1. Introduction
In the last few decades, new scientific fields have taken advantages of complex network approaches. This interest emerged, in part, by virtue of technological advances that generate new datasets in computational, social, biological, and others sciences [
1,
2,
3]. Examples include modern brain mapping techniques, such as functional magnetic resonance imaging (fMRI), that have provided previously inaccessible information about interaction patterns in the human brain [
4]. The theory of complex networks has proven to be a crucial tool to understand the interactions and dynamics in large systems.
Attempts to characterize new datasets bring up the challenge of extracting relevant features regarding the network’s structure. One of the main concerns is to identify the role of each node in the network and how the nodes cooperate to give rise to emergent behaviors. The majority of measurements that have been proposed in the last few decades allow the ranking of nodes’ importance by the number of connections, centrality, etc. [
5,
6,
7].
Instead of ranking a node’s relative importance, we ask how the nodes contribute locally to the global connectivity of the network, with the aim of better understanding the individual role played by each node in the network. We wish to characterize this role by quantifying the structural information of the diversity of influences among nodes.
The nodes in a network interact with their neighbors and, indirectly, with the neighbors of neighbors; and also with more distant nodes with even greater “neighborhood radius” (
Figure 1).
We aim to quantify the diversity of radii or distance of influences on a given node exercised by all other nodes in the network. For each node, we calculate the Shannon entropy functional [
