
High-Intensity Interval Cycling and Running Yield a Similar Myokine and Osteokine Response in Young Adult Females
 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Study Design and Procedures
2.3. Blood Collection and Biochemical Analysis
2.4. Statistical Analysis
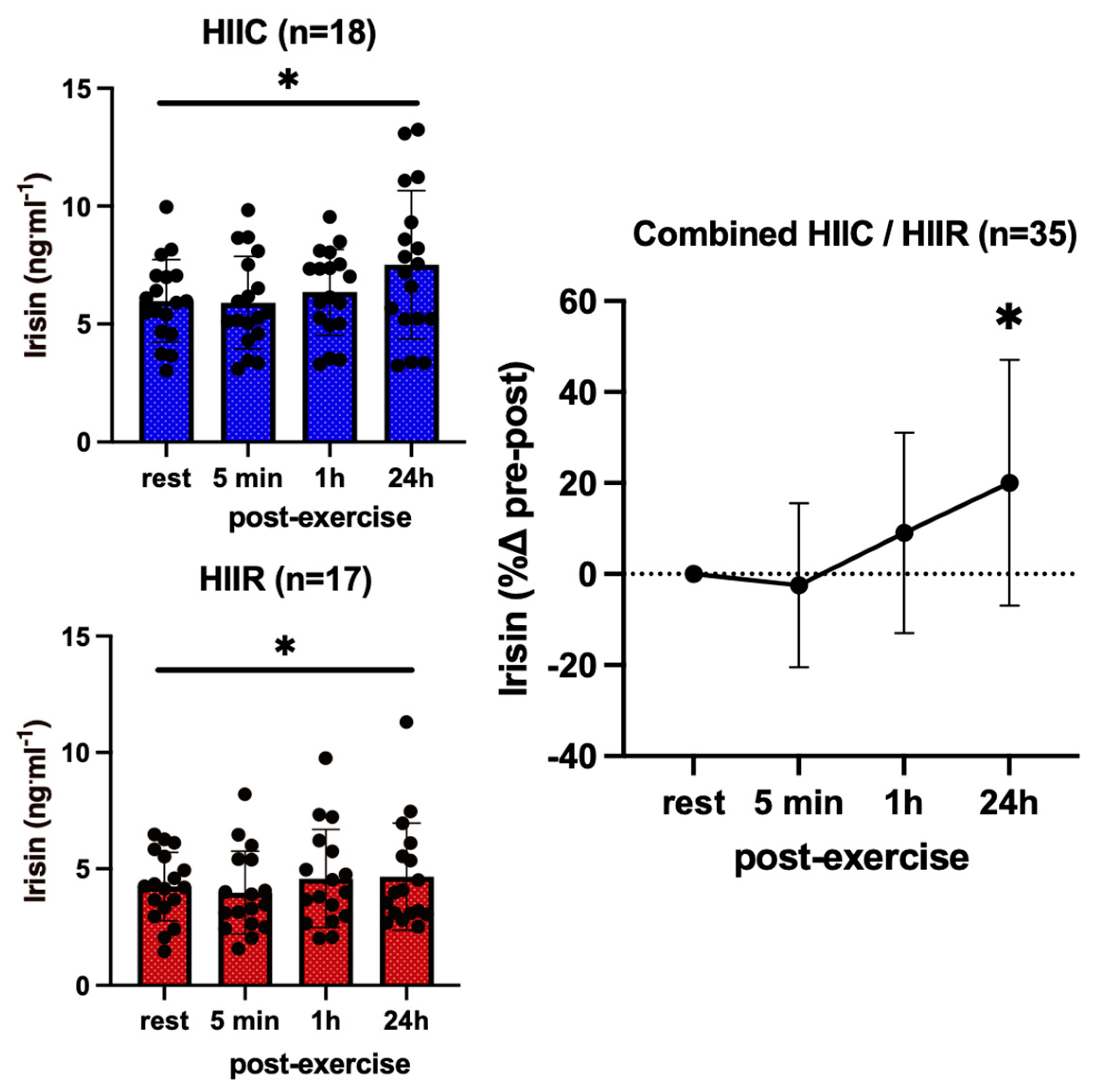
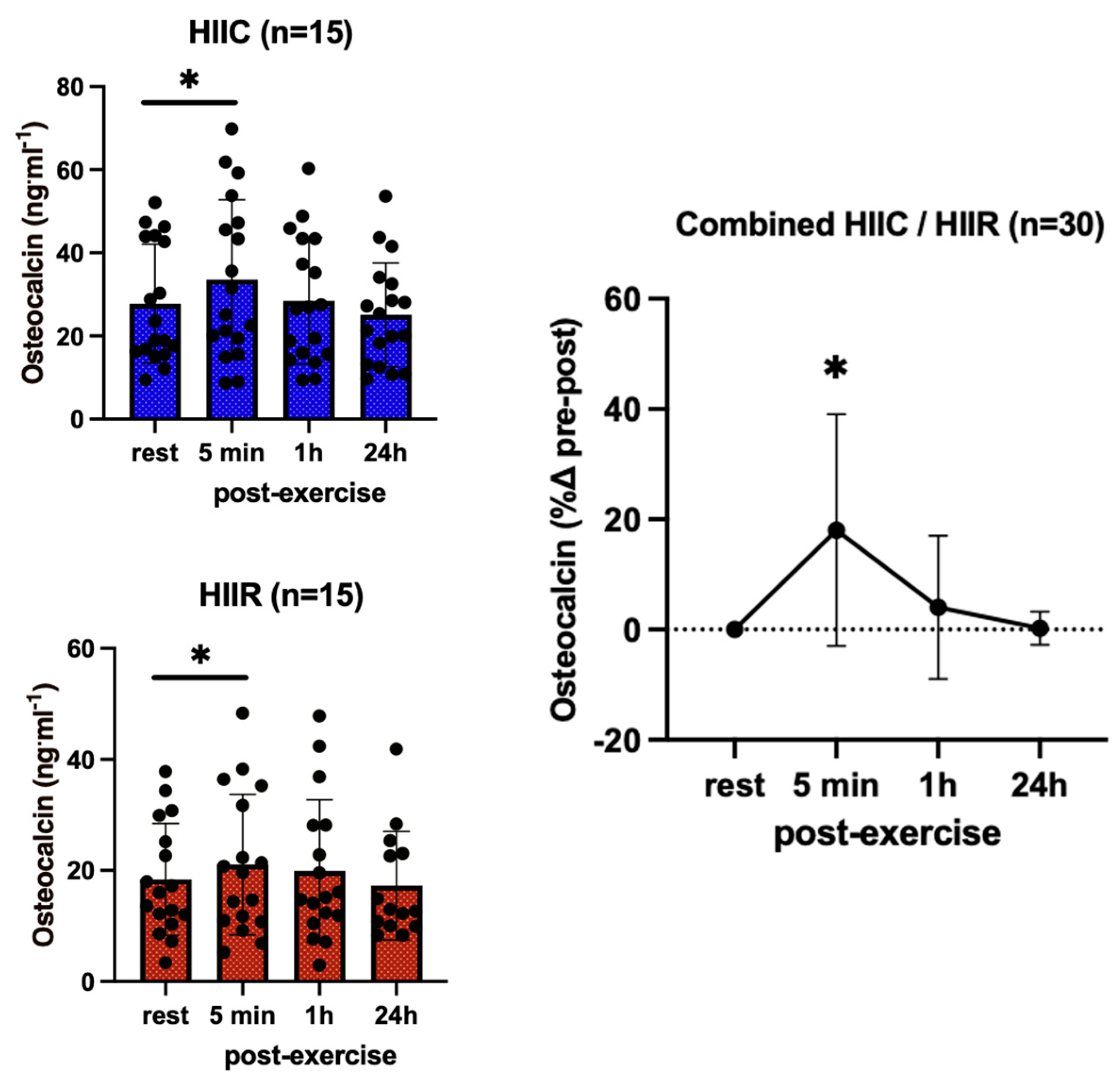
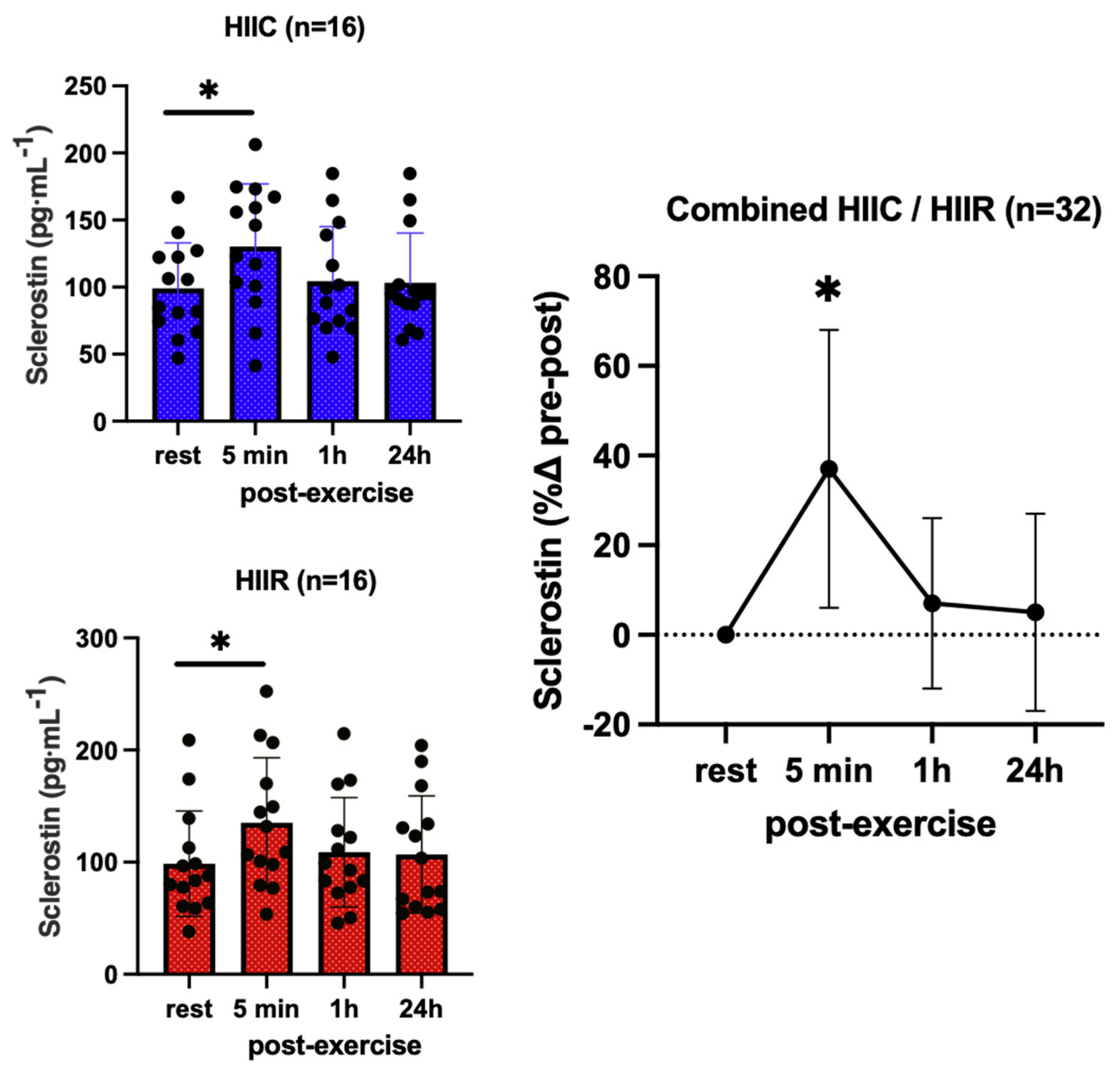
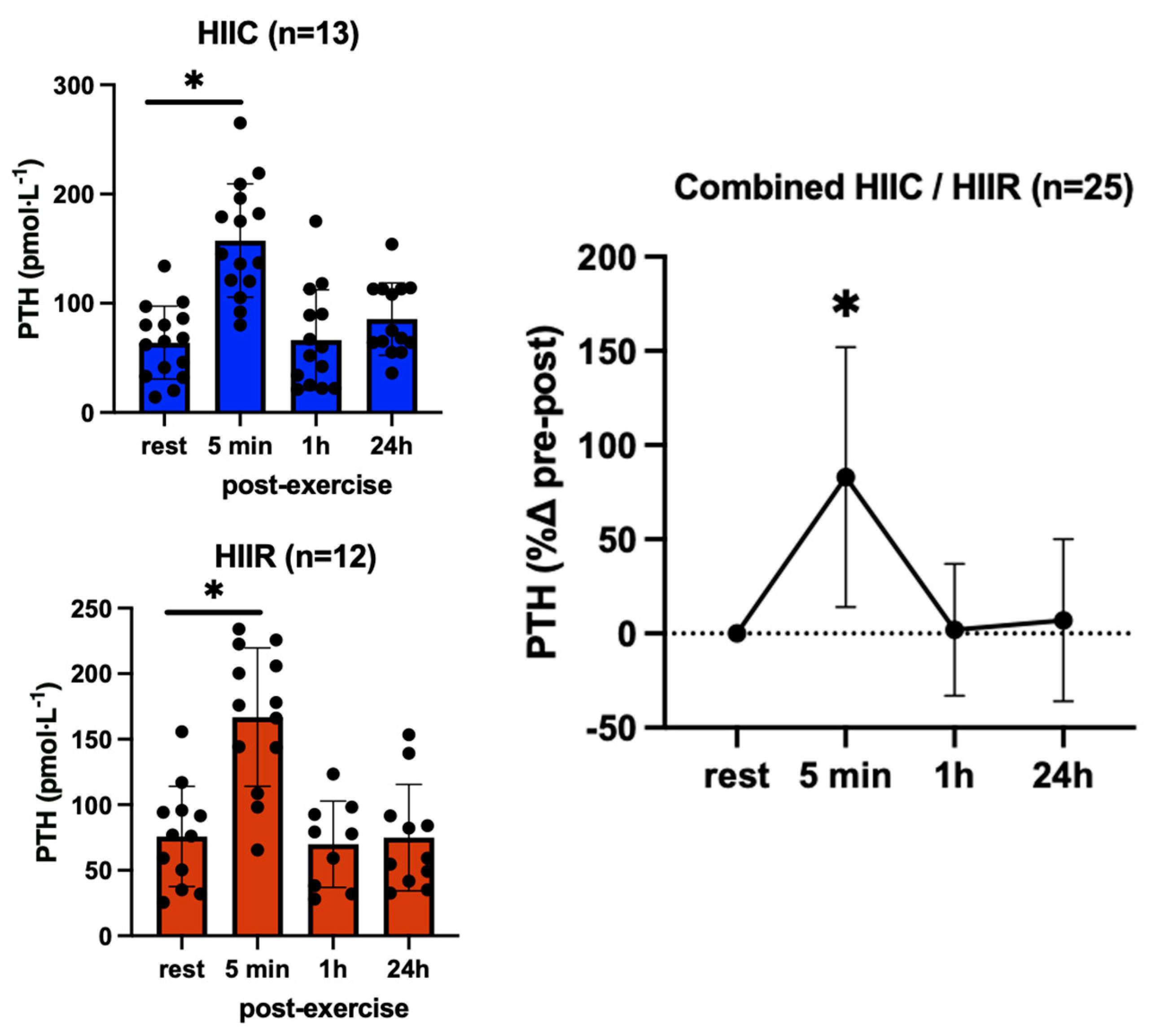
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Banfi, G.; Lombardi, G.; Colombini, A.; Lippi, G. Bone Metabolism Markers in Sports Medicine. J. Sports Med. 2012, 40, 697–714. [Google Scholar] [CrossRef]
- Lombardi, G.; Sanchis-Gomar, F.; Perego, S.; Sansoni, V.; Banfi, G. Implications of exercise-induced adipo-myokines in bone metabolism. Endocr. J. 2015, 54, 284–305. [Google Scholar] [CrossRef] [PubMed]
- Colaianni, G.; Cuscito, C.; Mongelli, T.; Pignataro, P.; Buccoliero, C.; Liu, P.; Lu, P.; Sartini, L.; Comite, M.; Mori, G.; et al. The myokine irisin increases cortical bone mass. Proc. Natl. Acad. Sci. USA 2015, 112, 12157–12162. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Valverde, P.; Zhu, X.; Murray, D.; Wu, Y.; Yu, L.; Jiang, H.; Dard, M.; Huang, J.; Xu, Z.; et al. Exercise-induced irisin in bone and systemic irisin administration reveal new regulatory mechanisms of bone metabolism. Bone Res. 2017, 5, 16056. [Google Scholar] [CrossRef]
- Kawao, N.; Moritake, A.; Tatsumi, K.; Kaji, H. Roles of Irisin in the Linkage from Muscle to Bone During Mechanical Unloading in Mice. Calcif. Tissue Int. 2018, 103, 24–34. [Google Scholar] [CrossRef]
- Li, G.; Cao, N.; Shang, P.; Xu, H. What the discovery of irisin receptor means to bone. Arch. Physiol. Biochem. 2020, 128, 1137–1139. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-W.; Seo, M.-W.; Jung, H.-C.; Song, J.-K. Effects of High-Impact Weight-Bearing Exercise on Bone Mineral Density and Bone Metabolism in Middle-Aged Premenopausal Women: A Randomized Controlled Trial. Appl. Sci. 2021, 11, 846. [Google Scholar] [CrossRef]
- Morseth, B.; Emaus, N.; Jørgensen, L. Physical activity and bone: The importance of the various mechanical stimuli for bone mineral density. A review. Nor. Epidemiol. 2011, 20, 173–178. [Google Scholar] [CrossRef]
