
Excessive Gestational Weight Gain Alters DNA Methylation and Influences Foetal and Neonatal Body Composition

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Anthropometric Assessment of the Pregnant Women
2.3. Foetal Body Composition
2.4. Anthropometry and Body Composition of Neonates
2.5. Sample Collection and DNA Extraction
2.6. Methylation Analysis
2.7. Data Analysis
3. Results
3.1. Characteristics of the Pregnant Women and Their Neonates
3.2. Anthropometry and Body Composition of the Pregnant Women and of Their Foetuses and Neonates
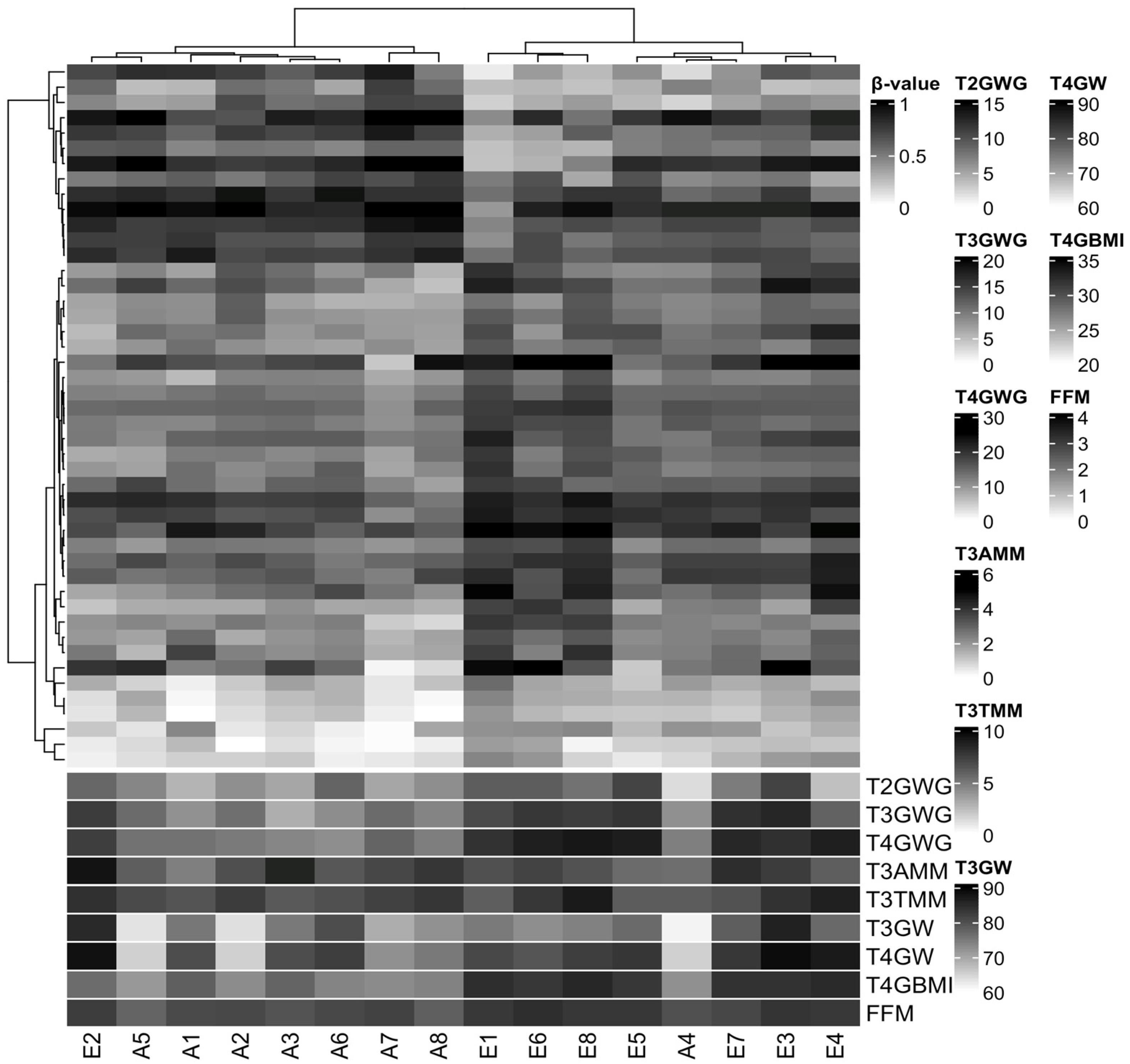
3.3. Characterization of DNA Methylation
3.4. DNA Methylation Changes Are Associated with Some Foetal and Neonatal Outcomes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Rasmussen, K.M.; Yaktine, A.L. (Eds.) Institute of Medicine (US) and National Research Council (US) Committee to Reexamine IOM Pregnancy Weight Guidelines. In Weight Gain during Pregnancy: Reexamining the Guidelines; National Academies Press: Washington, DC, USA, 2009. [Google Scholar]
- Guelinckx, I.; Devlieger, R.; Beckers, K.; Vansant, G. Maternal obesity: Pregnancy complications, gestational weight gain and nutrition. Obes. Rev. 2008, 9, 140–150. [Google Scholar] [PubMed]
- Centers for Disease Control and Prevention. QuickStats: Gestational weight gain* among women with full-term, singleton births, compared with recommendations—48 states and the District of Columbia, 2015. MMWR Morb. Mortal Wkly. Rep. 2016, 3, 2018. [Google Scholar]
- Goldstein, R.F.; Abell, S.K.; Ranasinha, S.; Misso, M.L.; Boyle, J.A.; Harrison, C.L.; Black, M.H.; Li, N.; Hu, G.; Corrado, F.; et al. Gestational weight gain across continents and ethnicity: Systematic review and meta-analysis of maternal and infant outcomes in more than one million women. BMC Med. 2018, 16, 153. [Google Scholar] [CrossRef] [PubMed]
- Carrilho, T.R.B.; Rasmussen, K.M.; Hutcheon, J.A.; Alves, R.F.S.; Farias, D.R.; Freitas-Costa, N.C.; Gonzalez, M.M.; Batalha, M.A.; Kac, G. Prevalence and temporal trends in prepregnancy nutritional status and gestational weight gain of adult women followed in the Brazilian Food and Nutrition Surveillance System from 2008 to 2018. Matern. Child Nutr. 2022, 18, e13240. [Google Scholar] [CrossRef]
- Barker, D.J. In utero programming of chronic disease. Clin. Sci. 1998, 95, 115–128. [Google Scholar] [CrossRef]
- Godfrey, K.M.; Sheppard, A.; Gluckman, P.D.; Lillycrop, K.A.; Burdge, G.C.; Mclean, C.; Rodford, J.; Slater-Jefferies, J.L.; Garratt, E.; Crozier, S.R.; et al. Epigenetic gene promoter methylation at birth is associated with child’s later adiposity. Diabetes 2011, 60, 1528–1534. [Google Scholar] [CrossRef]
- Portha, B.; Fournier, A.; Ah Kioon, M.D.; Mezger, V.; Movassat, J. Early environmental factors, alteration of epigenetic marks and metabolic disease susceptibility. Biochimie 2014, 97, 1–15. [Google Scholar] [CrossRef]
