
Dysregulation of miRNAs in DLBCL: Causative Factor for Pathogenesis, Diagnosis and Prognosis

 , ,
, , {kind=link}
{kind=link}
Abstract
:1. Introduction
2. miRNA Biogenesis
3. Delivery Platforms for miRNA
3.1. Viral Vectors
3.2. Non-Viral Vectors
4. Diffuse Large B-Cell Lymphoma
5. Etiology of DLBCL
6. Epigenetic Modifications in DLBCL
7. Immune Escape Pathways in DLBCL
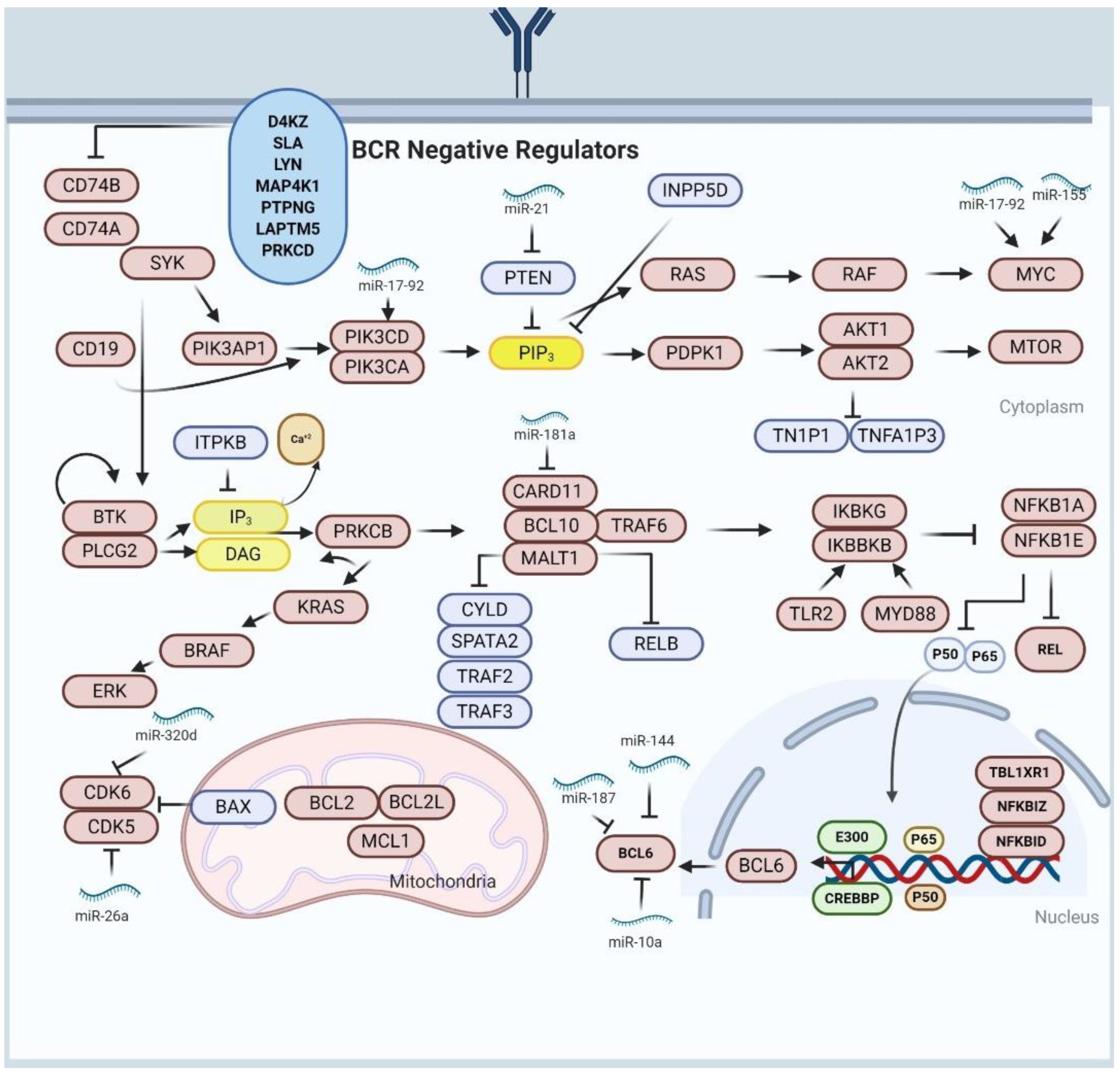
8. Alterations of NF-κβ Activity and BCR Signaling in DLBCL
9. Impaired BCL6 Activity in DLBCL
10. Blocking the Terminal Differentiation Pathway in DLBCL
11. Role of miRNA in DLBCL Pathogenesis
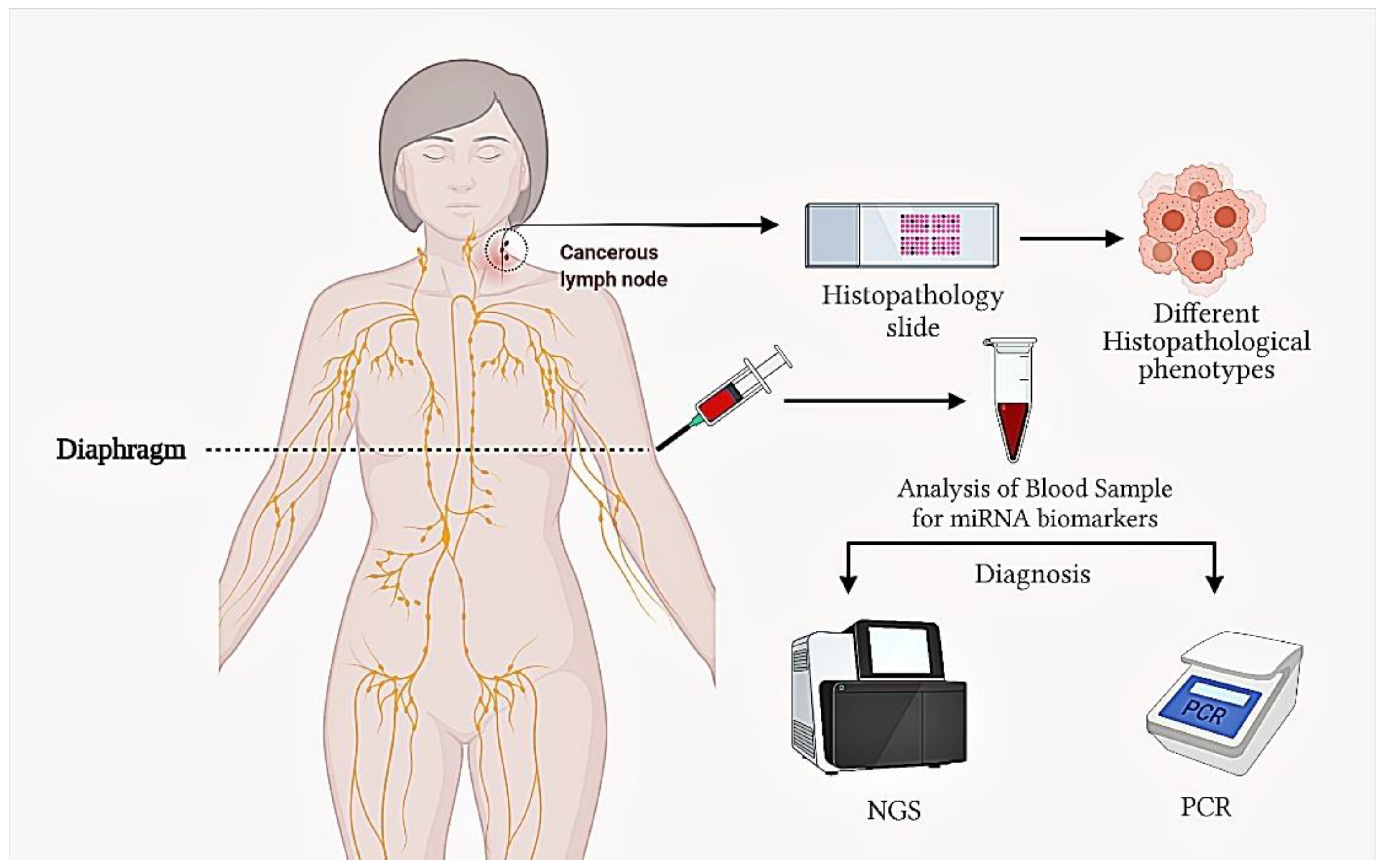
12. The Role of miRNAs in Diagnosis and Prognosis in DLBCL
13. Treatment
14. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vishnoi, A.; Rani, S. MiRNA Biogenesis and Regulation of Diseases: An Overview. In MicroRNA Profiling; Springer: Berlin/Heidelberg, Germany, 2017; pp. 1–10. [Google Scholar]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Joga, M.R.; Zotti, M.J.; Smagghe, G.; Christiaens, O. RNAi efficiency, systemic properties, and novel delivery methods for pest insect control: What we know so far. Front. Physiol. 2016, 7, 553. [Google Scholar] [CrossRef] [Green Version]
- Michlewski, G.; Cáceres, J.F. Post-transcriptional control of miRNA biogenesis. Rna 2019, 25, 1–6. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of microRNA biogenesis, mechanisms of actions, and circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Hammond, S.M.; Bernstein, E.; Beach, D.; Hannon, G.J. An RNA-directed nuclease mediates post-transcriptional gene silencing in Drosophila cells. Nature 2000, 404, 293–296. [Google Scholar] [CrossRef] [PubMed]
- Treiber, T.; Treiber, N.; Meister, G. Regulation of microRNA biogenesis and its crosstalk with other cellular pathways. Nat. Rev. Mol. Cell Biol. 2019, 20, 5–20. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Song, Y.; Shi, X.; Liu, J.; Xiong, S.; Chen, W.; Fu, Q.; Huang, Z.; Gu, N.; Zhang, R. The landscape of miRNA editing in animals and its impact on miRNA biogenesis and targeting. Genome Res. 2018, 28, 132–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daugaard, I.; Hansen, T.B. Biogenesis and function of ago-associated RNAs. Trends Genet. 2017, 33, 208–219. [Google Scholar] [CrossRef] [PubMed]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Greene, J.; Baird, A.M.; Brady, L.; Lim, M.; Gray, S.G.; McDermott, R.; Finn, S.P. Circular RNAs: Biogenesis, function and role in human diseases. Front. Mol. Biosci. 2017, 4, 38. [Google Scholar] [CrossRef]
- Vitsios, D.M.; Davis, M.P.; van Dongen, S.; Enright, A.J. Large-scale analysis of microRNA expression, epi-transcriptomic features and biogenesis. Nucleic Acids Res. 2017, 45, 1079–1090. [Google Scholar] [CrossRef] [Green Version]
- Asadi, M.; Shanehbandi, D.; Zafari, V.; Khaze, V.; Somi, M.H.; Hashemzadeh, S. Transcript level of MicroRNA processing elements in gastric cancer. J. Gastrointest. Cancer 2019, 50, 855–859. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Ahn, C.; Han, J.; Choi, H.; Kim, J.; Yim, J.; Lee, J.; Provost, P.; Rådmark, O.; Kim, S.; et al. The nuclear RNase III Drosha initiates microRNA processing. Nature 2003, 425, 415–419. [Google Scholar] [CrossRef]
- Xu, H.; Wang, X.; Wang, Z.; Li, J.; Xu, Z.; Miao, M.; Chen, G.; Lei, X.; Wu, J.; Shi, H.; et al. MicroRNA expression profile analysis in sperm reveals hsa-mir-191 as an auspicious omen of in vitro fertilization. BMC Genom. 2020, 21, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auyeung, V.C.; Ulitsky, I.; McGeary, S.E.; Bartel, D.P. Beyond secondary structure: Primary-sequence determinants license pri-miRNA hairpins for processing. Cell 2013, 152, 844–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, D.C.; Carneiro, F.D.; Almeida, K.C.; Fernandes-Santos, C. Role of miRNAs on the pathophysiology of cardiovascular diseases. Arq. Bras. Cardiol. 2018, 111, 738–746. [Google Scholar] [CrossRef] [PubMed]
- Selvarajan, S.; Vijayaraghavan, J.; Bobby, Z.; Ramalingam, J.; Porur, C. Micro RNAs—A review. J. Evol. Med. Dent. Sci. 2019, 8, 2918–2923. [Google Scholar] [CrossRef]
- Lund, E.; Dahlberg, J. (Eds.) Substrate Selectivity of Exportin 5 and Dicer in the Biogenesis of microRNAs. Cold Spring Harbor Symposia on Quantitative Biology; Cold Spring Harbor Laboratory Press: Long Island, NY, USA, 2006. [Google Scholar]
- Shomron, N.; Levy, C. MicroRNA-biogenesis and Pre-mRNA splicing crosstalk. J. Biomed. Biotechnol. 2009, 2009, 594678. [Google Scholar] [CrossRef] [PubMed]