- Lewis, R.D.; Modlesky, C.M. Nutrition, physical activity, and bone health in women. Int. J. Sport Nutr. Exerc. Metab. 1998, 8, 250–284. [Google Scholar] [CrossRef]
- Milgrom, C.; Finestone, A.; Levi, Y.; Simkin, A.; Ekenman, I.; Mendelson, S.; Millgram, M.; Nyska, M.; Benjuya, N.; Burr, D. Do high impact exercises produce higher tibial strains than running? Br. J. Sports Med. 2000, 34, 195–199. [Google Scholar] [CrossRef]
- Olmedillas, H.; González-Agüero, A.; Moreno, L.A.; Casajus, J.A.; Vicente-Rodríguez, G. Cycling and bone health: A systematic review. BMC Med. 2012, 10, 168. [Google Scholar] [CrossRef]
- Duncan, C.S.; Blimkie, C.J.R.; Kemp, A.; Higgs, W.; Cowell, C.T.; Woodhead, H.; Briody, J.N.; Howman-giles, R. Mid-femur geometry and biomechanical properties in 15- to 18-yr-old female athletes. Med. Sci. Sports Exerc. 2002, 34, 673–681. [Google Scholar] [PubMed]
- Torstveit, M.K.; Sundgot-Borgen, J. Low bone mineral density is two to three times more prevalent in non-athletic premenopausal women than in elite athletes: A comprehensive controlled study. Br. J. Sports Med. 2005, 39, 282–287. [Google Scholar] [CrossRef] [PubMed]
- Leigey, D.; Irrgang, J.; Francis, K.; Cohen, P.; Wright, V. Participation in High-Impact Sports Predicts Bone Mineral Density in Senior Olympic Athletes. Sports Health 2009, 1, 508–513. [Google Scholar] [CrossRef]
- Mezil, Y.A.; Allison, D.; Kish, K.; Ditor, D.; Ward, W.; Tsiani, E.; Klentrou, P. Response of Bone Turnover Markers and Cytokines to High-Intensity Low-Impact Exercise. Med. Sci. Sports Exerc. 2015, 47, 1495–1502. [Google Scholar] [CrossRef] [PubMed]
- Kouvelioti, R.; Kurgan, N.; Falk, B.; Ward, W.; Josse, A.R.; Klentrou, P. Response of Sclerostin and Bone Turnover Markers to High Intensity Interval Exercise in Young Women: Does Impact Matter? Biomed Res. Int. 2018, 2018, 4864952–4864958. [Google Scholar] [CrossRef]
- Kouvelioti, R.; Kurgan, N.; Falk, B.; Ward, W.; Josse, A.R.; Klentrou, P. Cytokine and Sclerostin Response to High-Intensity Interval Running versus Cycling. Med. Sci. Sports Exerc. 2019, 51, 2458–2464. [Google Scholar] [CrossRef]
- Kouvelioti, R.; LeBlanc, P.; Falk, B.; Ward, W.; Josse, A.R.; Klentrou, P. Effects of High-Intensity Interval Running Versus Cycling on Sclerostin, and Markers of Bone Turnover and Oxidative Stress in Young Men. Calcif. Tissue Int. 2019, 104, 582–590. [Google Scholar] [CrossRef] [PubMed]
