
Human Papillomavirus E7 and p16INK4a mRNA Multiplexed Quantification by a QuantiGeneTM Proof-of-Concept Assay Sensitively Detects Infection and Cervical Dysplasia Severity

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Clinical Study Samples
2.3. HPV DNA Testing
2.4. QuantiGeneTM 2.0 Plex Assay Design and Method Adaptation for QG-POC Assay
2.5. Statistical Analysis
3. Results
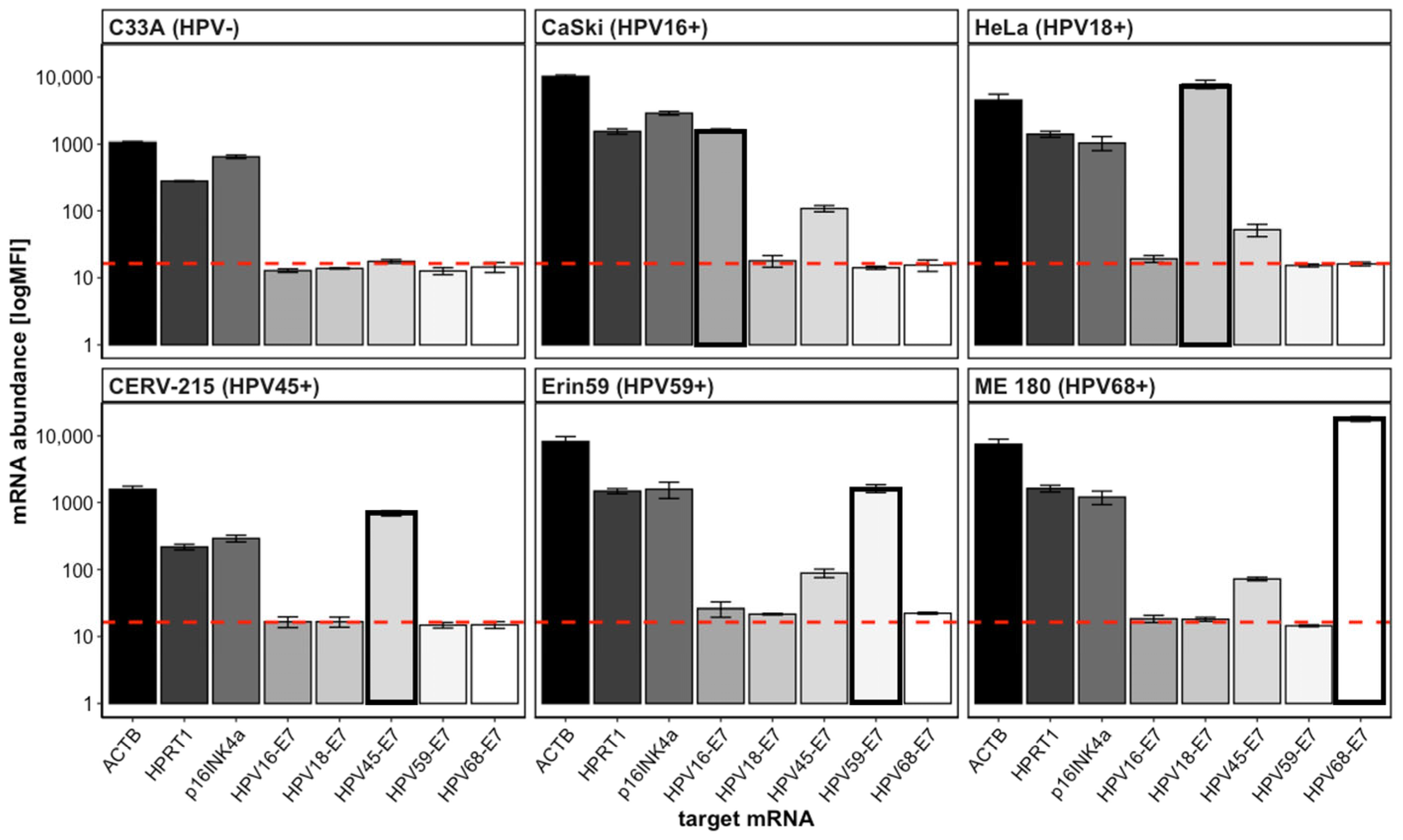
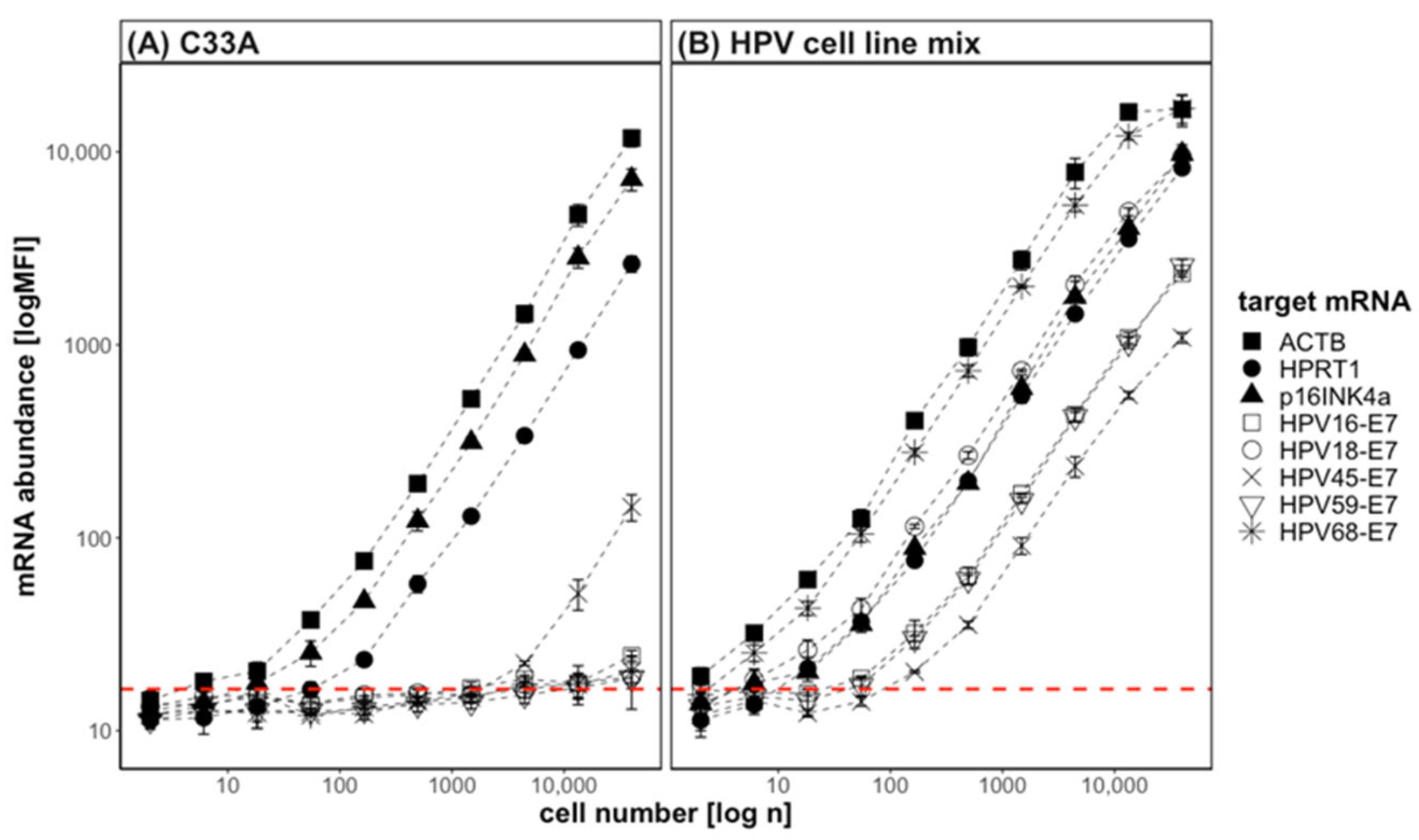
3.1. QG-POC Assay Detects mRNA Targets Specifically with Good Sensitivity in Cell-Line Samples
3.2. QG-POC Assay Detects HPV-E7 mRNA in Clinical Cervical Smear Samples
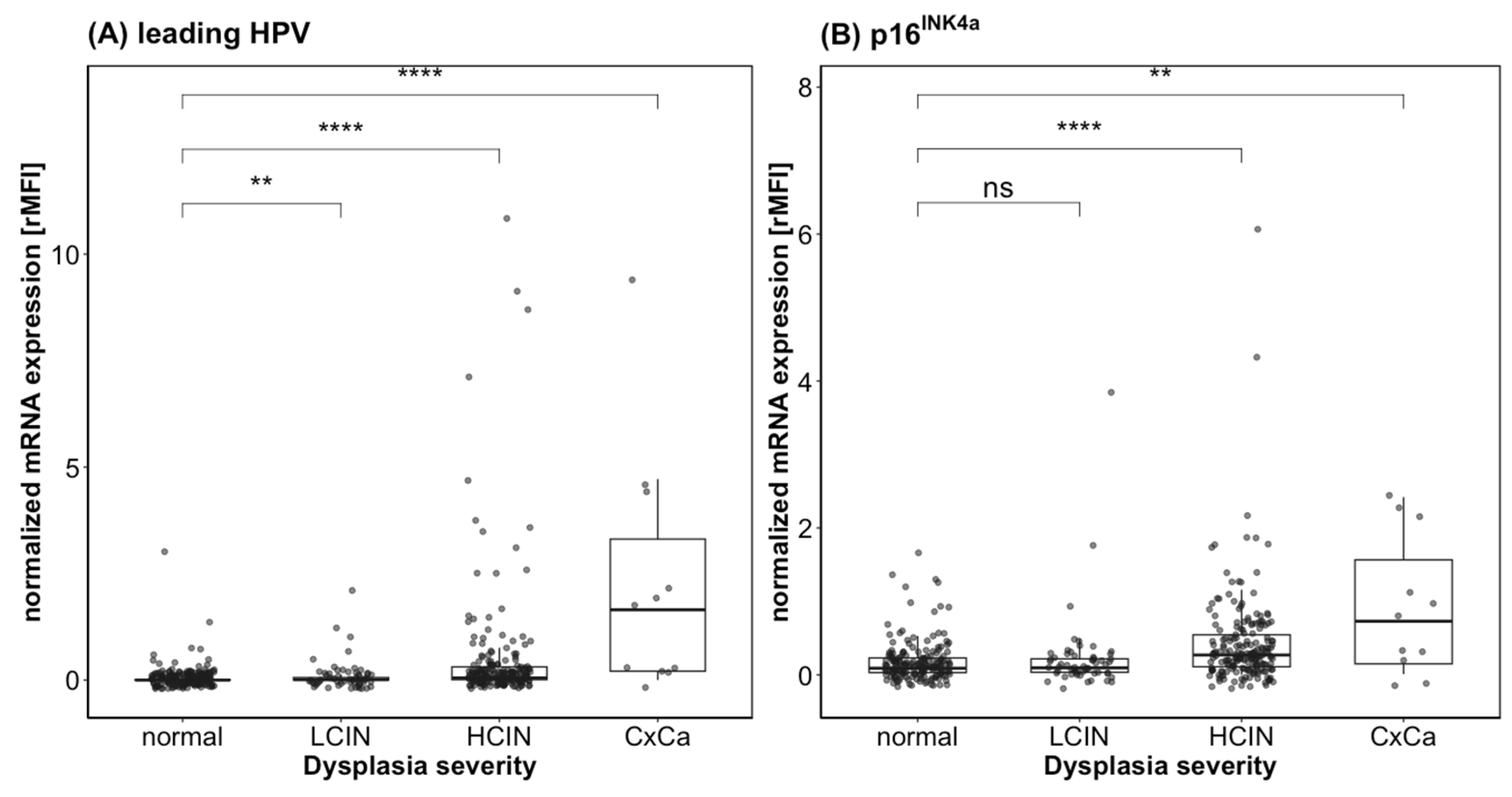
3.3. QG-POC Assay-Reported mRNA Expression Strength Indicates Dysplasia Presence and Severity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer. J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Bosch, F.X.; Qiao, Y.-L.; Castellsagué, X. The epidemiology of human papillomavirusinfection and its association with cervical cancer. Int. J. Gynecol. Obs. 2006, 94, S8–S21. [Google Scholar] [CrossRef] [PubMed]
- De Villiers, E.M.; Fauquet, C.; Broker, T.R.; Bernard, H.U.; zur Hausen, H. Classification of papillomaviruses. Virology 2004, 324, 17–27. [Google Scholar] [CrossRef] [Green Version]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. A Review of Human Carcinogens. IARC Monogr. Eval. Carcinog. Risks Hum. 2012, 100, 1–441. [Google Scholar]
- Dickinson, J.A.; Popadiuk, A.S.C.; Pogany, L.; Onysko, J.; Miller, A.B. Reduced cervical cancer incidence and mortality in Canada: National data from 1932 to 2006. BMC Public Health 2012, 12, 1471–2458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatzistamatiou, K.; Moysiadis, T.; Angelis, E.; Kaufmann, A.; Skenderi, A.; Jansen-Duerr, P.; Lekka, I.; Kilintzis, V.; Angelidou, S.; Katsiki, E.; et al. Diagnostic accuracy of high-risk HPV DNA genotyping for primary cervical cancer screening and triage of HPV-positive women, compared to cytology: Preliminary results of the PIPAVIR study. Arch. Gynecol. Obstet. 2017, 295, 1247–1257. [Google Scholar] [CrossRef]