- Nayerossadat, N.; Maedeh, T.; Ali, P.A. Viral and non-viral delivery systems for gene delivery. Adv. Biomed. Res. 2012, 1, 27. [Google Scholar] [CrossRef]
- Kamimura, K.; Suda, T.; Zhang, G.; Liu, D. Advances in gene delivery systems. Pharm. Med. 2011, 25, 293–306. [Google Scholar] [CrossRef]
- Andersson, M.G.; Haasnoot, P.J.; Xu, N.; Berenjian, S.; Berkhout, B.; Akusjaärvi, G. Suppression of RNA interference by adenovirus virus-associated RNA. J. Virol. 2005, 79, 9556–9565. [Google Scholar] [CrossRef] [Green Version]
- Lukac, D.M.; Garibyan, L.; Kirshner, J.R.; Palmeri, D.; Ganem, D. DNA binding by Kaposi’s sarcoma-associated herpesvirus lytic switch protein is necessary for transcriptional activation of two viral delayed early promoters. J. Virol. 2001, 75, 6786–6799. [Google Scholar] [CrossRef] [Green Version]
- Schultz, B.R.; Chamberlain, J.S. Recombinant adeno-associated virus transduction and integration. Mol. Ther. 2008, 16, 1189–1199. [Google Scholar] [CrossRef]
- Dhungel, B.P.; Bailey, C.G.; Rasko, J.E. Journey to the center of the cell: Tracing the path of AAV transduction. Trends Mol. Med. 2021, 27, 172–184. [Google Scholar] [CrossRef]
- Yang, Y.S.; Xie, J.; Chaugule, S.; Wang, D.; Kim, J.M.; Kim, J.; Tai, P.W.; Seo, S.K.; Gravallese, E.; Gao, G.; et al. Bone-targeting AAV-mediated gene silencing in osteoclasts for osteoporosis therapy. Mol. Ther. Methods Clin. Dev. 2020, 17, 922–935. [Google Scholar] [CrossRef]
- Liu, Y.P.; Berkhout, B. miRNA cassettes in viral vectors: Problems and solutions. Biochim. Biophys. Acta Gene Regul. Mech. 2011, 1809, 732–745. [Google Scholar] [CrossRef] [PubMed]
- Ye, D.; Wang, G.; Liu, Y.; Huang, W.; Wu, M.; Zhu, S.; Jia, W.; Deng, A.M.; Liu, H.; Kang, J. MiR-138 promotes induced pluripotent stem cell generation through the regulation of the p53 signaling. Stem Cells 2012, 30, 1645–1654. [Google Scholar] [CrossRef] [PubMed]
- Escors, D.; Breckpot, K. Lentiviral vectors in gene therapy: Their current status and future potential. Arch. Immunol. Ther. Exp. 2010, 58, 107–119. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.K.; Teng, Y.; Wong, H.K.; Ng, T.K.; Huang, L.; Lei, P.; Choy, K.W.; Liu, Y.; Zhang, M.; Lam, D.S.; et al. MicroRNA-145 regulates human corneal epithelial differentiation. PLoS ONE 2011, 6, e21249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craig, V.; Tzankov, A.; Flori, M.; Schmid, C.; Bader, A.; Müller, A. Systemic microRNA-34a delivery induces apoptosis and abrogates growth of diffuse large B-cell lymphoma in vivo. Leukemia 2012, 26, 2421–2424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, A.F.; Weirauch, U.; Thomas, M.; Grünweller, A.; Hartmann, R.K.; Aigner, A. MicroRNA replacement therapy for miR-145 and miR-33a is efficacious in a model of colon carcinoma. Cancer Res. 2011, 71, 5214–5224. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.W.L.; Paoletti, C.; Campisi, M.; Osaki, T.; Adriani, G.; Kamm, R.D.; Mattu, C.; Chiono, V. MicroRNA delivery through nanoparticles. J. Control. Release 2019, 313, 80–95. [Google Scholar] [CrossRef] [PubMed]
- Kristen, A.V.; Ajroud-Driss, S.; Conceição, I.; Gorevic, P.; Kyriakides, T.; Obici, L. Patisiran, an RNAi therapeutic for the treatment of hereditary transthyretin-mediated amyloidosis. Neurodegener. Dis. Manag. 2019, 9, 5–23. [Google Scholar] [CrossRef] [Green Version]
- Hong, D.S.; Kang, Y.K.; Borad, M.; Sachdev, J.; Ejadi, S.; Lim, H.Y.; Brenner, A.J.; Park, K.; Lee, J.L.; Kim, T.Y.; et al. Phase 1 study of MRX34, a liposomal miR-34a mimic, in patients with advanced solid tumours. Br. J. Cancer 2020, 122, 1630–1637. [Google Scholar] [CrossRef]
- Levin, A.A. Treating disease at the RNA level with oligonucleotides. N. Engl. J. Med. 2019, 380, 57–70. [Google Scholar] [CrossRef]
- Voutila, J.; Reebye, V.; Roberts, T.C.; Protopapa, P.; Andrikakou, P.; Blakey, D.C.; Habib, R.; Huber, H.; Saetrom, P.; Rossi, J.J.; et al. Development and mechanism of small activating RNA targeting CEBPA, a novel therapeutic in clinical trials for liver cancer. Mol. Ther. 2017, 25, 2705–2714. [Google Scholar] [CrossRef] [Green Version]
- Witten, L.; Slack, F.J. miR-155 as a novel clinical target for hematological malignancies. Carcinogenesis 2020, 41, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Aster, J. Disease of white blood cells, lymph nodes, spleen, thymus. In Robins and Cotran Pathologic Basis of Disease, 7th ed.; Kumar, V., Abbas, A.K., Fausto, N., Eds.; Elsevier Saunders: Philadelphia, PA, USA, 2005; pp. 702–705. [Google Scholar]