- Nikel, O.; Poundarik, A.A.; Bailey, S.; Vashishth, D. Structural role of osteocalcin and osteopontin in energy dissipation in bone. J. Biomech. 2018, 80, 45–52. [Google Scholar] [CrossRef]
- Gassel, L.C.; Schneider, S.; Banke, I.J.; Braun, K.F.; Volkering, C.; Zeeb, L.; Burgkart, R.H.H.; von Eisenhart-Rothe, R.; Biberthaler, P.; van Griensven, M.; et al. Dysregulation of Wnt signaling in bone of type 2 diabetes mellitus and diabetic Charcot arthropathy. BMC Musculoskelet. Disord. 2022, 23, 365. [Google Scholar] [CrossRef]
- Hiam, D.; Landen, S.; Jacques, M.; Voisin, S.; Alvarez-Romero, J.; Byrnes, E.; Chubb, P.; Levinger, I.; Eynon, N. Osteocalcin and its forms respond similarly to exercise in males and females. Bone 2012, 144, 115818. [Google Scholar] [CrossRef]
- Dror, N.; Carbone, J.; Haddad, F.; Falk, B.; Klentrou, P.; Radom-Aizik, S. Sclerostin and bone turnover markers response to cycling and running at the same moderate-to-vigorous exercise intensity in healthy men. J. Endocrinol. Investig. 2021, 45, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Chen, X.; Zhang, L.; Wu, J.; Guo, J.; Zou, D.; Chen, B.; Sun, Z.; Shen, C.; Zou, J. The roles of exercise in bone remodeling and in prevention and treatment of osteoporosis. Prog. Biophys. Mol. Biol. 2016, 122, 122–130. [Google Scholar] [CrossRef]
- Ziegler, S.; Niessner, A.; Richter, B.; Wirth, S.; Billensteiner, E.; Woloszczuk, W.; Slany, J.; Geyer, G. Endurance running acutely raises plasma osteoprotegerin and lowers plasma receptor activator of nuclear factor κ B ligand. Metab. Clin. Exp. 2005, 54, 935–938. [Google Scholar] [CrossRef] [PubMed]
- Gaudio, A.; Rapisarda, R.; Xourafa, A.; Zanoli, L.; Manfrè, V.; Catalano, A.; Signorelli, S.S.; Castellino, P. Effects of competitive physical activity on serum irisin levels and bone turnover markers. J. Endocrinol. Investig. 2021, 44, 2235–2241. [Google Scholar] [CrossRef]
- Baron, R.; Kneissel, M. WNT signaling in bone homeostasis and disease: From human mutations to treatments. Nat. Med. 2013, 19, 179–192. [Google Scholar] [CrossRef]
- Holdsworth, G.; Roberts, S.J.; Ke, H.Z. Novel actions of sclerostin on bone. J. Mol. Endocrinol. 2019, 62, R167–R185. [Google Scholar] [CrossRef]
