Intercropping the Sharp-Leaf Galangal with the Rubber Tree Exhibits Weak Belowground Competition

1
CAS Key Laboratory of Tropical Forest Ecology, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Menglun, Yunnan 666303, China
2
Center of Plant Ecology, Core Botanical Gardens, Chinese Academy of Sciences, Menglun, Yunnan 666303, China
3
University of Chinese Academy of Sciences, Beijing 100049, China
*
Author to whom correspondence should be addressed.
Forests 2019, 10(10), 924; https://doi.org/10.3390/f10100924
Submission received: 3 September 2019
/
Revised: 12 October 2019
/
Accepted: 18 October 2019
/
Published: 20 October 2019
(This article belongs to the Special Issue Agroforestry Systems: The Role of Trees in Ecosystem Services—A Special Issue in Collaboration with the 4th World Congress on Agroforestry)
Abstract
:Intercropping the sharp-leaf galangal with the rubber tree could help to improve the sustainability of the rubber tree planting industry. However, our understanding of belowground competition in such agroforestry systems is still limited. Therefore, we used stable isotope methods (i.e., water δ2H and δ18O and leaf δ13C) to investigate plant water-absorbing patterns and water use efficiency (WUE) in a monocultural rubber plantation and in an agroforestry system of rubber trees and sharp-leaf galangal. We also measured leaf carbon (C), nitrogen (N), and phosphorus (P) to evaluate the belowground competition effects on plant nutrient absorption status. Through a Bayesian mixing model, we found that the monocultural rubber trees and the intercropped sharp-leaf galangal absorbed much more surface soil water at a depth of 0–5 cm, while the rubber trees in the agroforestry system absorbed more water from the shallow and middle soil layers at a depth of 5–30 cm. This phenomenon verified the occurrence of plant hydrologic niche segregation, whereas the WUE of rubber trees in this agroforestry system suggested that the competition for water was weak. In addition, the negative correlation between the leaf P concentration of the rubber trees and that of the sharp-leaf galangal demonstrated their competition for soil P resources, but this competition had no obvious effects on the leaf nutrient status of the rubber trees. Therefore, this study verified that the belowground competition between rubber trees and sharp-leaf galangal is weak, and this weak competition may benefit their long-term intercropping.
1. Introduction
Natural rubber is an indispensable and essential raw material for a variety of industrial applications and products [1], and it has brought huge economic benefits to the cultivated regions of the rubber tree (Hevea brasiliensis (Willd. ex A. Juss.) Müll. Arg.), especially in mainland Southeast Asia [2]. However, negative impacts from the large-scale monocultural cultivation of rubber trees on the ecological environment has engendered an adverse reputation for the rubber tree, thus demonstrating a severe hindrance to the sustainable development of the rubber planting industry [3,4]. Apart from environmental problems, the present serious issue is the continuous low price of natural rubber [4]. Due to the direct impact of this issue on the livelihoods of rubber smallholders, rubber plantations in Xishuangbanna have a tendency to convert to other monocultural plantations, especially aging rubber plantations (i.e., with a planting history of more than 30 years) that exhibit lower rubber yields [5,6]. It is noteworthy that nearly half of rubber plantations in Xishuangbanna have grown old after decades of a rubber boom [7], and the smallholders of these aged rubber plantations have shown their willingness to plant another cash crop instead of replanting rubber trees [6]. Therefore, the future of the rubber planting industry seems pessimistic.
Seriously speaking, before a suitable substitute is found, natural rubber is still an indispensable national strategic resource, and the products of natural rubber are still part of our daily lives [1,8]. Moreover, aged rubber plantations also display lots of benefits. For example, they can store much more carbon [9,10], increase biodiversity [11], and be of great benefit to forest restoration [12]. Therefore, planning for aged rubber plantations needs more scrupulous and farsighted consideration. From a conservative perspective, it is especially necessary to extend the planting period of aged rubber plantations to acquire time for in-depth studies and final decisions. Meanwhile, corresponding countermeasures for reversing the current adverse trends of the rubber planting industry are urgently needed.
As a promising approach to achieve the green and sustainable cultivation of rubber plantations, the use of rubber-based agroforestry systems is conducive to improving the ecological environment, making full use of environmental resources, promoting the productivity of a system, and creating a favorable environment for transforming an aged rubber plantation into a secondary forest or another plantation [12,13]. In order to increase and stabilize the income of rubber smallholders, the economic value of intercropping plants is always considered in the design of rubber-based agroforestry systems, such as their medicinal value, edible value, industrial value, and ornamental value [12]. Therefore, successful intercrops should always exhibit high economic value, such as sharp-leaf galangal (Alpinia oxyphylla Miq.), an important medicinal herb that is frequently adopted in the treatment of dyspepsia, diarrhea, abdominal pain, and poor memory in East and Southeast Asia [14]. In addition, it is also an excellent ornamental plant. Because of its convenient management and maintenance, low economic cost and risk, handsome profit, and long-time attraction in the market, sharp-leaf galangal has been used as a suitable cash crop for intercropping with rubber trees [15]. Therefore, the rubber tree and sharp-leaf galangal agroforestry system (RS-AFS), which may help rubber smallholders pass through hard times of economic fluctuations in theory, has become one of the most promising rubber-based intercropping systems in the rubber-cultivating regions of China [13,16].
However, in addition to economic considerations, the suitability of this intercropping species is mainly evaluated through the adaptive capacity of low light availability and the aboveground growth characteristics of the intercropping species [12,13]. There have been few studies on the belowground competition between the rubber tree and this intercropping medicinal herb despite economic studies on such rubber-based agroforestry systems having been reported on for decades. Some studies have pointed out that competition between rubber trees and intercrops can be avoided by selecting intercrops with a differing root strategy, but this argument still needs to be verified [12]. As many lessons from intercropping history have suggested, ignoring belowground competition often leads to the failure of many agroforestry systems [16]. Therefore, before a large-scale extension of the rubber-based agroforestry system, more field studies on belowground competition are needed to verify the suitability of the species combination.
In general, soil water and various soil nutrients are the main resources plants compete for underground. Although plants have unique ways of competing for soil water and nutrients due to the different properties of these two resources, there are still many similarities in water and nutrient competition because both kinds of competition are related to root characteristics, especially the feeder root, which is commonly regarded as the main organ that absorbs water and nutrients in soil. However, due to the invisibility of root interaction, the complexity of plant resource use strategies, and the temporal and spatial heterogeneity of environmental resources [17,18], traditional methods for the investigation of plant belowground competition have had many limitations. For example, a root-digging method or other direct investigation of underground biomass is destructive for the subsequent sampling. Comparatively, the stable isotope tracer method shows many advantages, such as nondestructive sampling and precise and quantitative estimations of sources [19]. Because the processes of water absorption by plants do not discriminate between the stable isotopes of hydrogen and oxygen (i.e., 2H and 18O) in soil water [20], an analysis of the stable isotopic compositions (i.e., δ2H and δ18O) of plant xylem water and soil water could help recognize plant water use patterns belowground, thus assisting in the investigation of the distribution of plant feeder roots and plant competition for belowground resources [21,22]. In addition, nutrients are transported to plant roots mainly through three mechanisms: root interception, diffusion, and mass flow. Relatively speaking, root interception contributes less of a nutrient supply than diffusion and mass flow because it is not a major mechanism that supplies nutrients directly to roots, and diffusion and mass flow must use soil water as a medium. Meanwhile, their occurrences depend on plant water uptake and transpiration, which result in a difference in soil water potential and the nutrient concentrations of soil solutions within soil [23]. Therefore, an investigation of plant water use based on the stable isotope method is also conducive to understanding plant nutrient competition, because soil water regulates the distribution of root hairs and the mobility and availability of soil nutrients [24].
In order to investigate the belowground competition between rubber trees and the sharp-leaf galangal in an RS-AFS, we adopted stable hydrogen and oxygen isotopes to investigate plant niche differentiation in an RS-AFS. We also adopted stable carbon isotopes (i.e., plant leaf δ13C) to study water use efficiency (WUE) and the response to water stress of the rubber trees and the intercropped sharp-leaf galangal. In addition, plants can change the carbon (C), nitrogen (N), and phosphorus (P) concentrations and C/N/P ratios in their tissues through nutrient translocation and retranslocation immediately when the uptake of soil N and P is insufficient to support plant growth [23]. In order to understand the competition effects on plant leaf C/N/P stoichiometry, we also measured the leaf C, N, and P concentrations of rubber trees and intercropped sharp-leaf galangal in an RS-AFS to evaluate the competition effects on plant leaf nutrient status and related growth during the rainy and dry seasons of 2016. Then we put forward two main questions: (1) Does the intercropping of sharp-leaf galangal help to change the water use of rubber trees and improve the soil water condition of this agroforestry system? (2) Does the belowground competition between rubber trees and sharp-leaf galangal influence the nutrient absorption of the rubber trees? Due to the shallow root depth of sharp-leaf galangal within the soil [15], we hypothesized that competition between the sharp-leaf galangal and the rubber tree would not be intense enough to change the water use and nutrient absorption of rubber trees in an RS-AFS.
2. Materials and Methods
2.1. Study Site
All study sites were located in Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences (XTBG; 21°55′39″ N, 101°15′55″ E), in Xishuangbanna Prefecture, Yunnan Province, Southwestern China. The rainy season and the dry season are apparent in this region because the local climate is mainly controlled by the tropical southern monsoon of the Indian Ocean. The annual mean temperature is 21.8 °C, and the annual average precipitation is about 1550 mm. Despite the precipitation of this region being seemingly abundant, over 80% of the precipitation is concentrated in the rainy season, especially from May to August, and the dry season in this region exhibits less precipitation and a higher air temperature (the maximum air temperature in the daytime always reaches above 30 °C, especially from March to April; data were provided from the Xishuangbanna Station for Tropical Rainforest Ecosystem Studies).