- Baris, D.; Zahm, S.H. Epidemiology of lymphomas. Curr. Opin. Oncol. 2000, 12, 383–394. [Google Scholar] [CrossRef]
- Miao, Y.; Medeiros, L.J.; Li, J.; Young, K.H. Diffuse large B-cell lymphoma with molecular variations more than ABC and GCB classification. Gene Expr. 2018, 1400, 8. [Google Scholar] [CrossRef]
- Miao, Y.; Medeiros, L.J.; Li, Y.; Li, J.; Young, K.H. Genetic alterations and their clinical implications in DLBCL. Nat. Rev. Clin. Oncol. 2019, 16, 634–652. [Google Scholar] [CrossRef]
- Kokich, V.G. Surgical and orthodontic management of impacted maxillary canines. Am. J. Orthod. Dentofac. Orthop. 2004, 126, 278–283. [Google Scholar] [CrossRef]
- Pozzato, G.; Mazzaro, C.; Crovatto, M.; Modolo, M.L.; Ceselli, S.; Mazzi, G.; Sulfaro, S.; Franzin, F.; Tulissi, P.; Moretti, M. Low-grade malignant lymphoma, hepatitis C virus infection, and mixed cryoglobulinemia. Blood 1994, 84, 3047–3053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, D.Y.; Harris, N.L. Human Lymphoma: Clinical Implications of the REAL Classification; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Paya, C.V.; Fung, J.J.; Nalesnik, M.A.; Kieff, E.; Green, M.; Gores, G.; Habermann, T.M.; Wiesner, R.H.; Swinnen, L.J.; Woodle, E.S.; et al. Epstein-Barr virus-induced posttransplant lymphoproliferative disorders. Transplantation 1999, 68, 1517–1525. [Google Scholar] [CrossRef] [PubMed]
- Oksenhendler, E.; Boulanger, E.; Galicier, L.; Du, M.-Q.; Dupin, N.; Diss, T.C.; Hamoudi, R.; Daniel, M.T.; Agbalika, F.; Boshoff, C.; et al. High incidence of Kaposi sarcoma–associated herpesvirus–related non-Hodgkin lymphoma in patients with HIV infection and multicentric Castleman disease. Blood 2002, 99, 2331–2336. [Google Scholar] [CrossRef]
- Campo, E.; Swerdlow, S.H.; Harris, N.L.; Pileri, S.; Stein, H.; Jaffe, E.S. The 2008 WHO classification of lymphoid neoplasms and beyond: Evolving concepts and practical applications. Blood J. Am. Soc. Hematol. 2011, 117, 5019–5032. [Google Scholar] [CrossRef] [Green Version]
- Chapuy, B.; Stewart, C.; Dunford, A.J.; Kim, J.; Kamburov, A.; Redd, R.A.; Lawrence, M.S.; Roemer, M.G.; Li, A.J.; Ziepert, M.; et al. Molecular subtypes of diffuse large B cell lymphoma are associated with distinct pathogenic mechanisms and outcomes. Nat. Med. 2018, 24, 679–690. [Google Scholar] [CrossRef]
- Parsonnet, J.; Hansen, S.; Rodriguez, L.; Gelb, A.B.; Warnke, R.A.; Jellum, E.; Orentreich, N.; Vogelman, J.H.; Friedman, G.D. Helicobacter pylori infection and gastric lymphoma. N. Engl. J. Med. 1994, 330, 1267–1271. [Google Scholar] [CrossRef]
- Ok, C.Y.; Papathomas, T.G.; Medeiros, L.J.; Young, K.H. EBV-positive diffuse large B-cell lymphoma of the elderly. Blood J. Am. Soc. Hematol. 2013, 122, 328–340. [Google Scholar] [CrossRef] [Green Version]
- Shaffer, A.L.; Young, R.M.; Staudt, L.M. Pathogenesis of Human B Cell lymphomas. Annu. Rev. Immunol. 2012, 30, 565–610. [Google Scholar] [CrossRef] [PubMed]
- Morin, R.D.; Mendez-Lago, M.; Mungall, A.J.; Goya, R.; Mungall, K.L.; Corbett, R.D.; Johnson, N.A.; Severson, T.M.; Chiu, R.; Field, M.; et al. Frequent mutation of histone-modifying genes in non-Hodgkin lymphoma. Nature 2011, 476, 298–303. [Google Scholar] [CrossRef]
- Cerchietti, L.C.; Hatzi, K.; Caldas-Lopes, E.; Yang, S.N.; Figueroa, M.E.; Morin, R.D.; Hirst, M.; Mendez, L.; Shaknovich, R.; Cole, P.A.; et al. BCL6 repression of EP300 in human diffuse large B cell lymphoma cells provides a basis for rational combinatorial therapy. J. Clin. Investig. 2010, 120, 4569–4582. [Google Scholar] [CrossRef] [PubMed]
- Avantaggiati, M.L.; Ogryzko, V.; Gardner, K.; Giordano, A.; Levine, A.S.; Kelly, K. Recruitment of p300/CBP in p53-dependent signal pathways. Cell 1997, 89, 1175–1184. [Google Scholar] [CrossRef] [Green Version]
- Bereshchenko, O.R.; Gu, W.; Dalla-Favera, R. Acetylation inactivates the transcriptional repressor BCL6. Nat. Genet. 2002, 32, 606–613. [Google Scholar] [CrossRef] [PubMed]
- Ogryzko, V.V.; Schiltz, R.L.; Russanova, V.; Howard, B.H.; Nakatani, Y. The transcriptional coactivators p300 and CBP are histone acetyltransferases. Cell 1996, 87, 953–959. [Google Scholar] [CrossRef] [Green Version]
- Phan, R.T.; Dalla-Favera, R. The BCL6 proto-oncogene suppresses p53 expression in germinal-centre B cells. Nature 2004, 432, 635–639. [Google Scholar] [CrossRef]