- Sharp, G.C.; Lawlor, D.A.; Richmond, R.C.; Fraser, A.; Simpkin, A.; Suderman, M.; Shihab, H.A.; Lyttleton, O.; McArdle, W.; Ring, S.M.; et al. Maternal pre-pregnancy BMI and gestational weight gain, offspring DNA methylation and later offspring adiposity: Findings from the Avon Longitudinal Study of Parents and Children. Int. J. Epidemiol. 2015, 44, 1288–1304. [Google Scholar] [CrossRef]
- Olerup, O.; Zetterquist, H. HLA-DR typing by PCR amplification with sequence-specific primers (PCR-SSP) in 2 hours: An alternative to serological DR typing in clinical practice including donor-recipient matching in cadaveric transplantation. Tissue Antigens 1992, 39, 225–235. [Google Scholar] [CrossRef]
- Morris, T.J.; Butcher, L.M.; Feber, A.; Teschendorff, A.E.; Chakravarthy, A.R.; Wojdacz, T.K.; Beck, S. ChAMP: 450k Chip Analysis Methylation Pipeline. Bioinformatics 2014, 30, 428–430. [Google Scholar] [CrossRef]
- Nordlund, J.; Bäcklin, C.L.; Wahlberg, P.; Busche, S.; Berglund, E.C.; Eloranta, M.L.; Flaegstad, T.; Forestier, E.; Frost, B.-M.; Harila-Saari, A.; et al. Genome-wide signatures of differential DNA methylation in pediatric acute lymphoblastic leukemia. Genome Biol. 2013, 14, r105. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Laird, P.W.; Shen, H. Comprehensive characterization, annotation and innovative use of Infinium DNA methylation BeadChip probes. Nucleic Acids Res. 2017, 45, e22. [Google Scholar] [PubMed]
- Teschendorff, A.E.; Marabita, F.; Lechner, M.; Bartlett, T.; Tegner, J.; Gomez-Cabrero, D.; Beck, S. A beta-mixture quantile normalization method for correcting probe design bias in Illumina Infinium 450 k DNA methylation data. Bioinformatics 2013, 29, 189–196. [Google Scholar] [CrossRef]
- Teschendorff, A.E.; Menon, U.; Gentry-Maharaj, A.; Ramus, S.J.; Gayther, S.A.; Apostolidou, S.; Jones, A.; Lechner, M.; Beck, S.; Jacobs, I.J.; et al. An epigenetic signature in peripheral blood predicts active ovarian cancer. PLoS ONE 2009, 4, e8274. [Google Scholar]
- Johnson, W.E.; Li, C.; Rabinovic, A. Adjusting batch effects in microarray expression data using empirical Bayes methods. Biostatistics 2007, 8, 118–127. [Google Scholar] [CrossRef]
- Houseman, E.A.; Accomando, W.P.; Koestler, D.C.; Christensen, B.C.; Marsit, C.J.; Nelson, H.H.; Wiencke, J.K.; Kelsey, K.T. DNA methylation arrays as surrogate measures of cell mixture distribution. BMC Bioinform. 2012, 13, 86. [Google Scholar] [CrossRef]
- Smyth, G.K. Linear models and empirical bayes methods for assessing differential expression in microarray experiments. Stat. Appl. Genet. Mol. Biol. 2004, 3, 1. [Google Scholar]
- Jaffe, A.E.; Murakami, P.; Lee, H.; Leek, J.T.; Fallin, M.D.; Feinberg, A.P.; Irizarry, R.A. Bump hunting to identify differentially methylated regions in epigenetic epidemiology studies. Int. J. Epidemiol. 2012, 41, 200–209. [Google Scholar] [CrossRef]
- McLean, C.Y.; Bristor, D.; Hiller, M.; Clarke, S.L.; Schaar, B.T.; Lowe, C.B.; Wenger, A.M.; Bejerano, G. GREAT improves functional interpretation of cis-regulatory regions. Nat. Biotechnol. 2010, 28, 495–501. [Google Scholar] [CrossRef]
