
Trechus (Coleoptera: Carabidae) of Appalachia: A Phylogenetic Insight into the History of High Elevation Leaf Litter Communities

Department of Plant & Environmental Science, Clemson University, Clemson, SC 29634, USA
*
Author to whom correspondence should be addressed.
Diversity 2024, 16(4), 212; https://doi.org/10.3390/d16040212
Submission received: 19 February 2024
/
Revised: 19 March 2024
/
Accepted: 21 March 2024
/
Published: 30 March 2024
(This article belongs to the Section Biodiversity Conservation)
Abstract
:Elevation gradients provide a wealth of habitats for a wide variety of organisms. The southern Appalachian Mountains in eastern United States are known for their high biodiversity and rates of endemism in arthropods, including in high-elevation leaf-litter taxa that are often found nowhere else on earth. Trechus Clairville (Coleoptera: Carabidae) is a genus of litter inhabitants with a near-global distribution and over 50 Appalachian species. These span two subgenera, Trechus s. str. and Microtrechus Jeannel, largely restricted to north and south of the Asheville basin, respectively. Understanding the diversification of these 3–5 mm flightless beetles through geological time can provide insights into how the litter-arthropod community has responded to historical environments, and how they may react to current and future climate change. We identified beetles morphologically and sequenced six genes to reconstruct a phylogeny of the Appalachian Trechus. We confirmed the Asheville Basin as a biogeographical barrier with a split between the north and south occurring towards the end of the Pliocene. Finer scale biogeography, including mountain-range occupancy, was not a reliable indication of relatedness, with group ranges overlapping and many instances of species-, species group-, and subgeneric sympatry. This may be because of the recent divergence between modern species and species groups. Extensive taxonomic revision of the group is required for Trechus to be useful as a bioindicator, but their high population density and speciose nature make them worth additional time and resources.
1. Introduction
The Appalachian Mountains of eastern North America are a region defined by old mountains and valleys. While lower in elevation than their western counterparts, the Rockies and Sierra Nevada, their age has allowed for organisms to establish and evolve in association with the abundant microhabitats in the region [1]. The Pleistocene glacial cycles were especially important to the development of the fauna of the present Appalachian Mountains [2]. While the Laurentide Ice Sheet covered Canada and the northern United States, the southern Appalachian region was affected by the climate cycles accompanying the glaciers further north [1]. Many biologists hypothesize that the glaciers and the cooler regions associated with them served as a connection for tundra-adapted species, allowing range expansion and gene flow. As glaciers melted and temperatures rose, these organisms were forced back north or into higher elevations, where climates were cooler [3,4].
Glaciation was not a static process; throughout the Pleistocene, areas of glacier expanded and contracted, allowing periodic migration between areas that were subsequently divided. When the glaciers finally receded at the end of the Ice Age, surviving organisms south of their typical ranges were confined to microhabitats where the remnants of their ideal habitat could be found, a movement pattern that can be modelled for both restricted and extended ranges [5,6] and often includes “sky islands”, high-elevation areas that isolate organisms from one another across relatively short distances [7,8]. Whereas the mechanisms behind their origins may vary, “sky island” systems and the communities they sustain found in various parts of the world are facing similar challenges, requiring phylogenetic and conservation work to preserve them [9,10,11].
In pursuit of cool and wet environments, many montane Appalachian species also took refuge in caves and along streams [12,13]. Many of these taxa are still dependent on these geographical features, as they are now too low in elevation to survive elsewhere. Many of these lack obvious morphological adaptations to the dark and wet, closely resembling leaf-litter relatives [12].
Organisms are also limited in their ability to relocate or expand their ranges by geographical barriers, such as valleys and rivers, that change in form and degree over time. Whereas rivers themselves may not be absolute barriers, the lower elevation regions of unsuitable habitats surrounding them can be [14]. The Asheville depression, the modern valley of the French Broad River, is the classic example in the Southern Appalachians, but the Little Tennessee River and a range of smaller water courses no longer in existence created finer-scale genetic isolation [15].
Many microhabitat-limited Appalachian species are now short-range endemics under threat of extirpation by human activity, invasive species, and climate change [2,16,17]. Leaf-litter communities at these sites support high biodiversity, with frequent and ongoing studies continually bettering our taxonomic knowledge of these regions [15,18,19]. More broadly, systematic and taxonomic work in the southern Appalachian region has centered around rare, isolated, and habitat-specific taxa in an attempt to preserve the areas and species while they are still able to survive [3,13,20,21]. Whereas this is greatly informative of policy and crucial to the survival of rare species, including the spruce-fir moss spider, Microhexura montivaga [22], there are regional questions that may be better addressed by the common, highly speciose members of the community that have high variability in their environmental and climatic needs. Comprehensive regional conservation plans that take these under-documented elements of biodiversity into account will thereby become increasingly feasible.
The genus Trechus Clairville, 1806 has 52 species documented in southern Appalachia (Table 1). Adults are wingless and 3–5 mm in length, limiting dispersal ability. Where found, they can be incredibly abundant (hundreds of individuals) in litter samples and underneath rocks, providing a wealth of material for study. Because of their size, Trechus and beetles like them have been neglected as prospective ecological indicators in the region, where use of carabids is otherwise commonplace [23].
Three Trechus subgenera occur in the southeastern United States, two native, one introduced. Microtrechus Jeannel is found exclusively in Appalachia, typically southwest of the French Broad River in North Carolina, Tennessee, and northern Georgia. Trechus s. str. is a Holarctic subgenus with only a few members in Appalachia (in the regionally endemic ‘T. hydropicus’ group), usually found northeast of the French Broad. Trechus (Calotrechus) obtusus Erichson, 1837 has been recently introduced into Appalachia and other areas and, unlike the endemics, is winged [24].
Trechine beetles have attracted considerable interest around the globe because of their adaptable nature and frequent association with cryptic microhabitats, including caves. Caves have served as lower-elevation refugia in a variety of trechines, some of which have evolved remarkable troglobitic traits including long legs, a loss of pigmentation, and reduced eye size [25,26,27]. Resolving the relationships of Appalachian Trechus, where such adaptations have not arisen, will help flesh out the picture of Trechus evolution globally.
In Appalachia, the genus Trechus has garnered particular interest because of their sheer diversity (over 50 species) and their fine-scale distributions across the central and southern Appalachians. In the early twentieth century, Jeannel described three species within this range: Trechus beutenmuelleri (1931) (which would become a subspecies of T. hydropicus), T. vandykei (1927), and T. barberi (1931) [28]. This group was then largely untouched until the 1960s, when Thomas Barr Jr. began publishing his adventures in the high peaks and caves of Appalachia. Barr established the species groups and their subgroups that evolved into the classification in use today (Table 1, Figure 1), based on geographic and morphological hypotheses [12,29,30,31,32]. His meticulous notebooks and specimens were left to the Carnegie Museum of Natural History, where they were made available to researchers. Over his career, Barr described 35 species of Appalachian Trechus and left many other potential new species from additional localities unpublished. Martin Donabauer, an amateur entomologist from Europe, would publish many of these localities and others in the early 2000s, describing 18 novel species and revising Microtrechus higher organization into what is currently accepted (Table 1) [33,34,35]. Donabauer is still contributing to the global trechine literature, with a focus on European representatives [36,37].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Current taxonomy of Appalachian Trechus [30,32,33,35]. Taxa sampled in this study are indicated (*).
