
First Report and Genetic Analysis of the Invasive Species A. fruticosa L. in Greece: A Combined Genetic and Regeneration Study

 ,
,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. AFLP Procedure
2.3. Index of Regeneration
2.4. Data Collection and Statistical Analysis for AFLP Data
2.5. Habitat Information of 91E0*
3. Results
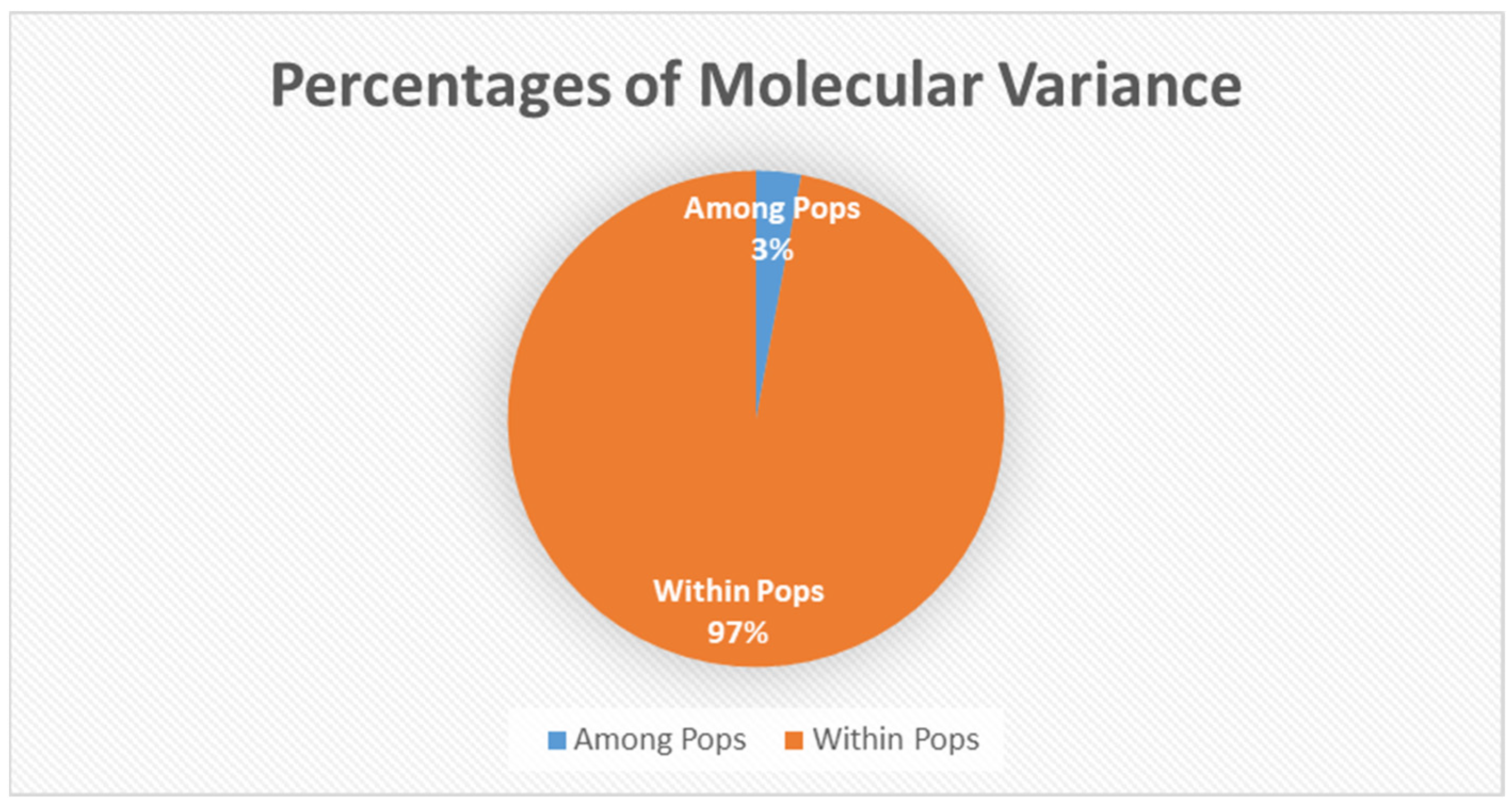
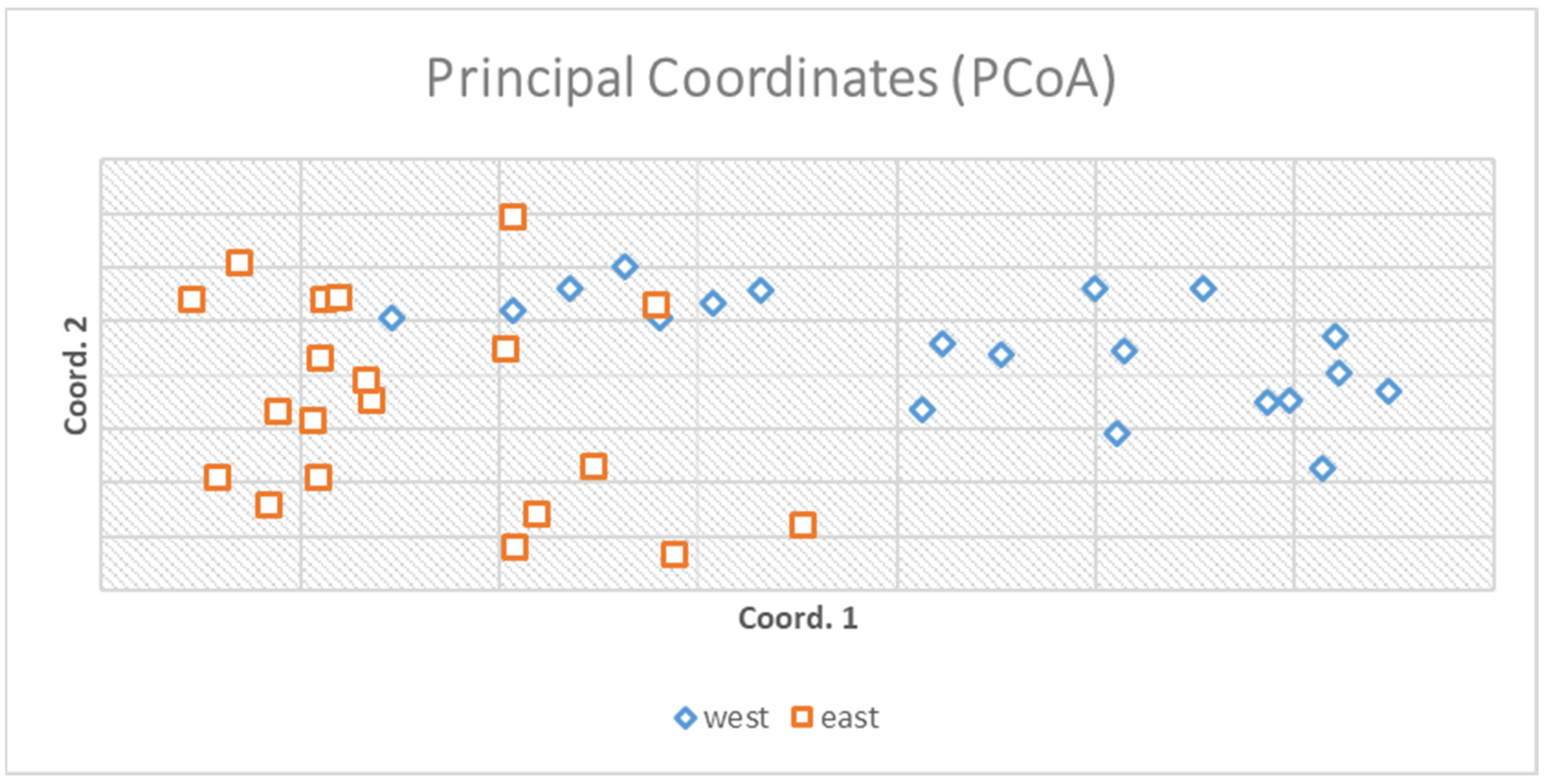
3.1. Genetic Results
3.2. Density and Index of Regeneration
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Richardson, D.M.; Pyšek, P.; Rejmanek, M.; Barbour, M.G.; Panetta, F.D.; West, C.J. Naturalization and invasion of alien plants: Concepts and definitions. Divers. Distrib. 2000, 6, 93–107. [Google Scholar] [CrossRef]
- Korakaki, E.; Legakis, A.; Katsanevakis, S.; Koulelis, P.; Avramidou, E.; Soulioti, N.; Petrakis, P. Invasive alien species of greece. Invasive Alien Species Obs. Issues Around World 2021, 3, 124–189. [Google Scholar]
- Miyawaki, S.; Washitani, I. Invasive alien plant species in riparian areas of japan: The contribution of agricultural weeds, revegetation species and aquacultural species. Glob. Environ. Res.-Engl. Ed. 2004, 8, 89–101. [Google Scholar]
- Callaway, R.M.; Aschehoug, E.T. Invasive plants versus their new and old neighbors: A mechanism for exotic invasion. Science 2000, 290, 521–523. [Google Scholar] [CrossRef] [PubMed]
- Ivanescu, B.; Lungu, C.; Vlase, L.; Gradinaru, A.C.; Tuchilus, C. Hplc analysis of phenolic compounds, antioxidant and antimicrobial activity of Amorpha fruticosa L. Extracts. J. Plant Dev. 2019, 26, 77. [Google Scholar] [CrossRef]
- Straub, S.C.; Sorrie, B.A.; Weakley, A.S. Amorpha confusa, a new name for an old Amorpha (Fabaceae: Amorpheae). J. Bot. Res. Inst. Tex. 2009, 151–155. [Google Scholar]
- Straub, S.C.; Doyle, J.J. Molecular phylogenetics of Amorpha (Fabaceae): An evaluation of monophyly, species relationships, and polyploid origins. Mol. Phylogenetics Evol. 2014, 76, 49–66. [Google Scholar] [CrossRef]
- Liu, J.; Wang, E.T.; Chen, W.X. Diverse rhizobia associated with woody legumes Wisteria sinensis, Cercis racemosa and Amorpha fruticosa grown in the temperate zone of China. Syst. Appl. Microbiol. 2005, 28, 465–477. [Google Scholar] [CrossRef]