The studies were performed in a rubber monoculture ((RM), as the control) and in an agroforestry system of rubber trees (H. brasiliensis) and sharp-leaf galangal (A. oxyphylla) (i.e., RS-AFS) (Figure S1). The rubber trees in all study sites were planted in 1973, they were planted in sets of double rows (rows were spaced 2 m apart, and the trees within the rows were spaced approximately 3 m apart), and each set of double rows was separated by an 18-m-wide gap. The sharp-leaf galangals in the RS-AFS were cultivated in this wide gap in 2010, and the planting pattern of this intercropped species was about 80 cm × 120 cm. After six years of intercropping, the sharp-leaf galangals grew densely and gradually covered the rubber tree planting rows through lateral extensions of their creeping stems. They had well-developed and thick rhizomes and stolons, but their rooting depths were about 25.46 ± 2.25 cm (n = 8). As a perennial herbaceous plant, sharp-leaf galangal produces clumps of leafy stems from a large creeping rhizome, and the aboveground heights of the sharp-leaf galangals were approximately 1.6 m. In addition, the heights of the rubber trees were more than 30 m during the studying period. The altitude of these two study sites was approximately 500 m above sea level, and the distances between study sites were less than 50 m. The terrain of all investigated sites was flat.
2.2. Sampling and Measuring Methods
Each study site was divided into several 6 × 9 m blocks (Figure S2). In total, there were 20 blocks in the RM and 14 blocks in the RS-AFS. On each sampling date, three blocks were selected randomly for sampling.
The leaves of the rubber trees and sharp-leaf galangals were sampled from the extremely dry season to the pronounced rainy season (i.e., 12 March 2016; 21 May 2016; 22 June 22 2016; and 20 July 2016). At noon of each sampling date, we collected the leaves from one individual rubber tree and one clump of the intercropped herbs at these three randomly selected blocks (6 × 9 m) in each study site (Figure S2). Therefore, on each sampling date, three individual rubber trees were selected for sampling in each site, and three clumps of sharp-leaf galangal were selected for sampling in the RS-AFS. We cut the shoots of rubber trees from the sunny slope of their canopy in each site and collected the leaves from these shoots. For the sharp-leaf galangals, we sampled the leaves directly. All leaf samples were dried (65 °C, 48 h) to a constant weight and then crushed with a pulverizer and sifted to fine powder with a 100-mesh sieve.
The leaf δ13C value was measured through an IsoPrime100 (Isoprime, Stockport, UK). The isotope ratios for 13C were expressed in parts per thousand relative to Vienna Pee Dee Belemnite (i.e., V-PDB). The total C and N concentrations of all samples were measured using the Vario MAX cube (Elementar; Hanau, Germany) at the Central Laboratory, XTBG (Xishuangbanna Tropical Botanical Garden). The total P concentration of plant tissue was determined through inductively coupled plasma atomic-emission spectrometry (Thermo Fisher; Waltham, USA) after digestion in HNO3–HClO4.
For the study of plant interspecific competition for water, we collected soil and plant xylem samples separately during the extremely dry season (on 12 March 2016) and the pronounced rainy season (on 20 July 2016) for a comparison of plant water-absorbing patterns in water-limited and water-abundant environments. This design mainly considers the occurrence of plant competition for water when water availability is low, and the best way to understand plant competition for water is to compare the functional traits of water use when water is limited and abundant [17]. To be specific, March is the driest period in this region, and it is also the leaf-flushing period of the rubber trees (Figure S3a,b), which suggests that they rubber trees have enough reserved water for their upcoming leaf expansion [25]. Therefore, the competition for water during this period may be very intense in an RS-AFS. In contrast, due to adequate rainfall in July (Figure S3b), soil water would be abundant in July, which may result in weak competition for water in an RS-AFS. Therefore, exploring plant water-absorbing patterns in an RS-AFS under these two extremely contrary situations is beneficial for understanding the plant competition for water in an agroforestry system.
On each sampling day, we randomly selected three blocks (6 × 9 m) in the RM and RS-AFS for sampling. We used an increment borer to obtain xylem samples with lengths of 10–15 cm from 3‒5 rubber tree trunks in each study site and collected four samples of mature rhizomes from sharp-leaf galangals in the RS-AFS at the same time. It is worth explaining that the sharp-leaf galangal has no xylem. Therefore, water within its belowground rhizome seemed more suitable for stable isotope analysis because the root water uptake of most plant species either does not lead to isotopic discrimination or the discrimination is too low to be observed [22].
All sampled rubber trees and intercropped herbs were selected randomly in the blocks, and the sampled materials of one plant species in one block were mixed to form one sample. Therefore, there were three samples for each plant species in each site. The phloem on the xylem samples of both the rubber trees and the intercropped herbs was removed, and the treated samples were stored in a 15-ml screw-cap glass vial, sealed with Parafilm, and frozen immediately (−20 °C). At the same time, we dug a straight hole through a soil auger (4.5 cm in diameter) in the middle location of each selected block in the RM and RS-AFS (three holes for each study site) and collected soil samples from the surface (0–5-cm depths), shallow (5–15-cm depths), middle (15–30-cm depths), and deep (30–80-cm depths) soil layers. This soil sampling design mainly considered the relatively shallow vertical root depths of sharp-leaf galangals at depths of less than 30 cm and considered that there is no significant variation in soil water isotope compositions below 80 cm of depth [26]. Therefore, there was no need to sample soil from deeper soil layers below 80 cm of depth because a 0‒80-cm deep soil layer was enough for a comparison of the water-absorbing patterns of the rubber tree and the sharp-leaf galangal. Therefore, we regarded the 30–80-cm soil layer as a deep soil layer. A small part of each sample from each soil layer was collected into a 15-ml screw-cap glass vial and sealed with Parafilm immediately. The rest of the sampled soil was sealed in LDPE (Low-Density Polyethylene) zip-lock bags to measure the gravimetric soil water content (SWC) via oven-drying (105 °C, 48 h).
The water of all plant xylem and soil samples was extracted by means of cryogenic vacuum distillation with liquid nitrogen. The negative pressure was set to be 2 Pa, and the heating temperature was set at 80 °C for stems and 105 °C for soils. The average extraction time was 75 min for stems and 60 min for soils. The δ2H and δ18O values of each water sample were also measured through an IsoPrime100 (Isoprime, Stockport, UK), and the isotope ratios were expressed in parts per thousand relative to Vienna Standard Mean Ocean Water (V-SMOW).
2.3. Calculations and Statistical Analyses
We used general linear models (GLMs) to analyze the differences in the SWCs and soil water δ2H and δ18O values for different seasons, sites, and depths (i.e., soil layers of different depths). Similarly, the differences in leaf δ13C and leaf nutrient (C, N, and P) concentrations and ratios (C/N, C/P, and N/P) for different seasons, sites, and species were also analyzed by GLMs. For other purposes, we selected various kinds of combinations of influencing factors as fixed factors (Table 1; Table 2). If the analyzed results attained a significant level (p ≤ 0.05), differences between groups were compared using a post hoc Tukey’s test.
MixSIAR, which is a Bayesian mixing model, was used to distinguish the water sources of the rubber tree and the sharp-leaf galangal quantitatively in R [27]. In this study, we used two isotopic values (i.e., δ2H and δ18O) to estimate the water-absorbing patterns of the rubber trees and the sharp-leaf galangal on the premise of considering individual effects. Because the process in which source water enters plant roots and differentiates very little in water 2H and 18O can be ignored totally [20,23], the isotopic discrimination of this mixing model was set at zero. The raw isotopic values in the xylem water were treated as the mixture data, and the mean ± SD of the soil water isotopic values in different soil layers was set as the source data. The MCMC (Markov chain Monte Carlo) run length was set as “very long”.
In addition, the soil water evaporation line of each site in each season was generated by fitting a linear regression to the isotope data in a dual isotope plot (i.e., δ2H and δ18O), and a Pearson correlation analysis was performed to display the relationships between the leaf δ13C values; the leaf C, N, and P concentrations; and the leaf C/N, C/P, and N/P ratios of the rubber trees and the intercropped species. Meanwhile, on the basis of foliar parameters, an analysis of Pearson correlation-based similarities between the plant species was conducted, and the statistical analyses were performed using R 3.5.1 [28].