- Dillner, J. Primary human papillomavirus testing in organized cervical screening. Curr. Opin. Obstet. Gynecol. 2013, 25, 11–16. [Google Scholar] [CrossRef]
- Wright, T.C.; Stoler, M.H.; Behrens, C.M.; Sharma, A.; Zhang, G.; Wright, T.L. Primary cervical cancer screening with human papillomavirus: End of study results from the ATHENA study using HPV as the first-line screening test. Gynecol. Oncol. 2015, 136, 189–197. [Google Scholar] [CrossRef] [Green Version]
- Ronco, G.; Dillner, J.; Elfström, K.M.; Tunesi, S.; Snijders, P.J.F.; Arbyn, M.; Kitchener, H.; Segnan, N.; Gilham, C.; Giorgi-Rossi, P.; et al. Efficacy of HPV-based screening for prevention of invasive cervical cancer: Follow-up of four European randomised controlled trials. Lancet 2014, 383, 524–532. [Google Scholar] [CrossRef]
- Rodriguez, A.C.; Schiffman, M.; Herrero, R.; Wacholder, S.; Hildesheim, A.; Castle, P.E.; Solomon, D.; Burk, R.; Proyecto Epidemiológico Guanacaste Group. Rapid clearance of human papillomavirus and implications for clinical focus on persistent infections. J. Natl. Cancer Inst. 2008, 100, 513–517. [Google Scholar] [CrossRef] [Green Version]
- Ciardiello, F.; Arnold, D.; Casali, P.G.; Cervantes, A.; Douillard, J.Y.; Eggermont, A.; Eniu, A.; McGregor, K.; Peters, S.; Piccart, M.; et al. Delivering precision medicine in oncology today and in future-the promise and challenges of personalised cancer medicine: A position paper by the European Society for Medical Oncology (ESMO). Ann. Oncol. 2014, 25, 1673–1678. [Google Scholar] [CrossRef]
- Wentzensen, N.; von Knebel Doeberitz, M. Biomarkers in cervical cancer screening. Dis. Markers 2007, 23, 315–330. [Google Scholar] [CrossRef]
- Solomon, D.; Davey, D.; Kurman, R.; Moriarty, A.; O’Connor, D.; Prey, M.; Raab, S.; Sherman, M.; Wilbur, D.; Wright, T., Jr.; et al. The 2001 Bethesda System: Terminology for Reporting Results of Cervical Cytology. JAMA 2002, 287, 2114–2119. [Google Scholar] [CrossRef]