8] of the probability distribution of the geodesic distances, that is, the shortest path lengths between each node and all other nodes. We call this measurement
geodesic entropy. Nodes with a great diversity of influences (i.e., with high geodesic entropy) may play an important role in, for example, to guaranteeing specialization of functional patterns. Besides, nodes with a low diversity of influences may indicate constraints relevant to network robustness. The statistical details of the distribution of distances, quantified by the Shannon entropy, may be a key to understanding how emergent behaviors arise.
We illustrate and apply our method to real network data. We use the geodesic entropy to analyze human brain functional networks under the influence of the psychedelic brew Ayahuasca.
Ayahuasca is a sacred brew from Amazonian indigenous culture made by decoction of two plants from Amazonian flora—the leaves of the bush Psychotria Viridis, that contains N, N-Dimethyltryptamine (DMT), and the vine Banisteriopsis caapi, that contains monoamine oxidase inhibitors MAOi [
9]. The DMT is a serotonergic psychedelic similar to LSD and mescaline [
10,
11] but fast metabolized by the human body. The MAOi’s act slowing down this degradation, allowing the DMT to cross the blood-brain barrier and enabling hours of psychedelic experience [
9]. For more information about Ayahuasca we refer [
12,
13,
14,
15,
16].
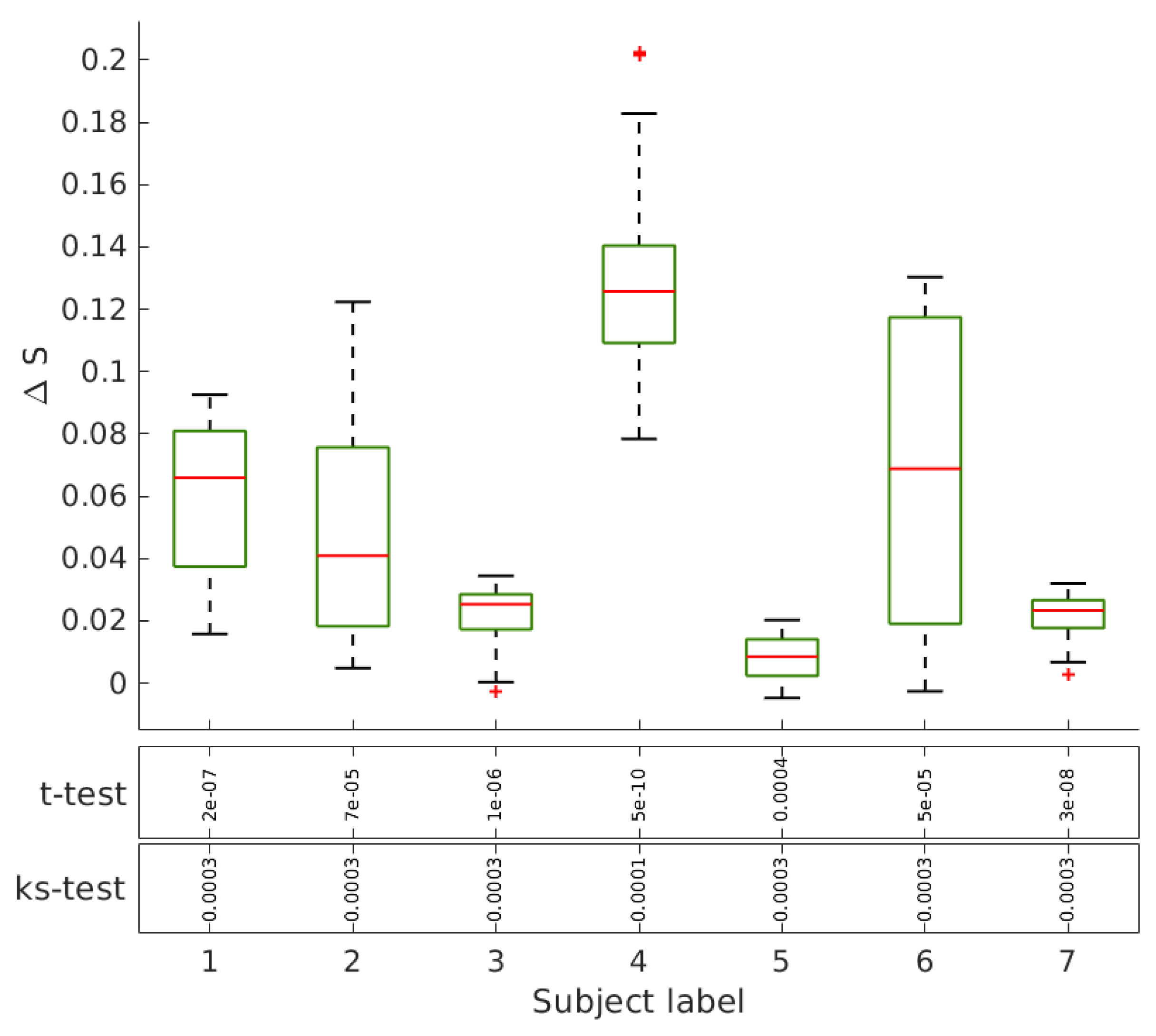
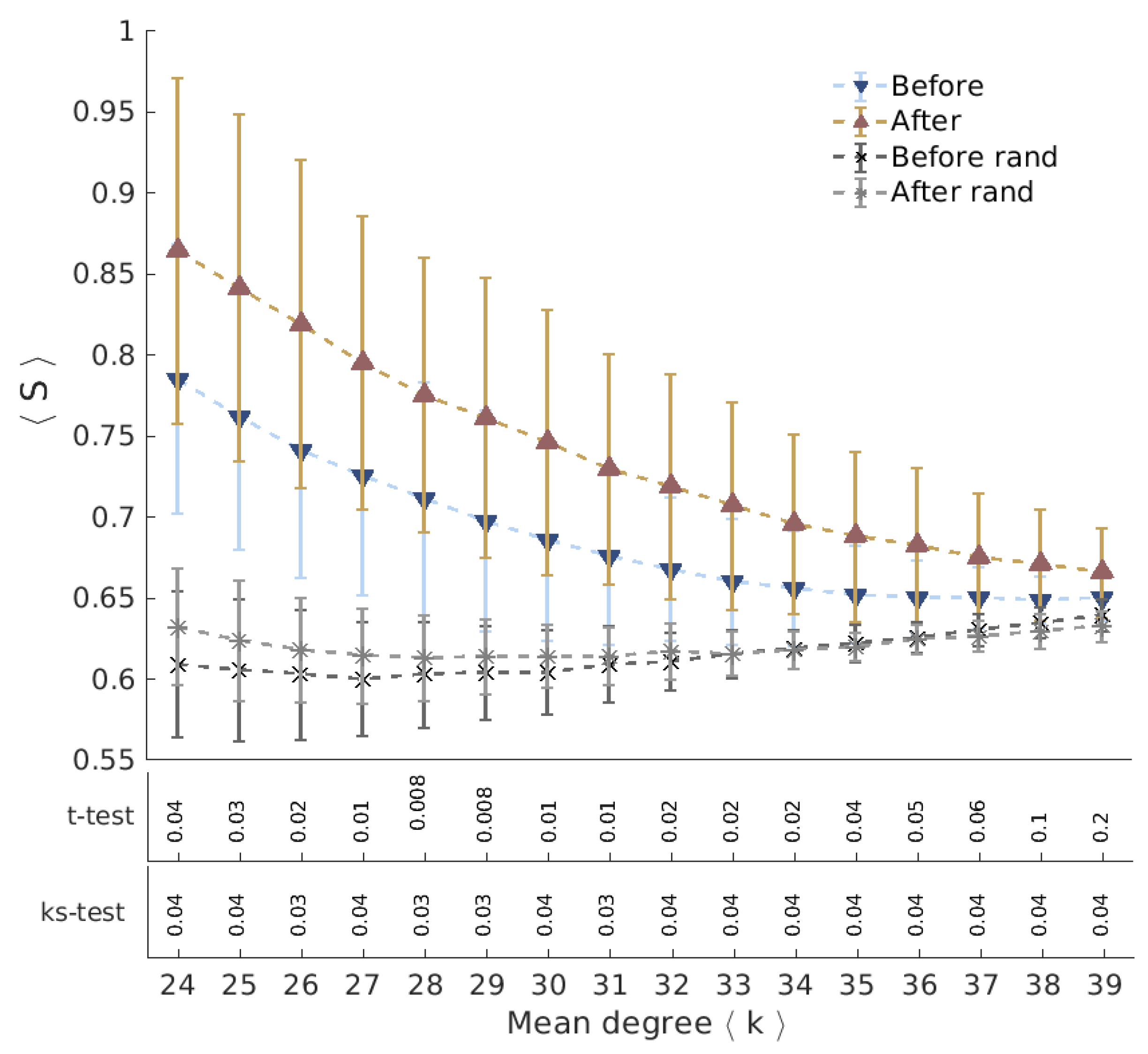
Ayahuasca ingestion may cause deep changes in the cognition and perceptions, promoting substantial alterations in the sense of the reality and the self [
16,
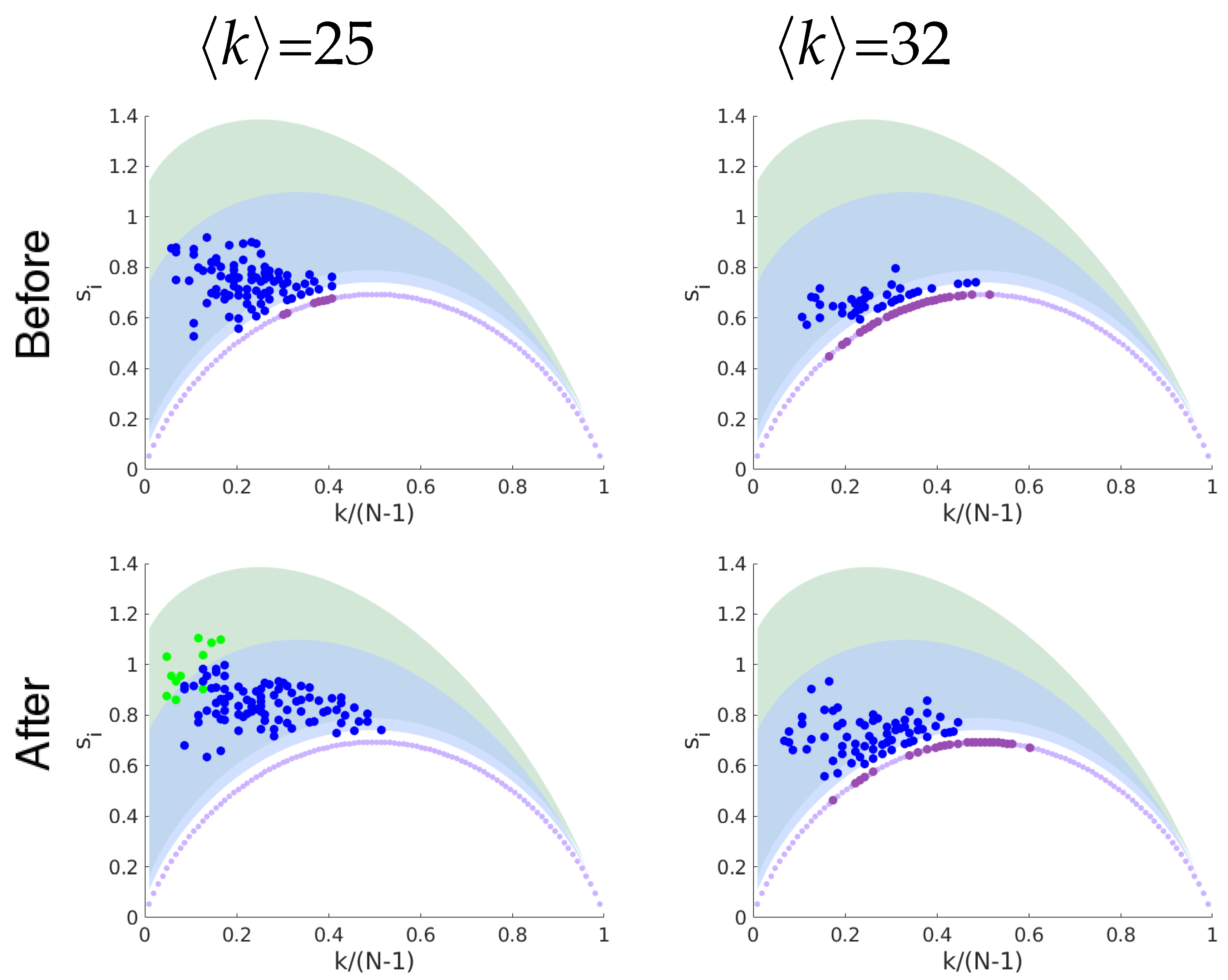