- Venkataraman, H.; Ram, U.; Craik, S.; Arungunasekaran, A.; Seshadri, S.; Saravanan, P. Increased fetal adiposity prior to diagnosis of gestational diabetes in South Asians: More evidence for the ‘thin-fat’ baby. Diabetologia 2017, 60, 399–405. [Google Scholar] [CrossRef]
- Yerlikaya-Schatten, G.; Feichtinger, M.; Stopp, T.; Huhn, E.A.; Chalubinski, K.; Husslein, P.; Eppel, W.; Schatten, C.; Göbl, C.S. Trajectories of fetal adipose tissue thickness in pregnancies after gastric bypass surgery. Obes. Surg. 2020, 30, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Favero, G.; Paini, A.; De Ciuceis, C.; Rodella, L.F.; Moretti, E.; Porteri, E.; Rossini, C.; Ministrini, S.; Solaini, L.; Stefano, C.; et al. Changes in extracellular matrix in subcutaneous small resistance arteries of patients with essential hypertension. Blood Press. 2018, 27, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Holzman, M.A.; Ryckman, A.; Finkelstein, T.M.; Landry-Truchon, K.; Schindler, K.A.; Bergmann, J.M.; Jeannotte, L.; Mansfield, J.H. HOXA5 participates in brown adipose tissue and epaxial skeletal muscle patterning and in brown adipocyte differentiation. Front. Cell Dev. Biol. 2021, 9, 311. [Google Scholar] [CrossRef]
- Lefort, N.; Glancy, B.; Bowen, B.; Willis, W.T.; Bailowitz, Z.; De Filippis, E.A.; Brophy, C.; Meyer, C.; Højlund, K.; Yi, Z.; et al. Increased reactive oxygen species production and lower abundance of complex I subunits and carnitine palmitoyltransferase 1B protein despite normal mitochondrial respiration in insulin-resistant human skeletal muscle. Diabetes 2010, 59, 2444–2452. [Google Scholar] [CrossRef] [PubMed]
- Walley, A.J.; Jacobson, P.; Falchi, M.; Bottolo, L.; Andersson, J.C.; Petretto, E.; Bonnefond, A.; Vaillant, E.; Lecoeur, C.; Vatin, V.; et al. Differential coexpression analysis of obesity-associated networks in human subcutaneous adipose tissue. Int. J. Obes. 2012, 36, 137–147. [Google Scholar]
- Tulyeu, J.; Kumagai, H.; Jimbo, E.; Watanabe, S.; Yokoyama, K.; Cui, L.; Osaka, H.; Mieno, M.; Yamagata, T. Probiotics prevents sensitization to oral antigen and subsequent increases in intestinal tight junction permeability in juvenile–young adult rats. Microorganisms 2019, 7, 463. [Google Scholar] [CrossRef]
- Mackay, D.J.G.; Callaway, J.L.A.; Marks, S.M.; White, H.E.; Acerini, C.L.; Boonen, S.E.; Dayanikli, P.; Firth, H.V.; Goodship, J.A.; Haemers, A.P.; et al. Hypomethylation of multiple imprinted loci in individuals with transient neonatal diabetes is associated with mutations in ZFP57. Nat. Genet. 2008, 40, 949–951. [Google Scholar] [CrossRef]
- Parkkola, A.; Turtinen, M.; Härkönen, T.; Ilonen, J.; Knip, M.; Knip, M. Family history of type 2 diabetes and characteristics of children with newly diagnosed type 1 diabetes. Diabetologia 2021, 64, 581–590. [Google Scholar]