3. Results
3.1. Plant Xylem Water and Soil Water
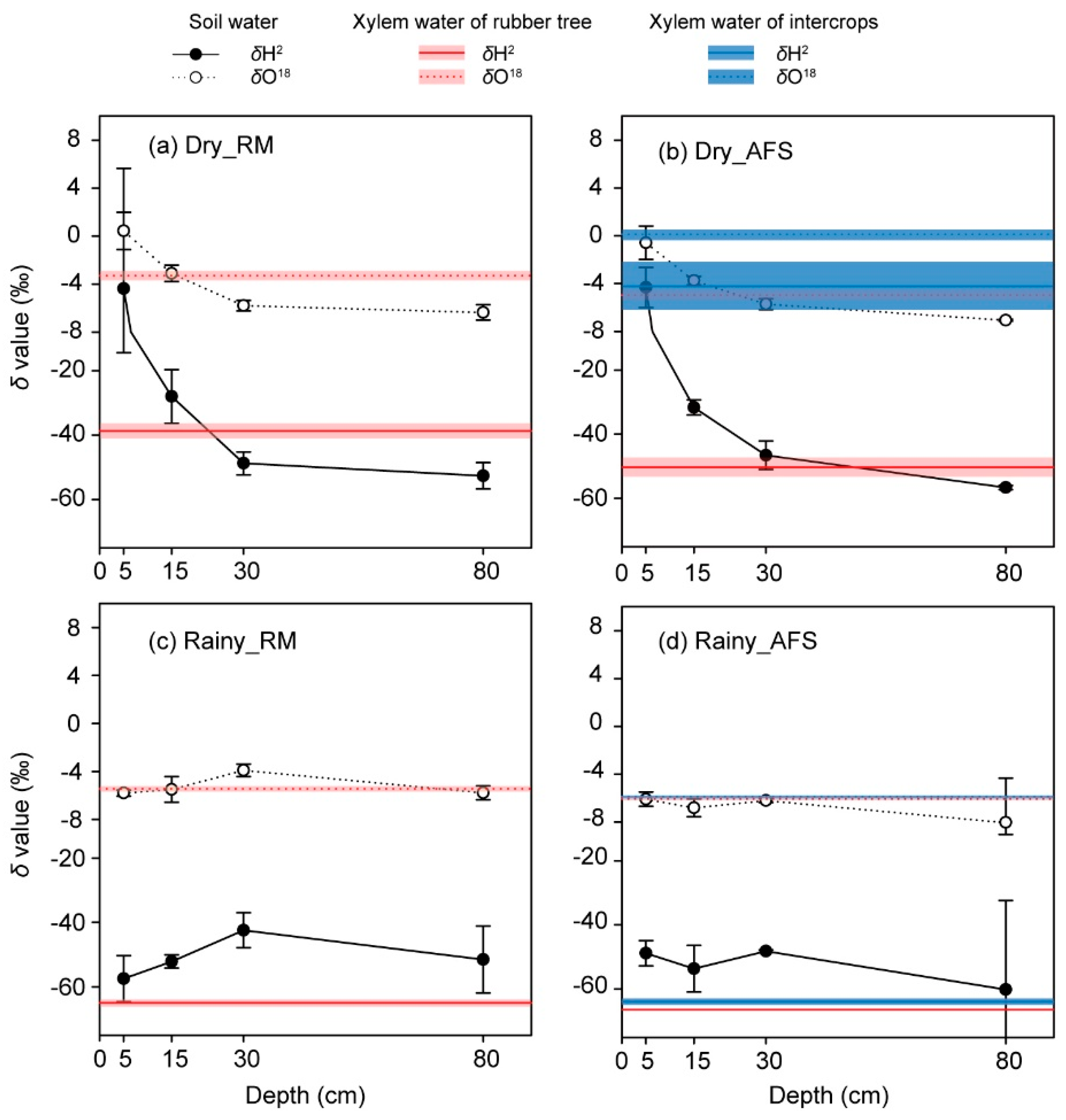
The xylem water δ2H and δ18O values of the rubber trees exhibited no significant differences between the sites either in the extremely dry season or in the pronounced rainy season (Figure 1). The xylem water δ2H and δ18O values of the rubber trees in the RS-AFS were significantly lower than the intercropped sharp-leaf galangal during the extremely dry season (p < 0.01, Figure 1), but the difference was not significant during the rainy season.
The δ2H and δ18O values of the soil water in both the RM and RS-AFS differed between the seasons and the sampling depths significantly (p < 0.01, Table 1). Similarly, the SWCs of each study site also exhibited significant variations between seasons and depths (p < 0.01, Table 1). In general, soil water δ2H and δ18O values were significantly higher in the extremely dry season than in the pronounced rainy season (p < 0.01), and the soil water isotope was significantly diminished from the surface soil layer to the deep soil layer during the extremely dry season, but the difference was not obvious in the pronounced rainy season (Figure 1c,d). In addition, the difference in the soil water isotope values between the sites was not significant (except for the δ18O values of the soil water in the pronounced rainy season; Table 1). In both the RM and RS-AFS, there existed a good linear relation between the δ2H and δ18O values of soil water.
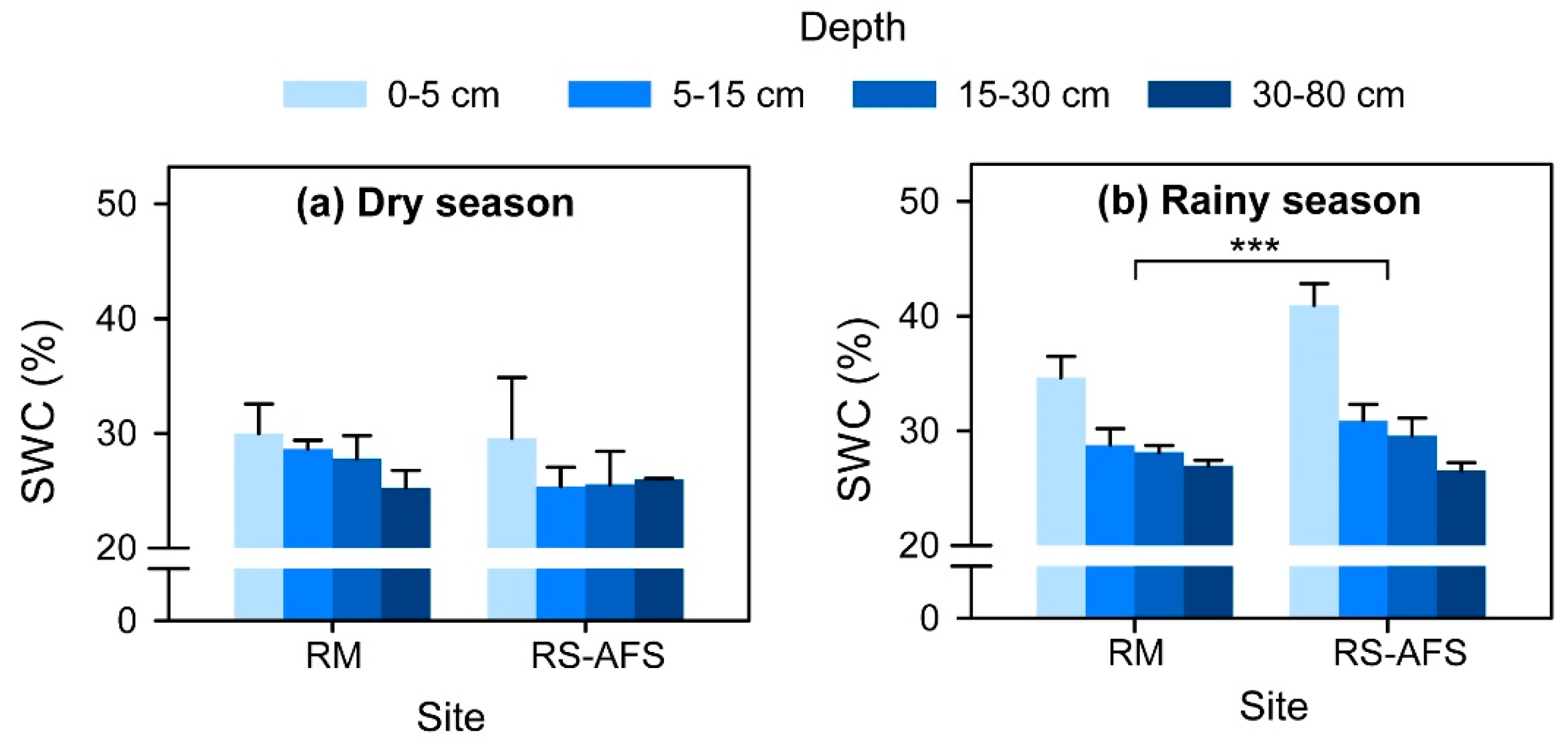
In the extremely dry season, the difference in the SWC between the sites was not significant (Table 1; Figure 2a). However, the SWC of the RS-AFS was significantly higher than that of the RM during the pronounced rainy season (p < 0.01; Figure 2b). In general, the SWC of both the RM and RS-AFS was significantly increased in the pronounced rainy season relative to the extremely dry season.
3.2. Plant Water Sources
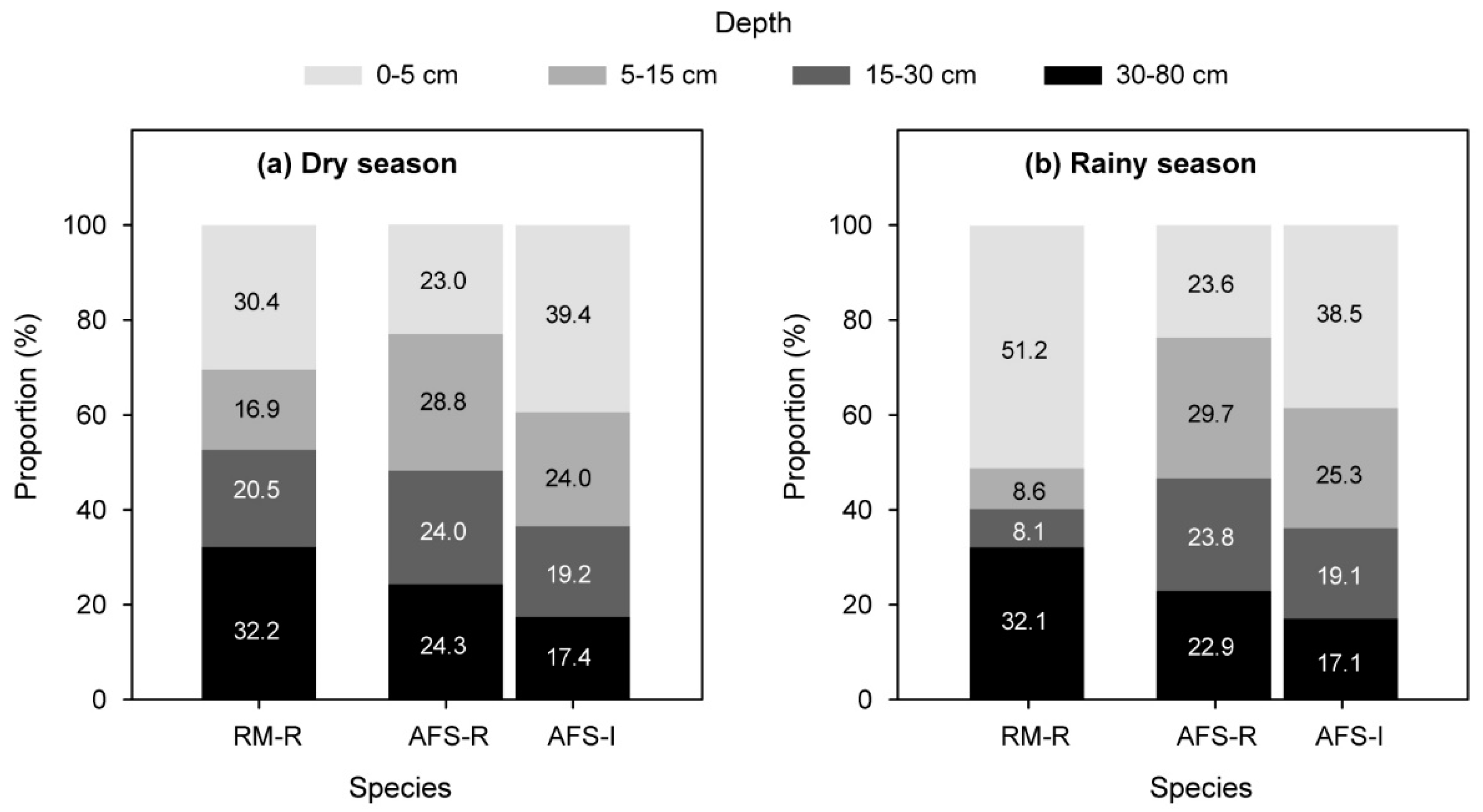
As the MixSIAR revealed, rubber trees in the RM absorbed a certain proportion of the soil water from the shallow and middle soil layers (5–30-cm depths) in the extremely dry season (37.4%, on average; Figure 3a), but the water-absorbing proportion from these soil layers decreased (16.7%, on average), and the main area of absorption shifted to the surface soil layer (0–5-cm depths) in the pronounced rainy season (51.2%, on average; Figure 3b).