- Pasqualucci, L.; Compagno, M.; Houldsworth, J.; Monti, S.; Grunn, A.; Nandula, S.V.; Aster, J.C.; Murty, V.V.; Shipp, M.A.; Dalla-Favera, R. Inactivation of the PRDM1/BLIMP1 gene in diffuse large B cell lymphoma. J. Exp. Med. 2006, 203, 311–317. [Google Scholar] [CrossRef] [Green Version]
- Lunning, M.A.; Green, M.R. Mutation of Chromatin Modifiers; An Emerging Hallmark of Germinal Center B-Cell Lymphomas. Blood Cancer J. 2015, 5, e361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasqualucci, L.; Dominguez-Sola, D.; Chiarenza, A.; Fabbri, G.; Grunn, A.; Trifonov, V.; Kasper, L.H.; Lerach, S.; Tang, H.; Ma, J.; et al. Inactivating mutations of acetyltransferase genes in B-cell lymphoma. Nature 2011, 471, 189–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shilatifard, A. The COMPASS Family of histone H3K4 methylases: Mechanisms of regulation in development and disease pathogenesis. Annu. Rev. Biochem. 2012, 81, 65–95. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Grubor, V.; Love, C.L.; Banerjee, A.; Richards, K.L.; Mieczkowski, P.A.; Dunphy, C.; Choi, W.; Au, W.Y.; Srivastava, G.; et al. Genetic heterogeneity of diffuse large B-cell lymphoma. Proc. Natl. Acad. Sci. USA 2013, 110, 1398–1403. [Google Scholar] [CrossRef] [Green Version]
- Pasqualucci, L.; Dalla-Favera, R. Genetics of diffuse large b-cell lymphoma. Blood 2018, 131, 2307–2319. [Google Scholar] [CrossRef]
- Sukswai, N.; Lyapichev, K.; Khoury, J.D.; Medeiros, L.J. Diffuse Large B-Cell Lymphoma Variants: An Update; Elsevier: Amsterdam, The Netherlands, 2020; pp. 53–67. [Google Scholar]
- Yap, D.B.; Chu, J.; Berg, T.; Schapira, M.; Cheng, S.W.G.; Moradian, A.; Morin, R.D.; Mungall, A.J.; Meissner, B.; Boyle, M.; et al. Somatic mutations at EZH2 Y641 act dominantly through a mechanism of selectively altered PRC2 catalytic activity, to increase H3K27 trimethylation. Blood 2011, 117, 2451–2459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knutson, S.K.; Wigle, T.J.; Warholic, N.M.; Sneeringer, C.J.; Allain, C.J.; Klaus, C.R.; Song, J.; Scott, M.P.; Jin, L.; Smith, J.J.; et al. A selective inhibitor of EZH2 blocks H3K27 methylation and kills mutant lymphoma cells. Nat. Chem. Biol. 2012, 8, 890–896. [Google Scholar] [CrossRef] [PubMed]
- Sneeringer, C.J.; Scott, M.P.; Kuntz, K.W.; Knutson, S.K.; Pollock, R.M.; Richon, V.M. Coordinated activities of wild-type plus mutant EZH2 drive tumor-associated hypertrimethylation of lysine 27 on histone H3 (H3K27) in human B-cell lymphomas. Proc. Natl. Acad. Sci. USA 2010, 107, 20980–20985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCabe, M.T.; Ott, H.M.; Ganji, G.; Korenchuk, S.; Thompson, C.; Aller, G.S.V.; Liu, Y.; Graves, A.P.; Diaz, E.; LaFrance, L.V.; et al. EZH2 inhibition as a therapeutic strategy for lymphoma with EZH2-activating mutations. Nature 2012, 492, 108–112. [Google Scholar] [CrossRef]
- Challa-Malladi, M.; Lieu, Y.K.; Califano, O.; Holmes, A.B.; Bhagat, G.; Murty, V.V.; Dominguez-Sola, D.; Pasqualucci, L.; Dalla-Favera, R. Combined genetic inactivation of β2-microglobulin and CD58 reveals frequent escape from immune recognition in diffuse large B cell lymphoma. Cancer Cell 2011, 20, 728–740. [Google Scholar] [CrossRef] [Green Version]
- Salles, G.; De Jong, D.; Xie, W.; Rosenwald, A.; Chhanabhai, M.; Gaulard, P.; Klapper, W.; Calaminici, M.; Sander, B.; Thorns, C.; et al. Prognostic significance of immunohistochemical biomarkers in diffuse large B-cell lymphoma: A study from the Lunenburg Lymphoma Biomarker Consortium. Blood 2011, 117, 7070–7078. [Google Scholar] [CrossRef]
- Steidl, C.; Shah, S.P.; Woolcock, B.W.; Rui, L.; Kawahara, M.; Farinha, P.; Johnson, N.A.; Zhao, Y.; Telenius, A.; Neriah, S.B.; et al. MHC class II transactivator CIITA is a recurrent gene fusion partner in lymphoid cancers. Nature 2011, 471, 377–383. [Google Scholar] [CrossRef] [Green Version]
- Moingeon, P.; Chang, H.C.; Wallner, B.P.; Stebbins, C.; Frey, A.Z.; Reinherz, E.L. CD2-mediated adhesion facilitates T lymphocyte antigen recognition function. Nature 1989, 339, 312–334. [Google Scholar] [CrossRef]
- Green, M.R.; Monti, S.; Rodig, S.J.; Juszczynski, P.; Currie, T.; O’Donnell, E.; Chapuy, B.; Takeyama, K.; Neuberg, D.; Golub, T.R.; et al. Integrative analysis reveals selective 9p24.1 amplification, increased PD-1 ligand expression, and further induction via JAK2 in nodular sclerosing Hodgkin lymphoma and primary mediastinal large B-cell lymphoma. Blood 2010, 116, 3268–3277. [Google Scholar] [CrossRef] [Green Version]