- Chinese, P. A Dictionary of Chinese Materia Medica; Chinese Science and Technology Press of Medicine: Beijing, China, 1997. [Google Scholar]
- Mitscher, L.A.; Park, Y.H.; Alshamma, A.; Hudson, P.B.; Haas, T. Amorfrutin a and b, bibenzyl antimicrobial agents from Amorpha fruticosa. Phytochemistry 1981, 20, 781–785. [Google Scholar] [CrossRef]
- Krpan, A.P.; Tomašić, Ž.; Bašić Palković, P. Biopotential of indigobusch (Amorpha fruticosa L.)–second year of investigation. Šumarski List. 2011, 135, 103–112. [Google Scholar]
- Kozuharova, E.; Matkowski, A.; Woźniak, D.; Simeonova, R.; Naychov, Z.; Malainer, C.; Mocan, A.; Nabavi, S.M.; Atanasov, A.G. Amorpha fruticosa–a noxious invasive alien plant in europe or a medicinal plant against metabolic disease? Front. Pharmacol. 2017, 8, 333. [Google Scholar] [CrossRef] [PubMed]
- Szigetvári, C. Initial steps in the regeneration of a floodplain meadow after a decade of dominance of an invasive transformer shrub, Amorpha fruticosa L. Tiscia 2002, 33, 67–77. [Google Scholar]
- Pedashenko, H.P.; Apostolova, I.I.; Vassilev, K.V. Amorpha fruticosa invasibility of different habitats in lower danube. Phytol. Balc. 2012, 18, 285–291. [Google Scholar]
- Arianoutsou, M.; Bazos, I.; Delipetrou, P.; Kokkoris, Y. The alien flora of Greece: Taxonomy, life traits and habitat preferences. Biol. Invasions 2010, 12, 3525–3549. [Google Scholar] [CrossRef]
- Samaras, A.G.; Koutitas, C.G. Modelling the impact on coastal morphology of the water management in transboundary river basins: The case of river Nestos. Manag. Environ. Qual. Int. J. 2008, 19, 455–466. [Google Scholar] [CrossRef]
- Décamps, H.; Décamps, O. Mediterranean Riparian Woodlands; Conservation of Mediterranean Wetlands; Tour du Valat: Arles, France, 2001. [Google Scholar]
- Mallinis, G.; Emmanoloudis, D.; Giannakopoulos, V.; Maris, F.; Koutsias, N. Mapping and interpreting historical land cover/land use changes in a natura 2000 site using earth observational data: The case of Nestos delta, Greece. Appl. Geogr. 2011, 31, 312–320. [Google Scholar] [CrossRef]