However, more than 50% of the absorbed water of rubber trees in the RS-AFS came from the shallow and middle soil layers (5–30-cm depths), both in the extremely dry season and the pronounced rainy season. Therefore, the water-absorbing patterns of rubber trees in the RS-AFS looked more stable relative to rubber trees in the RM (Figure 3).
Relative to the rubber trees in the RS-AFS, the absorbed water of the intercropped sharp-leaf galangal came mostly from the surface, shallow, and middle soil layers (close to 80%; Figure 3), and the water-absorbing proportions of each soil layer were also stable between the seasons.
3.3. Leaf δ13C, Nutrient Concentrations, and Ratios
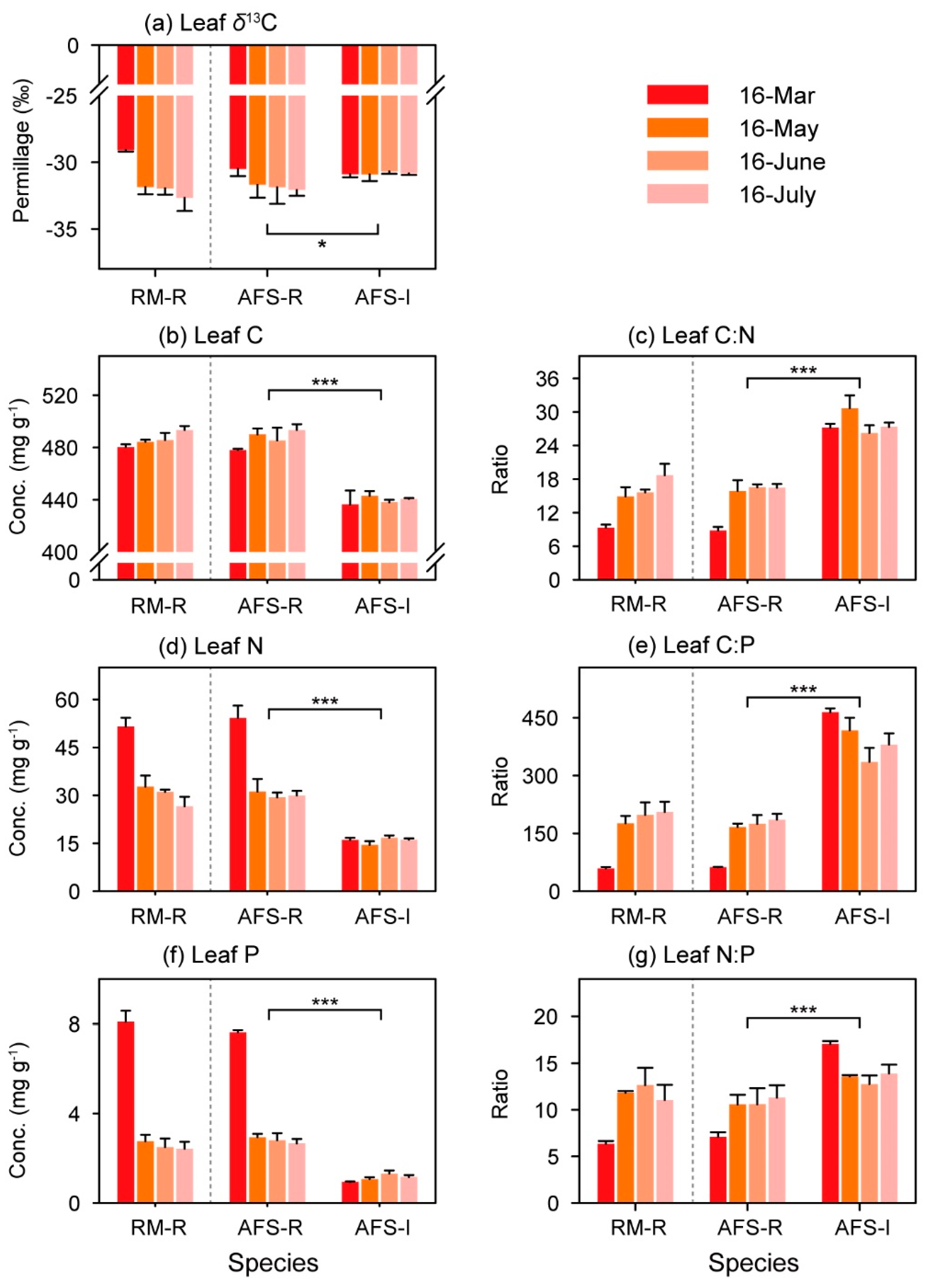
In the RM and RS-AFS, the mean leaf δ13C values of the rubber trees were −31.39‰ and −31.53‰, respectively. The mean leaf δ13C value was −30.83‰ for the sharp-leaf galangals in the RS-AFS. The rubber trees in the RM and the RS-AFS exhibited no significant differences in their leaf δ13C values (Table 2), but a significant decrease in the leaf δ13C values of the rubber trees was found from the dry season to the rainy season. In addition, rubber trees had significantly lower leaf δ13C values than did the intercropped sharp-leaf galangal in the RS-AFS (p < 0.05; Figure 4a). Furthermore, there was no significant difference in the leaf nutrient concentrations and ratios of the rubber trees between the two study sites (Table 2). In addition, in the RS-AFS, the leaf C, N, and P concentrations of the rubber trees were significantly higher than those of the sharp-leaf galangals, but the ratios of the leaf C/N, C/P, and N/P of the rubber trees were significantly lower than in the sharp-leaf galangals.
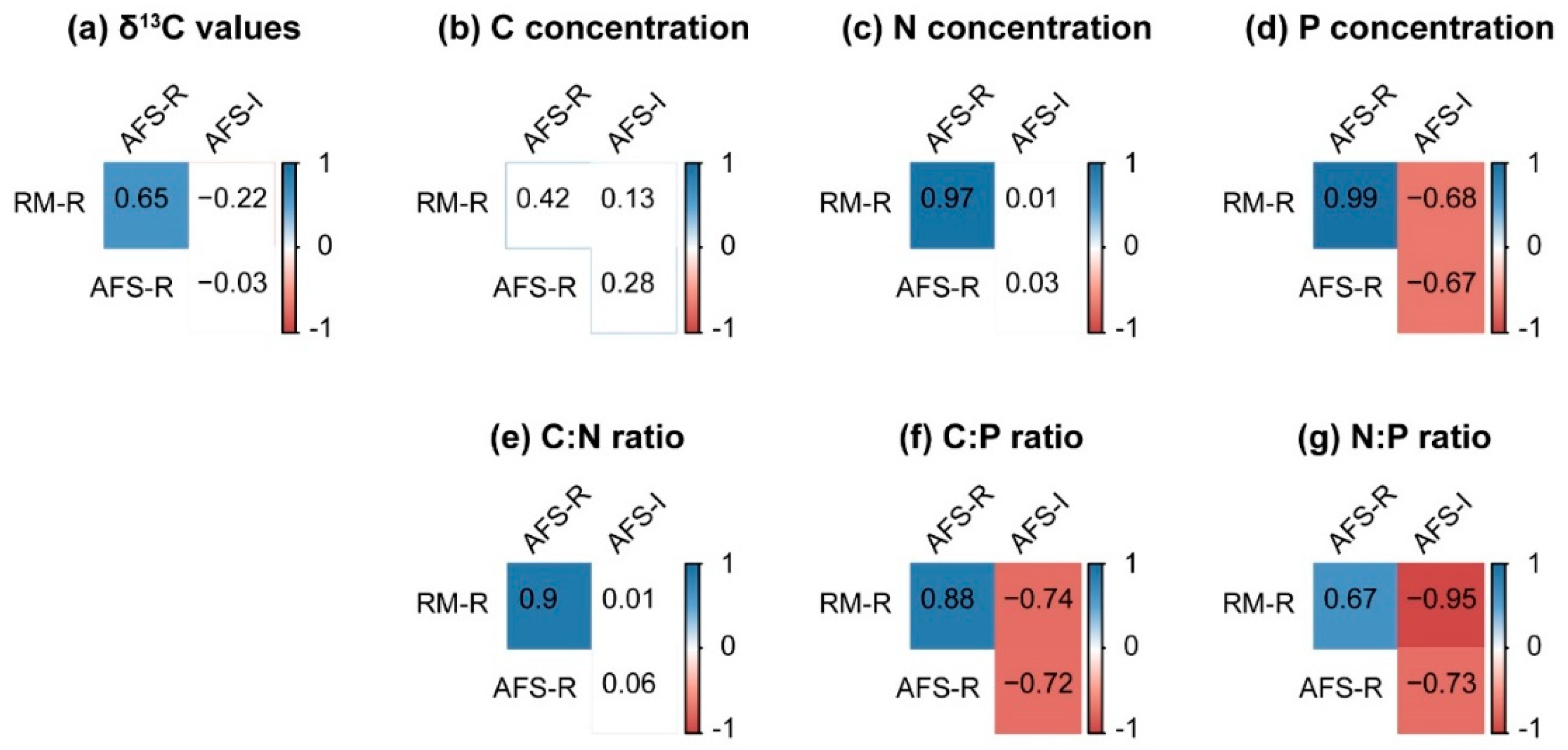
Furthermore, the leaf δ13C of the rubber tree was significantly correlated with the leaf nutrient concentration and leaf nutrient ratio in the RM (Figure 5a), but the correlation coefficients declined, and the significant correlations with leaf C, leaf C/P, and leaf N/P disappeared in the RS-AFS (Figure 5b). Like the rubber trees in the RS-AFS, the leaf δ13C of the intercrops was also significantly correlated with leaf N and P (Figure 5c). Except for leaf C, all leaf parameters of the rubber trees in the RM were significantly similar to those of rubber trees in the RS-AFS (Figure 6). However, in the RS-AFS, no significant similarity between the rubber trees and the intercropped sharp-leaf galangals was found in their leaf δ13C, leaf C and N, and leaf C/N (Figure 6a–c,e), but their leaf P concentrations and leaf C/P and N/P exhibited significant but opposite variations (Figure 6d,f,g).
