Optimization of Corn Steep Liquor Dosage and Other Fermentation Parameters for Ethanol Production by Saccharomyces cerevisiae Type 1 and Anchor Instant Yeast


Abstract
:1. Introduction
2. Results and Discussion
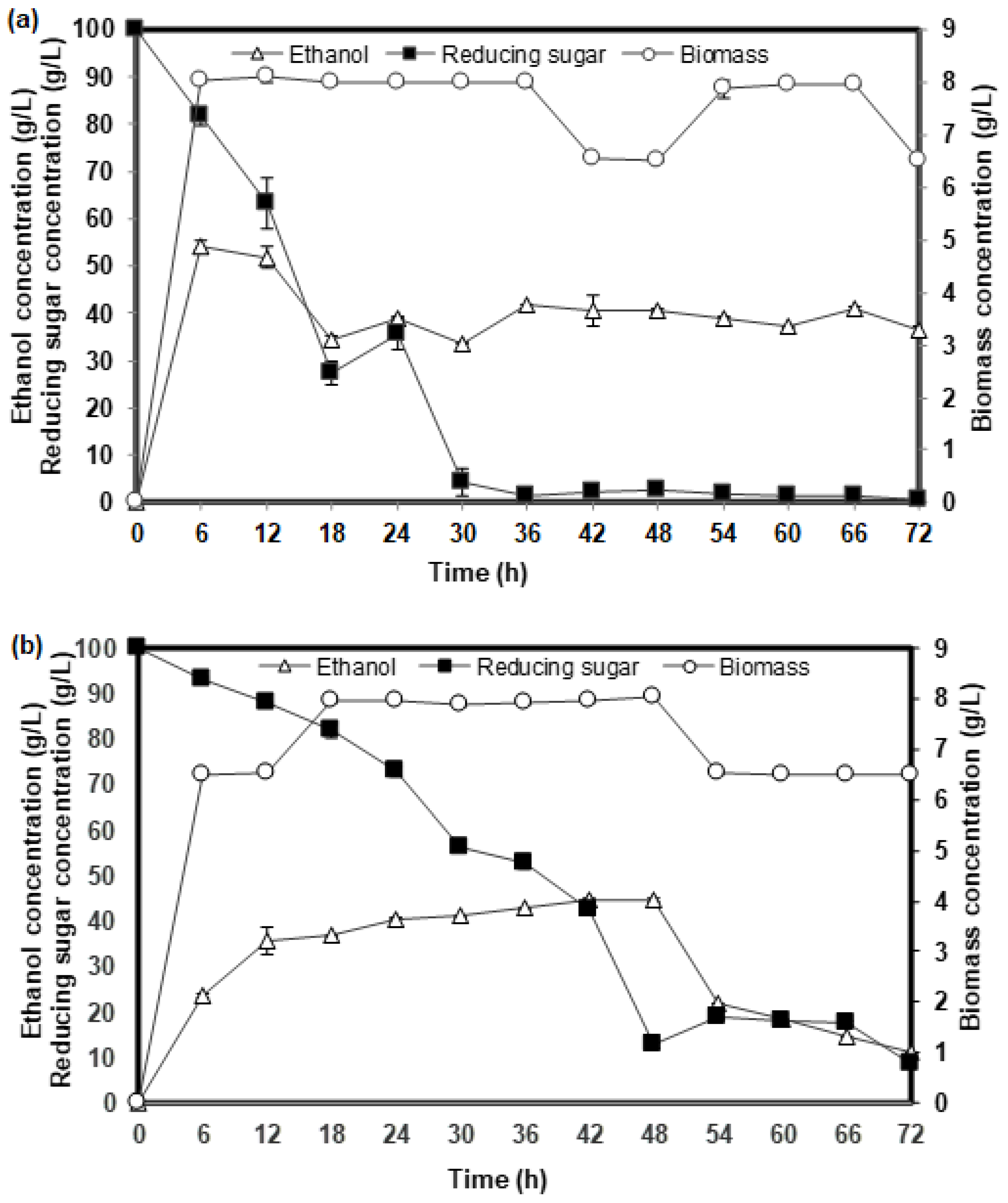
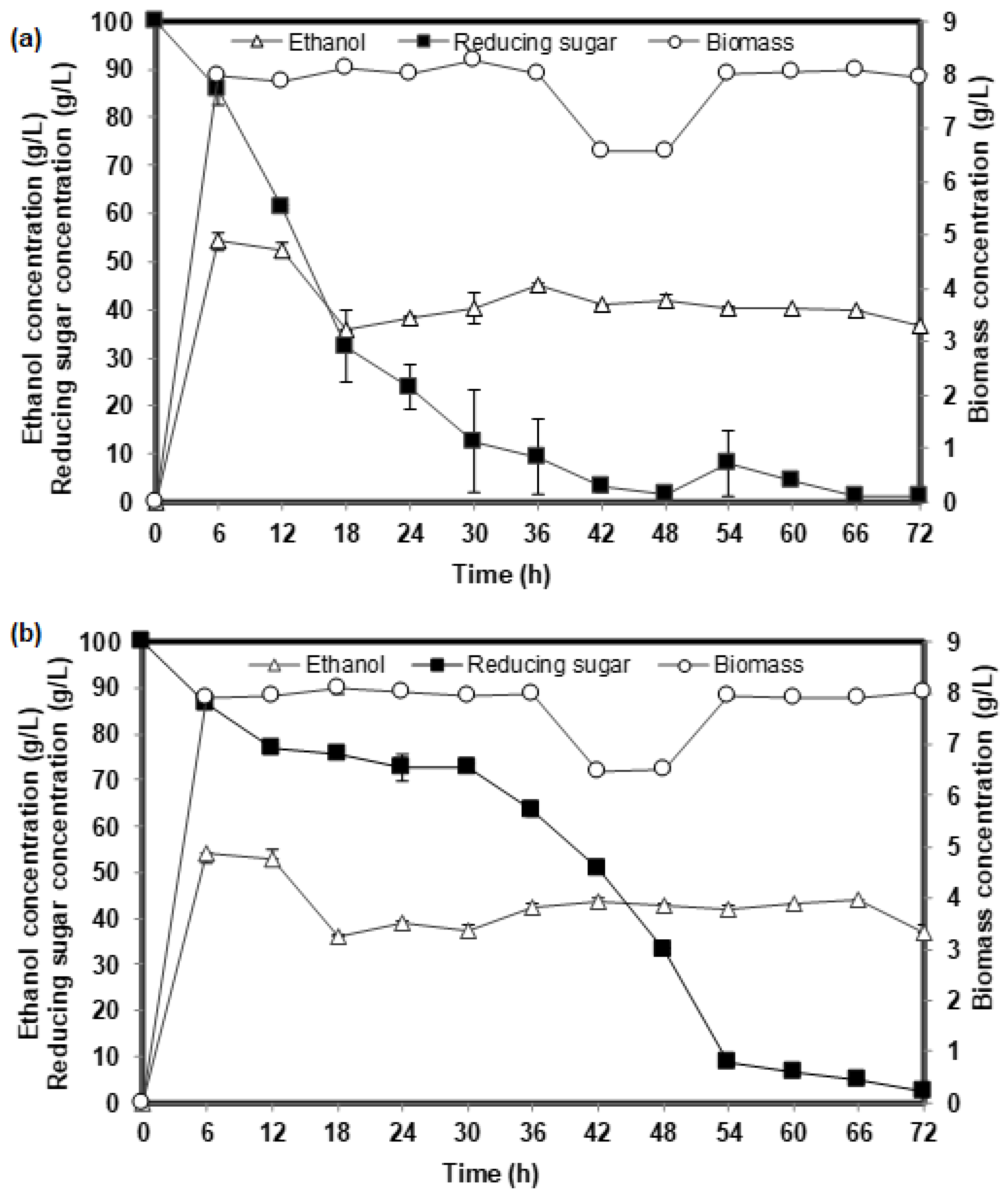
2.1. Comparison of the Fermentation Performance Based on Yeast Extract (YE) and Corn Steep Liquor (CSL) as Inoculum Component for Ethanol Fermentation of Two Yeast Strains
2.2. Ethanol Production from Glucose Fermentation Using CSL Inoculum: Optimization Studies
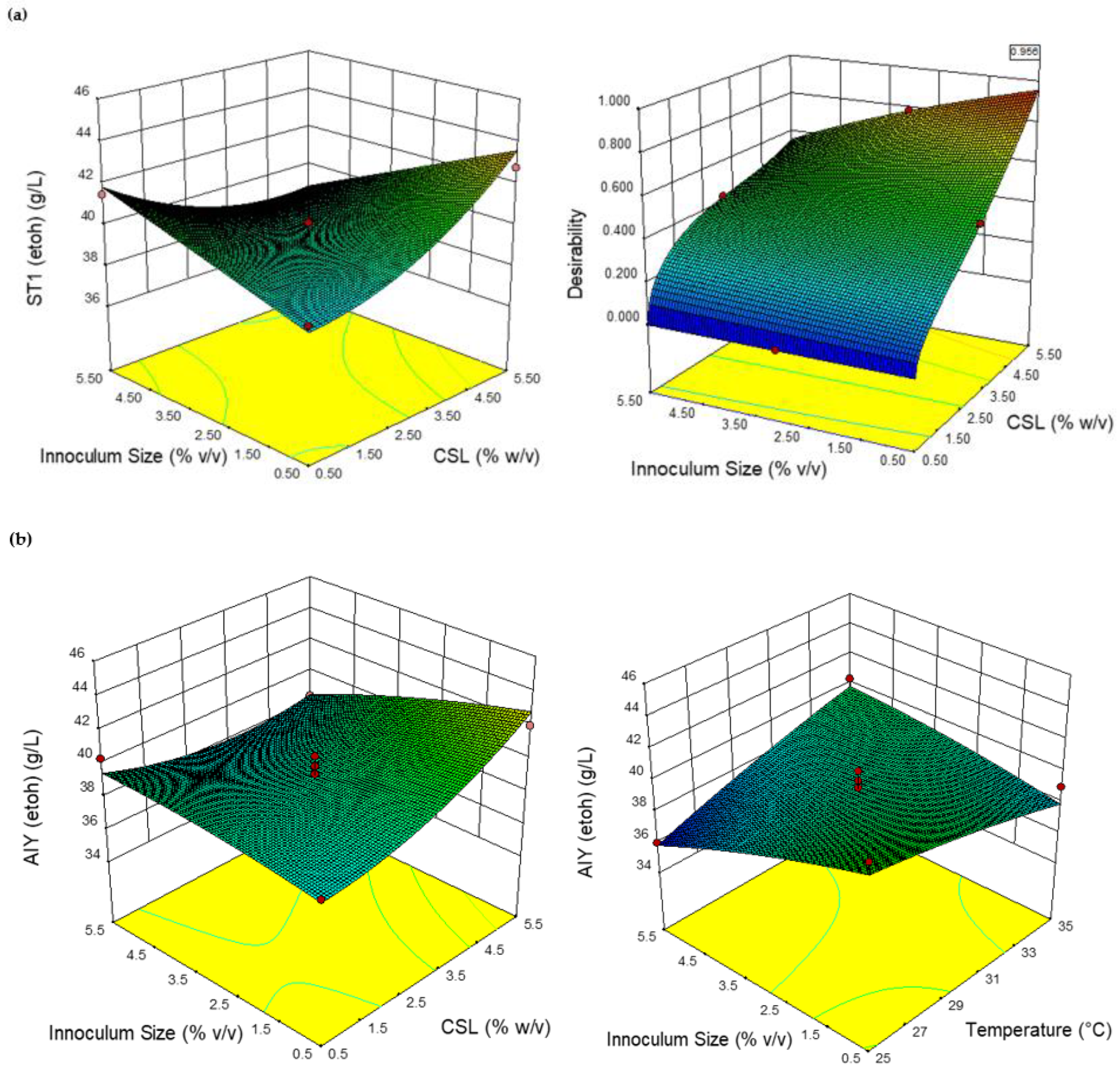
2.3. RSM Modeling Results for Ethanol Production
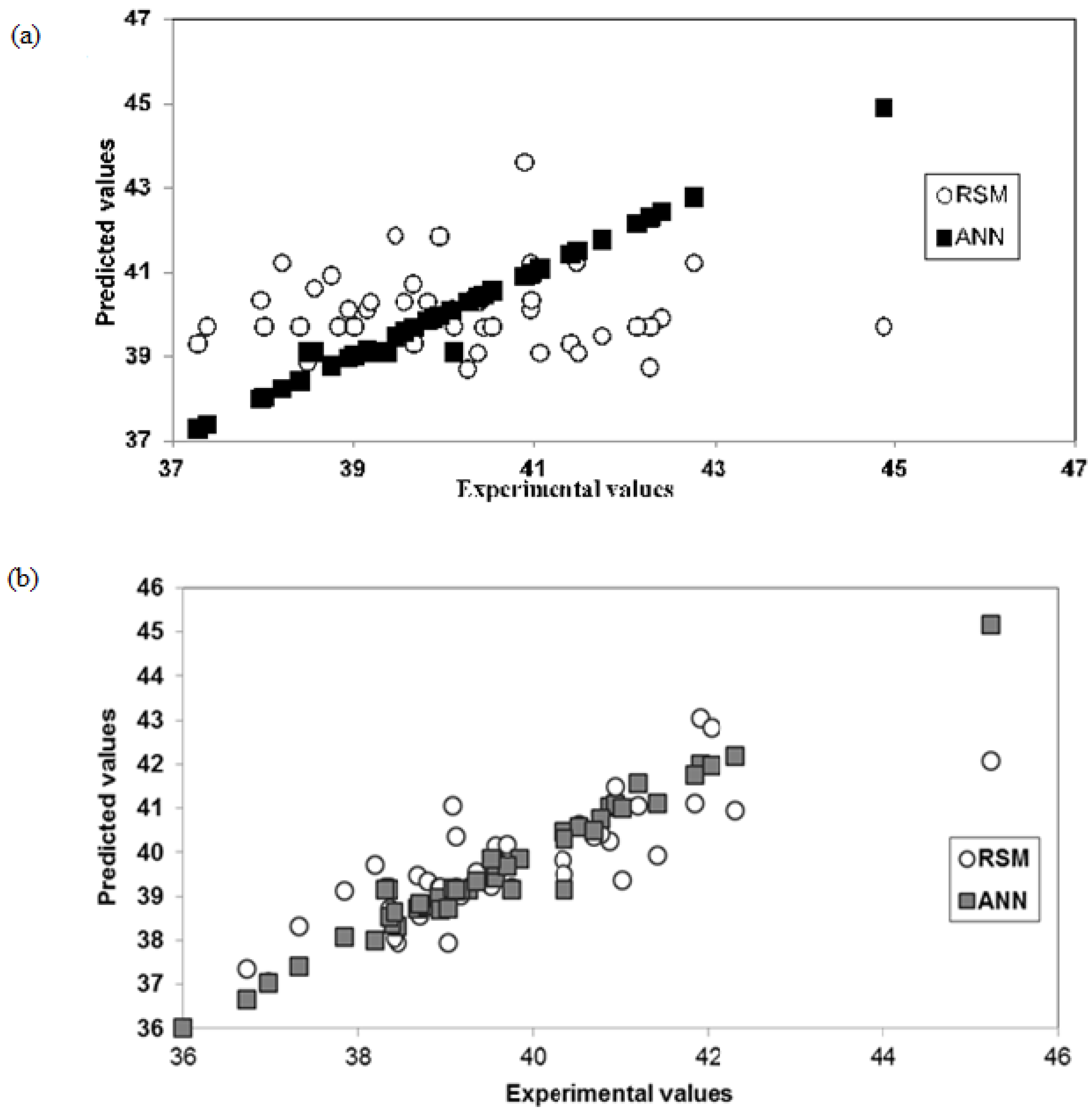
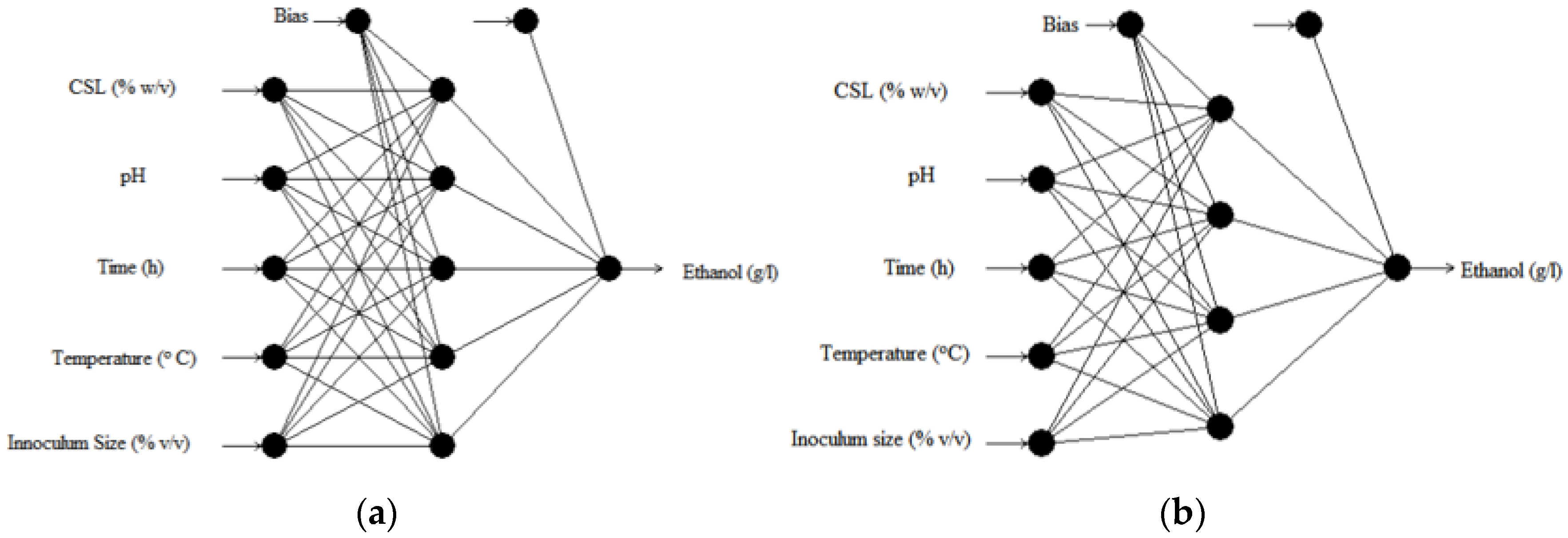
2.4. ANN Modeling Results for Ethanol Production
2.5. Comparing RSM and ANN Optimization for Ethanol Production from the Two Yeast Strains
3. Materials and Methods
3.1. Microorganism and Inoculum Preparation
3.2. Media Composition
3.3. Submerged Fermentation Study
3.4. Experimental Design
Response Surface Methodology
3.5. Analytical Procedures
3.5.1. Reducing Sugar Analysis
3.5.2. Ethanol Concentration Determination
3.5.3. Biomass Concentration Determination
3.6. Modeling Studies
3.6.1. RSM Analysis
3.6.2. ANN Analysis
3.6.3. Predictability of Model Evaluated in Artificial Neural Network (ANN)
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hahn, R.; Cecot, C. The benefits and costs of ethanol: An evaluation of the government’s analysis. J. Regul. Econ. 2009, 35, 275–295. [Google Scholar] [CrossRef]
- Sebayang, A.H.; Hassan, M.H.; Ong, H.C.; Dharma, S.; Silitonga, A.S.; Kusumo, F.; Mahlia, T.M.I.; Bahar, A.H. Optimization of reducing sugar production from manihot glaziovii starch using response surface methodology. Energies 2017, 10, 35. [Google Scholar] [CrossRef]
- Tan, J.; Jahim, J.; Wu, T.; Harun, S.; Mumtaz, T. Use of corn steep liquor as an economical nitrogen source for biosuccinic acid production by Actinobacillus succinogenes. IOP Conf. Ser. Earth Environ. Sci. 2016, 36. [Google Scholar] [CrossRef]
- Malherbe, S.; Bauer, F.; Du Toit, M. Understanding problem fermentations: A review. S. Afr. J. Enol. Vitic. 2007, 28, 169–186. [Google Scholar] [CrossRef]
- Li, Z.; Wang, D.; Shi, Y.-C. Effects of nitrogen source on ethanol production in very high gravity fermentation of corn starch. J. Taiwan Inst. Chem. Eng. 2017, 70, 229–235. [Google Scholar] [CrossRef]
- Andualem, B.; Gessesse, A. Production of microbial medium from defatted brebra (milletia ferruginea) seed flour to substitute commercial peptone agar. Asian Pac. J. Trop. Biomed. 2013, 3, 790–797. [Google Scholar] [CrossRef]
- Azhar, S.H.M.; Abdulla, R.; Jambo, S.A.; Marbawi, H.; Gansau, J.A.; Faik, A.A.M.; Rodrigues, K.F. Yeasts in sustainable bioethanol production: A review. Biochem. Biophys. Rep. 2017, 10, 52–61. [Google Scholar]
- Broach, J.R. Nutritional control of growth and development in yeast. Genetics 2012, 192, 73–105. [Google Scholar] [CrossRef] [PubMed]
- Albers, E.; Larsson, C.; Lidén, G.; Niklasson, C.; Gustafsson, L. Influence of the nitrogen source on Saccharomyces cerevisiae anaerobic growth and product formation. Appl. Environ. Microbiol. 1996, 62, 3187–3195. [Google Scholar] [PubMed]