- Akyol, P.; Albayrak, M.; Maral, S.; Canpolat, F.; Afacan Öztürk, H.B.; Sağlam, B.; Tığlıoğlu, M. Diffuse large B cell lymphoma progression with skin involvement: A case report. J. Tissue Viability 2020, 29, 216–217. [Google Scholar] [CrossRef]
- Eric Davis, R.; Brown, K.D.; Siebenlist, U.; Staudt, L.M. Constitutive nuclear factor κB activity is required for survival of activated B cell-like diffuse large B cell lymphoma cells. J. Exp. Med. 2001, 194, 1861–1874. [Google Scholar] [CrossRef]
- Compagno, M.; Lim, W.K.; Grunn, A.; Nandula, S.V.; Brahmachary, M.; Shen, Q.; Bertoni, F.; Ponzoni, M.; Scandurra, M.; Califano, A.; et al. Mutations of multiple genes cause deregulation of NF-B in diffuse large B-cell lymphoma. Nature 2009, 459, 717–721. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Shaffer, A.L.; Emre, N.C.T.; Ceribelli, M.; Zhang, M.; Wright, G.; Xiao, W.; Powell, J.; Platig, J.; Kohlhammer, H.; et al. Exploiting synthetic lethality for the therapy of ABC diffuse large B cell lymphoma. Cancer Cell 2012, 21, 723–737. [Google Scholar] [CrossRef] [Green Version]
- Davis, R.E.; Ngo, V.N.; Lenz, G.; Tolar, P.; Young, R.M.; Romesser, P.B.; Kohlhammer, H.; Lamy, L.; Zhao, H.; Yang, Y.; et al. Chronic active B-cell-receptor signalling in diffuse large B-cell lymphoma. Nature 2010, 463, 88–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontan, L.; Yang, C.; Kabaleeswaran, V.; Volpon, L.; Osborne, M.J.; Beltran, E.; Garcia, M.; Cerchietti, L.; Shaknovich, R.; Yang, S.N.; et al. MALT1 small molecule inhibitors specifically suppress ABC-DLBCL in vitro and in vivo. Cancer Cell 2012, 22, 812–824. [Google Scholar] [CrossRef] [Green Version]
- Ngo, V.N.; Young, R.M.; Schmitz, R.; Jhavar, S.; Xiao, W.; Lim, K.H.; Kohlhammer, H.; Xu, W.; Yang, Y.; Zhao, H.; et al. Oncogenically active MYD88 mutations in human lymphoma. Nature 2011, 470, 115–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, M.; Sanada, M.; Kato, I.; Sato, Y.; Takita, J.; Takeuchi, K.; Niwa, A.; Chen, Y.; Nakazaki, K.; Nomoto, J.; et al. Frequent inactivation of A20 in B-cell lymphomas. Nature 2009, 459, 712–716. [Google Scholar] [CrossRef] [PubMed]
- Lenz, G.; Davis, R.E.; Ngo, V.N.; Lam, L.; George, T.C.; Wright, G.W.; Dave, S.S.; Zhao, H.; Xu, W.; Rosenwald, A.; et al. Oncogenic CARD11 mutations in human diffuse large B cell lymphoma. Science 2008, 319, 1676–1679. [Google Scholar] [CrossRef] [PubMed]
- Basso, K.; Dalla-Favera, R. Roles of BCL6 in normal and transformed germinal center B cells. Immunol. Rev. 2012, 247, 172–183. [Google Scholar] [CrossRef]
- Pasqualucci, L.; Migliazza, A.; Basso, K.; Houldsworth, J.; Chaganti, R.S.K.; Dalla-Favera, R. Mutations of the BCL6 proto-oncogene disrupt its negative autoregulation in diffuse large B-cell lymphoma. Blood 2003, 101, 2914–2923. [Google Scholar] [CrossRef]
- Migliazza, A.; Martinotti, S.; Chen, W.; Fusco, C.; Ye, B.; Knowles, D.M.; Offit, K.; Chaganti, R.S.; Dalla-Favera, R. Frequent somatic hypermutation of the 5′ non-coding region of the BCL6 gene in B-cell lymphoma. Proc. Natl. Acad. Sci. USA 1995, 92, 12520–12524. [Google Scholar] [CrossRef] [Green Version]
- Duan, S.; Cermak, L.; Pagan, J.K.; Rossi, M.; Martinengo, C.; Di Celle, P.F.; Chapuy, B.; Shipp, M.; Chiarle, R.; Pagano, M. FBXO11 targets BCL6 for degradation and is inactivated in diffuse large B-cell lymphomas. Nature 2012, 481, 90–94. [Google Scholar] [CrossRef] [Green Version]
- Ying, C.Y.; Dominguez-Sola, D.; Fabi, M.; Lorenz, I.C.; Hussein, S.; Bansal, M.; Califano, A.; Pasqualucci, L.; Basso, K.; Dalla-Favera, R. MEF2B mutations lead to deregulated expression of the oncogene BCL6 in diffuse large B cell lymphoma. Nat. Immunol. 2013, 14, 1084–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cattoretti, G.; Pasqualucci, L.; Ballon, G.; Tam, W.; Nandula, S.V.; Shen, Q.; Mo, T.; Murty, V.V.; Dalla-Favera, R. Deregulated BCL6 expression recapitulates the pathogenesis of human diffuse large B cell lymphomas in mice. Cancer Cell 2005, 7, 445–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shapiro-Shelef, M.; Lin, K.I.; McHeyzer-Williams, L.J.; Liao, J.; McHeyzer-Williams, M.G.; Calame, K. Blimp-1 is required for the formation of immunoglobulin secreting plasma cells and pre-plasma memory B cells. Immunity 2003, 19, 607–620. [Google Scholar] [CrossRef] [Green Version]
- Mandelbaum, J.; Bhagat, G.; Tang, H.; Mo, T.; Brahmachary, M.; Shen, Q.; Chadburn, A.; Rajewsky, K.; Tarakhovsky, A.; Pasqualucci, L.; et al. BLIMP1 is a tumor suppressor gene frequently disrupted in activated B cell-like diffuse large B cell lymphoma. Cancer Cell 2010, 18, 568–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solé, C.; Arnaiz, E.; Lawrie, C.H. MicroRNAs as Biomarkers of B-Cell Lymphoma. Biomark. Insights 2018, 13, 1177271918806840. [Google Scholar] [CrossRef] [Green Version]