- Cameron, D.A.; Paschall, H.A.; Robinson, R.A. Changes in the Fine Structure of Bone Cells after the Administration of Parathyroid Extract. J. Mol. Cell Biol. 1967, 33, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Bellido, T.; Saini, V.; Pajevic, P.D. Effects of PTH on osteocyte function. Bone 2012, 54, 250–257. [Google Scholar] [CrossRef]
- Silva, B.C.; Bilezikian, J.P. Parathyroid hormone: Anabolic and catabolic actions on the skeleton. Curr. Opin. Pharmacol. 2015, 22, 41–50. [Google Scholar] [CrossRef]
- Kim, S.W.; Pajevic, P.D.; Selig, M.; Barry, K.J.; Yang, J.-Y.; Shin, C.S.; Baek, W.-Y.; Kim, J.-E.; Kronenberg, H.M. Intermittent parathyroid hormone administration converts quiescent lining cells to active osteoblasts. J. Bone Miner. 2012, 27, 2075–2084. [Google Scholar] [CrossRef] [PubMed]
- Falk, B.; Haddad, F.; Klentrou, P.; Ward, W.; Kish, K.; Mezil, Y.; Radom-Aizik, S. Differential sclerostin and parathyroid hormone response to exercise in boys and men. Osteoporos. Int. 2015, 27, 1245–1249. [Google Scholar] [CrossRef] [PubMed]
- Godin, G.; Shephard, R.J. A simple method to assess exercise behavior in the community. Can. J. Appl. Sci. 1985, 10, 141–146. [Google Scholar]
- Guzman, A.; Kurgan, N.; Moniz, S.C.; McCarthy, S.F.; Sale, C.; Heather Logan-Spenger Elliot-Sale, K.; Hazell, T.J.; Klentrou, P. Menstrual cycle related fluctuations in circulating markers of bone metabolism at rest and in response to running in eumenorrheic females. Calcif. Tissue Int. 2022, 111, 124–136. [Google Scholar] [CrossRef]
- Kargotich, S.; Goodman, C.; Keast, D.; Morton, A.R. The Influence of Exercise-Induced Plasma Volume Changes on the Interpretation of Biochemical Parameters Used for Monitoring Exercise, Training and Sport. J. Sports Med. 1998, 26, 101–117. [Google Scholar] [CrossRef]
- van Beaumont, W. Evaluation of hemoconcentration from hematocrit measurements. J. Appl. Physiol. 1972, 32, 712–713. [Google Scholar] [CrossRef]
- Lachenbruch, P.A.; Cohen, J. Statistical Power Analysis for the Behavioral Sciences. J. Am. Stat. Assoc. 1989, 84, 1096. [Google Scholar] [CrossRef]
- Lagzdina, R.; Rumaka, M.; Gersone, G.; Tretjakovs, P. Circulating Irisin in Healthy Adults: Changes after Acute Exercise, Correlation with Body Composition, and Energy Expenditure Parameters in Cross-Sectional Study. Medicina 2020, 56, 274. [Google Scholar] [CrossRef]