- Davis, L.; Rogers, P.; Pearce, J.; Peiris, P. Evaluation of zymomonas-based ethanol production from a hydrolysed waste starch stream. Biomass Bioenergy 2006, 30, 809–814. [Google Scholar] [CrossRef]
- Abbasiliasi, S.; Tan, J.S.; Ibrahim, T.A.T.; Bashokouh, F.; Ramakrishnan, N.R.; Mustafa, S.; Ariff, A.B. Fermentation factors influencing the production of bacteriocins by lactic acid bacteria: A review. RSC Adv. 2017, 7, 29395–29420. [Google Scholar] [CrossRef]
- Lawford, H.G.; Rousseau, J.D. Corn steep liquor as a cost-effective nutrition adjunct in high-performancezymomonas ethanol fermentations. Appl. Biochem. Biotechnol. 1997, 63, 287–304. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.-B.; Kim, S.S.; Lee, H.-Y.; Jung, K.-H. High-level production of ethanol during fed-batch ethanol fermentation with a controlled aeration rate and non-sterile glucose powder feeding of Saccharomyces cerevisiae. Biotechnol. Bioprocess Eng. 2009, 14, 591–598. [Google Scholar] [CrossRef]
- Edwinoliver, N.; Thirunavukarasu, K.; Purushothaman, S.; Rose, C.; Gowthaman, M.; Kamini, N. Corn steep liquor as a nutrition adjunct for the production of Aspergillus niger lipase and hydrolysis of oils thereof. J. Agric. Food Chem. 2009, 57, 10658–10663. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; An, M.; Liu, K.; Nagai, S.; Shigematsu, T.; Morimura, S.; Kida, K. Ethanol production from acid hydrolysate of wood biomass using the flocculating yeast Saccharomyces cerevisiae strain kf-7. Process Biochem. 2006, 41, 909–914. [Google Scholar] [CrossRef]
- Souza, A.F.; Rodriguez, D.M.; Ribeaux, D.R.; Luna, M.A.; Lima e Silva, T.A.; Andrade, R.F.S.; Gusmão, N.B.; Campos-Takaki, G.M. Waste soybean oil and corn steep liquor as economic substrates for bioemulsifier and biodiesel production by candida lipolytica UCP 0998. Int. J. Mol. Sci. 2016, 17, 1608. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Atiyeh, H.K.; Phillips, J.R.; Wilkins, M.R.; Huhnke, R.L. Development of low cost medium for ethanol production from syngas by clostridium ragsdalei. Bioresour. Technol. 2013, 147, 508–515. [Google Scholar] [CrossRef] [PubMed]
- Saxena, J.; Tanner, R.S. Optimization of a corn steep medium for production of ethanol from synthesis gas fermentation by clostridium ragsdalei. World J. Microbiol. Biotechnol. 2012, 28, 1553–1561. [Google Scholar] [CrossRef] [PubMed]
- Maddipati, P.; Atiyeh, H.K.; Bellmer, D.D.; Huhnke, R.L. Ethanol production from syngas by clostridium strain p11 using corn steep liquor as a nutrient replacement to yeast extract. Bioresour. Technol. 2011, 102, 6494–6501. [Google Scholar] [CrossRef] [PubMed]
- Kundiyana, D.K.; Huhnke, R.L.; Wilkins, M.R. Syngas fermentation in a 100-l pilot scale fermentor: Design and process considerations. J. Biosci. Bioeng. 2010, 109, 492–498. [Google Scholar] [CrossRef] [PubMed]
- Sreenath, H.K.; Jeffries, T.W. Effect of corn steep liquor on fermentation of mixed sugars by candida shehatae fpl-702. Appl. Biochem. Biotechnol. 1996, 57, 551–561. [Google Scholar] [CrossRef]