- Dafis, S.; Papastergiadou, E.; Georghiou, K.; Babalonas, D.; Georgiadis, T.; Papageorgiou, M.; Lazaridou, T.; Tsiaoussi, V. The Greek ‘habitat’project Natura 2000: An overview. In Life Contract B4-32000/94/756; Commission of the European Communities, The Goulandris Natural History Museum-Greek Biotope/Wetland Centre; Springer: Berlin/Heidelberg, Germany, 1996. [Google Scholar]
- Bianchi, L.; Paci, M. Dinamica evolutiva e gestione delle abetine toscane: Sintesi di quarant’anni di ricerche. For.-J. Silvic. For. Ecol. 2008, 5, 122. [Google Scholar] [CrossRef]
- Vekemans, X.; Beauwens, T.; Lemaire, M.; Roldán-Ruiz, I. Data from amplified fragment length polymorphism (AFLP) markers show indication of size homoplasy and of a relationship between degree of homoplasy and fragment size. Mol. Ecol. 2002, 11, 139–151. [Google Scholar] [CrossRef]
- Peakall, R.O.D.; Smouse, P.E. Genalex 6: Genetic analysis in excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Taftsoglou, M.; Valkaniotis, S.; Papathanassiou, G.; Klimis, N.; Dokas, I. A detailed liquefaction susceptibility map of nestos river delta, thrace, Greece based on surficial geology and geomorphology. Geosciences 2022, 12, 361. [Google Scholar] [CrossRef]
- DeHaan, L.; Ehlke, N.; Sheaffer, C.; Wyse, D.; DeHaan, R. Evaluation of diversity among north american accessions of false indigo (Amorpha fruticosa L.) for forage and biomass. Genet. Resour. Crop Evol. 2006, 53, 1463–1476. [Google Scholar] [CrossRef]
- Li, M.; Liu, Z.; Liu, C.; Zhu, F.; Wang, K.; Wang, Z.; Li, X.; Lan, X.; Guan, Q. Drought resistance of tobacco overexpressing the afnac1 gene of Amorpha fruticosa Linn. Front. Plant Sci. 2022, 13, 980171. [Google Scholar] [CrossRef] [PubMed]
- Dat, N.T.; Lee, J.-H.; Lee, K.; Hong, Y.-S.; Kim, Y.H.; Lee, J.J. Phenolic constituents of Amorpha fruticosa that inhibit nf-κb activation and related gene expression. J. Nat. Prod. 2008, 71, 1696–1700. [Google Scholar] [CrossRef] [PubMed]
- Song, F.; Qi, D.; Liu, X.; Kong, X.; Gao, Y.; Zhou, Z.; Wu, Q. Proteomic analysis of symbiotic proteins of Glomus mosseae and Amorpha fruticosa. Sci. Rep. 2015, 5, 18031. [Google Scholar] [CrossRef] [PubMed]