4. Discussion
4.1. Plant Water-Absorbing Patterns
Because of the significantly lower δ2H and δ18O values of the soil water in the deep soil layer in comparison to those in the shallow soil layers during the extremely dry season, the significantly higher xylem water δ2H and δ18O values of the sharp-leaf galangal relative to the rubber trees in the RS-AFS suggested that the sharp-leaf galangal absorbed much more water from the surface and shallow soil layers than the rubber tree did in the extremely dry season. Indeed, as demonstrated by the results of the MixSIAR model, the sharp-leaf galangals absorbed more water at a depth of 0‒5 cm (Figure 3a), and their water-absorbing proportion from the surface soil layer to the deep soil layer gradually decreased in the extremely dry season. Obviously, this was mainly due to the shallow distribution of their roots (less than 30 cm of depth). Meanwhile, from the extremely dry season to the pronounced rainy season, the vertical water-absorbing patterns of this intercrop merely changed a little (Figure 3b). Relatively speaking, rubber trees in the RS-AFS seemed to absorb more water from the middle and deep soil layers than the sharp-leaf galangal did, both in the extremely dry season and in the pronounced rainy season (Figure 3).
Essentially, the monocultural rubber trees mainly depend on surface and shallow resources, because approximately 55%–86% of their feeder roots are concentrated in the soil layers above 20 cm of depth, and they are sensitive to a soil water deficit [29,30,31,32]. Therefore, we found that the rubber trees in the RM in this study mainly depended on the soil water from the soil layer above 30 cm of depth, and their absorption of the surface soil water (0‒5-cm depths) was greatly enhanced from the extremely dry season to the pronounced rainy season (Figure 3). However, the main water-absorbing area of rubber trees shifted to the middle soil layer (i.e., 5–30-cm soil layers) when the sharp-leaf galangal was intercropped with them, whether it was in the extremely dry season or in the pronounced rainy season.
Despite the vertical rhizospheres of the sharp-leaf galangals being less than 30 cm, deep soil water (from 30–80-cm deep soil layers) was also taken as the water source of this intercropped species. This was mainly because rubber trees exhibit the function of hydraulic redistribution [33]. As a common phenomenon, hydraulic redistribution can be explained by the mechanism of vascular plant root systems transporting deep soil water to a shallow soil layer or transporting water from a water-abundant soil layer to a water-exhausted soil layer [34]. Therefore, as one possible water source, the deep soil water contributed a certain proportion to the absorbed water source of the sharp-leaf galangals in the RS-AFS (Figure 3). Such a phenomenon could not only benefit the water use of the intercropped sharp-leaf galangals in the extremely dry season, but also balance the SWCs of different soil layers, thus helping both the rubber trees and their intercrops form relatively stable water-absorbing patterns relative to those of rubber trees in the RM (Figure 3).
It is obvious that the flexible water uptake of the rubber trees (Figure 3) helped to form relatively complementary water-absorbing patterns in the RS-AFS. This was mainly because rubber roots demonstrate strong plasticity [30], and such plastic roots could detect and avoid neighboring roots [35]. Therefore, hydrological niche segregation, which is a phenomenon where coexisting plants use a water source through different strategies of water use [21], thus helped the rubber trees and the sharp-leaf galangals form relatively complementary water-absorbing patterns in the RS-AFS gradually. Therefore, the interspecific water competition between the rubber trees and the sharp-leaf galangals was particularly small in the RS-AFS, even in the extremely dry season. Moreover, the relatively deeper water-absorbing behavior of the rubber trees in the RS-AFS during the rainy season indicated a deeper rooting depth of rubber trees in the RS-AFS. Such deep rooting behavior (from rubber trees in an RS-AFS) benefits the hydrological processes within the soil in an RS-AFS. For example, a dye tracer experiment found that the deeper rooting phenomenon of rubber trees in rubber-based agroforestry systems could reduce soil compaction, optimize soil structure, improve soil water infiltration, enhance soil water-holding capacity, and facilitate soil water movement [36]. In addition, nutrient mass flow may also be enhanced because of enhanced infiltration, and rubber trees in an RS-AFS therefore may absorb more nutrients because deep roots imply the extension of root contact areas with soil nutrients [23]. Furthermore, the deep rooting depth of rubber trees in an RS-AFS could help improve their nutrient uptake efficiency from deep soil layers, thus effectively reducing nutrient losses through deep leaching [37] (Liu et al. found that a large proportion of P compounds were accumulated in the deep soil layers in rubber agroforestry systems [38]).
4.2. Plant Water Use Efficiency and Soil Water Conditions
Commonly, differences in leaf C isotope discrimination between C3 and C4 plants are great because of the great differences in their dark reactions in photosynthesis [39]. Therefore, the leaf δ13C values of C3 plants typically range between −20‰ and −37‰, and the leaf δ13C values of C4 plants mainly range between −12‰ and −16‰ [40,41]. That is, rubber trees and sharp-leaf galangals both belong to C3 plants (Figure 4a).
For C3 plants, their leaf δ13C values could represent their long-term WUEs and reflect the plant response to soil drought [40]. Commonly, high δ13C values always correspond to high WUEs [42,43]. Therefore, the similar leaf δ13C values of the rubber trees in the RM and the RS-AFS (Figure 4a; Figure 6a) suggest that the WUE of rubber trees remained the same with intercropping with sharp-leaf galangal. It was discovered that the SWC in the RS-AFS did not differ from the RM in the extremely dry season, and the SWC in the RS-AFS was even higher than in the RM in the pronounced rainy season (Figure 2). The results above imply that the competition for water in the RS-AFS was not too intense to reduce soil water availability. In addition, the WUE of the rubber trees and sharp-leaf galangals in the RS-AFS displayed no apparent difference in different seasons (Table 2). This phenomenon thereby implies that the internal microclimatic environment in the RS-AFS was stable, because environmental factors (e.g., moisture and temperature) also affect plant leaf δ13C greatly [43].
The soil water δ2H and δ18O values were higher in the extremely dry season than in the pronounced rainy season because the strong soil evaporation and low rainwater supply in the dry season generally result in the enrichment of the stable hydrogen and oxygen isotopes of soil water [19]. Similarly, the seasonal variations in rainfall also led to low SWC in the extremely dry season but high SWC in the pronounced rainy season. Theoretically speaking, the soil in rubber agroforestry systems could contain much more water because intercropping could increase interception, decrease runoff, and improve soil water infiltration and soil water-holding capacity [36]. Therefore, a significantly higher SWC was found in the RS-AFS in the pronounced rainy season. However, the SWC of the RS-AFS was as low as that of the RM in the extremely dry season. Excluding the causes of intense soil evaporation and low rainfall in this season (Figure 3a), these results might have been due to the extra consumption of the sharp-leaf galangal, which showed relatively lower WUEs compared to the rubber trees in the RS-AFS. In addition, rubber trees may also aggravate soil water consumption because they must access enough reserve water for leaf expansion during this season [25]. Anyway, the soil water in the RS-AFS was no less than that in the RM. However, the water in the middle soil layer of the RS-AFS consumed more relative to the RM in the extremely dry season (Figure 2a). This was probably because the rubber trees in the RS-AFS preferred to absorb water from the middle soil layers (Figure 3).
In short, intercropping sharp-leaf galangal could improve soil water conditions distinctly in the pronounced rainy season, but these benefits were not apparent in the extremely dry season. This finding was consistent with a previous similar study, which demonstrated that soil water shortages still existed in this kind of intercropping system in the dry season, and a soil drought would limit the normal growth and photosynthesis of sharp-leaf galangals [15]. Therefore, it seems essential to conduct appropriate irrigation in an RS-AFS for sharp-leaf galangals in the dry season, although such a shortage of soil water has limited impacts on rubber trees.