- Schmitt, M.; Dalstein, V.; Waterboer, T.; Clavel, C.; Gissmann, L.; Pawlita, M. The HPV16 transcriptome in cervical lesions of different grades. Mol. Cell Probes 2011, 25, 260–265. [Google Scholar] [CrossRef]
- Ishikawa, M.; Fujii, T.; Saito, M.; Nindly, I.; Ono, A.; Kubushiro, K.; Tsukazaki, K.; Mukai, M.; Nozawa, S. Overexpression of p16INK4a as an indicator for human papillomavirus oncogenic activity in cervical squamous neoplasia. Int. J. Gynecol. Cancer 2006, 16, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Scheffner, M.; Huibregtse, J.M.; Vierstra, R.D.; Howley, P.M. The HPV-16 E6 and E6-AP Complex Functions as a Ubiquitin-Protein Ligase in the Ubiquitination of p53. Cell 1993, 75, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Boyer, S.N.; Wazer, D.E.; Band, V. E7 Protein of Human Papilloma Virus-16 Induces Degradation of Retinoblastoma Protein through the Ubiquitin-Proteasome Pathway. Cancer Res. 1996, 56, 4620–4624. [Google Scholar]
- Szymonowicz, K.A.; Chen, J. Biological and clinical aspects of HPV-related cancers. Cancer Biol. Med. 2020, 17, 864–878. [Google Scholar] [CrossRef] [PubMed]
- Del Pino, M.; Garcia, S.; Fuste, V.; Alonso, I.; Fuste, P.; Torne, A.; Ordi, J. Value of p16(INK4a) as a marker of progression/regression in cervical intraepithelial neoplasia grade 1. Am. J. Obstet. Gynecol. 2009, 201, 488.E1–488.E7. [Google Scholar] [CrossRef] [PubMed]
- Wentzensen, N.; Schiffman, M.; Palmer, T.; Arbyn, M. Triage of HPV positive women in cervical cancer screening. J. Clin. Virol. 2016, 76, S49–S55. [Google Scholar] [CrossRef] [Green Version]
- Flagella, M.; Bui, S.; Zheng, Z.; Nguyen, C.T.; Zhang, A.; Pastor, L.; Ma, Y.; Yang, W.; Crawford, K.L.; McMaster, G.K.; et al. A multiplex branched DNA assay for parallel quantitative gene expression profiling. Anal. Biochem. 2006, 352, 50–60. [Google Scholar] [CrossRef]
- Dunbar, S.A. Applications of Luminex xMAP technology for rapid, high-throughput multiplexed nucleic acid detection. Clin. Chim. Acta 2006, 363, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Lehr, E.E.; Qadadri, B.; Brown, C.R.; Brown, D.R. Human papillomavirus type 59 immortalized keratinocytes express late