- Iyengar, N.M.; Zhou, X.K.; Mendieta, H.; El-Hely, O.; Giri, D.D.; Winston, L.; Falcone, D.J.; Wang, H.; Meng, L.; Ha, T.; et al. Effects of obesity on breast aromatase expression and systemic metabo-inflammation in women with BRCA1 or BRCA2 mutations. NPJ Breast Cancer 2021, 7, 18. [Google Scholar] [CrossRef]
- Sun, T.; Han, X. Death versus dedifferentiation: The molecular bases of beta cell mass reduction in type 2 diabetes. Semin. Cell Dev. Biol. 2020, 103, 76–82. [Google Scholar] [CrossRef]
- Jin, X.; Li, Y.; Guo, Y.; Jia, Y.; Qu, H.; Lu, Y.; Song, P.; Zhang, X.; Shao, Y.; Qi, D.; et al. ERα is required for suppressing OCT4-induced proliferation of breast cancer cells via DNMT1/ISL1/ERK axis. Cell Prolif. 2019, 52, e12612. [Google Scholar] [CrossRef]
- Wang, J.; Gao, F.; Zhao, X.; Cai, Y.; Jin, H. Integrated analysis of the transcriptome-wide m6A methylome in preeclampsia and healthy control placentas. PeerJ 2020, 8, e9880. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, D.; Johnston, A.D.; Nico, J.; Loh, L.N.; Jorge, Y.; Suzuki, M.; Macian, F.; Greally, J.M. Functional genomics of the pediatric obese asthma phenotype reveal enrichment of rho-gtpase pathways. Am. J. Respir. Crit. Care Med. 2020, 202, 259–274. [Google Scholar] [CrossRef]
- Shen, J.; Guo, T.; Wang, T.; Zhen, Y.; Ma, X.; Wang, Y.; Zhang, Z.-X.; Cai, J.-P.; Mao, W.; Zhu, F.-M.; et al. HLA-B∗07, HLA-DRB1∗07, HLA-DRB1∗12, and HLA-c∗03:02 strongly associate with BMI: Data from 1.3 million healthy Chinese adults. Diabetes 2018, 67, 861–871. [Google Scholar] [CrossRef]
- Karlsson, T.; Rask-Andersen, M.; Pan, G.; Höglund, J.; Wadelius, C.; Ek, W.E.; Johansson, Å. Contribution of genetics to visceral adiposity and its relation to cardiovascular and metabolic disease. Nat. Med. 2019, 25, 1390–1395. [Google Scholar] [CrossRef] [PubMed]
- Deane, C.S.; Willis, C.R.G.; Phillips, B.E.; Atherton, P.J.; Harries, L.W.; Ames, R.M.; Szewczyk, N.J.; Etheridge, T. Transcriptomic meta-analysis of disuse muscle atrophy vs. resistance exercise-induced hypertrophy in young and older humans. J. Cachexia Sarcopenia Muscle 2021, 12, 629–645. [Google Scholar] [CrossRef] [PubMed]
- Descipio, C.; Conlin, L.; Rosenfeld, J.; Tepperberg, J.; Pasion, R.; Patel, A.; McDonald, M.T.; Aradhya, S.; Ho, D.; Goldstein, J.; et al. Subtelomeric deletion of chromosome 10p15.3: Clinical findings and molecular cytogenetic characterization. Am. J. Med. Genet. Part A 2012, 158, 2152–2161. [Google Scholar] [CrossRef]
- Devarbhavi, P.; Telang, L.; Vastrad, B.; Tengli, A.; Vastrad, C.; Kotturshetti, I. Identification of key pathways and genes in polycystic ovary syndrome via integrated bioinformatics analysis and prediction of small therapeutic molecules. Reprod. Biol. Endocrinol. 2021, 19, 31. [Google Scholar] [CrossRef]
- Yang, I.V.; Lozupone, C.A.; Schwartz, D.A. The environment, epigenome, and asthma. J. Allergy Clin. Immunol. 2017, 140, 14–23. [Google Scholar] [CrossRef]