4.3. Plant Leaf Carbon, Nitrogen, and Phosphorus Concentrations and Ratios
Commonly, C, N, and P are the most essential nutrients in the composition of various proteins and genetic materials in plants [44]. In addition, C/N/P ratios are closely associated with many eco-physiological functions of plants [45,46], and thus some well-known hypotheses and concepts have been put forward, such as the growth rate hypothesis [47]. Therefore, the similar leaf nutrient concentrations and ratios between the rubber trees in the RM and the RS-AFS (Figure 4c–g; Figure 6c–g) suggest that the interspecific competition in the RS-AFS did not affect the leaf physiological functions of the rubber trees. This was probably because the water-absorbing patterns of the rubber trees in the RS-AFS were relatively fixed, and the main water-absorbing soil layers of plants are also the main soil layers where plants absorb soil N and P [48]. Therefore, the nutrient absorption of the rubber trees in the RS-AFS may not have been affected by the interspecific competition.
In addition, the significant and negative correlation between the leaf P of the rubber trees and the sharp-leaf galangals in the RS-AFS (Figure 5d) suggested their opposite P use strategies, and such P use was possibly affected by their water use behaviors, because their leaf δ13C was significantly and positively correlated with their leaf P (Figure 5b,c). Because intercropping can increase soil P mobility in a rubber-based agroforestry system, a large proportion of P compounds accumulate in the deep soil layers [38]. Therefore, P acquisition by rubber trees in the RS-AFS benefited from water-absorbing patterns (Figure 3). However, such phenomena might not be consistent with the P requirement of intercropped sharp-leaf galangal, which has very shallow absorption areas. Therefore, interspecific competition in an RS-AFS may have a great impact on the growth of sharp-leaf galangal [15].
As indicated by the growth rate hypothesis, fast-growing organisms generally exhibit low C/P and N/P ratios in their tissues [47]. Therefore, the significantly lower leaf C/P and N/P ratios of the rubber trees in the RS-AFS relative to their intercrops (Figure 4) seemed to indicate that their growth was relatively more vigorous than that of the sharp-leaf galangals. Additionally, the opposite variation tendency in the leaf C/P and N/P ratios of the rubber trees and the sharp-leaf galangals in the RS-AFS (Figure 6f,g) demonstrated that their life and competition strategies were quite different. Generally, slow-growing species exhibit lower rates of resource acquisition and longer leaf life than fast-growing species do [49,50], and plants with long-lived leaves can reduce their nutrient requirements to maintain their leaf areas in resource-limited environments [23]. These characteristics are essentially consistent with those of sharp-leaf galangals [15]. Therefore, as evergreen perennials, the sharp-leaf galangals in the RS-AFS seemed to have chosen a persistence competition strategy, which is regarded as a sit-and-wait approach for available resources due to their characteristics of long leaf lives, fixed water-absorbing patterns, and slow growth rates [51]. However, the rubber trees showed a foraging competition strategy, which reflected rapid plant growth toward available resource patches; the isolation of resources; and sustained, rapid, and controlled growth toward more available resources [51].
5. Conclusions
Rubber trees in the RM absorbed more water from the surface soil layers, but they would take up more water from the shallow and middle soil layers after six years of intercropping with the sharp-leaf galangals. This was because the intercropped sharp-leaf galangals mainly depended on surface and shallow soil water, and the root niches of the rubber trees and the sharp-leaf galangals in the RS-AFS seemed to overlap greatly at the beginning of the intercropping. Due to the competitive foraging strategy of the rubber trees in the RS-AFS, the feeder roots of the rubber tree would enter deeper soil layers. Then, a hydrologic niche separation occurred, and the stable and complementary water use patterns of the rubber trees and the sharp-leaf galangal were formed in the RS-AFS. Therefore, the competition for water in this agroforestry system was weak, and such competition did not change the WUE of the rubber trees. It is worth mentioning that the soil in the RS-AFS could contain more water during the pronounced rainy season, but the SWCs of the RS-AFS were also as low as those in the RM during the extremely dry season. Therefore, irrigation during the dry season is necessary to maintain the healthy growth of sharp-leaf galangals. In addition, competition for soil P between the rubber trees and the sharp-leaf galangals in the RS-AFS was obvious, but there was no distinct influence on the P requirement for the growth of rubber trees. Therefore, P fertilization should also be applied properly in the planting rows of sharp-leaf galangal, especially in the surface soil layer. In brief, the interspecific competition below the ground did not affect the water use and nutrient uptake of the rubber trees.
Supplementary Materials
The following are available online at https://www.mdpi.com/1999-4907/10/10/924/s1, Figure S1: Schematic diagrams of (a) the rubber monoculture and (b) the rubber tree and sharp-leaf galangal agroforestry system (RS-AFS). Figure S2: Randomly selected blocks for sampling. Figure S3: (a) Information on the phenophases of rubber trees and (b) the monthly precipitation and monthly mean temperature during the study period (data were provided from the Xishuangbanna Station for Tropical Rainforest Ecosystem Studies). FS, fruit setting; FR, fruit ripening; DS, dormant stage; LS, leaf shedding; LF, leaf flushing; LE, leaf expansion; and FP, flowering phase.
Author Contributions
J.W. and W.L. designed the experiments; J.W., H.Z., and C.C. conducted the experiment, analyzed the data, and wrote the first draft; W.L. and X.J. revised and edited the manuscript.
Funding
This study was supported by the National Natural Science Foundation of China (31570622, 41701029, and 31800356), the National Postdoctoral Program for Innovative Talents (BX201700278), the China Postdoctoral Science Foundation (2018M633437), the Fourth Oriented Training Foundation for Postdoctors of Yunnan Province, the Scientific Research Foundation for Postdoctors of Yunnan Province, the “PhD Plus” Talents Training Program Foundation of XTBG, the CAS 135 Program (2017XTBG-F01), and the Foundation for the CAS Key Laboratory of Tropical Forest Ecology (09KF001B04).
Acknowledgments
We thank Fu Y., Deng Y., Liu M.N., and the Central Laboratory of XTBG for the help provided.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cornish, K. Alternative natural rubber crops: Why should we care? Technol. Innov. 2017, 18, 244–255. [Google Scholar] [CrossRef]
- Mann, C.C. Addicted to rubber. Science 2009, 325, 564–566. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.; Castella, J.C. Expansion of rubber (Hevea brasiliensis) in Mainland Southeast Asia: What are the prospects for smallholders? J. Peasant Stud. 2013, 40, 155–170. [Google Scholar] [CrossRef]
- Wigboldus, S.; Hammond, J.; Xu, J.; Yi, Z.F.; He, J.; Klerkx, L.; Leeuwis, C. Scaling green rubber cultivation in Southwest China–An integrative analysis of stakeholder perspectives. Sci. Total Environ. 2017, 580, 1475–1482. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Kono, Y.; Kobayashi, S. The process of expansion in commercial banana cropping in tropical China: A case study at a Dai village, Mengla County. Agric. Syst. 2014, 124, 32–38. [Google Scholar] [CrossRef]
- Zhang, J.Q.; Corlett, R.T.; Zhai, D. After the rubber boom: Good news and bad news for biodiversity in Xishuangbanna, Yunnan, China. Reg. Environ. Chang. 2019, 19, 1–12. [Google Scholar] [CrossRef]
- Li, H.; Ma, Y.; Aide, T.M.; Liu, W. Past, present and future land-use in Xishuangbanna, China and the implications for carbon dynamics. For. Ecol. Manag. 2008, 255, 16–24. [Google Scholar] [CrossRef]
- Mann, C.C. Why We (Still) Can’t Live Without Rubber. Available online: https://www.nationalgeographic.com/magazine/2016/01/southeast-asia-rubber-boom/ (accessed on 20 September 2019).
- Song, Q.H.; Tan, Z.H.; Zhang, Y.P.; Sha, L.Q.; Deng, X.B.; Deng, Y.; Zhao, W. Do the rubber plantations in tropical China act as large carbon sinks? iForest 2013, 7, 42–47. [Google Scholar] [CrossRef]
- Blagodatsky, S.; Xu, J.; Cadisch, G. Carbon balance of rubber (Hevea brasiliensis) plantations: A review of uncertainties at plot, landscape and production level. Agric. Ecosyst. Environ. 2016, 221, 8–19. [Google Scholar] [CrossRef]
- Zhang, M.; Chang, C.; Quan, R. Natural forest at landscape scale is most important for bird conservation in rubber plantation. Biol. Conserv. 2017, 210, 243–252. [Google Scholar] [CrossRef]
- Langenberger, G.; Cadisch, G.; Martin, K.; Min, S.; Waibel, H. Rubber intercropping: A viable concept for the 21st century? Agrofor. Syst. 2017, 91, 577–596. [Google Scholar] [CrossRef]
- John, J.; Nair, A.M. Prospects of allelopathic research in multi-storey cropping systems. In Allelopathy in Ecological Agriculture and Forestry; Narwal, S.S., Hoagland, R.E., Dilday, R.H., Reigosa Roger, M.J., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 159–179. [Google Scholar]
- He, Z.H.; Ge, W.; Yue, G.G.L.; Bik-San Lau, C.; He, M.F.; But, P.P.H. Anti-angiogenic effects of the fruit of Alpinia oxyphylla. J. Ethnopharmacol. 2010, 132, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Liu, Q.F.; Yan, J.K.; Zhang, T.L.; Wang, Q.Y. Seasonal changes of photosynthetic characteristics of Alpinia oxyphylla growing under Hevea brasiliensis. Chin. J. Plant Ecol. 2018, 42, 585–594. [Google Scholar]
- FAO and IAEA. Management of Agroforestry Systems for Enhancing Resource use Efficiency and Crop Productivity. Available online: https://www.iaea.org/publications/8181/management-of-agroforestry-systems-for-enhancing-resource-use-efficiency-and-crop-productivity (accessed on 29 September 2019).