17]. According to the neural correlate hypothesis, we expect to find features of functional brain networks that can be correlated to this specific consciousness state. We evaluate the networks extracted from fMRI data acquired from the same group of subjects in two sections: before and 40 minutes after Ayahuasca intake. The geodesic entropy is able to identify a specific behavior for networks related to the psychedelic state of consciousness: the nodes of functional brain networks under Ayahuasca effects tend to have a greater geodesic entropy than the ordinary condition.
Our most important contribution here, regarding the method, is the geodesic entropy. The mean (or first moment) geodesic distance is a standard and fundamental quantity of interest in the study of complex networks. In addition to the first moment of the geodesic distance, however, there is much insight to be gained from studying other functionals (e.g., higher moments) of its distribution. In this context, one of the most important functionals of a probability distribution is the Shannon entropy. This crucial point motivates our definition of the geodesic entropy. Moreover, we define this entropy for each node. For a network of N nodes, the geodesic entropy of all nodes is an N-dimensional vector. Our second key contribution regarding methodological novelty is the entropy-degree diagram, which is a method for projecting the N-dimensional geodesic distance vectors onto a 2-dimensional plot, with node degree being the second dimension. The entropy-degree diagram is a visual representation of both the connectivity (i.e., degree) as well as the entropy for each node: 2 N-dimensional vectors are projected as N points in 2 dimensions.
This paper is organized as follow: In