- Samblas, M.; Milagro, F.I.; Mansego, M.L.; Marti, A.; Martinez, J.A. PTPRS and PER3 methylation levels are associated with childhood obesity: Results from a genome-wide methylation analysis. Pediatr. Obes. 2017, 13, 149–158. [Google Scholar] [CrossRef]
- Mohn, F.; Schübeler, D. Genetics and epigenetics: Stability and plasticity during cellular differentiation. Trends Genet. 2009, 25, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Rzehak, P.; Covic, M.; Saffery, R.; Reischl, E.; Wahl, S.; Grote, V.; Weber, M.; Xhonneux, A.; Langhendries, J.-P.; Ferre, N.; et al. DNA-methylation and body composition in preschool children: Epigenome-wide-analysis in the European Childhood Obesity Project (CHOP)-Study. Sci. Rep. 2017, 7, 14349. [Google Scholar] [CrossRef]
- Voerman, E.; Santos, S.; Golab, B.P.; Amiano, P.; Ballester, F.; Barros, H.; Bergström, A.; Charles, M.-A.; Chatzi, L.; Chevrier, C.; et al. Maternal body mass index, gestational weight gain, and the risk of overweight and obesity across childhood: An individual participant data meta-analysis. PLoS Med. 2019, 16, e1002744. [Google Scholar] [CrossRef] [PubMed]
- Castillo, H.; Santos, I.S.; Matijasevich, A. Relationship between maternal pre-pregnancy body mass index, gestational weight gain and childhood fatness at 6–7 years by air displacement plethysmography. Matern. Child Nutr. 2015, 11, 606–617. [Google Scholar] [CrossRef]
- Orsso, C.E.; Colin-Ramirez, E.; Field, C.J.; Madsen, L.H.; Prado, C.M.; Haqq, A.M. Adipose tissue development and expansion from the womb to adolescence: An overview. Nutrients 2020, 12, 2735. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; Schmidt, H.; Lai, B.; Ge, K. Transcriptional and epigenomic regulation of adipogenesis. Mol. Cell. Biol. 2019, 39, e00601-18. [Google Scholar] [CrossRef] [PubMed]
- Christodoulides, C.; Lagathu, C.; Sethi, J.K.; Vidal, A. Adipogenesis and WNT signalling. Trends Endocrinol. Metab. 2009, 20, 16–24. [Google Scholar] [CrossRef]
- Rosen, E.D.; MacDougald, O.A. Adipocyte differentiation from the inside out. Nat. Rev. Mol. Cell Biol. 2006, 7, 885–896. [Google Scholar] [CrossRef]
- Rishi, V.; Bhattacharya, P.; Chatterjee, R.; Rozenberg, J.; Zhao, J.; Glass, K.; Fitzgerald, P.; Vinson, C. CpG methylation of half-CRE sequences creates C/EBPα binding sites that activate some tissue-specific genes. Proc. Natl. Acad. Sci. USA 2010, 107, 20311–20316. [Google Scholar] [CrossRef]
- Bigaard, J.; Frederiksen, K.; Tjønneland, A.; Thomsen, B.L.; Overvad, K.; Heitmann, B.L.; Sørensen, T.I.A. Body fat and fat-free mass and all-cause mortality. Obes. Res. 2004, 12, 1042–1049. [Google Scholar] [CrossRef]
- Lin, J.; Gu, W.; Huang, H. Effects of paternal obesity on fetal development and pregnancy complications: A prospective clinical cohort study. Front. Endocrinol. 2022, 13, 826665. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Variables | Excessive Gestational Weight Gain (n = 30) | Adequate Gestational Weight Gain (n = 45) | p |
|---|---|---|---|