| Subgenus | Group | Subgroup | Species | Subspecies |
|---|---|---|---|---|
| T. (Microtrechus) | T. vandykei | T. barberi | T. subtilis Barr, 1962 * | |
| T. pseudosubtilis Donabauer, 2009 * | ||||
| T. tonitru Barr, 1962 | ||||
| T. barberi Jeannel, 1931 * | ||||
| T. pseudobarberi Donabauer, 2009 * | ||||
| T. bowlingi | T. bowlingi Barr, 1962 * | |||
| T. vandykei | T. tusquitee Barr, 1979 | |||
| T. pisgahensis Barr, 1979 * | ||||
| T. vandykei Jeannel, 1927 * | ||||
| T. haoe Barr, 1979 * | ||||
| T. uncifer | T. inexpectatus Barr, 1985 * | |||
| T. thunderheadensis Donabauer, 2005 * | ||||
| T. uncifer Barr, 1962 | ||||
| T. plottbalsamensis Donabauer, 2005 * | ||||
| T. aduncus Barr, 1962 * | ||||
| T. cowensis Barr, 1962 | ||||
| T. toxawayi Barr, 1979 * | ||||
| T. howellae Barr, 1979 | ||||
| T. satanicus Barr, 1962 * | ||||
| T. tusquitensis Donabauer, 2005 * | ||||
| T. talequah Barr, 1962 * | ||||
| T. nebulosus | T. balsamensis Barr, 1962 * | |||
| T. cheoahensis Donabauer, 2005 | ||||
| T. clingmanensis Donabauer, 2005 * | ||||
| T. haoeleadensis Donabauer, 2005 | ||||
| T. luculentus | T. l. cheoahbaldensis Donabauer, 2005 | |||
| T. l. joannabaldensis Donabauer, 2005 * | ||||
| T. l. luculentus Barr, 1962 * | ||||
| T. l. wayahensis Barr, 1979 | ||||
| T. nantahalae Barr, 1979 | ||||
| T. nebulosus Barr, 1962 * | ||||
| T. novaculosus Barr, 1962 * | ||||
| T. pseudonovaculosus Donabauer, 2005 | ||||
| T. ramseyensis Donabauer, 2005 * | ||||
| T. rosenbergi Barr, 1962 | ||||
| T. snowbirdensis Donabauer, 2005 | ||||
| T. stefanschoedli Donabauer, 2005 | ||||
| T. stupkai Barr, 1979 | ||||
| T. tennesseensis | T. t. tauricus Barr, 1962 * | |||
| T. t. tennesseensis Barr, 1962 * | ||||
| T. thomasbarri Donabauer, 2005 | ||||
| T. tobiasi Donabauer, 2005 * | ||||
| T. tuckaleechee Barr, 1962 * | ||||
| T. unicoi Barr, 1962 | ||||
| T. valentinei Barr, 1979 | ||||
| T. verus Barr, 1962 * | ||||
| T. wayahbaldensis Donabauer, 2005 * | ||||
| T. (Trechus) | T. hydropicus | T. hydropicus | T. h. hydropicus (Horn, 1883) * | |
| T. h. avus Barr, 1962 * | ||||
| T. h. beutenmuelleri Jeannel, 1931 * | ||||
| T. h. canus Barr Barr, 1962 * | ||||
| T. schwarzi | T. s. schwarzi Jeannel, 1931 | |||
| T. s. scopulosus Barr, 1979 * | ||||
| T. s. saludae Barr, 1979 * | ||||
| T. cumberlandus Barr, 1962 * | ||||
| T. mitchellensis Barr, 1962 * | ||||
| T. carolinae Schaeffer, 1901 * | ||||
| T. roanicus Barr, 1962 * | ||||
| T. calignis Barr, 1985 * |
Previous studies of Trechus have mostly focused on morphological variation in establishing the species limits and infrageneric taxa of these beetles. Whereas species groups are identifiable under the microscope through male genitalia and other somatic traits, relationships between them remain unknown. Kane et al., [25], a study evaluating the relationships of members of the T. vandykei and T. bowlingi subgroups using gel electrophoresis, is the only modern genetic assessment for at least a part of Microtrechus. Trechus pisgahensis was found to be the sister to the remainder, with T. bowlingi recovered as sister to T. tusquitee, and T. vandykei as sister to T. haoe. It was concluded that gene flow was nonexistent between species subgroups, and that altitude was a major factor in this isolation [25]. In a later taxonomic revision, it was speculated that the “T. bowlingi” sampled therein were a mixture of multiple species, bringing these results into question [35]. Whereas the species subgroup taxonomy and preferred genetic analyses have changed over the last 30 years, their hypotheses regarding the movement of Trechus across Appalachia have provided a perspective inaccessible through morphological analysis.
In this study, we evaluated the large-scale evolutionary history of Appalachian Trechus beetles. We first ask how many independent lineages are represented among Appalachian Trechus, as their current classification into two subgenera, one shared with other Holarctic taxa, does not suggest monophyly. We reconstruct a dated, multigene phylogeny for the southern Appalachian Trechus to examine environmental and historical factors implicated in their isolation and related evolutionary patterns, including the Pleistocene glacial cycles, earlier climatic events, and physical barriers such as the Asheville depression. By sampling across the region, we explore genetic differences between peaks and mountain ranges to determine the timings of possible patterns of dispersal and occupancy. This intensive look at how a tiny, flightless leaf-litter resident has navigated geological history provides a different perspective of regional changes.
2. Materials and Methods
2.1. Specimen Collection and Identification
Southern Appalachia includes Virginia, West Virginia, Kentucky, Tennessee, North and South Carolina, Georgia, and Alabama. Trechus is not known to reside in Alabama.
Adults and larvae were collected into ethanol and live in leaf-litter samples throughout the known range of Appalachian Trechus. Sampling localities were selected based on the previous literature, type localities for species of interest, and a general rule that Trechus are typically found above 5000′ elevation (though as we have found, this is not absolute). Leaf litter was sifted using an 8 mm mesh litter sifter and placed in Berlese funnels for 8–12 h to collect beetles. Adults were also hand-collected by flipping rocks and logs, peering under moss, and peeling apart leaf packs within and along streams.
Adult males were identified morphologically by aedeagus morphology using the primary literature (Table 1, Figure 1) [12,29,30,31,32,33,34,35]. A subset (3–5) of those collected from each site, including males, females, and larvae, were selected for sequencing, with more included where morphological variability, such as in body size or in aedeagal morphology (detectable when slightly extruded from the apex of the abdomen, or through the abdominal sternites of less darkly pigmented specimens) were apparent.
Extralimital Trechus, necessary for assessing the monophyly of Appalachian taxa, were sampled from other published studies, totaling 233 specimens. Because the monophyly of global Trechus is questionable, individuals from several related genera were included as well. The final data set included 200 Appalachian Trechus individuals, 214 extralimital Trechus, and 46 non-Trechus outgroups.
2.2. DNA Extraction and Amplification
Six genes were chosen for this study based on their prevalence in the worldwide trechine literature [37,38,39]. All Appalachian specimens were sequenced for the COI barcoding or mini-barcoding region following lab protocol [24]. Wingless, 18S, 28S, CAD4, and Topoisomerase were sequenced for a subset of specimens, one individual per species or species group (Table 2). Outgroup sequences were obtained from Genbank (National Library of Medicine, Bethesda, MD, USA) and other Trechini works in progress [40].