- Winn, N.C.; Grunewald, Z.I.; Liu, Y.; Heden, T.D.; Nyhoff, L.M.; Kanaley, J.A. Plasma Irisin Modestly Increases during Moderate and High-Intensity Afternoon Exercise in Obese Females. PLoS ONE 2017, 12, e0170690. [Google Scholar] [CrossRef]
- Tsuchiya, Y.; Daisuke, A.; Kazushige, G.; Masataka, K.; Mitsuya, Y. High-Intensity Exercise Causes Greater Irisin Response Compared with Low-Intensity Exercise under Similar Energy Consumption. Tohoku J. Exp. Med. 2014, 233, 135–140. [Google Scholar] [CrossRef]
- Qiu, S.; Bosnyák, E.; Treff, G.; Steinacker, J.M.; Nieß, A.M.; Krüger, K.; Mooren, F.C.; Zügel, M.; Schumann, U. Acute exercise-induced irisin release in healthy adults: Associations with training status and exercise mode. Eur. J. Sport Sci. 2018, 18, 1226–1233. [Google Scholar] [CrossRef]
- Kraemer, R.R.; Shockett, P.E.; Webb, N.D.; Shah, U.; Castracane, V.D. A Transient Elevated Irisin Blood Concentration in Response to Prolonged, Moderate Aerobic Exercise in Young Men and Women. Med. Sci. Sports Exerc. 2014, 46, 404. [Google Scholar] [CrossRef]
- Kim, H.; Wrann, C.D.; Jedrychowski, M.; Vidoni, S.; Kitase, Y.; Nagano, K.; Zhou, C.; Chou, J.; Parkman, V.-J.A.; Novick, S.J.; et al. Irisin Mediates Effects on Bone and Fat via αV Integrin Receptors. Cell 2018, 175, 1756–1768.e17. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Qiao, X.; Ma, Y.; Deng, H.; Xu, C.C.; Xu, L. Disordered Metabolism in Mice Lacking Irisin. Sci. Rep. 2020, 10, 17368. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Li, X.; Wang, X.; Chen, T.; Tao, F.; Liu, C.; Tu, Q.; Shen, G.; Chen, J.J. Irisin Deficiency Disturbs Bone Metabolism. J. Cell Physiol. 2021, 236, 664–676. [Google Scholar] [CrossRef]
- Falk, B.; Klentrou, P. Elevation in Sclerostin After Exercise: Is It Affected by Age and Sex? Calcif. Tissue Int. 2017, 102, 380–381. [Google Scholar] [CrossRef] [PubMed]
- Stunes, A.K.; Brobakken, C.L.; Sujan, M.A.J.; Aagård, N.; Brevig, M.S.; Wang, E.; Syversen, U.; Mosti, M.P. Acute Effects of Strength and Endurance Training on Bone Turnover Markers in Young Adults and Elderly Men. Front. Endocrinol. 2022, 13, 915241. [Google Scholar] [CrossRef]