- Pereira, F.B.; Guimarães, P.M.; Teixeira, J.A.; Domingues, L. Optimization of low-cost medium for very high gravity ethanol fermentations by Saccharomyces cerevisiae using statistical experimental designs. Bioresour. Technol. 2010, 101, 7856–7863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Açıkel, Ü.; Erşan, M.; Açıkel, Y.S. Optimization of critical medium components using response surface methodology for lipase production by rhizopus delemar. Food Bioprod. Process 2010, 88, 31–39. [Google Scholar] [CrossRef]
- Baş, D.; Boyacı, İ.H. Modeling and optimization ii: Comparison of estimation capabilities of response surface methodology with artificial neural networks in a biochemical reaction. J. Food Eng. 2007, 78, 846–854. [Google Scholar] [CrossRef]
- Osunkanmibi, O.B.; Owolabi, T.O.; Betiku, E. Comparison of artificial neural network and response surface methodology performance on fermentation parameters optimization of bioconversion of cashew apple juice to Gluconic acid. Int. J. Food Eng. 2015, 11, 393–403. [Google Scholar] [CrossRef]
- Hull, S.R.; Yang, B.Y.; Venzke, D.; Kulhavy, K.; Montgomery, R. Composition of corn steep water during steeping. J. Agric. Food Chem. 1996, 44, 1857–1863. [Google Scholar] [CrossRef]
- Oyeniran, O.; Taiwo, A.; Betiku, E. A modeling study by response surface methodology on the culture parameters optimization of citric acid bioproduction from sweet potato peel. Ife J. Technol. 2013, 22, 21–25. [Google Scholar]
- Senanayake, S.N.; Shahidi, F. Lipase-catalyzed incorporation of Docosahexaenoic acid (DHA) into borage oil: Optimization using response surface methodology. Food Chem. 2002, 77, 115–123. [Google Scholar] [CrossRef]
- Ebrahimpour, A.; Rahman, R.N.Z.R.A.; Ch’ng, D.H.E.; Basri, M.; Salleh, A.B. A modeling study by response surface methodology and artificial neural network on culture parameters optimization for thermostable lipase production from a newly isolated thermophilic Geobacillus sp. strain ARM. BMC Biotechnol. 2008, 8, 96. [Google Scholar] [CrossRef] [PubMed]
- Dumancas, G.G.; Adrianto, I.; Bello, G.; Dozmorov, M. Current developments in machine learning techniques in biological data mining. Bioinf. Biol. Insights 2017, 11, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Wesolowski, M.; Suchacz, B. Artificial neural networks: Theoretical background and pharmaceutical applications: A review. J. AOAC Int. 2012, 95, 652–668. [Google Scholar] [CrossRef] [PubMed]
- Betiku, E.; Omilakin, O.R.; Ajala, S.O.; Okeleye, A.A.; Taiwo, A.E.; Solomon, B.O. Mathematical modeling and process parameters optimization studies by artificial neural network and response surface methodology: A case of non-edible neem (Azadirachta indica) seed oil biodiesel synthesis. Energy 2014, 72, 266–273. [Google Scholar] [CrossRef]
- Ighose, B.O.; Adeleke, I.A.; Damos, M.; Junaid, H.A.; Okpalaeke, K.E.; Betiku, E. Optimization of biodiesel production from thevetia peruviana seed oil by adaptive neuro-fuzzy inference system coupled with genetic algorithm and response surface methodology. Energy Convers. Manag. 2017, 132, 231–240. [Google Scholar] [CrossRef]
