ, USA, 1998. [Google Scholar]
- Coulombe, M.L.; Huot, J.; Massé, A.; Côté, S.D. Influence of forage biomass and cover on deer space use at a fine scale: A controlled-density experiment. Ecoscience 2011, 18, 262–272. [Google Scholar] [CrossRef]
- Anderwald, P.; Haller, R.M.; Filli, F. Heterogeneity in primary productivity influences competitive interactions between Red deer and Alpine chamois. PLoS ONE 2016, 11, e0146458. [Google Scholar] [CrossRef] [Green Version]
- Giroux, M.-A.; Dussault, C.; Tremblay, J.P.; Côté, S.D. Winter severity modulates the benefits of using a habitat temporally uncoupled from browsing. Ecosphere 2016, 7, e01432. [Google Scholar] [CrossRef]
- Provincia Autonoma di Trento. Quaderno di Nivologia n°29, Stagione Invernale 2011-’12; Pubblicazione interna a cura dell’Ufficio Previsioni e Pianificazione, Servizio Prevenzione Rischi; Dipartimento Protezione Civile: Trento (TN), Italy, 2015.
- Renaud, P.; Verheyden-Tixier, H.; Dumont, B. Damage to saplings by red deer (Cervus elaphus): Effect of foliage height and structure. For. Ecol. Manag. 2003, 181, 31–37. [Google Scholar] [CrossRef]
- Kamler, J.; Homolka, M. Needles in faeces: An index of quality of wild ungulate winter diet. Folia Zool. 2011, 60, 63–69. [Google Scholar] [CrossRef]
- Stewart, K.M.; Bowyer, R.T.; Dick, B.L.; Kie, J.G. Effects of density dependence on diet composition of North American elk Cervus elaphus and mule deer Odocoileus hemionus: An experimental manipulation. Wildlife Biol. 2011, 17, 417–430. [Google Scholar] [CrossRef] [Green Version]
- Barbier, S.; Gosselin, F.; Belandier, P. Influence of tree species on understory vegetation diversity and mechanisms involved—A critical review for temperate and boreal forests. For. Ecol. Manag. 2008, 254, 1–15. [Google Scholar] [CrossRef]
- Herfindal, I.; Tremblay, J.-P.; Hester, A.J.; Lande, U.S.; Wam, H.K. Associational relationships at multiple spatial scales affect forest damage by moose. For. Ecol. Manag. 2015, 348, 97–107. [Google Scholar] [CrossRef] [Green Version]
- Dumont, A.; Ouellet, J.P.; Crête, M.; Huot, J. Winter foraging strategy of white-tailed deer at the northern limit of its range. Écoscience 2005, 12, 476–484. [Google Scholar] [CrossRef]
- Anderson, B.T.; McNamara, J.P.; Marshall, H.; Flores, A.N. Insights into the physical processes controlling correlations between snow distribution and terrain properties. Water Resour. Res. 2014, 50, 4545–4563. [Google Scholar] [CrossRef]
- Gilbert, S.L.; Hundertmark, K.J.; Person, D.K.; Lindberg, M.S.; Boyce, M.S. Behavioral plasticity in a variable environment: Snow depth and habitat interactions drive deer movement in winter. J. Mammal. 2017, 98, 246–259. [Google Scholar] [CrossRef] [Green Version]
- Parker, K.L.; Robbins, C.T.; Hanley, T.A. Energy expenditures for locomotion by mule deer and elk. J. Wildl. Manage. 1984, 48, 474–488. [Google Scholar] [CrossRef]
- Ceacero, F.; Garcia, A.J.; Landete-Castillejos, T.; Bartošová, J.; Bartoš, L.; Gallego, L. Benefits for Dominant Red Deer Hinds under a Competitive Feeding System: Food Access Behavior, Diet and Nutrient Selection. PLoS ONE 2012, 7, e32780. [Google Scholar] [CrossRef] [Green Version]
- Lochhead, K.D.; Comeau, P.G. Relationships between forest structure, understorey light and regeneration in complex Douglas-fir dominated stands in south-eastern British Columbia. For. Ecol. Manag. 2012, 284, 12–22. [Google Scholar] [CrossRef]