Section 2 we define the metric geodesic entropy and introduce the entropy-degree diagram. In
Section 3 we present the results regarding the use of geodesic entropy to comparing functional brain networks of individual before and after Ayahuasca intake. We present our conclusion and discussion in
Section 4. In
Section 5, we give information about the fMRI data and its processing to generate the functional networks.
2. Geodesic Entropy
A complex network is a schematic representation of the relations (links) between elements (nodes) of a system with a nontrivial topology of interactions [
1,
18,
19]. Consider a non-weighted undirected network
, where
V is a set with
N nodes and
E is the set of links. It is represented numerically by a
adjacency matrix
: If a pair of nodes
i and
j are connected, the matrix element is
and
otherwise. The nodes are connected if the elements that they represent share some kind of information, or have potential to share, have some kind of link, correlation or mutual influences. The number of links that have each node is termed degree. The statistics of the degrees in a network is quantified by the degree distribution, a histogram of degrees considering the whole network [
18].
Nodes directly connected are called nearest first neighbors. A node can also influence and be influenced by the neighbors of its neighbors, called second nearest neighbors. Considering a connected network, the influences may be extended to all neighborhood radius. Our goal is to quantify the amount of information involved in the diversity of influence extending over the network. For this purpose, we calculate the Shannon entropy [
8] considering the statistics of distances between a node and all their neighborhood radius.