- Betiku, E.; Taiwo, A.E. Modeling and optimization of bioethanol production from breadfruit starch hydrolyzate vis-à-vis response surface methodology and artificial neural network. Renew. Energy 2015, 74, 87–94. [Google Scholar] [CrossRef]
- Tu, J.V. Advantages and disadvantages of using artificial neural networks versus logistic regression for predicting medical outcomes. J. Chronic Dis. 1996, 49, 1225–1231. [Google Scholar] [CrossRef]
- Schmidhuber, J. Deep learning in neural networks: An overview. Neural Netw. 2015, 61, 85–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díaz-Montaño, D.M. Continuous agave juice fermentation for producing bioethanol. In Biomass Now-Sustainable Growth and Use; InTech: London, UK, 2013. [Google Scholar]
- Sarks, C.; Jin, M.; Sato, T.K.; Balan, V.; Dale, B.E. Studying the rapid bioconversion of lignocellulosic sugars into ethanol using high cell density fermentations with cell recycle. Biotechnol. Biofuels 2014, 7, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rebnegger, C.; Vos, T.; Graf, A.B.; Valli, M.; Pronk, J.T.; Daran-Lapujade, P.; Mattanovich, D. Pichia pastoris exhibits high viability and a low maintenance energy requirement at near-zero specific growth rates. Appl. Environ. Microbiol. 2016, 82, 4570–4583. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, T.; Morley, A. An evolutionary perspective on the crabtree effect. Front. Mol. Biosci. 2014, 1, 17. [Google Scholar] [CrossRef] [PubMed]
- Van Urk, H.; Postma, E.; Scheffers, W.A.; Van Dijken, J.P. Glucose transport in crabtree-positive and crabtree-negative yeasts. J. Gen. Microbiol. 1989, 135, 2399–2406. [Google Scholar] [CrossRef] [PubMed]
- Mezule, L.; Dalecka, B. Adjustment of yeast growth media for the fermentation of lignocellulosic sugars. Chem. Eng. Trans. 2017, 57, 25–30. [Google Scholar]
- Betiku, E.; Alade, O. Media evaluation of bioethanol production from cassava starch hydrolysate using Saccharomyces cerevisiae. Energy Sources Part A 2014, 36, 1990–1998. [Google Scholar] [CrossRef]
- Jin, H.; Liu, R.; He, Y. Kinetics of batch fermentations for ethanol production with immobilized Saccharomyces cerevisiae growing on sweet sorghum stalk juice. Procedia Environ. Sci. 2012, 12, 137–145. [Google Scholar] [CrossRef]
- Sarteshnizi, R.A.; Hosseini, H.; Bondarianzadeh, D.; Colmenero, F.J. Optimization of prebiotic sausage formulation: Effect of using β-glucan and resistant starch by d-optimal mixture design approach. LWT-Food Sci. Technol. 2015, 62, 704–710. [Google Scholar] [CrossRef]
- Tesfaw, A.; Assefa, F. Current trends in bioethanol production by Saccharomyces cerevisiae: Substrate, inhibitor reduction, growth variables, coculture, and immobilization. Int. Sch. Res. Not. 2014. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Sumbhate, S.V.; Nayak, S.; Goupale, D.; Tiwari, A.; Jadon, R.S. Colorimetric method for the estimation of ethanol in alcoholic-drinks. J. Anal. Tech. 2012, 1, 1–6. [Google Scholar]