- Morgan, A.L.; Weiss, J.; Kelley, E.T. Bone Turnover Response to Acute Exercise with Varying Impact Levels: A preliminary investigation. Int. J. Exerc. Sci. 2015, 8, 154–163. [Google Scholar]
- Troy, K.L.; Mancuso, M.E.; Butler, T.A.; Johnson, J.E. Exercise Early and Often: Effects of Physical Activity and Exercise on Women’s Bone Health. Int. J. Environ. Res. Public Health 2018, 15, 878. [Google Scholar] [CrossRef]
- Oniszczuk, A.; Kaczmarek, A.; Kaczmarek, M.; Ciałowicz, M.; Arslan, E.; Silva, A.F.; Murawska-Ciałowicz, E. Sclerostin as a biomarker of physical exercise in osteoporosis: A narrative review. Front. Endocrinol. 2022, 13, 954895. [Google Scholar] [CrossRef]
- Pickering, M.-E.; Simon, M.; Sornay-Rendu, E.; Chikh, K.; Carlier, M.-C.; Raby, A.-L.; Szulc, P.; Confavreux, C.B. Serum Sclerostin Increases After Acute Physical Activity. Calcif. Tissue Int. 2017, 101, 170–173. [Google Scholar] [CrossRef] [PubMed]
- Heinonen, I.; Kemppainen, J.; Kaskinoro, K.; Langberg, H.; Knuuti, J.; Boushel, R.; Kjaer, M.; Kalliokoski, K.K. Bone blood flow and metabolism in humans: Effect of muscular exercise and other physiological perturbations. J. Bone Miner. 2013, 28, 1068–1074. [Google Scholar] [CrossRef]
- Drüeke, T.B.; Massy, Z.A. Changing bone patterns with progression of chronic kidney disease. Kidney Int. 2016, 89, 289–302. [Google Scholar] [CrossRef] [PubMed]
- Oldknow, K.J.; MacRae, V.E.; Farquharson, C. Endocrine role of bone: Recent and emerging perspectives beyond osteocalcin. J. Endocrinol. 2015, 225, R1–R19. [Google Scholar] [CrossRef]
- Fulzele, K.; Lai, F.; Dedic, C.; Saini, V.; Uda, Y.; Shi, C.; Tuck, P.; Aronson, J.L.; Liu, X.; Spatz, J.M.; et al. Osteocyte-Secreted Wnt Signaling Inhibitor Sclerostin Contributes to Beige Adipogenesis in Peripheral Fat Depots. J. Bone Miner. 2017, 32, 373–384. [Google Scholar] [CrossRef]
- Kurgan, N.; Islam, H.; Matusiak, J.B.L.; Baranowski, B.J.; Stoikos, J.; Fajardo, V.A.; MacPherson, R.E.K.; Gurd, B.J.; Klentrou, P. Subcutaneous adipose tissue sclerostin is reduced and Wnt signaling is enhanced following 4-weeks of sprint interval training in young men with obesity. Physiol. Rep. 2022, 10, e15232. [Google Scholar] [CrossRef]