| Pregnant women | |||
| Age (years) | 25.9 ± 6.0 | 29.0 ± 6.4 | 0.064 |
| Ethnicity | |||
| White | 10 (33.3%) | 15 (33.3%) | 0.351 |
| Black | 03 (10.0%) | 10 (22.2%) | |
| Brown | 17 (56.7%) | 20 (44.4%) | |
| Marital status | |||
| Single/without partner | 00 (0.0%) | 04 (8.8%) | 0.245 |
| Married/with partner | 30 (100.0%) | 41 (91.1%) | |
| Education | |||
| Elementary school | 01 (3.3%) | 09 (20.0%) | 0.115 |
| High school degree | 24 (80.0%) | 30 (66.7%) | |
| University degree | 05 (16.7%) | 06 (13.3%) | |
| Parity | |||
| 0 | 05 (16.7%) | 07 (15.6%) | 0.991 |
| 1 | 19 (63.3%) | 29 (64.4%) | |
| 2 a 4 | 06 (20.0%) | 09 (20.0%) | |
| Neonates | |||
| Age (weeks) | 39.9 ± 1.1 | 39.2 ± 1.4 | 0.162 |
| Sex | |||
| Female | 17 (56.6%) | 16 (35.6%) | 0.071 |
| Male | 13 (43,3%) | 29 (64.4%) | |
| Variables | Excessive Gestational Weight Gain (n = 30) | Adequate Gestational Weight Gain (n = 45) | p | ||
|---|---|---|---|---|---|
| Pregnant women | Mean | SD | Mean | SD | |
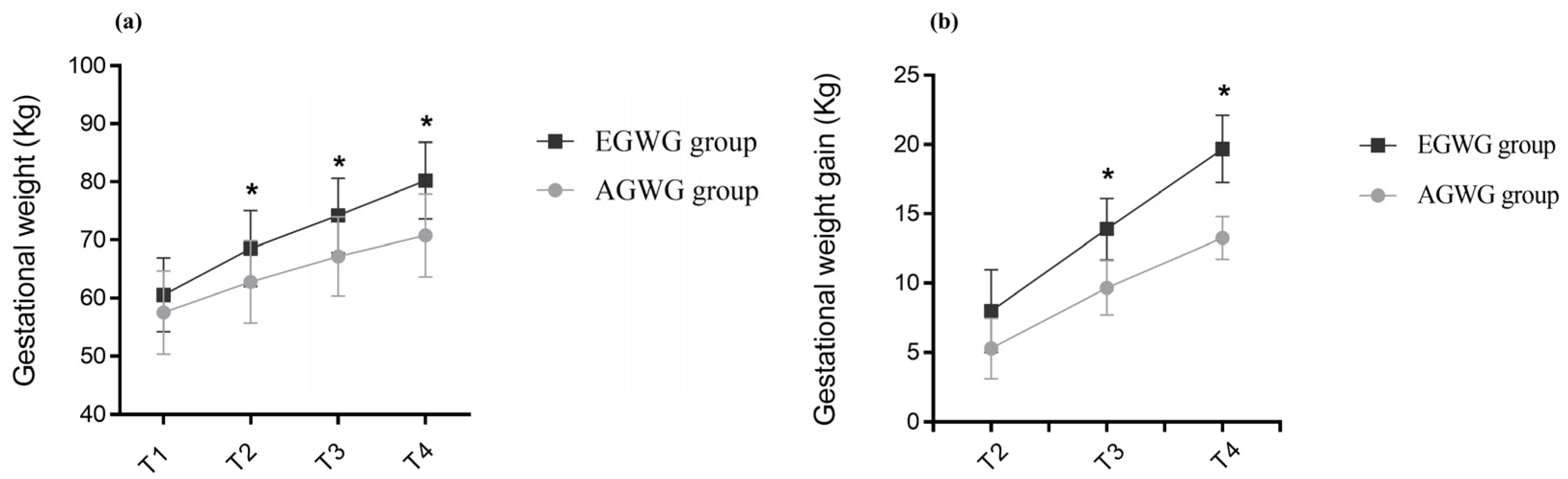
| T1 Pre-pregnancy weight (kg) | 60.54 | 6.37 | 57.52 | 7.17 | 0.682 |
| T1 Height (cm) | 163.71 | 6.56 | 161.39 | 6.93 | 0.785 |
| T1 Pre-pregnancy BMI (kg/m2) | 22.68 | 1.74 | 22.06 | 1.79 | 0.405 |
| T4 BMI (kg/m2) | 29.90 | 2.01 | 27.15 | 1.84 | 0.010 |
| T4 Total gestational weight gain (kg) | 19.60 | 2.43 | 13.26 | 1.54 | 0.010 |
| Pre-pregnancy fat mass (%) | 29.32 | 4.03 | 26.80 | 4.85 | 0.262 |
| Pre-pregnancy fat mass (kg) | 18.75 | 4.19 | 15.82 | 4.40 | 0.167 |
| Pre-pregnancy fat-free body mass (kg) | 44.17 | 4.60 | 42.26 | 3.85 | 0.188 |
| Pre-pregnancy muscle mass (kg) | 41.94 | 4.35 | 40.04 | 3.58 | 0.135 |
| Foetuses | Mean | SD | Mean | SD | |
| T2 Foetal weight (g) | 629.30 | 204.02 | 598.87 | 186.40 | 0.875 |
| T3 Foetal weight (g) | 2172.82 | 353.43 | 2132.18 | 457.47 | 0.514 |
| T2 SCFT (mm) | 2.84 | 0.52 | 2.95 | 0.56 | 0.721 |
| T3 SCFT (mm) | 4.13 | 0.76 | 4.07 | 1.07 | 0.521 |
| T2 Total thigh tissue (cm3) | 5.23 | 1,78 | 5.12 | 1.53 | 0.945 |
| T3 Total thigh tissue (cm3) | 13.37 | 2.97 | 13.53 | 3.25 | 0.955 |
| T2 Thigh muscle mass (cm3) | 2.97 | 1.04 | 2.90 | 0.93 | 0.729 |
| T3 Thigh muscle mass (cm3) | 7.69 | 1.72 | 7.54 | 1.80 | 0.643 |
| T2 Subcutaneous thigh fat (cm3) | 2.26 | 0.84 | 2.27 | 0.76 | 0.991 |
| T3 Subcutaneous thigh fat (cm3) | 5.68 | 1.64 | 6.03 | 1.61 | 0.225 |
| T2 Total arm tissue (cm3) | 3.05 | 0.93 | 2.85 | 0.83 | 0.358 |
| T3 Total arm tissue (cm3) | 7.01 | 1.55 | 7.07 | 1.87 | 0.860 |
| T2 Arm muscle mass (cm3) | 1.57 | 0.50 | 1.46 | 0.46 | 0.224 |
| T3 Arm muscle mass (cm3) | 3.45 | 0.82 | 3.52 | 0.98 | 0.683 |
| T2 Subcutaneous arm fat (cm3) | 1.46 | 0.52 | 1.46 | 0.63 | 0.991 |
| T3 Subcutaneous arm fat (cm3) | 3.55 | 0.93 | 3.56 | 1.02 | 0.928 |
| Neonates | Mean | SD | Mean | SD | |
| T5 Weight (g) | 3354.87 | 298.47 | 3068.50 | 386.57 | 0.027 |
| T5 Length (cm) | 50.03 | 1.78 | 48.80 | 2.33 | 0.182 |