- Häsler, H.; Senn, J. Ungulate browsing on European silver fir Abies alba: The role of occasions, food shortage and diet preferences. Wildlife Biol. 2012, 18, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Beier, P.; McCullough, D.R. Factors influencing white-tailed deer activity patterns and habitat use. Wildl. Monogr. 1990, 109, 5–51. [Google Scholar]
- Webb, S.L.; Gee, K.L.; Strickland, B.K.; Demarais, S.; De Young, R.W. Measuring fine-scale white-tailed deer movements and environmental influences using GPS collars. Int. J. Ecol. 2010, 2010, 459610. [Google Scholar] [CrossRef] [Green Version]
- Bonnot, N.; Morellet, N.; Verheyden, H.; Cargnelutti, B.; Lourtet, B.; Klein, F.; Hewison, A.M. Habitat use under predation risk: Hunting, roads and human dwellings influence the spatial behaviour of roe deer. Eur. J. Wildl. Res. 2013, 59, 185–193. [Google Scholar] [CrossRef]
- Williams, D.M.; Quinn, A.C.D.; Porter, W.F. Landscape effects on scales of movement by white-tailed deer in an agricultural–forest matrix. Landsc. Ecol. 2012, 27, 45–57. [Google Scholar] [CrossRef]
- Jara-Guerrero, A.; Escribano-Avila, G.; Espinosa, C.I.; De la Cruz, M.; Méndez, M. White-tailed deer as the last megafauna dispersing seeds in Neotropical dry forests: The role of fruit and seed traits. Biotropica 2018, 50, 169–177. [Google Scholar] [CrossRef]
- Albert, A.; Auffret, A.G.; Cosyns, E.; Cousins, S.A.O.; D’hondt, B.; Eichberg, C.; Eycott, A.E.; Heinken, T.; Hoffmann, M.; Jaroszewicz, B.; et al. Seed dispersal by ungulates as an ecological filter: A trait-based meta-analysis. Oikos 2015, 124, 1109–1120. [Google Scholar] [CrossRef]
- Simončič, T.; Bončina, A.; Jarni, K.; Klopčič, M. Assessment of the long-term impact of deer on understory vegetation in mixed temperate forests. J. Veg. Sci. 2019, 30, 108–120. [Google Scholar] [CrossRef] [Green Version]
- Lecomte, X.; Fedriani, J.M.; Caldeira, M.C.; Clemente, A.S.; Olmi, A.; Bugalho, M.N. Too Many Is Too Bad: Long-Term Net Negative Effects of High Density Ungulate Populations on a Dominant Mediterranean Shrub. PLoS One 2016, 11, e0158139. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Variable | Classification | 1994 | 2003 | 2008 | 2014 |
|---|---|---|---|---|---|
| % Damaged Trees | browsing | 11 (20) | 16 (27) | 24 (31) | 16 (26) |
| fraying | 4 (10) | 2 (9) | 3 (10) | 4 (12) | |
| bark stripping | 0 | 1 (4) | 1 (5) | 3 (11) | |
| % Browsed Trees | A | 22 (28) | 27 (28) | 42 (33) | 36 (29) |
| B | 22 (23) | 28 (33) | 36 (34) | 21 (28) | |
| C | 3 (9) | 6 (17) | 13 (27) | 4 (14) | |
| D | 0 (0) | 1 (4) | 2 (3) | 2 (11) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Aprile, D.; Vacchiano, G.; Meloni, F.; Garbarino, M.; Motta, R.; Ducoli, V.; Partel, P. Effects of Twenty Years of Ungulate Browsing on Forest Regeneration at Paneveggio Reserve, Italy. Forests 2020, 11, 612. https://doi.org/10.3390/f11060612
D’Aprile D, Vacchiano G, Meloni F, Garbarino M, Motta R, Ducoli V, Partel P. Effects of Twenty Years of Ungulate Browsing on Forest Regeneration at Paneveggio Reserve, Italy. Forests. 2020; 11(6):612. https://doi.org/10.3390/f11060612
Chicago/Turabian StyleD’Aprile, Davide, Giorgio Vacchiano, Fabio Meloni, Matteo Garbarino, Renzo Motta, Vittorio Ducoli, and Piergiovanni Partel. 2020. "Effects of Twenty Years of Ungulate Browsing on Forest Regeneration at Paneveggio Reserve, Italy" Forests 11, no. 6: 612. https://doi.org/10.3390/f11060612