- Kohrt, W.M.; Wherry, S.J.; Wolfe, P.; Sherk, V.D.; Wellington, T.; Swanson, C.M.; Weaver, C.M.; Boxer, R.S. Maintenance of Serum Ionized Calcium During Exercise Attenuates Parathyroid Hormone and Bone Resorption Responses. J. Bone Miner. Res. 2018, 33, 1326–1334. [Google Scholar] [CrossRef] [PubMed]
- Townsend, R.; Elliott-Sale, K.J.; Pinto, A.J.; Thomas, C.; Scott, J.P.R.; Currell, K.; Fraser, W.D.; Sale, C. Parathyroid Hormone Secretion Is Controlled by Both Ionized Calcium and Phosphate During Exercise and Recovery in Men. J. Clin. Endocr. 2016, 101, 3231–3239. [Google Scholar] [CrossRef]
- Scott, J.P.R.; Sale, C.; Greeves, J.P.; Casey, A.; Dutton, J.; Fraser, W.D. The role of exercise intensity in the bone metabolic response to an acute bout of weight-bearing exercise. J. Appl. Physiol. 2011, 110, 423–432. [Google Scholar] [CrossRef]
- Śliwicka, E.; Cisoń, T.; Pilaczyńska-Szcześniak, Ł.; Ziemba, A.; Straburzyńska-Lupa, A. Effects of marathon race on selected myokines and sclerostin in middle-aged male amateur runners. Sci. Rep. 2021, 11, 2813. [Google Scholar] [CrossRef]
- Maïmoun, L.; Manetta, J.; Couret, I.; Dupuy, A.M.; Mariano-Goulart, D.; Micallef, J.P.; Peruchon, E.; Rossi, M. The Intensity Level of Physical Exercise and the Bone Metabolism Response. J. Appl. Physiol. 2016, 27, 105–111. [Google Scholar] [CrossRef]
- Guerriere, K.I.; Hughes, J.M.; Gaffney-Stomberg, E.; Staab, J.S.; Matheny, R.W. Circulating sclerostin is not suppressed following a single bout of exercise in young men. Physiol. Rep. 2018, 6, e13695. [Google Scholar] [CrossRef] [PubMed]
- Kemmler, W.; Shojaa, M.; Kohl, M.; von Stengel, S. Effects of different types of exercise on bone mineral density in postmenopausal women: A systematic review and meta-analysis. Calc. Tissue Int. 2020, 107, 409–439. [Google Scholar] [CrossRef] [PubMed]
- Onyia, J.E.; Helvering, L.M.; Gelbert, L.; Wei, T.; Huang, S.; Chen, P.; Dow, E.R.; Maran, A.; Zhang, M.; Lotinun, S.; et al. Molecular profile of catabolic versus anabolic treatment regimens of parathyroid hormone (PTH) in rat bone: An analysis by DNA microarray. J. Cell. Biochem. 2005, 95, 403–418. [Google Scholar] [CrossRef] [PubMed]
- Bouassida, A.; Latiri, I.; Bouassida, S.; Zalleg, D.; Zaouali, M.; Feki, Y.; Gharbi, N.; Zbidi, A.; Tabka, Z. Parathyroid hormone and physical exercise: A brief review. J. Sports Sci. Med. 2006, 5, 367–374. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | Trial | Pre-Exercise | Post-Exercise | ||