| T5 Fat-free mass percentage (%) | 90.39 | 3.98 | 91.57 | 5.65 | 0.120 |
| T5 Fat mass percentage (%) | 9.61 | 3.98 | 8.43 | 5.65 | 0.120 |
| T5 Fat-free mass (kg) | 3.08 | 0.19 | 2.76 | 0.27 | 0.218 |
| T5 Fat mass (kg) | 0.34 | 0.13 | 0.26 | 0.21 | 0.039 |
| Terms Name | Binom Raw p-Value | Binom Fold Enrichment |
|---|---|---|
| Transient neonatal diabetes mellitus | 0.0010 | 1041.1 |
| Neonatal insulin-dependent diabetes mellitus | 0.0015 | 656.2 |
| Severe failure to thrive | 0.0037 | 269.7 |
| Insulin-resistant diabetes mellitus | 0.0185 | 53.7 |
| Insulin resistance | 0.0236 | 42.0 |
| Breast carcinoma | 0.0255 | 38.7 |
| Neoplasm of the breast | 0.0272 | 36.4 |
| Hyperglycemia | 0.0275 | 35.9 |
| Dehydration | 0.0379 | 25.9 |
| T2 Total thigh tissue | Β | r² | p | 95% CI |
| DMR 2 | 9.172 | 0.853 | 0.014 | 2.340; 16.005 |
| Gestational weight gain | −0.010 | 0.843 | −0.127; 0.106 | |
| Pre-pregnancy BMI | −0.780 | 0.005 | −1.249; −0.310 | |
| Maternal age | −0.048 | 0.400 | −0.172; 0.076 | |
| Sex | 1.026 | 0.092 | −0.205; 2.257 | |
| Gestational age | 0.833 | 0.002 | 0.411; 1.255 | |
| DMR 6 | 21.516 | 0.820 | 0.039 | 1.407; 41.625 |
| Gestational weight gain | −0.072 | 0.322 | −0.228; 0.084 | |
| Pre-pregnancy BMI | −0.786 | 0.008 | −1.316; −0.256 | |
| Maternal age | −0.044 | 0.489 | −0.181; 0.093 | |
| Sex | 1.313 | 0.070 | −0.131; 2.757 | |
| Gestational age | 0.939 | 0.002 | 0.443; 1.434 | |
| T3 Total thigh tissue | Β | r² | p | 95% CI |
| DMR 2 | 8.265 | 0.715 | 0.018 | 1.790; 14.740 |
| Gestational weight gain | 0.045 | 0.371 | −0.064; 0.154 | |
| Pre-pregnancy BMI | −0.393 | 0.080 | −0.844; 0.058 | |
| Maternal age | 0.036 | 0.487 | −0.077; 0.150 | |
| Sex | 0.679 | 0.127 | −0.235; 1.593 | |
| Gestational age | 0.472 | 0.155 | −0215; 1.160 | |
| T2 Thigh muscle mass | Β | r² | p | 95% CI |
| DMR 2 | 5.314 | 0.814 | 0.021 | 1.006; 9.622 |
| Gestational weight gain | −0.026 | 0.440 | −0.100; 0.047 | |
| Pre-pregnancy BMI | −0.416 | 0.011 | −0.712; −0.120 | |
| Maternal age | −0.012 | 0.739 | −0.090; 0.066 | |
| Sex | 0.773 | 0.051 | −0.003; 1.549 | |
| Gestational age | 0.431 | 0.005 | 0.165; 0.697 | |
| T3 Thigh muscle mass | Β | r² | p | 95% CI |
| DMR 2 | 6.373 | 0.687 | 0.032 | 0.694; 12.052 |
| Gestational weight gain | 0.067 | 0.147 | −0.029; 0.162 | |
| Pre-pregnancy BMI | −0.177 | 0.339 | −0.572; 0.219 | |
| Maternal age | 0.036 | 0.429 | −0.063; 0.136 | |
| Sex | 0.442 | 0.244 | −0.360; 1.243 | |
| Gestational age | 0.358 | 0.213 | −0.246; 0.961 | |
| T2 Subcutaneous thigh fat | Β | r² | p | 95% CI |
| DMR 2 | 3.858 | 0.846 | 0.029 | 0.506; 7.211 |
| Gestational weight gain | 0.016 | 0.549 | −0.041; 0.073 | |
| Pre-pregnancy BMI | −0.364 | 0.006 | −0.594; −0.133 | |
| Maternal age | −0.037 | 0.207 | −0.097; 0.024 | |
| Sex | 0.254 | 0.367 | −0.350; 0.858 | |
| Gestational age | 0.402 | 0.002 | 0.195; 0.609 | |
| DMR 6 | 10.933 | 0.862 | 0.017 | 2.494; 19.372 |
| Gestational weight gain | −0.019 | 0.532 | −0.084; 0.047 | |