Beetles were extracted using a combination of spin-column (GeneJET, Thermo-Scientific, Waltham, MA, USA) protocols and magnetic bead (Omega Bio-Tek Norcross, Norcross, GA, USA) purification. Non-mitochondrial loci were commercially Sanger sequenced by Psomagen (Rockville, MD, USA), while COI sequences were generated through both Sanger and next-generation techniques (Nanopore [46] and Illumina (https://illumina.com accessed on 10 January 2024) sequencing). Sequences were trimmed in GENEIOUS 8.1.8 (https://www.geneious.com accessed on 10 January 2024) before being aligned both with MUSCLE [47] and manually in Mesquite [48].
2.3. Tree Building
Before analysis, individuals identical in all sequences were merged in PAUP* for simplification. To infer the topology and monophyly of the taxa in question, a maximum likelihood analysis of all sequences available was performed with IQ-Tree on CIPRES (www.phylo.org accessed on 10 January 2024) with 50 bootstrap replicates and the TestMerge option, creating a consensus tree, returning a rooted phylogram with maximum-likelihood support values. Finding the monophyly of Appalachian Trechus to be strongly supported, the data set was then trimmed to Appalachian representatives and two outgroups (Pseudanophthtalmus Jeannel, 1920, and the extralimital T. obtusus), and the process was repeated. These outgroups were chosen over the more closely related Duvalius Delarouzée, 1859, because we were able to sequence all six genes from them. This trimmed phylogram was used as the model tree for BEAST2 analyses, constraining the topology to the above. Outgroups were limited in this analysis because of their variable genetic coverage. A dated phylogeny was created in BEAST2 with a standard carabid mutation rate for COI [49,50] of 0.0145 mutations per site per million years. A fossil parameter was included, creating a mean of 25 MYA for the separation of Trechus s.l. from other genera, with an offset of 15 and reverse log normal distribution. This was consistent with the dates obtained in the recent trechine literature [50,51]. The six genes were partitioned with unique rates and a single tree. The models were determined using IQ-Tree, which designated GTR for COI, TN94 for a concatenation of Topo and Wg, and Jukes–Cantor for the remainder of the genes (18S, 28S, and CAD4). Gamma site variability for all models was set to four categories. Bayesian analyses were completed on CIPRES in BEAST 2.6.6 with 100,000,000 mcmc generations and a tree-sampling frequency of 10,000. Trees were combined to obtain a consensus in TreeAnnotator, discarding 10% as burn-in.
A lineage-through-time plot was constructed to visualize the timing of diversification frequency and correlate these values with potential environmental cues. The plot was constrained to the interval between the Appalachian Trechus split and the timing of the shallowest branches with 90% ML support.
3. Results
The broadest maximum likelihood analysis provided 100% support for Appalachian Trechus monophyly, and for a sister group relationship between Microtrechus and the Appalachian members of Trechus s. str. (Supplementary Figure S1). Other genera were found to be interspersed with the Trechus outgroups but did not impact the taxa of interest’s reciprocal monophyly.
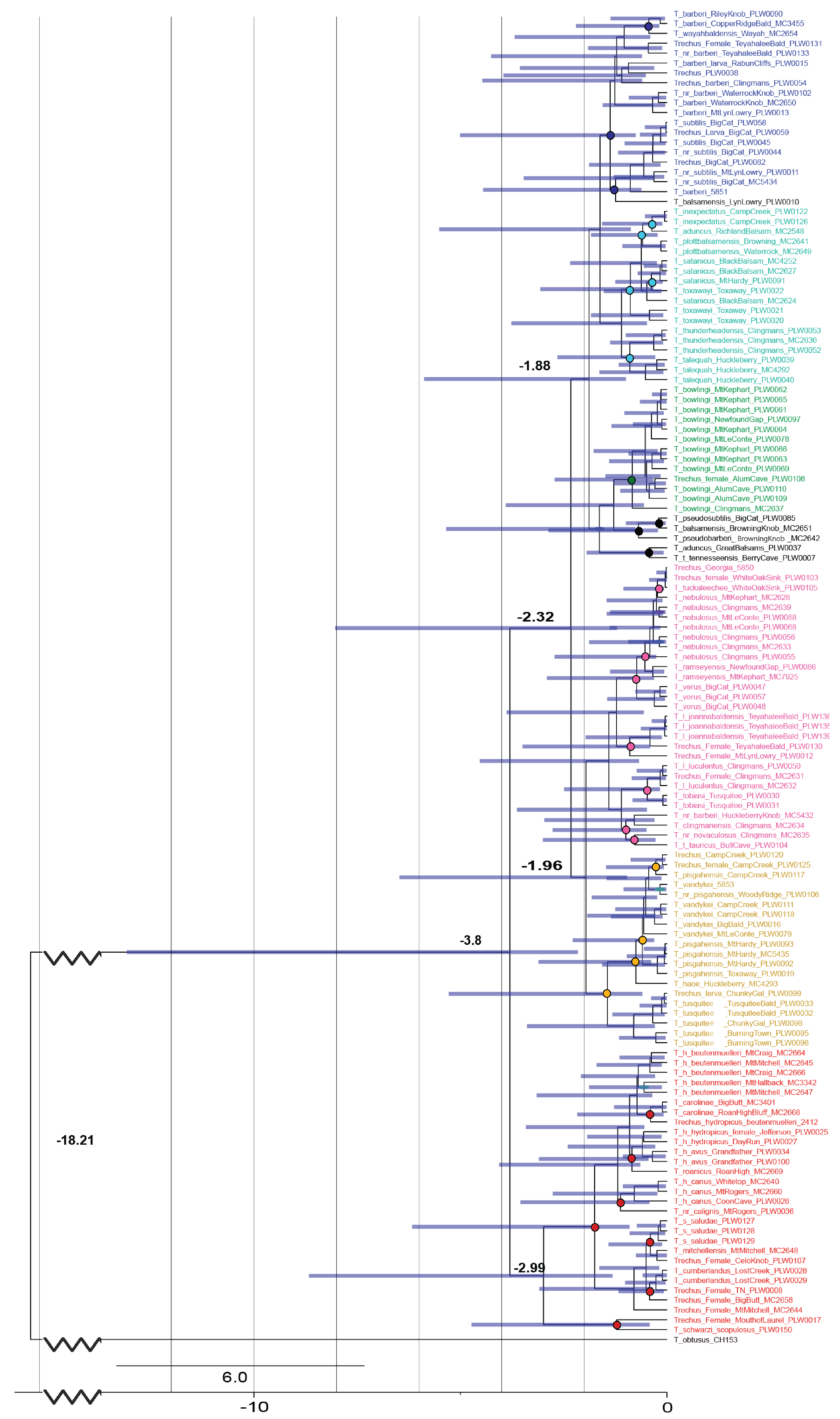
Through Bayesian clock analyses on the constrained ML tree (Figure 2) from the reduced data set, we found that the split between Microtrechus and Appalachian members of Trechus s.str. is estimated at 3.8 MYA with a wide margin of error (2 MYA to 15 MYA). Species subgroups diverged from one another between 1 and 3 MYA, with species-level differentiation falling largely within the last million years (Figure 3 and Figure 4). ESS values were high for all parameters except for COI tree likelihood (ESS = 126), which may demonstrate some incongruence between COI and the other genes.