|---|---|---|---|---|---|
| 5 min | 1 h | 24 h | |||
| Irisin (ng·mL−1) | Cycling (N = 18) | 6.0 ± 1.7 # | 5.9 ± 2.0 # | 6.3 ± 1.8 # | 7.5 ± 3.1 #* |
| Running (N = 17) | 4.2 ± 1.5 # | 4.0 ± 1.8 # | 4.6 ± 2.1 # | 5.3 ± 2.3 #* | |
| Osteocalcin (ng·mL−1) | Cycling (N = 15) | 23.9 ± 12.2 | 29.4 ± 18.1 * | 24.4 ± 12.5 * | 20.8 ± 8.2 |
| Running (N = 15) | 22.7 ± 10.0 | 26.8 ± 16.7 * | 24.6 ± 11.4 * | 20.2 ± 8.7 | |
| Sclerostin (pg·mL−1) | Cycling (N = 18) | 99.1 ± 33.9 | 133.8 ± 54.2 * | 104.5 ± 40.5 | 103.1 ± 37.1 |
| Running (N = 18) | 98.6 ± 47.0 | 135.1 ± 58.0 * | 108.9 ± 48.8 | 106.7 ± 52.3 | |
| Parathyroid Hormone (pmol·L−1) | Cycling (N = 13) | 91.8 ± 66.4 | 178.5 ± 62.0 * | 94.7 ± 72.6 | 88.5 ± 58.7 |
| Running (N = 12) | 87.2 ± 58.6 | 166.8 ± 70.9 * | 88.2 ± 60.8 | 85.2 ± 66.5 | |
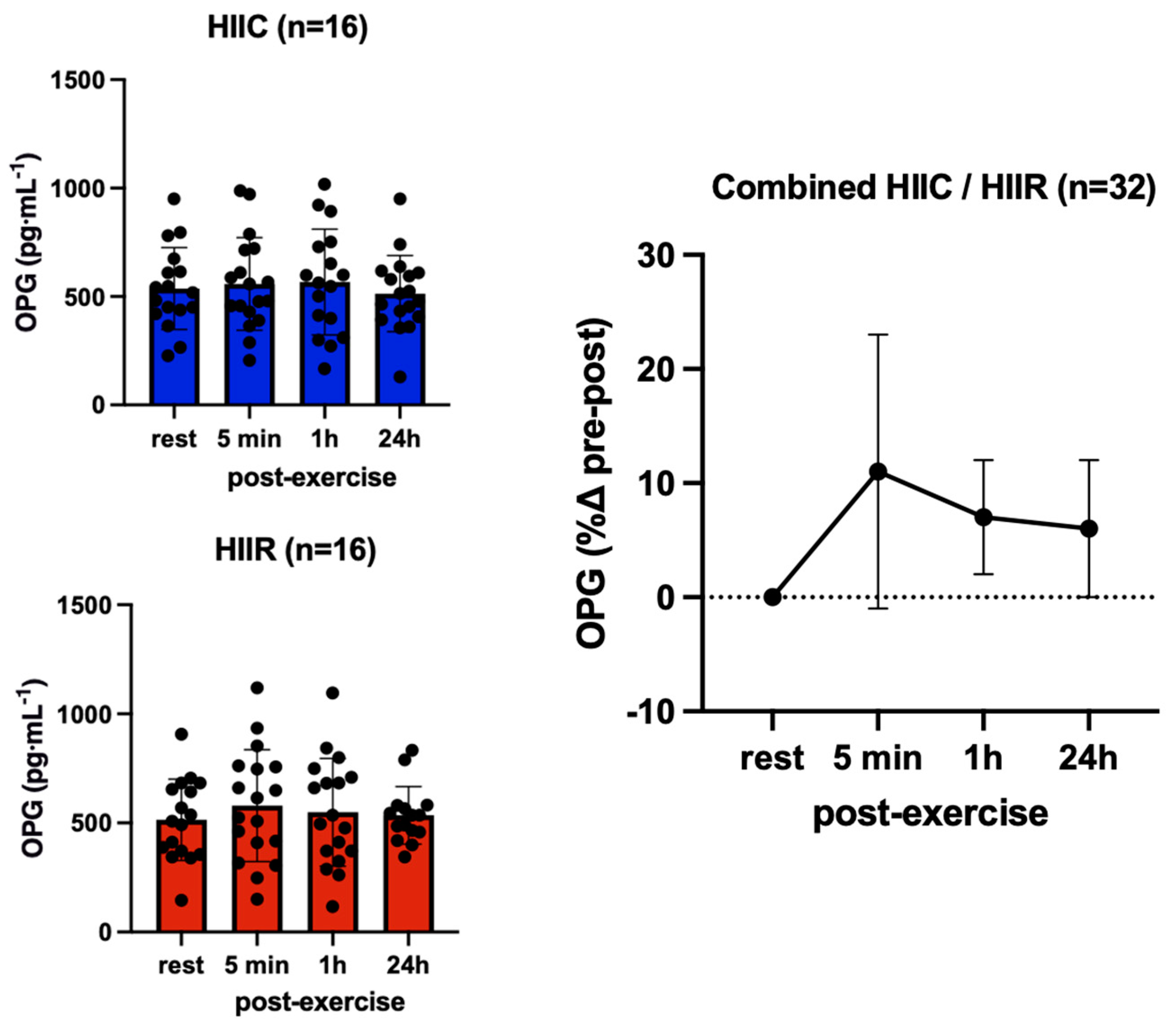
| Osteoprotegerin (pg·mL−1) | Cycling (N = 17) | 514.7 ± 206.1 | 540.2 ± 227.6 | 542.7 ± 257.3 | 515.3 ± 261.8 |
| Running (N = 17) | 508.2 ± 132.5 | 608.7 ± 227.5 | 601.5 ± 235.5 | 509.5 ± 218.6 | |
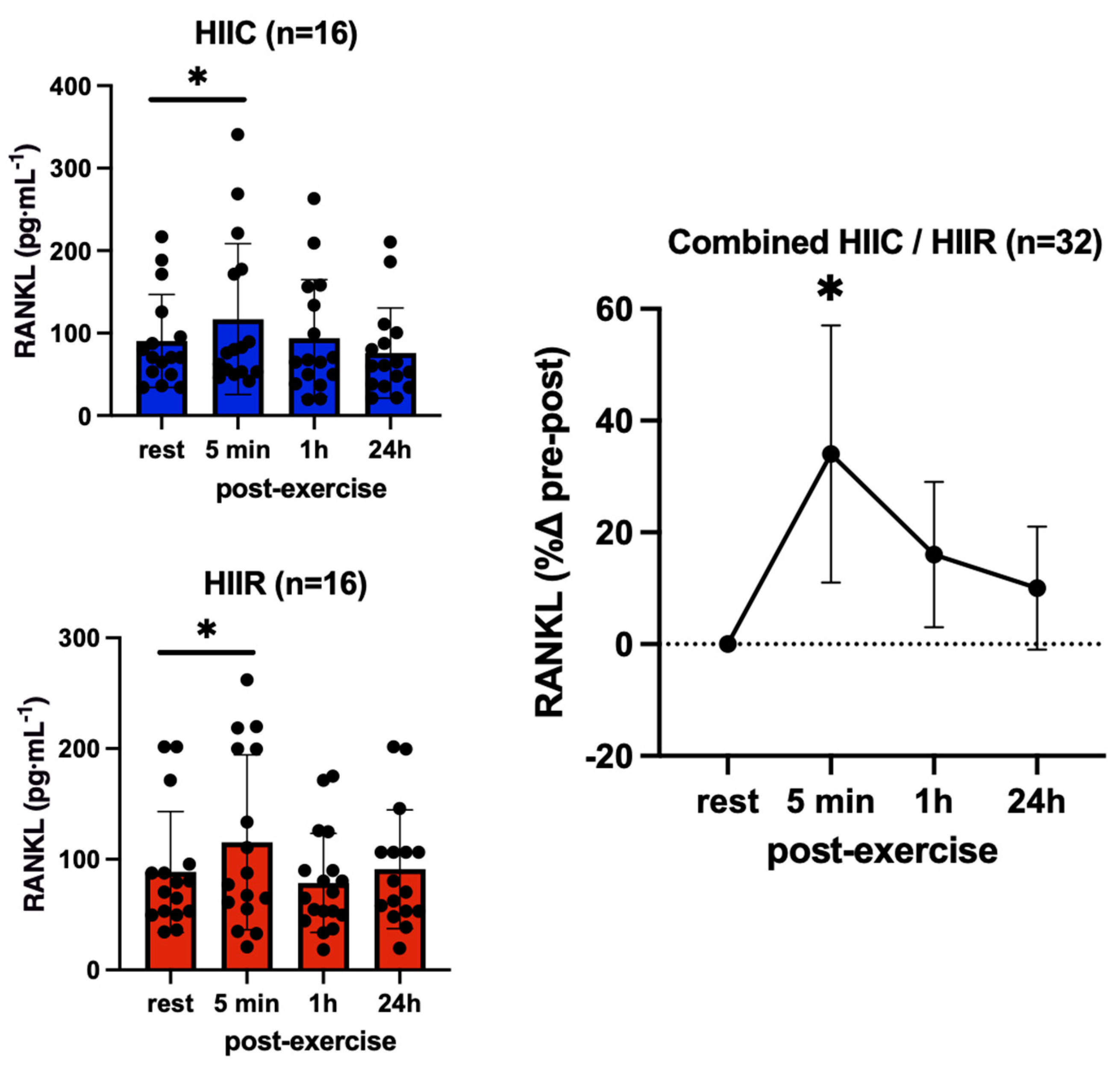
| Receptor Activator Nuclear Factor Kappa-β Ligand (pg·mL−1) | Cycling (N = 16) | 93.2 ± 57.7 | 121.1 ± 93.1 * | 96.7 ± 72.2 | 93.8 ± 73.9 |
| Running (N = 16) | 90.1 ± 56.2 | 122.3 ± 94.0 * | 95.6 ± 78.7 | 95.3 ± 54.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borzooeian, G.; Kottaras, S.; Kouvelioti, R.; Bell, M.; Ward, W.E.; Tsiani, E.; Klentrou, P. High-Intensity Interval Cycling and Running Yield a Similar Myokine and Osteokine Response in Young Adult Females. Endocrines 2023, 4, 312-326. https://doi.org/10.3390/endocrines4020025
Borzooeian G, Kottaras S, Kouvelioti R, Bell M, Ward WE, Tsiani E, Klentrou P. High-Intensity Interval Cycling and Running Yield a Similar Myokine and Osteokine Response in Young Adult Females. Endocrines. 2023; 4(2):312-326. https://doi.org/10.3390/endocrines4020025
Chicago/Turabian StyleBorzooeian, Giti, Steven Kottaras, Rozalia Kouvelioti, Madison Bell, Wendy E. Ward, Evangelia Tsiani, and Panagiota Klentrou. 2023. "High-Intensity Interval Cycling and Running Yield a Similar Myokine and Osteokine Response in Young Adult Females" Endocrines 4, no. 2: 312-326. https://doi.org/10.3390/endocrines4020025