viral proteins and infectious virus after calcium stimulation. Virology 2003, 314, 562–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agorastos, T.; Chatzistamatiou, K.; Moysiadis, T.; Kaufmann, A.M.; Skenderi, A.; Lekka, I.; Koch, I.; Soutschek, E.; Boecher, O.; Kilintzis, V.; et al. Human papillomavirus E7 protein detection as a method of triage to colposcopy of HPV positive women, in comparison to genotyping and cytology. Final results of the PIPAVIR study. Int. J. Cancer 2017, 141, 519–530. [Google Scholar] [CrossRef] [Green Version]
- Agorastos, T.; Koutikas, V.; Falelakis, M.; Lekka, I.; Mikos, T.; Delopoulos, A.; Mitkas, P.A.; Tantsis, A.; Weyers, S.; Coorevits, P.; et al. Semantic Integration of Cervical Cancer Data Repositories to Facilitate Multicenter Association Studies: The ASSIST Approach. Cancer Inform. 2009, 8, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, M.; Bravo, I.G.; Snijders, P.J.; Gissmann, L.; Pawlita, M.; Waterboer, T. Bead-based multiplex genotyping of human papillomaviruses. J. Clin. Microbiol. 2006, 44, 504–512. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, M.; Dondog, B.; Waterboer, T.; Pawlita, M. Homogeneous amplification of genital human alpha papillomaviruses by PCR using novel broad-spectrum GP5+ and GP6+ primers. J. Clin. Microbiol. 2008, 46, 1050–1059. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Wickham, H.; François, R.; Henry, L.; Müller, K. dplyr: A Grammar of Data Manipulation; Posit, PBC: Boston, MA, USA, 2020. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Cohen, J. A Power Primer. Quant. Methods Psychol. 1992, 112, 155–159. [Google Scholar] [CrossRef]
- Nakazawa, M. fmsb: Functions for Medical Statistics Book with some Demographic Data; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Landis, J.R.; Koch, G.G. The Measurement of Observer Agreement for Categorical Data. Biometrics 2012, 33, 159–174. [Google Scholar] [CrossRef] [Green Version]
- Robin, X.; Turck, N.; Hainard, A.; Tiberti, N.; Lisacek, F.; Sanchez, J.-C.; Müller, M. pROC: An open-source package for R and S+ to analyze and compare ROC curves. BMC Bioinform. 2011, 12, 77. [Google Scholar] [CrossRef]