- Craine, J.M.; Dybzinski, R. Mechanisms of plant competition for nutrients, water and light. Funct. Ecol. 2013, 27, 833–840. [Google Scholar] [CrossRef]
- Zechmeister-Boltenstern, S.; Keiblinger, K.M.; Mooshammer, M.; Peñuelas, J.; Richter, A.; Sardans, J.; Wanek, W. The application of ecological stoichiometry to plant-microbial-soil organic matter transformations. Ecol. Monogr. 2015, 85, 133–155. [Google Scholar] [CrossRef]
- Fry, B. Stable Isotope Ecology; Springer: New York, NY, USA, 2006. [Google Scholar]
- White, J.W.; Cook, E.R.; Lawrence, J.R. The DH ratios of sap in trees: Implications for water sources and tree ring DH ratios. Geochim. Cosmochim. Acta 1985, 49, 237–246. [Google Scholar] [CrossRef]
- Silvertown, J.; Araya, Y.; Gowing, D. Hydrological niches in terrestrial plant communities: A review. J. Ecol. 2015, 103, 93–108. [Google Scholar] [CrossRef]
- Youri, R.; Mathieu, J. Reviews and syntheses: Isotopic approaches to quantify root water uptake: A review and comparison of methods. Biogeosciences 2017, 14, 2199–2224. [Google Scholar]
- Chapin, F.S., III; Matson, P.A.; Vitousek, P. Principles of Terrestrial Ecosystem Ecology; Springer: New York, NY, USA, 2011. [Google Scholar]
- Everard, K.; Seabloom, E.W.; Harpole, W.S.; de Mazancourt, C. Plant water use affects competition for nitrogen: Why drought favors invasive species in California. Am. Nat. 2009, 1, 85–97. [Google Scholar] [CrossRef]
- Guardiola-Claramonte, M.; Troch, P.A.; Ziegler, A.D.; Giambelluca, T.W.; Durcik, M.; Vogler, J.B.; Nullet, M.A. Hydrologic effects of the expansion of rubber (Hevea brasiliensis) in a tropical catchment. Ecohydrol. 2010, 3, 306–314. [Google Scholar] [CrossRef]
- Wu, J.; Liu, W.; Chen, C. Can intercropping with the world’s three major beverage plants help improve the water use of rubber trees? J. Appl. Ecol. 2016, 53, 1787–1799. [Google Scholar] [CrossRef]
- Parnell, A.C.; Phillips, D.L.; Bearhop, S.; Semmens, B.X.; Ward, E.J.; Moore, J.W.; Inger, R. Bayesian stable isotope mixing models. Environmetrics 2013, 24, 387–399. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing Computer Software; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- George, S.; Suresh, P.R.; Wahid, P.A.; Nair, R.B.; Punnoose, K.I. Active root distribution pattern of Hevea brasiliensis determined by radioassay of latex serum. Agrofor. Syst. 2009, 76, 275–281. [Google Scholar] [CrossRef]
- Priyadarshan, P. Biology of Hevea Rubber; CABI: Wallingford, UK, 2011. [Google Scholar]
- Kobayashi, N.; Kumagai, T.O.; Miyazawa, Y.; Matsumoto, K.; Tateishi, M.; Lim, T.K.; Yin, S. Transpiration characteristics of a rubber plantation in central Cambodia. Tree Physiol. 2014, 34, 285–301. [Google Scholar] [CrossRef] [Green Version]
- Giambelluca, T.W.; Mudd, R.G.; Liu, W.; Ziegler, A.D.; Kobayashi, N.; Kumagai, T.O.; Yin, S. Evapotranspiration of rubber (Hevea brasiliensis) cultivated at two plantation sites in Southeast Asia. Water Resour. Res. 2016, 52, 660–679. [Google Scholar] [CrossRef]
- Wu, J.; Liu, W.; Chen, C. How do plants share water sources in a rubber-tea agroforestry system during the pronounced dry season. Agric. Ecosyst. Environ. 2017, 236, 69–77. [Google Scholar] [CrossRef]
- Bleby, T.M.; Mcelrone, A.J.; Jackson, R.B. Water uptake and hydraulic redistribution across large woody root systems to 20 m depth. Plant Cell Environ. 2010, 33, 2132–2148. [Google Scholar] [CrossRef]
- Callaway, R.M. The detection of neighbors by plants. Trends Ecol. Evol. 2002, 17, 104–105. [Google Scholar] [CrossRef]
- Jiang, X.J.; Liu, W.; Wu, J.; Wang, P.; Liu, C.; Yuan, Z.Q. Land Degradation Controlled and Mitigated by Rubber-based Agroforestry Systems through Optimizing Soil Physical Conditions and Water Supply Mechanisms: A Case Study in Xishuangbanna, China. Land Degrad. Dev. 2017, 28, 2277–2289. [Google Scholar] [CrossRef]
- Pierret, A.; Maeght, J.L.; Clément, C.; Montoroi, J.P.; Hartmann, C.; Gonkhamdee, S. Understanding deep roots and their functions in ecosystems: An advocacy for more unconventional research. Ann. Bot. 2016, 118, 621–635. [Google Scholar] [CrossRef]
- Liu, C.; Jin, Y.; Liu, C.; Tang, J.; Wang, Q.; Xu, M. Phosphorous fractions in soils of rubber-based agroforestry systems: Influence of season, management and stand age. Sci. Total Environ. 2018, 616, 1576–1588. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, M.H. Carbon isotopes in photosynthesis. BioScience 1988, 38, 328–336. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Kohn, M.J. Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo) ecology and (paleo) climate. Proc. Natl. Acad. Sci. USA 2010, 107, 19691–19695. [Google Scholar] [CrossRef] [PubMed]
- Ehleringer, J.R.; Roden, J.; Dawson, T.E. Assessing ecosystem-level water relations through stable isotope ratio analyses. In Methods in Ecosystem Science; Sala, O.E., Jackson, R.B., Mooney, H.A., Howarth, R.W., Eds.; Springer: New York, NY, USA, 2000; pp. 181–198. [Google Scholar]
- Marshall, J.D.; Brooks, J.R.; Lajtha, K. Sources of variation in the stable isotopic composition of plants. In Stable Isotopes in Ecology and Environmental Science; Michener, R., Lajtha, K., Eds.; John Wiley Sons: Chichester, UK, 2007; pp. 22–60. [Google Scholar]
- Sardans, J.; Rivas-Ubach, A.; Penuelas, J. The elemental stoichiometry of aquatic and terrestrial ecosystems and its relationships with organismic lifestyle and ecosystem structure and function: A review and perspectives. Biogeochemistry 2012, 111, 1–39. [Google Scholar] [CrossRef]
- Sasaki, T.; Yoshihara, Y.; Jamsran, U.; Ohkuro, T. Ecological stoichiometry explains larger-scale facilitation processes by shrubs on species coexistence among understory plants. Ecol. Eng. 2010, 36, 1070–1075. [Google Scholar] [CrossRef]
- Sterner, R.W.; Elser, J.J. Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere; Princeton University Press: Oxford, UK, 2002. [Google Scholar]
- Elser, J.J.; Sterner, R.W.; Gorokhova, E.A.; Fagan, W.F.; Markow, T.A.; Cotner, J.B.; Weider, L.W. Biological stoichiometry from genes to ecosystems. Ecol. Lett. 2000, 3, 540–550. [Google Scholar] [CrossRef] [Green Version]
- Barber, S.A. Soil Nutrient Bioavailability: A Mechanistic Approach; John Wiley Sons: New York, NY, USA, 1995. [Google Scholar]
- Craine, J.M.; Fargione, J.; Sugita, S. Supply pre-emption, not concentration reduction, is the mechanism of competition for nutrients. New Phytol. 2005, 166, 933–940. [Google Scholar] [CrossRef]
- Craine, J.M. Competition for nutrients and optimal root allocation. Plant Soil. 2006, 285, 171–185. [Google Scholar] [CrossRef]
- Keddy, P.; Fraser, L.H.; Wisheu, I.C. A comparative approach to examine competitive response of 48 wetland plant species. J. Veg. Sci. 1998, 9, 777–786. [Google Scholar] [CrossRef]
Figure 1.
The δ2H and δ18O values of the consumers (i.e., the xylem water of rubber trees and intercrops) and their water sources (i.e., soil water within four different depths) at each site during the extremely dry season (a,b) and the pronounced rainy season (c,d). The bold lines inside of the shaded bars indicate mean values, and the width of the shaded bar indicates the SD. All data are expressed as the mean ± SD. “Dry_RM” and “Dry_AFS” indicate the rubber monoculture (RM) and the rubber tree and sharp-leaf galangal agroforestry system (RS-AFS) in the extremely dry season, respectively. Similarly, “Rainy_RM” and “Rainy_AFS” respectively indicate those two sites in the pronounced rainy season.
Figure 1.
The δ2H and δ18O values of the consumers (i.e., the xylem water of rubber trees and intercrops) and their water sources (i.e., soil water within four different depths) at each site during the extremely dry season (a,b) and the pronounced rainy season (c,d). The bold lines inside of the shaded bars indicate mean values, and the width of the shaded bar indicates the SD. All data are expressed as the mean ± SD. “Dry_RM” and “Dry_AFS” indicate the rubber monoculture (RM) and the rubber tree and sharp-leaf galangal agroforestry system (RS-AFS) in the extremely dry season, respectively. Similarly, “Rainy_RM” and “Rainy_AFS” respectively indicate those two sites in the pronounced rainy season.

Figure 2.
Soil water content (SWC) within the soil profiles at each site during (a) the dry season and (b) the rainy season. “RM” and “RS-AFS” indicate the rubber monoculture and the rubber tree and sharp-leaf galangal agroforestry system, respectively. Data are expressed as the mean ± SD. * p < 0.05, ** p < 0.01, *** p < 0.001.
Figure 2.
Soil water content (SWC) within the soil profiles at each site during (a) the dry season and (b) the rainy season. “RM” and “RS-AFS” indicate the rubber monoculture and the rubber tree and sharp-leaf galangal agroforestry system, respectively. Data are expressed as the mean ± SD. * p < 0.05, ** p < 0.01, *** p < 0.001.