Distances in network theory are related to the path lengths. By definition, a path length
is the number of consecutive links between the pair of nodes
i and
j, following a specific trail. The shortest path length (
) defines geodesic distance between two nodes [
20]. The geodesic distance has been used in several network characterizations such as small-world networks [
21].
By looking at the distribution of geodesic distances for a given node, we can know how far or near that particular node is to all other nodes and thereby obtain a qualitative understanding of how that node fits into the network overall.
We define
as a probability mass function of find a node in the neighborhood ratio
r of the node
i, that is, the probability that, in a random choice, one selects a node
j from the set of the remaining nodes
with the geodesic distance
. This probability is defined as:
where the neighborhood radius
r assumes values according to the interval
with
. See an illustration in
Figure 1.
The distribution contains information about the connectivity across multiple links of a network. For illustration, consider hypercubic lattices of dimension D with links only between neighboring nodes. The distribution scales according to , because nodes a fixed distance r away lie on the (hyper)surface of constant distance to the node i, where in D dimensions, this surface has dimension . Hence, it makes sense that the characterization of the distribution has the potential to provide insights into network connectivity.
The geodesic entropy is given by:
The value of
does not depend on the network size for larger networks (
. The characteristic geodesic entropy of a network is defined by:
Distinct from the entropy of the degree distribution that quantifies the constraints imposed by the network degree distribution [
22], the geodesic entropy quantifies the information due to the intrinsic configuration of network structure. Networks with different structures can share the same degree distribution, that is, they can be degenerate in the entropy of the degree distribution. The characteristic geodesic entropy can lift the degeneracy of those networks. In this sense, the geodesic entropy is a more appropriate metric for characterizing the interdependence of influences in the network. For a comparison of the geodesic entropy with other similar metrics, we refer the reader to the discussion section.