- Todorova, T.; Boyadzhiev, K.; Shkondrov, A.; Parvanova, P.; Dimitrova, M.; Ionkova, I.; Krasteva, I.; Kozuharova, E.; Chankova, S. Screening of Amorpha fruticosa and Ailanthus altissima extracts for genotoxicity/antigenotoxicity, mutagenicity/antimutagenicity and carcinogenicity/anticarcinogenicity. BioRisk 2022, 17, 201–212. [Google Scholar] [CrossRef]
- Cvetković, D.M.; Jovankić, J.V.; Milutinović, M.G.; Nikodijević, D.D.; Grbović, F.J.; Ćirić, A.R.; Topuzović, M.D.; Marković, S.D. The anti-invasive activity of Robinia pseudoacacia L. and Amorpha fruticosa L. On breast cancer mda-mb-231 cell line. Biologia 2019, 74, 915–928. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Mean and SE over Loci for Each Population | Na | Ne | I | He | |
|---|---|---|---|---|---|
| Western population | Mean | 1.671 | 1.184 | 0.239 | 0.136 |
| SE | 0.042 | 0.011 | 0.010 | 0.007 | |
| Eastern population | Mean | 1.661 | 1.155 | 0.216 | 0.119 |
| SE | 0.042 | 0.010 | 0.009 | 0.006 |
| River Side | Mean Height (m) | Mean Density (Trees ha −1) | Mean IR (cm m−2) |
|---|---|---|---|
| West | 1.01 | 1070.46 | 10.17 |
| East | 1.45 | 84.88 | 1.23 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avramidou, E.V.; Korakaki, E.; Malliarou, E.; Solomou, A.D.; Mantakas, G.; Karetsos, G. First Report and Genetic Analysis of the Invasive Species A. fruticosa L. in Greece: A Combined Genetic and Regeneration Study. Ecologies 2023, 4, 627-635. https://doi.org/10.3390/ecologies4030041
Avramidou EV, Korakaki E, Malliarou E, Solomou AD, Mantakas G, Karetsos G. First Report and Genetic Analysis of the Invasive Species A. fruticosa L. in Greece: A Combined Genetic and Regeneration Study. Ecologies. 2023; 4(3):627-635. https://doi.org/10.3390/ecologies4030041
Chicago/Turabian StyleAvramidou, Evangelia V., Evangelia Korakaki, Ermioni Malliarou, Alexandra D. Solomou, George Mantakas, and George Karetsos. 2023. "First Report and Genetic Analysis of the Invasive Species A. fruticosa L. in Greece: A Combined Genetic and Regeneration Study" Ecologies 4, no. 3: 627-635. https://doi.org/10.3390/ecologies4030041