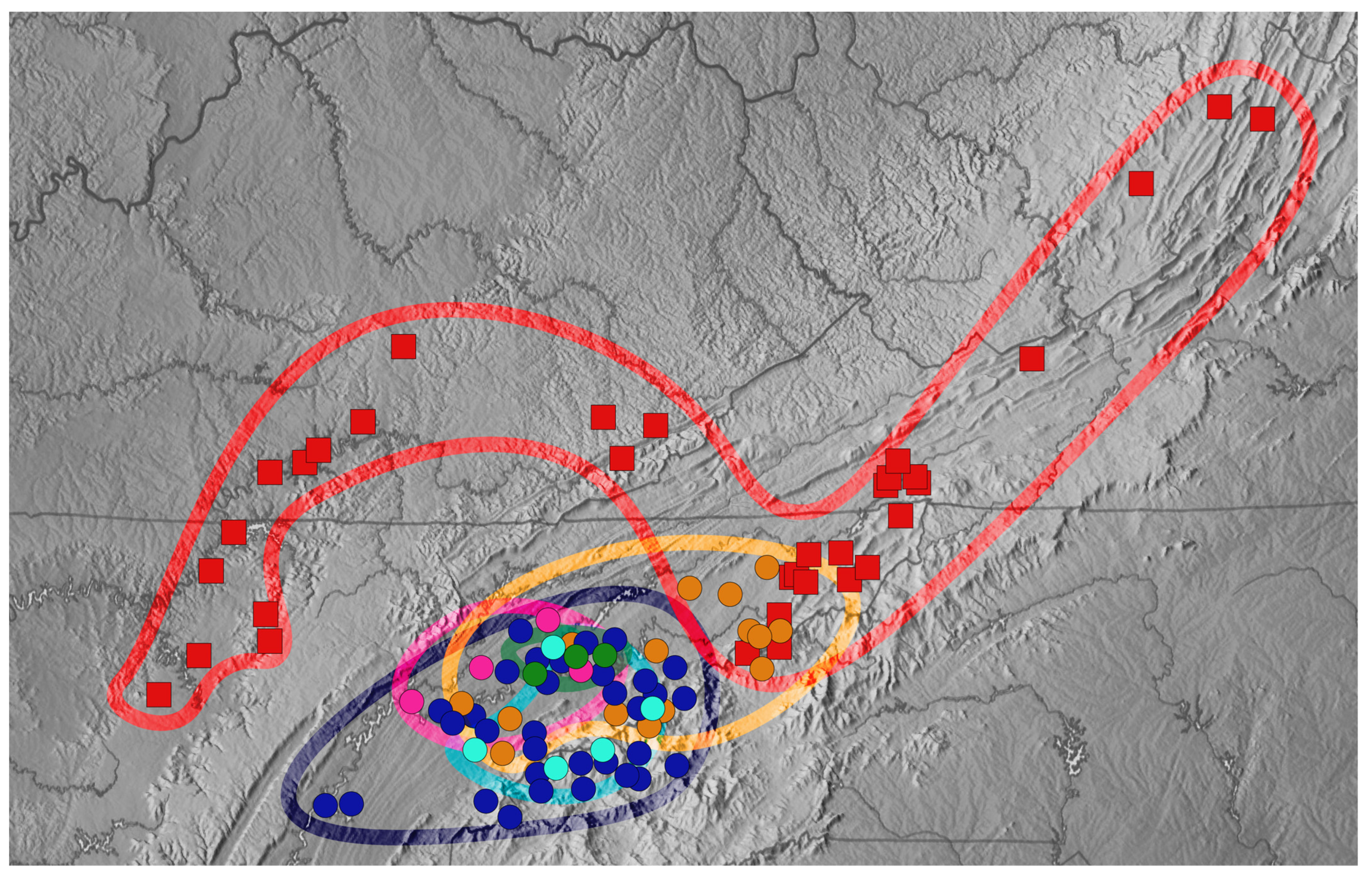
The range of Trechus s.str. in Appalachia (the T. hydropicus group) was confirmed to largely be confined to the north of the Asheville depression. Those not conforming to this pattern were found in the Cumberland Plateau, where they are known exclusively from caves and cave entrances. This group also contains a few lower-elevation representatives from wet riparian areas (subspecies of T. schwarzi Barr).
Microtrechus was subdivided into five well-supported lineages that exhibit considerable geographic overlap, though mostly west of the Asheville depression, and thus seem to represent old and simultaneously diversifying groups in the region (Figure 5). Members of the T. vandykei subgroup, however, extend across the Asheville depression, where they overlap with the T. (s.str.) hydropicus group. The T. bowlingi subgroup was confined to the Great Smoky Mountains and overlapped with all other Microtrechus groups and subgroups. The T. barberi subgroup extends the furthest south, into the mountains of north Georgia, but also overlaps with all other Microtrechus groupings. The T. uncifer and T. nebulosus groups share a minor overlap in the Great Smoky Mountains, which otherwise serve as a border between their ranges.
The Great Smoky Mountains was the range exhibiting the highest diversity and overlap in subgroups, with members of all five Microtrechus subgroups present at Clingmans Dome or a neighboring peak (e.g., Mount LeConte). The time of speciation amongst and within these subgroups is inconsistent, as is the diversity within them. Trechus subtilis exhibited several haplotypes, even though most specimens were recovered from only Big Cataloochee Mountain. Its widespread sister, T. barberi, had low genetic variability within sample sites, leading to one haplotype representing most localities.
At lower levels, there are a number of clear geographical patterns in speciation. From north (Camp Creek Bald and Big Bald) to south (Mount Hardy and Toxaway Mountain), there is a clear split in the T. vandykei subgroup, dated at ~0.75 MYA. From east (Balsam Mountains and Toxaway Mountain) to west (Clingmans Dome and Huckleberry Knob), the T. uncifer group appears to have split at least 0.25 MYA.