Entropy-Degree Diagram
Next, we introduce the entropy-degree diagram, a visualization tool to depict each node along two axes: (i) geodesic entropy and (ii) node degree. Entropy-degree diagram is built plotting the geodesic entropy (
)
versus the nodal degree
k normalized by the maximum number of connections possible,
, for all nodes belonging to the network. This normalization allows us to compare networks with different sizes.
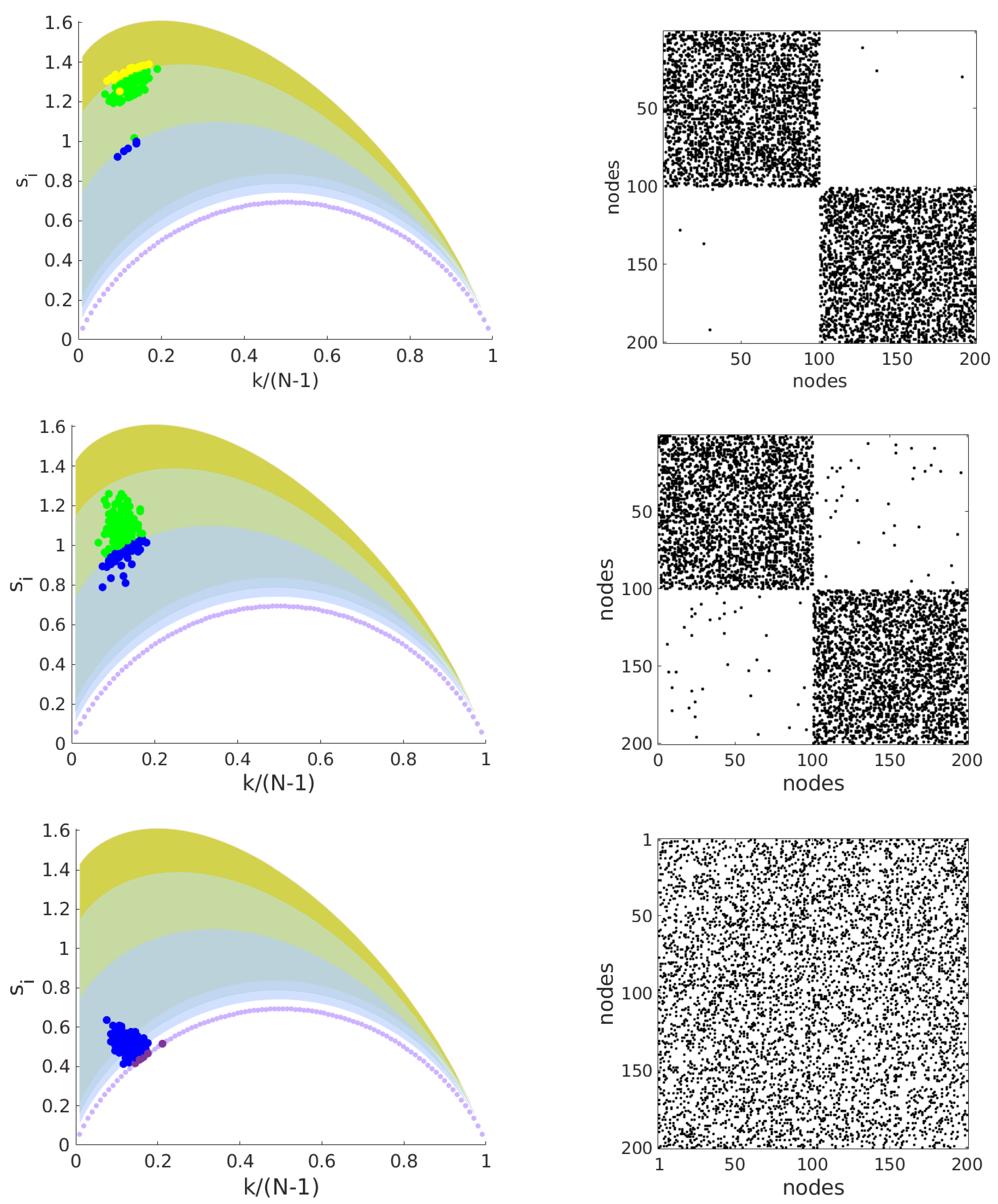
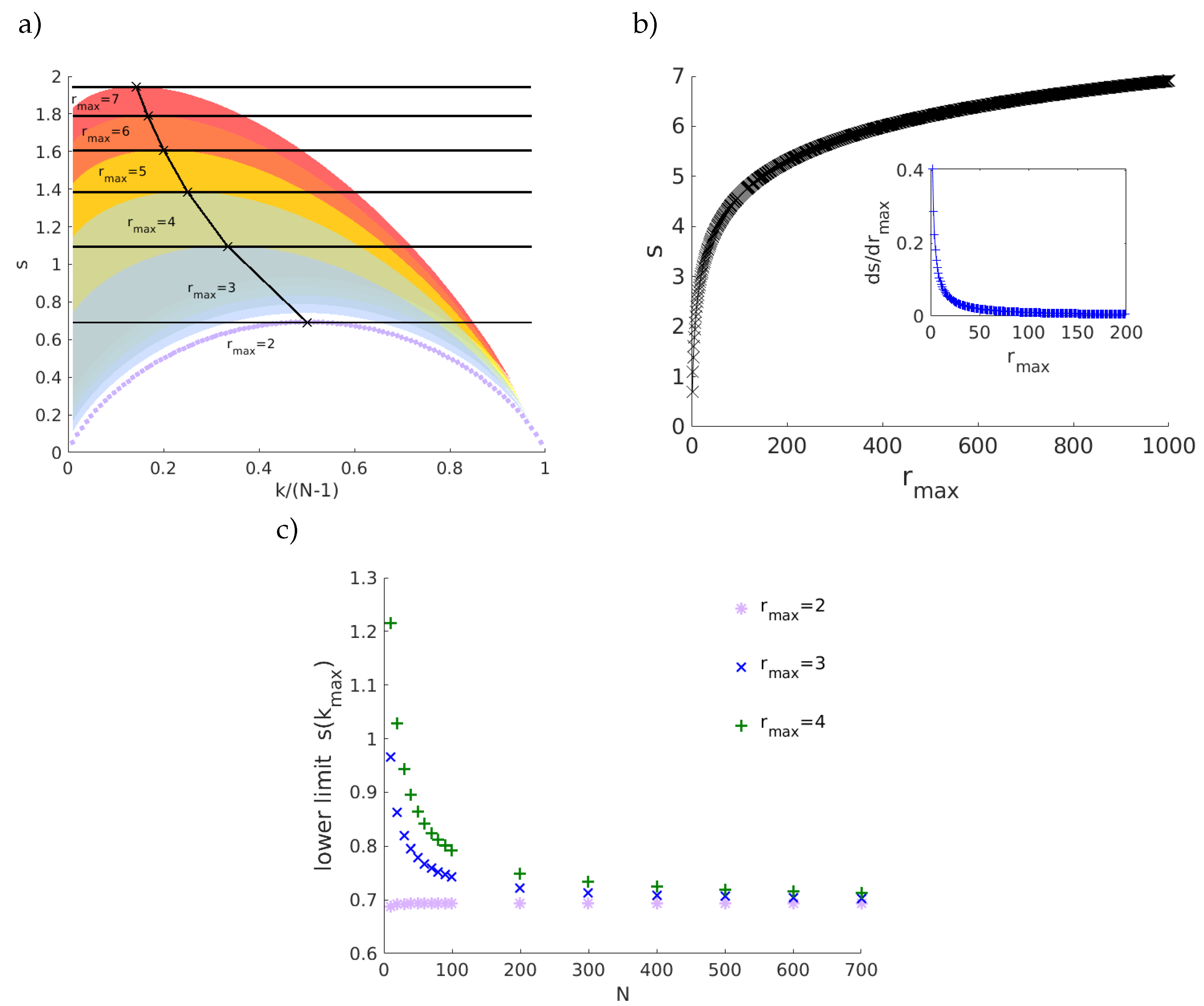
Figure 2 shows the entropy-degree diagram for 3 networks that share the same number of nodes and links, have the same degree distribution but have different structures. Each marker (•) represents a node. Colors are used as a didactic artifact to improve the accessibility of visualization. They correspond to the maximum neighborhood radius (
), that is, the greatest geodesic distance between the given node and the remaining nodes. The shaded regions follow the same colors and mark the space of possibilities for each value of
. For example, the purple curve delimits the possible positions on the diagram for nodes with first and second neighbors. The region in blue indicates the positions for nodes with first, second and third neighbors and it follows for the others regions. The upper bounds are peaked at (
,
). Note the values do not depend on the network size; rather they depend only on the network structure. The magnitude of the increment in the geodesic entropy due to the increase of
is inversely proportional to
, (
. This means that there is a limit in which the increase of maximum geodesic distances (increase the sparsity or large world behavior) contributes significantly to the network entropy. The lower limits will be affected by the size of the network and converge to the first curve (
) for large networks. See
Figure 3. The entropy-degree diagram is a bidimensional representation of the statistics of distances and connectivity of all nodes. Both local and global properties of the entire network are visually represented. Higher geodesic entropy reflects a greater diversity of influences regarding the more homogeneous distribution in the population density across the neighborhood