- Ghaffari, A.; Abdollahi, H.; Khoshayand, M.; Bozchalooi, I.S.; Dadgar, A.; Rafiee-Tehrani, M. Performance comparison of neural network training algorithms in modeling of bimodal drug delivery. Int. J. Pharm. 2006, 327, 126–138. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, M.G.; Ahmad, F.B.H.; Basri, M.; Rahman, M.B.A. Artificial neural network modeling studies to predict the yield of enzymatic synthesis of betulinic acid ester. Electron. J. Biotechnol. 2010, 13, 3–4. [Google Scholar] [CrossRef]
- Basri, M.; Rahman, R.N.Z.R.A.; Ebrahimpour, A.; Salleh, A.B.; Gunawan, E.R.; Rahman, M.B.A. Comparison of estimation capabilities of response surface methodology (RSM) with artificial neural network (ANN) in lipase-catalyzed synthesis of palm-based wax ester. BMC Biotechnol. 2007, 7, 53. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Glucose (g/L) | Type of Yeast | Ethanol (g/L) | Yield (%) | CSL (% w/v) | Scale | Author |
|---|---|---|---|---|---|---|
| 100 | Saccharomyces Type 1 | 43.39 | 85 | 5.5 | Shake Flask | This study |
| 100 | Anchor Instant Yeast | 45.36 | 89 | 5.36 | Shake Flask | This study |
| 107 | Candida shehatae FPL-702 | 40.1 | 73 | 5 | Shake Flask | [21] |
| 300 | Saccharomyces cerevisiae | 125 | 82 | 110 | Shake Flask | [22] |
| Runs | CSL (% w/v) | pH | Time (h) | Temperature (°C) | Inoculum Size (% v/v) | Experimental Values (g/L) | Predicted Values (g/L) | Experimental Values (g/L) | Predicted Values (g/L) | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ST1 | RSM | ANN | AIY | RSM | ANN | ||||||
| 1 | 0.5 | 5.0 | 24 | 30 | 3.0 | 40.96 | 40.92 | 40.96 | 40.34 | 39.82 | 40.47 |
| 2 | 0.5 | 4.5 | 36 | 30 | 3.0 | 42.30 | 40.30 | 42.30 | 42.03 | 42.83 | 41.98 |
| 3 | 0.5 | 4.5 | 12 | 30 | 3.0 | 39.69 | 40.30 | 39.69 | 40.88 | 40.26 | 41.03 |
| 4 | 0.5 | 4.0 | 24 | 30 | 3.0 | 37.99 | 39.69 | 37.99 | 42.32 | 40.94 | 42.19 |
| 5 | 0.5 | 4.5 | 24 | 30 | 5.5 | 41.49 | 41.86 | 41.49 | 40.95 | 41.48 | 41.09 |
| 6 | 0.5 | 4.5 | 24 | 35 | 3.0 | 39.47 | 40.30 | 39.47 | 36.99 | 37.05 | 37.03 |
| 7 | 0.5 | 4.5 | 24 | 30 | 0.5 | 39.02 | 38.74 | 39.02 | 38.94 | 39.20 | 38.70 |
| 8 | 0.5 | 4.5 | 24 | 25 | 3.0 | 41.50 | 40.30 | 41.50 | 39.75 | 39.19 | 39.14 |
| 9 | 3.0 | 5.0 | 24 | 30 | 5.5 | 39.90 | 39.91 | 39.90 | 38.46 | 37.94 | 38.32 |
| 10 | 3.0 | 4.0 | 24 | 25 | 3.0 | 38.77 | 39.09 | 38.77 | 40.53 | 40.63 | 40.56 |
| 11 | 3.0 | 4.5 | 12 | 30 | 5.5 | 40.39 | 39.30 | 40.39 | 38.19 | 39.70 | 38.00 |
| 12 | 3.0 | 4.0 | 36 | 30 | 3.0 | 36.99 | 39.09 | 36.99 | 38.69 | 39.47 | 38.73 |
| 13 | 3.0 | 4.5 | 24 | 30 | 3.0 | 39.38 | 39.70 | 39.11 | 41.90 | 43.03 | 41.99 |
| 14 | 3.0 | 4.5 | 24 | 30 | 3.0 | 38.84 | 39.70 | 39.10 | 38.80 | 39.33 | 38.77 |
| 15 | 3.0 | 4.5 | 24 | 35 | 0.5 | 39.97 | 40.11 | 39.97 | 39.57 | 39.82 | 39.41 |
| 16 | 3.0 | 4.5 | 12 | 25 | 3.0 | 39.16 | 39.70 | 39.16 | 39.84 | 39.84 | 39.85 |
| 17 | 3.0 | 4.5 | 24 | 30 | 3.0 | 38.51 | 39.70 | 39.11 | 39.27 | 39.19 | 39.14 |
| 18 | 3.0 | 4.5 | 12 | 30 | 0.5 | 40.41 | 40.11 | 40.41 | 41.42 | 39.93 | 41.12 |
| 19 | 3.0 | 4.5 | 24 | 25 | 0.5 | 40.10 | 40.11 | 40.10 | 39.12 | 40.36 | 39.13 |
| 20 | 3.0 | 4.5 | 36 | 30 | 5.5 | 40.96 | 39.30 | 40.96 | 38.71 | 38.55 | 38.84 |
| 21 | 3.0 | 4.5 | 24 | 25 | 5.5 | 37.28 | 39.30 | 37.28 | 38.93 | 39.20 | 38.95 |