- Lawrie, C.H.; Chi, J.; Taylor, S.; Tramonti, D.; Ballabio, E.; Palazzo, S.; Saunders, N.J.; Pezzella, F.; Boultwood, J.; Wainscoat, J.S.; et al. Expression of microRNAs in diffuse large B cell lymphoma is associated with immunophenotype, survival and transformation from follicular lymphoma. J. Cell. Mol. Med. 2009, 13, 1248–1260. [Google Scholar] [CrossRef]
- Solé, C.; Larrea, E.; Di Pinto, G.; Tellaexte, M.; Lawrie, C.H. miRNAs in B-Cell Lymphoma: Molecular Mechanisms and Biomarker Potential. Cancer Lett. 2017, 405, 79–89. [Google Scholar]
- Jablonska, E.; Gorniak, P.; Szydlowski, M.; Sewastianik, T.; Bialopiotrowicz, E.; Polak, A.; Warzocha, K.; Juszczynski, P. MiR-17-92 represses PTPROt and PP2A phosphatases and amplifies tonic BCR signaling in DLBCL cells. Exp. Hematol. 2017, 46, 56–61. [Google Scholar] [CrossRef]
- Wang, H.; Wang, A.; Hu, Z.; Xu, X.; Liu, Z.; Wang, Z. A critical role of miR-144 in diffuse large B-cell lymphoma proliferation and invasion. Cancer Immunol. Res. 2016, 4, 337–344. [Google Scholar] [CrossRef]
- Huang, F.; Jin, Y.; Wei, Y. Microrna-187 induces diffuse large B-cell lymphoma cell apoptosis via targeting BCL6. Oncol. Lett. 2016, 11, 2845–2850. [Google Scholar] [CrossRef] [Green Version]
- Fn, Q.; Meng, X.; Liang, H.; Zhang, H.; Liu, X.; Li, L.; Li, W.; Sun, W.; Zhang, H.; Zen, K.; et al. miR-10a inhibits cell proliferation and promotes cell apoptosis by targeting BCL6 in diffuse large B-cell lymphoma. Protein Cell. 2016, 7, 899–912. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Wang, H.; Ji, M. Overexpression of miR-222-3p promotes the proliferation and inhibits the apoptosis of diffuse large B-cell lymphoma cells via suppressing PPP2R2A. Technol. Cancer Res. Treat. 2019, 18, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Li, X.D.; Li, X.M.; Gu, J.W.; Sun, X.C. MiR-155 regulates lymphoma cell proliferation and apoptosis through targeting SOCS3/JAK-STAT3 signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 5153–5159. [Google Scholar] [PubMed]
- Costinean, S.; Zanesi, N.; Pekarsky, Y.; Tili, E.; Volinia, S.; Heerema, N.; Croce, C.M. Pre-B cell proliferation and lymphoblastic leukemia/high-grade lymphoma in Eμ-miR155 transgenic mice. Proc. Natl. Acad. Sci. USA 2006, 103, 7024–7029. [Google Scholar] [CrossRef] [Green Version]
- Kozloski, G.A.; Jiang, X.; Bhatt, S.; Ruiz, J.; Vega, F.; Shaknovich, R. MiR-181a negatively regulates NF-κB signaling and affects activated B-cell-like diffuse large B-cell lymphoma pathogenesis. Blood 2016, 127, 2856–2866. [Google Scholar] [CrossRef] [Green Version]
- He, M.; Gao, L.; Zhang, S.; Tao, L.; Wang, J.; Yang, J. Prognostic significance of miR-34a and its target proteins of FOXP1, p53, and BCL2 in gastric MALT lymphoma and DLBCL. Gastric Cancer 2014, 17, 431–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.R.; Zhang, X.; Zhang, Y. MiR-214 prevents the progression of diffuse large B-cell lymphoma by targeting PD-L1. Cell. Mol. Biol. Lett. 2019, 24, 68. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.M.; Lian, G.Y.; Song, Y.; Huang, Y.F.; Gong, Y. LncRNA MALAT1 promotes tumorigenesis and immune escape of diffuse large B cell lymphoma by sponging miR-195. Life Sci. 2019, 231, 116335. [Google Scholar] [CrossRef]
- Qian, C.S.; Li, L.J.; Huang, H.W.; Yang, H.F.; Wu, D.P. MYC-regulated lncRNA NEAT1 promotes B cell proliferation and lymphomagenesis via the miR-34b-5p-GLI1 pathway in diffuse large B-cell lymphoma. Cancer Cell Int. 2020, 20, 87. [Google Scholar] [CrossRef] [Green Version]
- Farina, F.M.; Inguscio, A.; Kunderfranco, P.; Cortesi, A.; Elia, L.; Quintavalle, M. MicroRNA-26a/cyclin-dependent kinase 5 axis controls proliferation, apoptosis and in vivo tumor growth of diffuse large B-cell lymphoma cell lines. Cell Death Dis. 2017, 8, e2890. [Google Scholar] [CrossRef]
- Su, H.; Chang, J.; Xu, M.; Sun, R.; Wang, J. CDK6 overexpression resulted from microRNA-320d downregulation promotes cell proliferation in diffuse large B-cell lymphoma. Oncol. Rep. 2019, 42, 321–327. [Google Scholar] [CrossRef]
- Xu, M.; Xu, T.A.O. Expression and clinical significance of miR-23a and MTSS1 in diffuse large B-cell lymphoma. Oncol. Lett. 2018, 16, 371–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Shen, J.; Su, N.; Wang, Q.; Zhang, M.; Liu, C. MiR-645 regulates the proliferation and apoptosis of diffuse large B-cell lymphoma by targeting DACH1. Human Cell 2020, 33, 1091–1098. [Google Scholar] [CrossRef]