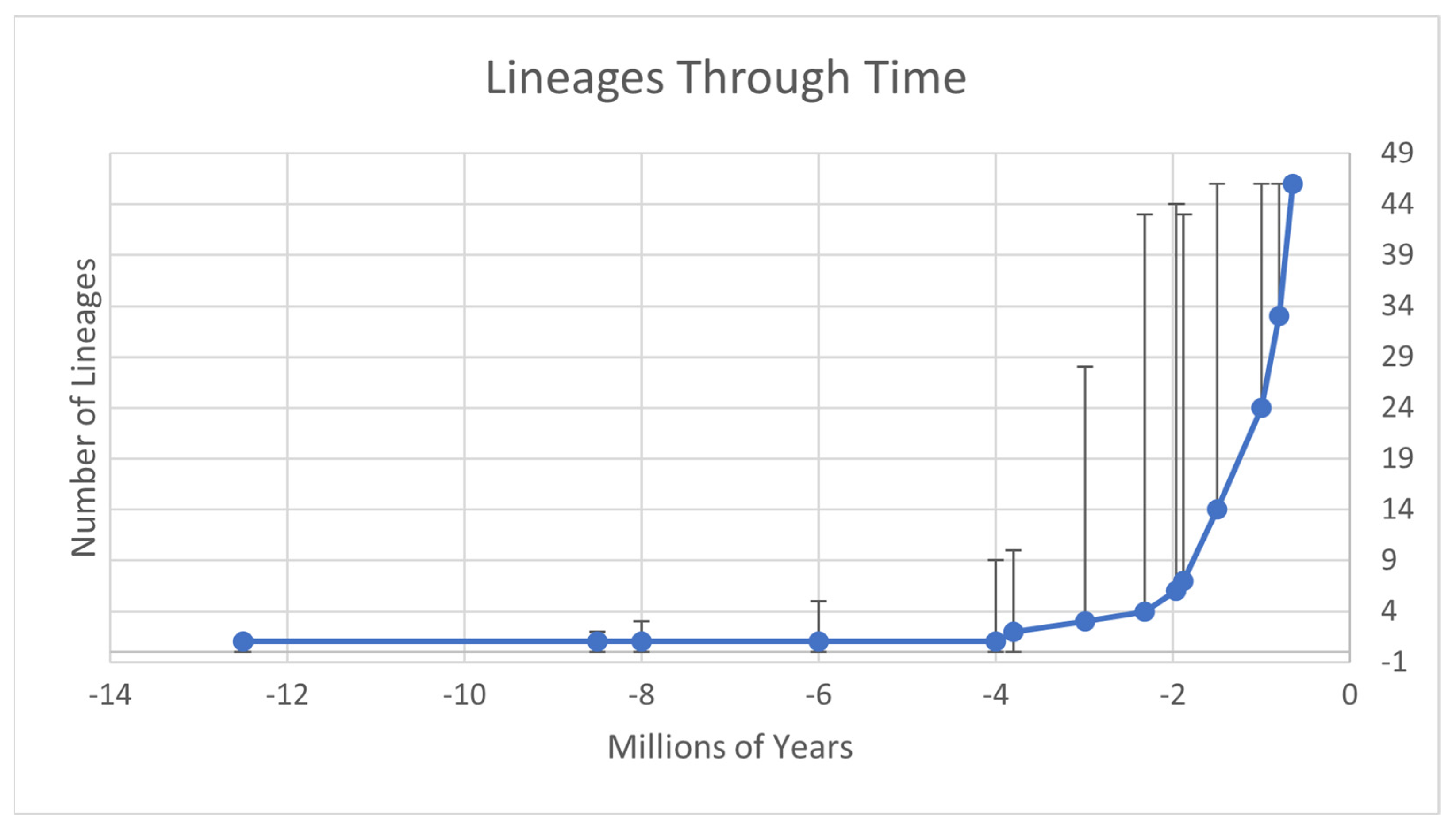
The lineage-through-time plot demonstrates that Appalachian Trechus, after splitting at the end of the Pliocene, had limited diversification for the next 2 MYA, followed by a period of increasingly rapid diversification, totaling 46 sampled lineages at the ~species-level cut-off of 0.65 MYA. Nearly half of the total current-lineage diversity apparently arose within the past million years.
4. Discussion
In this study, we used the morphological and genetic traits of Trechus across southern Appalachia to better our understanding of how the Pleistocene glacial cycles, older climatic events, and physical barriers have shaped the diversification and distribution of extant Trechus.
Subgenera of Trechus in southern Appalachia are reciprocally monophyletic and are the result of one larger radiation, rather than two smaller ones or a polyphyletic species assemblage. The presence of Microtrechus and their relationship with the T. hydropicus group reinforces that global Trechus s.str. is not monophyletic, with the Microtrechus and the T. hydropicus group being sisters to one another [36,52].
The split between these two groups occurred approximately 4 MYA, in the late Pliocene, with finer divisions occurring throughout the Pleistocene. Further differentiation into the species groups and subgroups of Microtrechus recognized today occurred approximately 2 MYA, with T. hydropicus differentiating considerably earlier (3 MYA). The type-species of this group is multiply paraphyletic and will require more extensive revision (namely the elevation of the subspecies) than any of the groups of Microtrechus (Wooden and Caterino in prep).
The division into major lineages within Microtrechus and the T. hydropicus group occurred largely within the early Pleistocene epoch. The dates of speciation within these lineages are highly variable, consistent with multiple periods of gene flow and isolation across the region (Figure 3). Within Microtrechus, the T. barberi subgroup was recovered as sister to the T. uncifer group. This confounds the hypothesis of T. vandykei group monophyly (Table 1), and is interesting morphologically, as there is little genitalic difference between T. barberi and T. bowlingi, and T. uncifer group members have morphologically complex male genitalia (Figure 1). The sister relationship between the T. vandykei subgroup and the T. nebulosus group carries similar implications. The T. uncifer and T. nebulosus groups also speciated later than lineages exhibiting more conserved morphology (Figure 3), which may indicate disruptive selection pressure in the later Pleistocene, possibly related to a reintroduction into sympatry.
The groups and subgroups exhibit considerable overlap, so their ancestors were evidently able to move across the landscape during this time, with many populations becoming endemic as the ranges expanded and retracted (Figure 4). This is atypical for southern Appalachian fauna that are often found in population clusters, e.g., one mountain range, habitat type, or area with little to no sympatry [20,21,22,53,54]. This uniqueness upholds our expectation that studying a mix of narrowly distributed taxa within a widespread genus with multiple habitat preferences yields a different picture than a focus on small-range endemics. Trechus, while useful for reconstructing recent biogeographic events, did not differentiate substantially before the late Pliocene and therefore cannot provide a picture of older climatic events. This late Pliocene split coincides with the development of the Asheville Basin around 4 MYA, a pervasive physical barrier.
The T. hydropicus group, to the north and west, exhibits a much larger geographic range than Microtrechus. This may be because southern ancestors, unable to travel north due to this basin, were more dependent on refugial microhabitats than their northern counterparts, and this is reflected in the results where morphology has been largely conserved, and individuals at the periphery of the range (such as T. cumberlandus in central Tennessee) have recently split from their relatives. Southern Microtrechus, as post-Pleistocene climates warmed, moved up the same mountain ranges as their ancestors, whereas T. hydropicus was able to expand to the west.
The speciation of Trechus was rapid and recent when compared to the ancient mountains in which they reside (Figure 3). Some other studies of Appalachian fauna have similarly found major splits in the late Pliocene and early Pleistocene, such as the eastern tiger salamander (Ambystoma tigrinum tigrinum), whose clades became reciprocally isolated in the early Pleistocene and have undergone extensive population differentiation in the last 500,000 years [5]. Unlike many other arthropods, Trechus species groups largely overlap geographically, which may be due to this late explosion of speciation and dynamic migration patterns, resulting in the sympatry seen today (at Clingmans Dome, for example). Species groups diverged approximately 2 MYA (1–6 MYA according to confidence interval), but most species divergences occurred later, in the late Pleistocene, allowing for distantly related species to re-establish on and inhabit the same mountaintop. Smaller historical riverine barriers that limited movement for older radiations [4] cannot reliably predict cladistic relationships in this case.