| 22 | 3.0 | 5.0 | 36 | 30 | 3.0 | 39.58 | 40.32 | 39.58 | 38.40 | 38.51 | 38.36 |
| 23 | 3.0 | 4.5 | 24 | 35 | 5.5 | 40.45 | 39.30 | 40.45 | 37.84 | 39.12 | 38.08 |
| 24 | 3.0 | 4.5 | 36 | 25 | 3.0 | 39.84 | 39.70 | 39.84 | 36.00 | 35.84 | 36.02 |
| 25 | 3.0 | 5.0 | 12 | 30 | 3.0 | 38.03 | 40.32 | 38.03 | 40.36 | 39.50 | 40.31 |
| 26 | 3.0 | 4.5 | 36 | 35 | 3.0 | 40.30 | 39.70 | 40.30 | 38.36 | 38.73 | 39.14 |
| 27 | 3.0 | 5.0 | 24 | 35 | 3.0 | 42.15 | 40.32 | 42.15 | 41.20 | 41.06 | 41.56 |
| 28 | 3.0 | 4.5 | 12 | 35 | 3.0 | 40.97 | 39.70 | 40.97 | 40.78 | 40.41 | 40.77 |
| 29 | 3.0 | 5.0 | 24 | 30 | 0.5 | 40.55 | 40.72 | 40.55 | 38.30 | 39.19 | 39.14 |
| 30 | 3.0 | 4.0 | 24 | 35 | 3.0 | 39.68 | 39.09 | 39.68 | 39.08 | 41.06 | 39.17 |
| 31 | 3.0 | 4.0 | 24 | 30 | 5.5 | 41.07 | 38.69 | 41.07 | 39.70 | 40.17 | 39.68 |
| 32 | 3.0 | 4.0 | 24 | 30 | 0.5 | 40.28 | 39.49 | 40.28 | 39.12 | 39.19 | 39.14 |
| 33 | 3.0 | 4.5 | 24 | 30 | 3.0 | 38.58 | 39.70 | 39.11 | 38.36 | 39.19 | 38.54 |
| 34 | 3.0 | 4.5 | 36 | 30 | 0.5 | 42.29 | 40.11 | 42.29 | 38.42 | 38.06 | 38.65 |
| 35 | 3.0 | 4.0 | 12 | 30 | 3.0 | 38.95 | 39.09 | 38.95 | 39.52 | 39.22 | 39.85 |
| 36 | 3.0 | 4.5 | 24 | 30 | 3.0 | 39.20 | 39.70 | 39.11 | 41.02 | 39.37 | 40.99 |
| 37 | 3.0 | 4.5 | 24 | 30 | 3.0 | 40.13 | 39.70 | 39.11 | 39.04 | 37.93 | 38.71 |
| 38 | 3.0 | 5.0 | 24 | 25 | 3.0 | 38.42 | 40.32 | 38.42 | 39.35 | 39.55 | 39.33 |
| 39 | 5.5 | 4.5 | 36 | 30 | 3.0 | 42.42 | 41.22 | 42.42 | 36.74 | 37.36 | 36.65 |
| 40 | 5.5 | 5.0 | 24 | 30 | 3.0 | 44.88 | 41.84 | 44.88 | 37.33 | 38.31 | 37.41 |
| 41 | 5.5 | 4.5 | 24 | 30 | 5.5 | 37.39 | 38.86 | 37.39 | 39.57 | 40.15 | 39.71 |
| 42 | 5.5 | 4.5 | 12 | 30 | 3.0 | 41.42 | 41.22 | 41.42 | 39.18 | 39.02 | 39.14 |
| 43 | 5.5 | 4.5 | 24 | 35 | 3.0 | 38.23 | 41.22 | 38.23 | 40.35 | 39.19 | 39.14 |
| 44 | 5.5 | 4.5 | 24 | 25 | 3.0 | 41.77 | 41.22 | 41.77 | 41.84 | 41.10 | 41.74 |
| 45 | 5.5 | 4.5 | 24 | 30 | 0.5 | 42.77 | 43.59 | 42.77 | 45.23 | 42.08 | 45.17 |
| 46 | 5.5 | 4.0 | 24 | 30 | 3.0 | 40.90 | 40.61 | 40.90 | 40.70 | 40.36 | 40.50 |
| Runs | CSL (% w/v) | pH | Time (h) | Temperature (°C) | Inoculum Size (% v/v) | Saccharomyces Type 1 (ST1) α | Anchor Instant Yeast (AIY) α | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Experimental | Predicted | Experimental | Predicted | ||||||||
| (g/L) | RSM | ANN | (g/L) | RSM | ANN | ||||||
| 1 | 0 | 1 | 0 | 0 | 1 | * 39.9 | 39.91 | 39.9 | * 38.46 | 37.94 | 38.32 |
| 2 | 1 | 0 | 1 | 0 | 0 | 42.42 | 41.22 | 42.42 | 36.74 | 37.36 | 36.65 |
| 3 | −1 | 1 | 0 | 0 | 0 | 40.96 | 40.92 | 40.96 | 40.34 | 39.82 | 40.47 |
| 4 | 0 | −1 | 0 | −1 | 0 | * 38.77 | 39.09 | 38.77 | * 40.53 | 40.63 | 40.56 |
| 5 | 0 | 0 | −1 | 0 | 1 | 40.39 | 39.3 | 40.39 | 38.19 | 39.7 | 38 |
| 6 | 0 | −1 | 1 | 0 | 0 | 36.99 | 39.09 | 36.99 | 38.69 | 39.47 | 38.73 |
| 7 | 0 | 0 | 0 | 0 | 0 | * 39.38 | 39.7 | 39.11 | * 41.9 | 43.03 | 41.99 |
| 8 | 0 | 0 | 0 | 0 | 0 | * 38.84 | 39.7 | 39.11 | 38.8 | 39.33 | 38.77 |
| 9 | 1 | 1 | 0 | 0 | 0 | 44.88 | 41.84 | 44.88 | 37.33 | 38.31 | 37.41 |
| 10 | 0 | 0 | 0 | 1 | −1 | 39.97 | 40.11 | 39.97 | 39.57 | 39.82 | 39.41 |