- Huang, Y.; Zou, Y.; Lin, L.; Ma, X.; Zheng, R. MiR-101 regulates cell proliferation and apoptosis by targeting KDM1A in diffuse large B cell lymphoma. Cancer Manag. Res. 2019, 11, 2739–2746. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Wang, L.; Cheng, L. Aberrant ERG expression associates with downregulation of miR-4638-5p and selected genomic alterations in a subset of diffuse large B-cell lymphoma. Mol. Carcinog. 2019, 58, 1846–1854. [Google Scholar] [CrossRef] [PubMed]
- Mazan-Mamczarz, K.; Zhao, X.F.; Dai, B.; Steinhardt, J.J.; Peroutka, R.J.; Berk, K.L.; Landon, A.L.; Sadowska, M.; Zhang, Y.; Lehrmann, E.; et al. Down-Regulation of eIF4GII by miR-520c-3p Represses Diffuse Large B Cell Lymphoma Development. PLoS Genet. 2014, 10, e1004105. [Google Scholar] [CrossRef]
- Battistella, M.; Romero, M.; Castro-Vega, L.J.; Gapihan, G.; Bouhidel, F.; Bagot, M.; Feugeas, J.P.; Janin, A. The high expression of the microRNA 17-92 cluster and its paralogs, and the downregulation of the target gene PTEN, is associated with primary cutaneous B-cell lymphoma progression. J. Investig. Dermatol. 2015, 135, 1659–1667. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Z.; Li, X.; Zhu, Y.; Gu, W.; Xie, X.; Jisxang, J. Prognostic significance of MiRNA in patients with diffuse large B-cell lymphoma: A meta-analysis. Cell. Physiol. Biochem. 2016, 9, 1891–1904. [Google Scholar] [CrossRef] [Green Version]
- Takashima, Y.; Kawaguchi, A.; Iwadate, Y.; Hondoh, H.; Fukai, J.; Kajiwara, K.; Hayano, A.; Yamanaka, R. MiR-101, miR-548b, miR-554, and miR-1202 are reliable prognosis predictors of the miRNAs associated with cancer immunity in primary central nervous system lymphoma. PLoS ONE 2020, 15, e0229577. [Google Scholar] [CrossRef] [PubMed]
- Swerdlow, S.H.; Campo, E.; Pileri, S.A.; Lee Harris, N.; Stein, H.; Siebert, R.; Advani, R.; Ghielmini, M.; Salles, G.A.; Zelenetz, A.D.; et al. The 2016 Revision of the World Health Organization Classification of Lymphoid Neoplasms. Blood 2016, 127, 2375–2390. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, J.; Shen, Y.; Huang, X.; Liu, Y.; Wake, L.; Liu, C.; Deffenbacher, K.; Lachel, C.M.; Wang, C.; Rohr, J.; et al. Global microRNA expression profiling uncovers molecular markers for classification and prognosis in aggressive B-cell lymphoma. Blood 2015, 125, 1137–1145. [Google Scholar] [CrossRef]
- Lenze, D.; Leoncini, L.; Hummel, M.; Volinia, S.; Liu, C.G.; Amato, T.; De Falco, G.; Githanga, J.; Horn, H.; Nyagol, J.; et al. The different epidemiologic subtypes of Burkitt lymphoma share a homogenous micro RNA profile distinct from diffuse large B-cell lymphoma. Leukemia 2011, 25, 1869–1876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillar, N.; Bairey, O.; Goldschmidt, N.; Fellig, Y.; Rosenblat, Y.; Shehtman, I.; Haguel, D.; Raanani, P.; Shomron, N.; Siegal, T. MicroRNAs as predictors for CNS relapse of systemic diffuse large B-cell lymphoma. Oncotarget 2017, 8, 86020–86030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandhu, S.K.; Volinia, S.; Costinean, S.; Galasso, M.; Neinast, R.; Santhanam, R.; Parthun, M.R.; Perrotti, D.; Marcucci, G.; Garzon, R.; et al. MiR-155 targets histone deacetylase 4 (HDAC4) and impairs transcriptional activity of B-cell lymphoma 6 (BCL6) in the Eμ-miR-155 transgenic mouse model. Proc. Natl. Acad. Sci. USA 2012, 109, 20047–20052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Wei, B.; Hu, H.; Liu, F.; Tu, Y.; Zhao, M.; Wu, D. Preliminary study on decreasing the expression of FOXP3 with miR-155 to inhibit diffuse large B-cell lymphoma. Oncol. Lett. 2017, 14, 1711–1718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawrie, C.H.; Saunders, N.J.; Soneji, S.; Palazzo, S.; Dunlop, H.M.; Cooper, C.D.O.; Brown, P.J.; Troussard, X.; Mossafa, H.; Enver, T.; et al. MicroRNA Expression in Lymphocyte Development and Malignancy. Leukemia 2008, 22, 1440–1446. [Google Scholar] [CrossRef]
- Gu, L.; Song, G.; Chen, L.; Nie, Z.; He, B.; Pan, Y. Inhibition of miR-21 induces biological and behavioral alterations in diffuse large B-cell lymphoma. Acta Haematol. 2013, 130, 87–94. [Google Scholar] [CrossRef]
- Song, G.; Gu, L.; Li, J.; Tang, Z.; Liu, H.; Chen, B.; Sun, X.; He, B.; Pan, Y.; Wang, S.; et al. Serum microRNA expression profiling predict response to R-CHOP treatment in diffuse large B cell lymphoma patients. Ann. Hematol. 2014, 93, 1735–1743. [Google Scholar] [CrossRef]