A recent species explosion is not unprecedented for Trechus, as species in the Canary Islands have undergone a similar expansion within the last 4 million years [55]. In Ptomaphagus (Coleoptera: Leiodidae), with numerous cave and soil endemic species in the southeast, Appalachian representatives have been found to have diversified in similar waves, with a split between the Cumberland Plateau lineages (TN, AL) and rest of their Appalachian range (GA, KY) [56]. Whereas the Asheville depression is not implicated in the isolation of these beetles, the changes in the Cumberland Plateau that are hypothesized to be behind their isolation would have been a factor for western lineages of Trechus as well. In Odontotaenius disjunctus (Coleoptera: Passalidae), Appalachia is home to widespread sympatric lineages thought to be divided by niche-partitioning, allowing for sympatry [57]. Although this is not a high-elevation endemic, it was affected by Pleistocene cycles and experienced divergences similar to those seen in this study. This passalid has defied numerous geographical barriers in southeastern United States, but its lineages still felt the effects of glacial cycles [57].
The Asheville Basin, which contains the French Broad River, has long been considered a biogeographic barrier for high-elevation taxa that may have prevented their retreat to the north as climates warmed. The generalization that species of Trechus s. str. are found north of this barrier and Microtrechus largely to the south (Figure 4) agrees with the existing literature [3,5,15,18,19]. Previous Trechus work cites the Asheville Basin as the prominent divider in Appalachian species [25]. However, there are exceptions in both directions. T. hydropicus group members are found in the Cumberland Plateau to the west, where extant individuals are largely troglobitic. Trechus vandykei is found on both sides of the basin, indicating recent leakage. This anomaly has been a common source of debate, and it is thought that when met with the divide, members of the T. hydropicus group moved westward and T. vandykei east [25,35].
Clingmans Dome, the highest peak in the Smokies, and neighboring Mount LeConte, are common hosts for endemic flora and fauna. Sedges [8], grasses [54], and other sedentary taxa have members only present there, and the sympatry found in Trechus agrees. The variability in the time of differentiation in Clingmans Dome residents indicates multiple dispersal events to the mountain within the last million years, which is consistent with the rapid diversification seen in the lineage-through-time plot. Kane et al. [25] speculated that the Smokies were special because of their patchiness and high abundance of high-elevation microhabitats. They hypothesized that this may have caused rapid niche divergence in the region as populations competed for resources and space. We see evidence in support of this hypothesis in the region’s sympatry, high diversity, and T. subtilis’ high genotype diversity on Big Cataloochee Mountain. Our study upholds that the Smokies are an important focus for Appalachian conservation, and that further niche modelling is required to understand what makes this range so special.
Three lineages of Appalachian Trechus are known exclusively from caves. Trechus cumberlandus (T. hydropicus group) diverged approximately 0.3 MYA. Trechus tennesseensis tauricus (T. nebulosus group) diverged around the same period, and T. t. tennesseensis (which does not appear to be closely related to its other subspecies) was more recent, around 0.2 MYA. This supports previous work suggesting that Trechus migrated into caves opportunistically, rather than coming to the surface from them. This movement into caves from montane habitats is found in other Trechines (namely Pseudanophthalmus) and reflects multiple waves into the Cumberland Plateau from the Black, Craggy, and Smoky mountains [29]. This recent deviation into caves is also supported by their lack of troglobitic morphology [32].
Two lineages are associated with riparian microclimates, found in lower elevations in leaf packs along and within riffles of perennial streams. T. schwarzi saludae (T. hydropicus group) and T. luculentus joannabaldensis (T. nebulosus group), while distantly related, both diverged around 0.25 MYA, implying that an environmental event, such as warming climatic cycles, may have sent them into these cooler, moist streams. Barr had also recognized that streams may be crucial to the lower-elevation success of these beetles, remarking that what he found below 4500 feet were proximal to mountain streams [29]. Trechus schwarzi saludae’s type and sampling locality are at a mere 1250 feet above sea level.
Appalachian Trechus contains multiple widespread species—namely T. barberi, T. vandykei, and T. hydropicus. There is little in their morphology to suggest why or for how long they have been more mobile than their relatives, and an ecological analysis is needed to understand the evolution of habitat preferences in the tolerant, riparian, and troglobitic members of the genus.
Southern Appalachian fauna have largely responded to the same geographical stressors, but the type of response and subsequent success of descendants is as varied as the region itself. Leaf-litter taxa are and continue to be a conduit for understanding the processes of the past and potential future. In pursuit of conserving the taxa known to science and describing those currently unknown to science, combining the perspectives of rare and common taxa in the context of phylogeography and niche partitioning may provide the most comprehensive picture for policy decisions in this microhabitat-rich landscape.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d16040212/s1, Figure S1: Placement of Appalachian Trechus in global trechine phylogeny; Table S1: Table of GenBank accession number and notes on methodology.
Author Contributions
Both authors conducted fieldwork, generated data, and completed identifications. P.L.S.W. confirmed identifications against type material. Both participated in the drafting of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by NSF DEB 1916263, the Clemson University Experiment Station (SC-1700596), and Clemson University’s E.W. King Award, Dissertation Completion Grant, and Wade Stackhouse Fellowship. This represents Technical Contribution # 7285 of the Clemson University Experiment Station.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
All data generated during this study are available on GenBank (sequence data; Accession #s pending) or the Clemson University Arthropod Collection’s search page on the SCAN (Symbiota Collections of Arthropods Network) database website (occurrence data).