Figure 3.
Mean water-absorbing proportion of each plant species in (a) the extremely dry season and (b) the pronounced rainy season. “RM-R” indicates rubber trees in the rubber monoculture. “AFS-R” and “AFS-I” indicate rubber trees and intercrops in the agroforestry system, respectively.
Figure 3.
Mean water-absorbing proportion of each plant species in (a) the extremely dry season and (b) the pronounced rainy season. “RM-R” indicates rubber trees in the rubber monoculture. “AFS-R” and “AFS-I” indicate rubber trees and intercrops in the agroforestry system, respectively.

Figure 4.
(a) Leaf δ13C; (b–d) leaf C, N, and P concentrations’ and (e–g) leaf C/N, C/P, and N/P ratios of each plant species at each site. Data are expressed as the mean ± SD. * p < 0.05, ** p < 0.01, *** p < 0.001. See Figure 3 for abbreviations.
Figure 4.
(a) Leaf δ13C; (b–d) leaf C, N, and P concentrations’ and (e–g) leaf C/N, C/P, and N/P ratios of each plant species at each site. Data are expressed as the mean ± SD. * p < 0.05, ** p < 0.01, *** p < 0.001. See Figure 3 for abbreviations.

Figure 5.
The Pearson correlations between all leaf parameters (i.e., leaf δ13C values; leaf C, N, and P concentrations; and leaf C/N, C/P, and N/P ratios) of (a) the rubber trees in the RM and the leaves of (b) the rubber trees and (c) their intercrops in the RS-AFS. The color indicates the sign of the correlation (blue indicates a positive correlation, and red indicates a negative correlation). All the correlation coefficients in the red and blue boxes reached a significant level (p < 0.05). See Figure 3 for abbreviations.
Figure 5.
The Pearson correlations between all leaf parameters (i.e., leaf δ13C values; leaf C, N, and P concentrations; and leaf C/N, C/P, and N/P ratios) of (a) the rubber trees in the RM and the leaves of (b) the rubber trees and (c) their intercrops in the RS-AFS. The color indicates the sign of the correlation (blue indicates a positive correlation, and red indicates a negative correlation). All the correlation coefficients in the red and blue boxes reached a significant level (p < 0.05). See Figure 3 for abbreviations.

Figure 6.
The Pearson correlation-based similarities between RM-R, AFS-R, and AFS-I, which were analyzed through the (a) leaf δ13C values; (b‒d) leaf C, N, and P concentrations; and (e‒g) leaf C/N, C/P, and N/P ratios of the rubber trees and their intercrops. The color indicates the sign of the correlation (blue indicates a positive correlation, and red indicates a negative correlation). All the correlation coefficients in the red and blue boxes were significant (p < 0.05).
Figure 6.
The Pearson correlation-based similarities between RM-R, AFS-R, and AFS-I, which were analyzed through the (a) leaf δ13C values; (b‒d) leaf C, N, and P concentrations; and (e‒g) leaf C/N, C/P, and N/P ratios of the rubber trees and their intercrops. The color indicates the sign of the correlation (blue indicates a positive correlation, and red indicates a negative correlation). All the correlation coefficients in the red and blue boxes were significant (p < 0.05).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Results of a general linear model testing the effects on soil water content (SWC) and soil water δ2H and δ18O. F-values and significance are reported. The results included the following: (A) the effects of season, site, and depth; the effects of season and depth (B) in the RM and (C) in the RS-AFS; and the effects of site and depth (D) in the dry season and (E) in the rainy season. “d.f.” indicates the degree of freedom, “F” indicates F value and “P” indicates P value. * p < 0.05, ** p < 0.01, *** p < 0.001.
Table 1.
Results of a general linear model testing the effects on soil water content (SWC) and soil water δ2H and δ18O. F-values and significance are reported. The results included the following: (A) the effects of season, site, and depth; the effects of season and depth (B) in the RM and (C) in the RS-AFS; and the effects of site and depth (D) in the dry season and (E) in the rainy season. “d.f.” indicates the degree of freedom, “F” indicates F value and “P” indicates P value. * p < 0.05, ** p < 0.01, *** p < 0.001.
| Tested Effects | δ2H | δ18O | SWC | |||||
|---|---|---|---|---|---|---|---|---|
| d.f. | F | P | F | P | F | P | ||
| (A) | Season | 1 | 47.66 | *** | 35.02 | *** | 35.66 | *** |
| Site | 1 | 0.39 | 9.5 | ** | 0.78 | |||
| Depth | 3 | 18.69 | *** | 21.29 | *** | 31.05 | *** | |
| Season * Site | 1 | 0.01 | 2.02 | 9.48 | ** | |||
| Season * Depth | 3 | 21.22 | *** | 18.52 | *** | 6.73 | ** | |
| Site * Depth | 3 | 0.74 | 0.25 | 1.92 | ||||
| Season * Site * Depth | 3 | 0.52 | 0.95 | 2.11 | ||||
| (B) | Season | 1 | 37.21 | *** | 21.43 | *** | 7.19 | * |
| Depth | 3 | 9.85 | *** | 17.94 | *** | 16.55 | *** | |
| Season * Depth | 3 | 22.71 | *** | 29.47 | *** | 2.7 | ||
| (C) | Season | 1 | 17.79 | *** | 17.62 | *** | 28.88 | *** |
| Depth | 3 | 9.66 | *** | 8.56 | ** | 16.46 | *** | |
| Season * Depth | 3 | 5.52 | ** | 3.64 | * | 5.12 | * | |
| (D) | Site | 1 | 0.38 | 2.68 | 1.54 | |||
| Depth | 3 | 106 | *** | 72.3 | *** | 2.85 | ||
| Site * Depth | 3 | 0.47 | 0.45 | 0.74 | ||||
| (E) | Site | 1 | 0.16 | 6.83 | * | 18.13 | *** | |
| Depth | 3 | 0.95 | 1.7 | 76.91 | *** | |||
| Site * Depth | 3 | 0.66 | 0.66 | 6.64 | ** | |||
Table 2.
Results of a general linear model testing the effects on leaf δ13C, leaf nutrient (C, N, and P) concentration, and leaf ratios (C/N, C/P, and N/P). F-values and significance are reported. The results included the following: (A) time effects on the rubber trees in the RM; (B) time effects on the rubber trees in the RS-AFS; (C) time effects on the intercrops in the RS-AFS; (D) the effects of time and site on the rubber trees; and (E) the effects of time and species on the plants in the RS-AFS. “d.f.” indicates the degree of freedom, “F” indicates F value and “P” indicates P value. * p < 0.05, ** p < 0.01, *** p < 0.001.
Table 2.
Results of a general linear model testing the effects on leaf δ13C, leaf nutrient (C, N, and P) concentration, and leaf ratios (C/N, C/P, and N/P). F-values and significance are reported. The results included the following: (A) time effects on the rubber trees in the RM; (B) time effects on the rubber trees in the RS-AFS; (C) time effects on the intercrops in the RS-AFS; (D) the effects of time and site on the rubber trees; and (E) the effects of time and species on the plants in the RS-AFS. “d.f.” indicates the degree of freedom, “F” indicates F value and “P” indicates P value. * p < 0.05, ** p < 0.01, *** p < 0.001.
| Tested Effects | δ13C | C | N | P | C/N | C/P | N/P | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| d.f. | F | P | F | P | F | P | F | P | F | P | F | P | F | P | ||
| (A) | Time | 3 | 19.86 | *** | 7.66 | ** | 50.44 | *** | 162.91 | *** | 24.62 | *** | 25.82 | *** | 15.21 | ** |
| (B) | Time | 3 | 2.01 | 3.82 | 50.58 | *** | 400.68 | *** | 34.6 | *** | 47.41 | *** | 7.12 | * | ||
| (C) | Time | 3 | 0.39 | 0.79 | 4.01 | 8.24 | ** | 5.57 | * | 10.81 | ** | 22.85 | *** | |||
| (D) | Time | 3 | 13.96 | *** | 8.8 | ** | 99.61 | *** | 436.11 | *** | 54.91 | *** | 63.05 | *** | 20.82 | |
| Site | 1 | 0.21 | 0.15 | 0.38 | 0.28 | 0.17 | 2.47 | 1.3 | ||||||||
| Time * Site | 3 | 2.06 | 0.77 | 1.41 | 2.06 | 2.23 | 0.57 | 1.65 | ||||||||
| (E) | Time | 3 | 1.58 | 3.79 | * | 47.78 | *** | 298.91 | *** | 18.42 | *** | 3.37 | * | 0.88 | ||
| Species | 1 | 7.05 | * | 395.01 | *** | 534.04 | *** | 1893.82 | *** | 665.24 | *** | 731.62 | *** | 119.17 | *** | |
| Time * Species | 3 | 2.1 | 0.9 | 46.58 | *** | 366.16 | *** | 14.35 | *** | 32.76 | *** | 20.8 | *** | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wu, J.; Zeng, H.; Chen, C.; Liu, W.; Jiang, X. Intercropping the Sharp-Leaf Galangal with the Rubber Tree Exhibits Weak Belowground Competition. Forests 2019, 10, 924. https://doi.org/10.3390/f10100924
AMA Style
Wu J, Zeng H, Chen C, Liu W, Jiang X. Intercropping the Sharp-Leaf Galangal with the Rubber Tree Exhibits Weak Belowground Competition. Forests. 2019; 10(10):924. https://doi.org/10.3390/f10100924
Chicago/Turabian StyleWu, Junen, Huanhuan Zeng, Chunfeng Chen, Wenjie Liu, and Xiaojin Jiang. 2019. "Intercropping the Sharp-Leaf Galangal with the Rubber Tree Exhibits Weak Belowground Competition" Forests 10, no. 10: 924. https://doi.org/10.3390/f10100924
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.