radius.The influence of nodes with high entropy into the networks are more diverse, perhaps more intricate and with greater variability. They are embedded in the structure in a site where the diversity of influences is more smoothly distributed. The overall relation of this node with the remain nodes is less trivial to predict. The opposite can be affirmed to nodes with low entropy.
4. Discussion and Conclusions
In a networked system, the role of each element depends on its relative position inside the overall network architecture. In such a connected system, the elements (represented by nodes) are influenced not only by first neighbors but may also be influenced indirectly by more distant nodes. The geodesic entropy is a statistical quantity that measures, in the frame of reference of a given node, the level of constraints in the (average of the) aggregated influences imposed by the distribution of neighborhood radius.
Functional networks are usually defined according to statistical dependencies between brain regions activities. Assumed as a fully connected network, all pairs of brain regions will have some interdependence. This can be measured directly, by means of the correlation between their activities, or inferred indirectly from the network structure. In this precise sense, the increase of the characteristic geodesic entropy in the functional networks after Ayahuasca intake indicates that the overall interdependencies are less constrained than before Ayahuasca ingestion.
We briefly compare and relate the geodesic entropy to similar quantities that have been used to study networks. The use of geodesic distances to evaluate Shannon entropy was proposed by Chen and collaborators [
24]. Instead of defining the entropy per node, they defined a global entropy considering only one specific value
r of geodesic distance. A recent work from Stella and Domenico proposes to calculate the Shannon entropy of distances to characterize centrality [
25]. Their entropy is insensitive to the maximum neighborhood radius variability, due to normalization in the formula, by the factor log(
). In contrast with the above methods, the geodesic entropy proposed in this work allows to evaluate the statistics of influences diversity for each node in a network accounting the full information of its geometry.