- Sing, T.; Sander, O.; Beerenwinkel, N.; Lengauer, T. ROCR: Visualizing classifier performance in R. Bioinformatics 2005, 21, 7881. [Google Scholar] [CrossRef] [PubMed]
- Wang-Johanning, F.; Lu, D.W.; Wang, Y.; Johnson, M.R.; Johanning, G.L. Quantitation of human papillomavirus 16 E6 and E7 DNA and RNA in residual material from ThinPrep Papanicolaou tests using real-time polymerase chain reaction analysis. Cancer 2002, 94, 2199–2210. [Google Scholar] [CrossRef]
- Quint, W.; Jenkins, D.; Molijn, A.; Struijk, L.; van de Sandt, M.; Doorbar, J.; Mols, J.; Van Hoof, C.; Hardt, K.; Struyf, F.; et al. One virus, one lesion--individual components of CIN lesions contain a specific HPV type. J. Pathol. 2012, 227, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Scerri, J.; Baldacchino, S.; Saliba, C.; Scerri, C.; Grech, G. Bead-based RNA multiplex panels for biomarker detection in oncology samples. Methods 2018, 158, 86–91. [Google Scholar] [CrossRef]
- McCredie, M.R.E.; Sharples, K.J.; Paul, C.; Baranyai, J.; Medley, G.; Jones, R.W.; Skegg, D.C.G. Natural history of cervical neoplasia and risk of invasive cancer in women with cervical intraepithelial neoplasia 3: A retrospective cohort study. Lancet Oncol. 2008, 9, 425–434. [Google Scholar] [CrossRef]
- Longuet, M.; Beaudenon, S.; Orth, G. Two Novel Genital Human Papillomavirus (HPV) Types, HPV68 and HPV70, Related to the Potentially Oncogenic HPV39. J. Clin. Microbiol. 1996, 34, 738–744. [Google Scholar] [CrossRef] [Green Version]
- Jaworek, H.; Kubanova, K.; Koudelakova, V.; Slavkovsky, R.; Drabek, J.; Hajduch, M. Pitfalls of commercially available HPV tests in HPV68a detection. PLoS ONE 2019, 14, e0220373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klaes, R.; Woerner, S.M.; Ridder, R.; Wentzensen, N.; Duerst, M.; Schneider, A.; Lotz, B.; Melsheimer, P.; von Knebel Doeberitz, M. Detection of High-Risk Cervical Intraepithelial Neoplasia and Cervical Cancer by Amplification of Transcripts Derived from Integrated Papillomavirus Oncogenes1. Cancer Res. 1999, 59, 6132–6136. [Google Scholar] [PubMed]