Acknowledgments
We would like to acknowledge the Carnegie Museum of Natural History, Louisiana State Arthropod Museum, Smithsonian Museum of Natural History, and Harvard Museum of Comparative Zoology for access to specimens and loans. Adam Haberski, Curt Harden, Michael Ferro, Frank Etzler, and Derek Wooden were instrumental in the field portion of this project. Ernesto Recuero contributed much appreciated guidance on software use. David Maddison provided outgroup data access.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Yarnell, S.L. The Southern Appalachians: A History of the Landscape; US Department of Agriculture Forest Service: Asheville, NC, USA, 1998. [Google Scholar]
- Delcourt, P.A.; Delcourt, H.R. Paleoecological Insights on Conservation of Biodiversity: A Focus on Species, Ecosystems, and Landscapes. Ecol. Appl. 2017, 8, 921–934. [Google Scholar] [CrossRef]
- Hedin, M.; McCormack, M. Biogeographical evidence for common vicariance and rare dispersal in a southern Appalachian harvestman (Sabaconidae, Sabacon cavicolens). J. Biogeogr. 2017, 44, 1665–1678. [Google Scholar] [CrossRef]
- Thomas, S.M.; Hedin, M. Multigenic phylogeographic divergence in the paleoendemic southern Appalachian opilionid Fumontana deprehendor Shear (Opiliones, Laniatores, Triaenonychidae). Mol. Phylogen. Evol. 2008, 46, 645–658. [Google Scholar] [CrossRef]
- Church, S.A.; Kraus, J.M.; Mitchell, J.C.; Church, D.R.; Taylor, D.R. Evidence for multiple pleistocene refugia in the postglacial expansion of the eastern tiger salamander, Ambystoma tigrinum tigrinum. Evolution 2008, 57, 372–383. [Google Scholar] [CrossRef]
- Walker, M.J.; Stockman, A.K.; Marek, P.E.; Bond, J.E. Pleistocene glacial refugia across the appalachian mountains and coastal plain in the millipede genus Narceus: Evidence from population genetic, phylogeographic, and paleoclimatic data. BMC Evol. Biol. 2009, 9, 25. [Google Scholar] [CrossRef]
- Wheeler, Q. Notes on Host Associations and Habitats of Dasyceridae (Coleoptera) in the Southern Appalachian Mountains. Coleopt. Bull. 1984, 38, 227–231. [Google Scholar]
- Estes, D. Carex fumosimontana (Cyperaceae), a new endemic from the Great Smoky Mountains National Park, North Carolina and Tennessee. Brittonia 2013, 65, 200–207. [Google Scholar] [CrossRef]
- Dirnböck, T.; Essl, F.; Rabitsch, W. Disproportional risk for habitat loss of high-altitude endemic species under climate change. Glob. Chang. Biol. 2011, 17, 990–996. [Google Scholar] [CrossRef]
- Mccain, C.M.; Colwell, R.K. Assessing the threat to montane biodiversity from discordant shifts in temperature and precipitation in a changing climate. Ecol. Lett. 2011, 14, 1236–1245. [Google Scholar] [CrossRef]
- Wiens, J.J.; Camacho, A.; Goldberg, A.; Jezkova, T.; Kaplan, M.E.; Lambert, S.M.; Miller, E.C.; Streicher, J.W.; Walls, R.L. Climate change, extinction, and Sky Island biogeography in a montane lizard. Mol. Ecol. 2019, 28, 2610–2624. [Google Scholar] [CrossRef] [PubMed]
- Barr, T.C. The Pseudanophthalmus of the Appalachian Valley (Coleoptera: Carabidae). Am. Midl. Nat. 1965, 73, 41–72. [Google Scholar] [CrossRef]
- Hedin, M.; Milne, M.A. New species in old mountains: Integrative taxonomy reveals ten new species and extensive short-range endemism in Nesticus spiders (Araneae, Nesticidae) from the southern Appalachian Mountains. ZooKeys 2023, 1145, 1. [Google Scholar] [CrossRef]
- Graham, C.H.; Carnaval, A.C.; Cadena, C.D.; Zamudio, K.R.; Roberts, T.E.; Parra, J.L.; Mccain, C.M.; Bowie, R.C.K.; Moritz, C.; Baines, S.B.; et al. The origin and maintenance of montane diversity: Integrating evolutionary and ecological processes. Ecography 2014, 37, 711–719. [Google Scholar] [CrossRef]
- Caterino, M.S.; Recuero, E. Shedding light on dark taxa in sky-island Appalachian leaf litter: Assessing patterns of endemicity using large-scale, voucher-based barcoding. Insect Conserv. Divers. 2023, 17, 16–30. [Google Scholar] [CrossRef]
- Jenkins, M.A. Impact of the Balsam Woolly Adelgid (Adelges piceae Ratz.) on an Abies fraseri (Pursh) Poir. Dominated Stand near the Summit of Mount LeConte, Tennessee. Castanea 2003, 68, 109–118. [Google Scholar]
- McManamay, R.H.; Resler, L.M.; Campbell, J.B.; McManamay, R.A. Assessing the impacts of balsam woolly adelgid (Adelges piceae Ratz.) and anthropogenic disturbance on the stand structure and mortality of Fraser fir [Abies fraseri (pursh) Poir.] in the Black Mountains, North Carolina. Castanea 2011, 76, 1–19. [Google Scholar] [CrossRef]
- Dukes, C.D.; Janssens, F.; Recuero, E.; Caterino, M.S. Specific and Intraspecific Diversity of Symphypleona and Neelipleona (Hexapoda: Collembola) in Southern High Appalachia (USA). Diversity 2022, 14, 847. [Google Scholar] [CrossRef]
- Caterino, M.S.; Recuero, E. Molecular diversity of Protura in southern High Appalachian leaf litter. Biodivers. Data J. 2023, 11, e113342. [Google Scholar] [CrossRef]
- Graham, M.R.; Flint, W.D.; Powell, A.M.; Fet, V.; Pauley, T.K. Phylogeography of the Cow Knob Salamander (Plethodon punctatus Highton): Populations on isolated Appalachian mountaintops are disjunct but not divergent. Front. Amphib. Reptile Sci. 2023, 1, 1175492. [Google Scholar] [CrossRef]
- Herman, T.A.; Bouzat, J.L. Range-wide phylogeography of the four-toed salamander: Out of Appalachia and into the glacial aftermath. J. Biogeogr. 2016, 43, 666–678. [Google Scholar] [CrossRef]
- Hedin, M.; Carlson, D.; Coyle, F. Sky island diversification meets the multispecies coalescent—Divergence in the spruce-fir moss spider (Microhexura montivaga, Araneae, Mygalomorphae) on the highest peaks of southern Appalachia. Mol. Ecol. 2015, 24, 3467–3484. [Google Scholar] [CrossRef]
- Ortiz, C.C.; Browne, R.A. Carabidae (ground beetle) species composition of southern Appalachian spruce-fir forests. Southeast. Nat. 2011, 10, 591–608. [Google Scholar] [CrossRef]
- Harden, C.W.; Caterino, M.S.; Malabad, T.E. First Records of the Palearctic Species Trechus obtusus Erichson from the Appalachian Region of the Southeastern United States (Coleoptera: Carabidae: Trechinae: Trechini). Coleopt. Bull. 2022, 76, 61–69. [Google Scholar] [CrossRef]
- Kane, C.; Barr, T.C.; Stratton, E. Genetic patterns and population structure in Appalachian Trechus of the vandykei group. Brimleyana 1990, 16, 133–150. [Google Scholar]
- Faille, A.; Ribera, I.; Deharveng, L.; Bourdeau, C.; Garnery, L.; Queinnec, E.; Deuve, T. A molecular phylogeny shows the single origin of the Pyrenean subterranean Trechini ground beetles (Coleoptera: Carabidae). Mol. Phylogen. Evol. 2010, 54, 97–106. [Google Scholar] [CrossRef]
- Huang, S.; Tian, M.; Faille, A. A preliminary phylogeny of cave trechine beetles from China (Coleoptera: Carabidae: Trechini). In ARPHA Conference Abstracts; Pensoft Publishers: Sofia, Bulgaria, 2020; Volume 3. [Google Scholar] [CrossRef]
- Erwin, T.L.; House, G.N. A catalogue of the primary types of Carabidae (incl. Cicindelinae) in the collections of the United States National Museum of Natural History (USNM) (Coleoptera). Coleopt. Bull. 1978, 32, 231–256. [Google Scholar]