| Pre-pregnancy BMI | −0.385 | 0.004 | −0.607; −1.162 | |
| Maternal age | −0.035 | 0.202 | −0.093; 0.023 | |
| Sex | 0.424 | 0.148 | −0.182; 1.030 | |
| Gestational age | 0.463 | 0.001 | 0.255; 0.671 | |
| T3 Subcutaneous thigh fat | Β | r² | p | 95% CI |
| DMR 10 | 7.604 | 0.596 | 0.033 | 0.763; 14.445 |
| Gestational weight gain | −0.034 | 0.267 | −0.100; 0.031 | |
| Pre-pregnancy BMI | −0.247 | 0.064 | −0.511; 0.018 | |
| Maternal age | 0.015 | 0.619 | −0.052; 0.083 | |
| Sex | 0.257 | 0.311 | −0.284; 0.797 | |
| Gestational age | −0.004 | 0.982 | −0.401; 0.393 | |
| T3 Total arm tissue | Β | r² | p | 95% CI |
| DMR 6 | −25.640 | 0.804 | 0.002 | −39.368; −11.911 |
| Gestational weight gain | 0.115 | 0.039 | 0.007; 0.222 | |
| Pre-pregnancy BMI | 0.410 | 0.043 | 0.016; 0.805 | |
| Maternal age | 0.038 | 0.400 | −0.059; 0.134 | |
| Sex | −0.269 | 0.460 | −1.059; 0.521 | |
| Gestational age | 0.311 | 0.257 | −0.271; 0.893 | |
| T3 Subcutaneous arm fat | Β | r² | p | 95% CI |
| DMR 6 | −17.433 | 0.667 | 0.010 | −29.597; −5.270 |
| Gestational weight gain | 0.078 | 0.097 | −0.017; 0.172 | |
| Pre-pregnancy BMI | 0.233 | 0.165 | −0.116; 0.583 | |
| Maternal age | 0.014 | 0.716 | −0.071; 0.099 | |
| Sex | −0.339 | 0.302 | −1.039; 0.361 | |
| Gestational age | 0.175 | 0.461 | −0.340; 0.691 | |
| T5 Fat mass percentage | Β | r² | p | 95% CI |
| DMR 2 | −20.299 | 0.761 | 0.039 | −39.362; −1.236 |
| Gestational weight gain | 0.500 | 0.013 | 0.135; 0.865 | |
| Pre-pregnancy BMI | 0.651 | 0.275 | −0.615; 1.916 | |
| Maternal age | 0.071 | 0.530 | −0.175; 0.318 | |
| Sex | −5.207 | 0.002 | −7.968; −2.446 | |
| Gestational age | −0.653 | 0.196 | −1.710; 0.403 | |
| T5 Fat mass | Β | r² | p | 95% CI |
| DMR 2 | −0.719 | 0.780 | 0.040 | −0.395; −0.042 |
| Gestational weight gain | 0.019 | 0.009 | 0.006; 0.032 | |
| Pre-pregnancy BMI | 0.026 | 0.216 | −0.018; 0.071 | |
| Maternal age | 0.003 | 0.391 | −0.005; 0.012 | |
| Sex | −0.180 | 0.002 | −0.279; 0.082 | |
| Gestational age | −0.016 | 0.374 | −0.053; 0.022 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Argentato, P.P.; Guerra, J.V.d.S.; Luzia, L.A.; Ramos, E.S.; Maschietto, M.; Rondó, P.H.d.C. Excessive Gestational Weight Gain Alters DNA Methylation and Influences Foetal and Neonatal Body Composition. Epigenomes 2023, 7, 18. https://doi.org/10.3390/epigenomes7030018
Argentato PP, Guerra JVdS, Luzia LA, Ramos ES, Maschietto M, Rondó PHdC. Excessive Gestational Weight Gain Alters DNA Methylation and Influences Foetal and Neonatal Body Composition. Epigenomes. 2023; 7(3):18. https://doi.org/10.3390/epigenomes7030018
Chicago/Turabian StyleArgentato, Perla Pizzi, João Victor da Silva Guerra, Liania Alves Luzia, Ester Silveira Ramos, Mariana Maschietto, and Patrícia Helen de Carvalho Rondó. 2023. "Excessive Gestational Weight Gain Alters DNA Methylation and Influences Foetal and Neonatal Body Composition" Epigenomes 7, no. 3: 18. https://doi.org/10.3390/epigenomes7030018