- Yuan, W.X.; Gui, Y.X.; Na, W.N.; Chao, J.; Yang, X. Circulating microRNA-125b and microRNA-130a expression profiles predict chemoresistance to R-CHOP in diffuse large B-cell lymphoma patients. Oncol. Lett. 2016, 11, 423–432. [Google Scholar] [CrossRef] [Green Version]
- Troppan, K.; Wenzl, K.; Pichler, M.; Pursche, B.; Schwarzenbacher, D.; Feichtinger, J.; Thallinger, G.G.; Beham-Schmid, C.; Neumeister, P.; Deutsch, A. miR-199a and miR-497 are associated with better overall survival due to increased chemosensitivity in diffuse large b-cell lymphoma patients. Int. J. Mol. Sci. 2015, 16, 18077–18095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leivonen, S.-K.; Icay, K.; Jäntti, K.; Siren, I.; Liu, C.; Alkodsi, A.; Cervera, A.; Ludvigsen, M.; Hamilton-Dutoit, S.J.; d’Amore, F.; et al. MicroRNAs regulate key cell survival pathways and mediate chemosensitivity during progression of diffuse large B-cell lymphoma. Blood Cancer J. 2017, 7, 654. [Google Scholar] [CrossRef] [PubMed]
- Lawrie, C.H.; Gal, S.; Dunlop, H.M.; Pushkaran, B.; Liggins, A.P.; Pulford, K.; Banham, A.H.; Pezzella, F.; Boultwood, J.; Wainscoat, J.S.; et al. Detection of elevated levels of tumor-associated microRNAs in serum of patients with diffuse large B-cell lymphoma. Br. J. Haematol. 2008, 141, 672–675. [Google Scholar] [CrossRef] [PubMed]
- Khare, D.; Goldschmidt, N.; Bardugo, A.; Gur-Wahnon, D.; Ben-Dov, I.Z.; Avni, B. Plasma microRNA profiling: Exploring better biomarkers for lymphoma surveillance. PLoS ONE 2017, 12, e0187722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchesi, F.; Regazzo, G.; Palombi, F.; Terrenato, I.; Sacconi, A.; Spagnuolo, M.; Donzelli, S.; Marino, M.; Ercolani, C.; Di Benedetto, A.; et al. Serum miR-22 as potential non-invasive predictor of poor clinical outcome in newly diagnosed, uniformly treated patients with diffuse large B-cell lymphoma: An explorative pilot study. J. Exp. Clin. Cancer Res. 2018, 37, 95. [Google Scholar] [CrossRef] [Green Version]
- Meng, Y.; Quan, L.; Liu, A. Identification of key microRNAs associated with diffuse large B-cell lymphoma by analyzing serum microRNA expressions. Gene 2018, 642, 205–211. [Google Scholar] [CrossRef]
- Di Rocco, A.; De Angelis, F.; Ansuinelli, M.; Foà, R.; Martelli, M. Is now the time for molecular driven therapy for diffuse large B-cell lymphoma? Expert Rev. Hematol. 2017, 10, 761–774. [Google Scholar] [CrossRef] [Green Version]
- Chaganti, S.; Illidge, T.; Barrington, S.; Mckay, P.; Linton, K.; Cwynarski, K.; McMillan, A.; Davies, A.; Stern, S.; Peggs, K. Guidelines for the management of diffuse large B-cell lymphoma. Br. J. Haematol. 2016, 174, 43–56. [Google Scholar] [CrossRef] [Green Version]
- Martelli, M.; Ferreri, A.J.; Agostinelli, C.; Di Rocco, A.; Pfreundschuh, M.; Pileri, S.A. Diffuse large B-cell lymphoma. Crit. Rev. Oncol. Hematol. 2013, 87, 146–171. [Google Scholar] [CrossRef]
- Zhong, X.; Zhu, F.; Xu, X.; He, J.; Yu, X.; Chen, X.; Xu, K. The expression and clinical significance of microRNA-34a in diffuse lager B-cell lymphoma. Int. J. Clin. Exp. Med. 2016, 9, 10170–10180. [Google Scholar]
- Bai, H.; Wei, J.; Deng, C.; Yang, X.; Wang, C.; Xu, R. MicroRNA-21 regulates the sensitivity of diffuse large B-cell lymphoma cells to the CHOP chemotherapy regimen. Int. J. Hematol. 2013, 97, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Witten, L.W.; Cheng, C.J.; Slack, F.J. miR-155 drives oncogenesis by promoting and cooperating with mutations in the c-Kit oncogene. Oncogene 2019, 38, 2151–2161. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alsaadi, M.; Khan, M.Y.; Dalhat, M.H.; Bahashwan, S.; Khan, M.U.; Albar, A.; Almehdar, H.; Qadri, I. Dysregulation of miRNAs in DLBCL: Causative Factor for Pathogenesis, Diagnosis and Prognosis. Diagnostics 2021, 11, 1739. https://doi.org/10.3390/diagnostics11101739
Alsaadi M, Khan MY, Dalhat MH, Bahashwan S, Khan MU, Albar A, Almehdar H, Qadri I. Dysregulation of miRNAs in DLBCL: Causative Factor for Pathogenesis, Diagnosis and Prognosis. Diagnostics. 2021; 11(10):1739. https://doi.org/10.3390/diagnostics11101739
Chicago/Turabian StyleAlsaadi, Mohammed, Muhammad Yasir Khan, Mahmood Hassan Dalhat, Salem Bahashwan, Muhammad Uzair Khan, Abdulgader Albar, Hussein Almehdar, and Ishtiaq Qadri. 2021. "Dysregulation of miRNAs in DLBCL: Causative Factor for Pathogenesis, Diagnosis and Prognosis" Diagnostics 11, no. 10: 1739. https://doi.org/10.3390/diagnostics11101739