- Von Keyserling, H.; Kuhn, W.; Schneider, A.; Bergmann, T.; Kaufmann, A.M. p16INK(4)a and p14ARF mRNA expression in Pap smears is age-related. Mod. Pathol. 2012, 25, 465–470. [Google Scholar] [CrossRef] [Green Version]
- Santin, A.D.; Zhan, F.; Bignotti, E.; Siegel, E.R.; Cane, S.; Bellone, S.; Palmieri, M.; Anfossi, S.; Thomas, M.; Burnett, A.; et al. Gene expression profiles of primary HPV16- and HPV18-infected early-stage cervical cancers and normal cervical epithelium: Identification of novel candidate molecular markers for cervical cancer diagnosis and therapy. Virology 2005, 331, 269–291. [Google Scholar] [CrossRef] [Green Version]
- Goulart, A.P.S.; Goncalves, M.A.G.; Da-Silva, V.D. Evaluation of Telomerase (hTert), Ki67 and p16ink4a expressions in low and high-grade cervical intraepithelial lesions. Rev. Col. Bras. Cir. 2017, 44, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Clad, A.; Reuschenbach, M.; Weinschenk, J.; Grote, R.; Rahmsdorf, J.; Freudenberg, N. Performance of the Aptima high-risk human papillomavirus mRNA assay in a referral population in comparison with Hybrid Capture 2 and cytology. J. Clin. Microbiol. 2011, 49, 1071–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szarewski, A.; Mesher, D.; Cadman, L.; Austin, J.; Ashdown-Barr, L.; Ho, L.; Terry, G.; Liddle, S.; Young, M.; Stoler, M.; et al. Comparison of seven tests for high-grade cervical intraepithelial neoplasia in women with abnormal smears: The Predictors 2 study. J. Clin. Microbiol. 2012, 50, 1867–1873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorji, N.; Tshering, S.; Choden, S.; Chhetri, M.; Bhujel, D.; Wangden, T.; Pradhan, B.; Bhutia, P.C.; Tshomo, U. Evaluation of the diagnostic performance of colposcopy in the diagnosis of histologic cervical intraepithelial neoplasia 2+ (CIN2+). BMC Cancer 2022, 22, 930. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target mRNA | Gene Accession Number |
|---|---|
| HPV6-E7 | L41216 |
| HPV16-E7 | K02718 |