| 11 | 0 | 0 | −1 | −1 | 0 | 39.16 | 39.7 | 39.16 | 39.84 | 39.84 | 39.85 |
| 12 | 1 | 0 | 0 | 0 | 1 | 37.39 | 38.86 | 37.39 | * 39.57 | 40.15 | 39.71 |
| 13 | 0 | 0 | 0 | 0 | 0 | * 38.51 | 39.7 | 39.11 | * 39.27 | 39.19 | 39.14 |
| 14 | −1 | 0 | 1 | 0 | 0 | 42.3 | 40.3 | 42.3 | 42.03 | 42.83 | 41.98 |
| 15 | 0 | 0 | −1 | 0 | −1 | 40.41 | 40.11 | 40.41 | 41.42 | 39.93 | 41.12 |
| 16 | 0 | 0 | 0 | −1 | −1 | * 40.1 | 40.11 | 40.1 | 39.12 | 40.36 | 39.13 |
| 17 | 0 | 0 | 1 | 0 | 1 | 40.96 | 39.3 | 40.96 | 38.71 | 38.55 | 38.84 |
| 18 | 0 | 0 | 0 | −1 | 1 | 37.28 | 39.3 | 37.28 | 38.93 | 39.2 | 38.95 |
| 19 | −1 | 0 | −1 | 0 | 0 | * 39.69 | 40.3 | 39.69 | * 40.88 | 40.26 | 41.03 |
| 20 | 0 | 1 | 1 | 0 | 0 | 39.58 | 40.32 | 39.58 | 38.4 | 38.51 | 38.36 |
| 21 | −1 | −1 | 0 | 0 | 0 | 37.99 | 39.69 | 37.99 | 42.32 | 40.94 | 42.19 |
| 22 | 0 | 0 | 0 | 1 | 1 | * 40.45 | 39.3 | 40.45 | * 37.84 | 39.12 | 38.08 |
| 23 | 1 | 0 | −1 | 0 | 0 | 41.42 | 41.22 | 41.42 | 39.18 | 39.02 | 39.14 |
| 24 | 1 | 0 | 0 | 1 | 0 | 38.23 | 41.22 | 38.23 | 40.35 | 39.19 | 39.14 |
| 25 | −1 | 0 | 0 | 0 | 1 | * 41.49 | 41.86 | 41.49 | * 40.95 | 41.48 | 41.09 |
| 26 | −1 | 0 | 0 | 1 | 0 | 39.47 | 40.3 | 39.47 | 36.99 | 37.05 | 37.03 |
| 27 | 0 | 0 | 1 | −1 | 0 | 39.84 | 39.7 | 39.84 | 36 | 35.84 | 36.02 |
| 28 | 0 | 1 | −1 | 0 | 0 | * 38.03 | 40.32 | 38.03 | * 40.36 | 39.5 | 40.31 |
| 29 | 0 | 0 | 1 | 1 | 0 | 40.3 | 39.7 | 40.3 | 38.36 | 38.73 | 39.14 |
| 30 | 0 | 1 | 0 | 1 | 0 | 42.15 | 40.32 | 42.15 | 41.2 | 41.06 | 41.56 |
| 31 | 0 | 0 | −1 | 1 | 0 | * 40.97 | 39.7 | 40.97 | * 40.78 | 40.41 | 40.77 |
| 32 | 0 | 1 | 0 | 0 | −1 | 40.55 | 40.72 | 40.55 | 38.3 | 39.19 | 39.14 |
| 33 | 0 | −1 | 0 | 1 | 0 | 39.68 | 39.09 | 39.68 | 39.08 | 41.06 | 39.17 |
| 34 | 0 | −1 | 0 | 0 | 1 | * 41.07 | 38.69 | 41.07 | * 39.7 | 40.17 | 39.68 |
| 35 | 0 | −1 | 0 | 0 | −1 | 40.28 | 39.49 | 40.28 | * 39.12 | 39.19 | 39.14 |
| 36 | 1 | 0 | 0 | −1 | 0 | 41.77 | 41.22 | 41.77 | 41.84 | 41.1 | 41.74 |
| 37 | 1 | 0 | 0 | 0 | −1 | * 42.77 | 43.59 | 42.77 | * 45.23 | 42.08 | 45.17 |
| 38 | 1 | −1 | 0 | 0 | 0 | 40.9 | 40.61 | 40.9 | 40.7 | 40.36 | 40.5 |
| 39 | 0 | 0 | 0 | 0 | 0 | 38.58 | 39.7 | 39.11 | 38.36 | 39.19 | 38.54 |
| 40 | −1 | 0 | 0 | 0 | −1 | * 39.02 | 38.74 | 39.02 | * 38.94 | 39.2 | 38.7 |
| 41 | 0 | 0 | 1 | 0 | −1 | 42.29 | 40.11 | 42.29 | 38.42 | 38.06 | 38.65 |
| 42 | 0 | −1 | −1 | 0 | 0 | 38.95 | 39.09 | 38.95 | 39.52 | 39.22 | 39.85 |
| 43 | −1 | 0 | 0 | −1 | 0 | * 41.5 | 40.3 | 41.5 | * 39.75 | 39.19 | 39.14 |
| 44 | 0 | 0 | 0 | 0 | 0 | 39.2 | 39.7 | 39.11 | 41.02 | 39.37 | 40.99 |
| 45 | 0 | 0 | 0 | 0 | 0 | 40.13 | 39.7 | 39.11 | 39.04 | 37.93 | 38.71 |
| 46 | 0 | 1 | 0 | −1 | 0 | * 38.42 | 40.32 | 38.42 | * 39.35 | 39.55 | 39.33 |
| Saccharomyces Type 1 Yeast (ST1) | Anchor Instant Yeast (AIY) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Source | SS | df | MS | F value | p value | SS | df | MS | F value | p value |
| Model | 39.11 | 5 | 7.82 | 4.070 | <0.0001 | 84.80 | 20 | 4.24 | 2.74 | <0.0001 |
| CSL | 3.39 | 1 | 3.39 | 1.76 | 0.1916 | 10.84 | 1 | 10.84 | 6.99 | <0.0001 |
| pH | 6.05 | 1 | 6.05 | 3.15 | 0.0836 | 2.15 | 1 | 2.15 | 1.39 | 0.2503 |