We have evaluated the geodesic entropy of functional brain network of subjects in the resting state before and after the ingestion of the psychedelic brew Ayahuasca. We have found that during Ayahuasca experience nodes of the functional network tend to have greater geodesic entropy than in the ordinary condition, resulting in networks with higher characteristic geodesic entropy. Hence, the geodesic distances between nodes become less constrained on average, i.e., their distribution becomes “wider”. In a previous work, we showed that the entropy of the degree distribution of brain functional connectivity networks under the influence of Ayahuasca is greater than in the ordinary state [
22]. The entropy of degree distribution is a global measurement and networks with different patters can share the same degree distribution. The results presented here suggest that the structural patterns are less constrained under the influence of Ayahuasca compared to the ordinary condition. The geodesic distances are more broadly distributed, thus contributing to a high diversity of influences among the network nodes.
The hypothesis of an entropy increase in response to psychedelic states has been discussed in the literature [
26,
27,
28]. This hypothesis predicts that the psychedelics state is associated with greater entropy compared to the ordinary state. The hypothesis of brain entropy increase claims to explain the increased flexibility in thoughts, facility to access suppressed memory, increase of creativity, among others [
26].
In conclusion, we have shown how the geodesic entropy quantifies locally the overall influences of a network. Furthermore, we have used entropy-degree diagrams to evaluate features of each node in the network, giving a clearer view of the network topology and global connectivity. The application to fMRI-based functional connectivity networks sheds insights on how the brain changes under the influence of external influences. Although in this study we used Ayahuasca, there is in fact no reason why the method cannot be applied to study a variety of drugs, psychopathological symptoms, meditative states, etc. We hope that these ideas and methods find use in furthering our understanding of brain function networks and complex networks in general.

 and
and 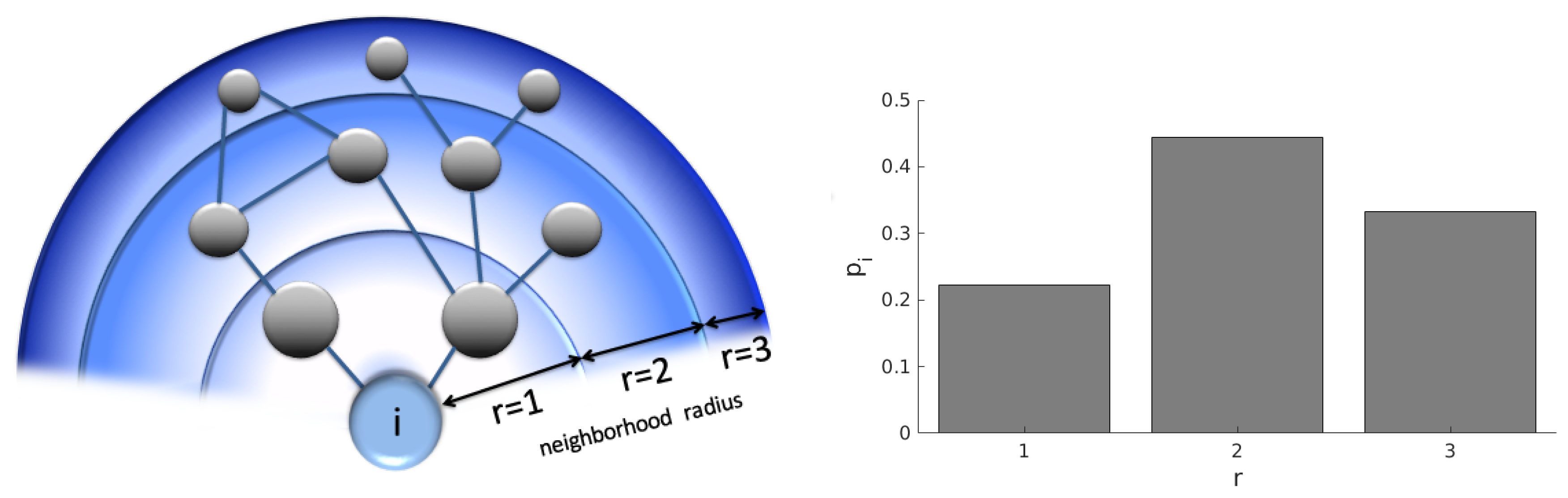{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