| HPV18-E7 | X05015 |
| HPV31-E7 | J04353 |
| HPV45-E7 | X74479 |
| HPV59-E7 | EU918767 |
| HPV68-E7 | Y14591 |
| ACTB | NM_001101 |
| HPRT1 | NM_000077 |
| p16INK4a | NM_000194 |
| ASSIST Score 1 | Clinical Group | HPV Genotype 2 (n (%)) | No HPV Detectable | |||||
|---|---|---|---|---|---|---|---|---|
| 16 | 18 | 31 | 59 | 68 | 6 | |||
| 0 | Normal (n = 177) | 113 (52.6) | 15 (6.9) | 6 (2.8) | 26 (12.1) | 2 (0.9) | 5 (2.3) | 41 (23.2) |
| 1 | LCIN (n = 58) | 48 (63.2) | 6 (7.9) | 8 (10.5) | 5 (8.6) | 4 (5.3) | 2 (2.6) | 0 (0) |
| 2 | HCIN (n = 195) | 151 (62.7) | 25 (10.4) | 24 (10.0) | 22 (9.1) | 4 (1.7) | 5 (2.1) | 0 (0) |
| 3 | CxCa (n = 11) | 10 (90.9) | 1 (9.1) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| total | 441 | 322 (73.0) | 47 (10.7) | 38 (8.6) | 53 (12.0) | 8 (1.8) | 12 (2.7) | 41 (9.1) |
| ASSIST Score 1 | Clinical Group | HPV Genotype * (n (%)) | No HPV Detectable | |||||
|---|---|---|---|---|---|---|---|---|
| 16 | 18 | 31 | 59 | 68 | 6 | |||
| 0 | Normal (n = 177) | 46 (26.0) | 8 (4.5) | 11 (6.2) | 16 (9.0) | 3 (1.7) | 5 (2.8) | 106 (60.0) |
| 1 | LCIN (n = 58) | 27 (46.6) | 4 (6.9) | 5 (8.6) | 12 (20.7) | 2 (3.4) | 3 (5.1) | 21 (36.2) |
| 2 | HCIN (n = 195) | 113 (57.9) | 20 (10.3) | 26 (13.3) | 23 (11.8) | 4 (2.1) | 6 (3.1) | 44 (22.6) |
| 3 | CxCa (n = 11) | 8 (72.7) | 2 (18.2) | 0 (0.0) | 3 (27.3) | 1 (9.1) | 0 (0.0) | 1 (9.1) |
| total | 441 | 194 (44.0) | 34 (7.8) | 42 (9.5) | 54 (12.2) | 10 (2.3) | 14 (3.2) | 172 (39.0) |
| Cohen’s Kappa | ||||
|---|---|---|---|---|
| HPV Type | Concordance (%) | Κ | CI (95%) | Agreement + |
| 6 | 98.41 | 0.66 *** | 0.44–0.87 | Substantial |
| 16 | 65.53 | 0.35 *** | 0.26–0.43 | Fair |
| 18 | 95.46 | 0.73 *** | 0.61–0.84 | Substantial |
| 31 | 93.42 | 0.60 *** | 0.46–0.74 | Moderate |
| 59 | 79.37 | 0.03 | −0.14–0.20 | Slight |
| 68 | 95.92 | −0.02 | −0.48–0.44 | No |
| QG-POC HPV genotypes # | 70.29 | 0.27 *** | 0.17–0.38 | Fair |
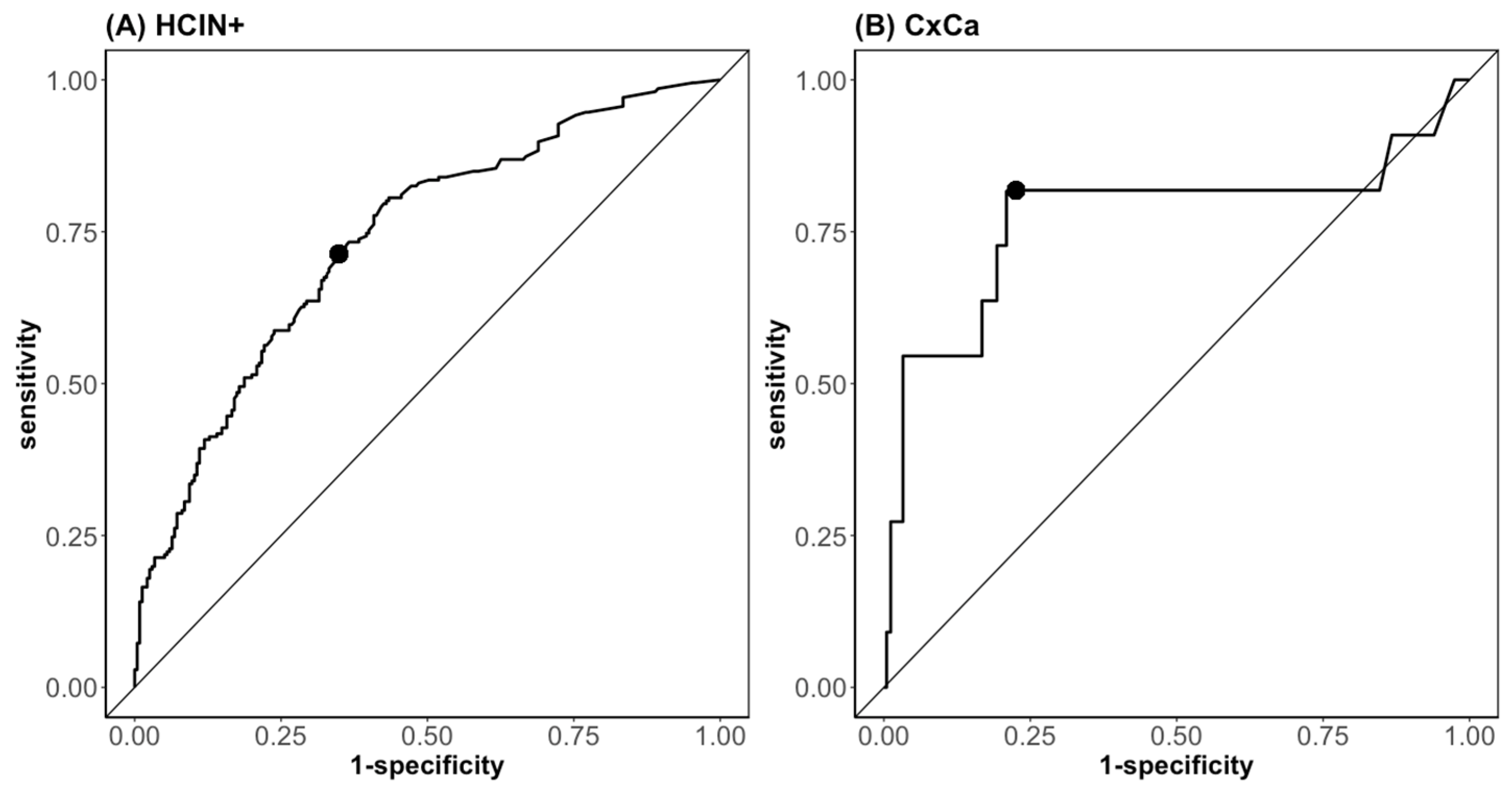
| Biomarker | AUC (CI 95%) | Cut-off | Sensitivity (%) | Specificity (%) | Univariate Regression OR (CI 95%) | Multivariate Regression OR (CI 95%) |