| Time | - | - | - | - | - | 16.28 | 1 | 16.28 | 10.51 | <0.0001 |
| Temperature | - | - | - | - | - | 0.65 | 1 | 0.65 | 0.42 | 0.5224 |
| Inoculum | 2.61 | 1 | 2.61 | 1.36 | 0.2505 | 11.32 | 1 | 11.32 | 7.31 | <0.0001 |
| CSL× Inoculum | 15.39 | 1 | 15.39 | 8.00 | <0.0001 | 6.97 | 1 | 6.97 | 4.50 | <0.0001 |
| Temperature × inoculum | - | - | - | - | - | 12.75 | 1 | 12.75 | 8.23 | <0.0001 |
| CSL2 | 11.67 | 1 | 11.67 | 6.07 | <0.0001 | 7.65 | 1 | 7.65 | 4.94 | <0.0001 |
| Inoculum2 | - | - | - | - | - | 0.68 | 1 | 0.68 | 0.44 | 0.5152 |
| Residual | 76.87 | 40 | 1.92 | - | - | 38.73 | 25 | 1.55 | - | - |
| Lack of fit | 75.04 | 35 | 2.14 | 5.88 | 0.0280 | 35.58 | 20 | 1.78 | 2.82 | 0.1266 |
| Pure Error | 1.82 | 5 | 0.37 | - | - | 3.15 | 5 | 0.63 | - | - |
| Correlation Total | 115.97 | 45 | - | - | - | 123.54 | 45 | - | - | - |
| Strain | Transfer Function Topology | RMSE | MSE | R2 | AAD% | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Training | Testing | Training | Testing | Training | Testing | Training | Testing | |||
| ST1 | Sigmoid | 5-4-1 | 0.2164 | 0.1641 | 0.04681 | 0.02693 | 0.9844 | 0.9834 | 0.1613 | 0.1402 |
| AIY | Tanh | 5-5-1 | 0.4333 | 0.1952 | 0.1878 | 0.0381 | 0.9219 | 0.9860 | 0.8260 | 0.3406 |
| Strain | RSM | ANN | ||||||
|---|---|---|---|---|---|---|---|---|
| R2 | AAD% | RMSE | MSE | R2 | AAD% | RMSE | MSE | |
| ST1 | 0.34 | 2.52 | 1.29 | 1.68 | 0.98 | 0.15 | 0.19 | 0.039 |
| AIY | 0.98 | 1.72 | 0.97 | 0.84 | 0.99 | 0.46 | 0.29 | 0.089 |
| Yeast Strain | Tool | CSL (% w/v) | pH | Time (h) | Temperature (°C) | Inoculum (% v/v) | Predicted (g/L) | Validated (g/L) |
|---|---|---|---|---|---|---|---|---|
| AIY | RSM | 5.36 | 4.06 | 12.99 | 28.14 | 0.72 | 45.48 | 45.36 ± 0.02 |
| ANN | 0.50 | 4.99 | 35.99 | 34.17 | 5.49 | 45.73 | 40.60 ± 0.01 | |
| ST1 | RSM | 5.50 | 5.00 | 26.83 | 32.68 | 0.50 | 44.20 | 43.38 ± 0.0002 |
| ANN | 0.50 | 4.99 | 36 | 34.17 | 5.49 | 45.73 | 41.30 ± 0.001 |
| Coded Factor Levels | |||||
|---|---|---|---|---|---|
| Variables | Units | Symbols | −1 | 0 | 1 |
| CSL | (% w/v) | X1 | 0.5 | 5 | 5.5 |
| pH | - | X2 | 4 | 4.5 | 5 |
| Time | (h) | X3 | 12 | 24 | 36 |
| Temperature | (°C) | X4 | 25 | 30 | 35 |
| Inoculum Size | (% v/v) | X5 | 0.5 | 3 | 5.5 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taiwo, A.E.; Madzimbamuto, T.N.; Ojumu, T.V. Optimization of Corn Steep Liquor Dosage and Other Fermentation Parameters for Ethanol Production by Saccharomyces cerevisiae Type 1 and Anchor Instant Yeast. Energies 2018, 11, 1740. https://doi.org/10.3390/en11071740
Taiwo AE, Madzimbamuto TN, Ojumu TV. Optimization of Corn Steep Liquor Dosage and Other Fermentation Parameters for Ethanol Production by Saccharomyces cerevisiae Type 1 and Anchor Instant Yeast. Energies. 2018; 11(7):1740. https://doi.org/10.3390/en11071740
Chicago/Turabian StyleTaiwo, Abiola Ezekiel, Tafirenyika Nyamayaro Madzimbamuto, and Tunde Victor Ojumu. 2018. "Optimization of Corn Steep Liquor Dosage and Other Fermentation Parameters for Ethanol Production by Saccharomyces cerevisiae Type 1 and Anchor Instant Yeast" Energies 11, no. 7: 1740. https://doi.org/10.3390/en11071740