|---|---|---|---|---|---|---|
| HPV16-E7 | 0.68 (0.63–0.72) | 0.025 | 47.6 | 84.7 | 6.03 *** (2.55–18.37) | - |
| HPV18-E7 | 0.54 (0.50–0.57) | 0.015 | 16.9 | 90.2 | 3.13 (1.29–18.89) | - |
| HPV31-E7 | 0.53 (0.49–0.57) | 0.005 | 17.5 | 88.9 | 1.30 (0.22–9.57) | - |
| HPV59-E7 | 0.50 (0.48–0.53) | 0.005 | 1.9 | 100 | 3.88 (0.85–140.64) | - |
| Leading HPV-E7 | 0.74 (0.70–0.79) | 0.490 | 63.6 | 75.5 | 4.53 *** (2.44–9.78) | 2.78 ** (1.55–5.97) |
| p16INK4a | 0.71 (0.66–0.76) | 0.175 | 64.0 | 68.9 | 5.81 *** (3.04–12.05) | 3.31 ** (1.64–7.13) |
| Biomarker | AUC (CI 95%) | Cut-off | Sensitivity (%) | Specificity (%) | Univariate Regression OR (CI 95%) | Multivariate Regression OR (CI 95%) |
|---|---|---|---|---|---|---|
| Leading HPV-E7 | 0.79 (0.62–0.97) | 0.185 | 81.8 | 78.6 | 1.51 *** (1.21–1.88) | 1.49 * (1.06–2.01) |
| p16INK4a | 0.69 (0.48–0.91) | 0.725 | 54.5 | 90.2 | 2.08 ** (1.19–3.56) | 1.06 (0.48–2.31) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skof, A.S.; Rotenberg, L.; Hannemann, P.V.F.; Thies, S.; Boschetti-Grützmacher, E.; Kaufmann, A.M. Human Papillomavirus E7 and p16INK4a mRNA Multiplexed Quantification by a QuantiGeneTM Proof-of-Concept Assay Sensitively Detects Infection and Cervical Dysplasia Severity. Diagnostics 2023, 13, 1135. https://doi.org/10.3390/diagnostics13061135
Skof AS, Rotenberg L, Hannemann PVF, Thies S, Boschetti-Grützmacher E, Kaufmann AM. Human Papillomavirus E7 and p16INK4a mRNA Multiplexed Quantification by a QuantiGeneTM Proof-of-Concept Assay Sensitively Detects Infection and Cervical Dysplasia Severity. Diagnostics. 2023; 13(6):1135. https://doi.org/10.3390/diagnostics13061135
Chicago/Turabian StyleSkof, Anna Sophie, Lina Rotenberg, Paul Viktor Felix Hannemann, Sarah Thies, Eleonora Boschetti-Grützmacher, and Andreas M. Kaufmann. 2023. "Human Papillomavirus E7 and p16INK4a mRNA Multiplexed Quantification by a QuantiGeneTM Proof-of-Concept Assay Sensitively Detects Infection and Cervical Dysplasia Severity" Diagnostics 13, no. 6: 1135. https://doi.org/10.3390/diagnostics13061135