- Barr, T.C. The genus Trechus (Coleoptera: Carabidae: Trechini) in the southern Appalachians. Coleopt. Bull. 1962, 16, 65–92. [Google Scholar]
- Barr, T.C. Revision of Appalachian Trechus (Coleoptera: Carabidae). Brimleyana 1979, 2, 29–75. [Google Scholar]
- Barr, T.C. New Trechine Beetles (Coleoptera: Carabidae) from the Appalachian Region. Brimleyana 1985, 11, 119–132. [Google Scholar]
- Barr, T.C. Pattern and process in speciation of trechine beetles in eastern North America (Coleoptera: Carabidae: Trechinae). In Taxonomy, Phylogeny and Zoogeography of Beetles and Ants; Ball, G.E., Ed.; Dr W Junk Publishers: The Hague, The Netherlands, 1985. [Google Scholar]
- Donabauer, M. New Species and Subspecies of the Trechus (Microtrechus) nebulosus-group from the Southern Appalachians (Coleoptera: Carabidae: Trechinae). Z. Der Arbeitsgemeinschaft Der Osterr. Entomol. 2005, 57, 65–92. [Google Scholar]
- Donabauer, M. New species of the Trechus (Microtrechus) uncifer-group from the Southern Appalachians (Coleoptera: Carabidae). Z. Der Arbeitsgemeinschaft Der Osterr. Entomol. 2005, 57, 51–62. [Google Scholar]
- Donabauer, M. New species of the Trechus (Microtrechus) vandykei group from the southern Appalachians (Coleoptera: Carabidae: Trechinae). Z. Der Arbeitsgemeinschaft Der Osterr. Entomol. 2009, 61, 129–141. [Google Scholar]
- Donabauer, M. A taxonomic reorganization of European Trechus CLAIRVILLE, 1806 (Coleoptera: Carabidae: Trechinae). Z. Der Arbeitsgem. Osterr. Entomol. 2019, 71, 87–117. [Google Scholar]
- Möst, M.H.; Donabauer, M.; Arthofer, W.; Schlick-Steiner, B.C.; Steiner, F.M. Towards an evolutionary history of European-Alpine Trechus ground beetles: Species groups and wing reduction. Mol. Phylogen. Evol. 2020, 149, 106822. [Google Scholar] [CrossRef]
- Maddison, D.R.; Porch, N. A preliminary phylogeny and review of the genus Tasmanitachoides, with descriptions of two new species (Coleoptera, carabidae, bembidarenini). ZooKeys 2021, 1044, 153–196. [Google Scholar] [CrossRef]
- Maddison, D.R.; Kanda, K.; Boyd, O.F.; Faille, A.; Porch, N.; Erwin, T.L.; Rogi-Junent, S. Phylogeny of the beetle supertribe Trechitae (Coleoptera: Carabidae): Unexpected clades, isolated lineages, and morphological convergence. Mol. Phylogen. Evol. 2019, 132, 151–176. [Google Scholar] [CrossRef]
- Harden, C.W.; Davidson, R.L.; Malabad, T.E.; Caterino, M.S.; Maddison, D.R. Biology Phylogenetic systematics of the enigmatic genus Horologion Valentine, 1932 (Coleoptera, Carabidae, Trechinae, Horologionini), with description of a new species from Bath County, Virginia. Subterr. Biol. 2024, 49, 1–49. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome C oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Elbrecht, V.; Leese, F. Validation and development of freshwater invertebrate metabarcoding COI primers for environmental impact assessment. Front. Environ. Sci. 2017, 5, 1–11. [Google Scholar]
- Wild, A.L.; Maddison, D.R. Evaluating nuclear protein-coding genes for phylogenetic utility in beetles. Mol. Phylogen. Evol. 2008, 48, 877–891. [Google Scholar] [CrossRef]
- Shull, V.L.; Vogler, A.P.; Baker, M.D.; Maddison, D.R.; Hammond, P.M. Sequence alignment of 18S ribosomal RNA and the basal relationships of Adephagan beetles: Evidence for monophyly of aquatic families and the placement of Trachypachidae. Syst. Biol. 2011, 50, 945–969. [Google Scholar] [CrossRef]
- Maddison, D.R. Systematics of the North American beetle subgenus Pseudoperyphus (Coleoptera: Carabidae: Bembidion) based upon morphological, chromosomal, and molecular data. Ann. Carnegie Mus. 2008, 77, 147–193. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, Y.; Bollas, A.; Wang, Y.; Au, K.F. Nanopore sequencing technology, bioinformatics and applications. Nat. Biotechnol. 2021, 39, 1348–1365. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Molecular Analysis. 2001. Available online: https://www.mesquiteproject.org/home.html (accessed on 10 January 2024).
- Andújar, C.; Serrano, J.; Gámez-Zurita, J. Winding up the molecular clock in the genus Carabus (Coleoptera: Carabidae): Assessment of methodological decisions on rate and node age estimation. BMC Evol. Biol. 2012, 12, 40. [Google Scholar] [CrossRef]
- Faille, A.; Hofmann, S.; Merene, Y.; Hauth, D.; Opgenoorf, L.; Woldehawariat, Y.; Schmidt, J. Explosive radiation versus old relicts: The complex history of Ethiopian Trechina, with description of a new genus and a new subgenus. Dtsch. Entomol. Z. 2023, 70, 311–335. [Google Scholar] [CrossRef]
- Schmidt, J.; Belousov, I.; Michalik, P. X-ray microscopy reveals endophallic structures in a new species of the ground beetle genus Trechus Clairville, 1806 from Baltic amber (Coleoptera, Carabidae, Trechini). Zookeys 2016, 614, 113–127. [Google Scholar] [CrossRef]
- Faille, A.; Fresneda, J.; Bourdeau, C. Reconciling morphological and molecular data in a highly convergent group: The Pyrenean radiation of hypogean Trechini (Coleoptera: Carabidae). Integr. Syst. 2023, 6, 9–37. [Google Scholar] [CrossRef]
- Crespi, E.J.; Rissler, L.J.; Browne, R.A. Testing Pleistocene refugia theory: Phylogeographical analysis of Desmognathus wrighti, a high-elevation salamander in the southern Appalachians. Mol. Ecol. 2003, 12, 969–984. [Google Scholar] [CrossRef]
- Hermans, G.C.D.; Moya, O.; Oromi, P.; Juan, C. Evolution and diversification of the forest and hypogean ground-beetle genus Trechus in the Canary Islands. Mol. Phylogen. Evol. 2007, 42, 687–699. [Google Scholar]
- Leray, V.L.; Caravas, J.; Friedrich, M.; Zigler, K.S. Mitochondrial sequence data indicate “Vicariance by Erosion” as a mechanism of species diversification in North American Ptomaphagus (Coleoptera, Leiodidae, Cholevinae) cave beetles. Subterr. Biol. 2019, 29, 35–57. [Google Scholar] [CrossRef]
- Whitaker, M.; Procter, T.; Fontanella, F. Phylogeography and demographic expansion in the widely distributed horned passalus beetle, Odontotaenius disjunctus (Coleoptera: Passalidae). Mitochondrial DNA Part A 2021, 32, 85–97. [Google Scholar] [CrossRef]
- Anderson, W.A. A new species of Glyceria from the Great Smoky Mountains. Rhodora 1933, 35, 320–322. [Google Scholar]
Figure 1.
Appalachian Trechus (A). Adult habitus dorsal view (B–G). Aedeagi of group representatives (B). Trechus barberi (T. barberi subgroup) (C). T. pisgahensis (T. vandykei subgroup) (D). T. bowlingi (T. bowlingi subgroup) (E). T. nebulosus (T. nebulosus group) (F). T. hydropicus (T. hydropicus group) (G). T. thunderheadensis (T. uncifer group).
Figure 1.
Appalachian Trechus (A). Adult habitus dorsal view (B–G). Aedeagi of group representatives (B). Trechus barberi (T. barberi subgroup) (C). T. pisgahensis (T. vandykei subgroup) (D). T. bowlingi (T. bowlingi subgroup) (E). T. nebulosus (T. nebulosus group) (F). T. hydropicus (T. hydropicus group) (G). T. thunderheadensis (T. uncifer group).

Figure 2.
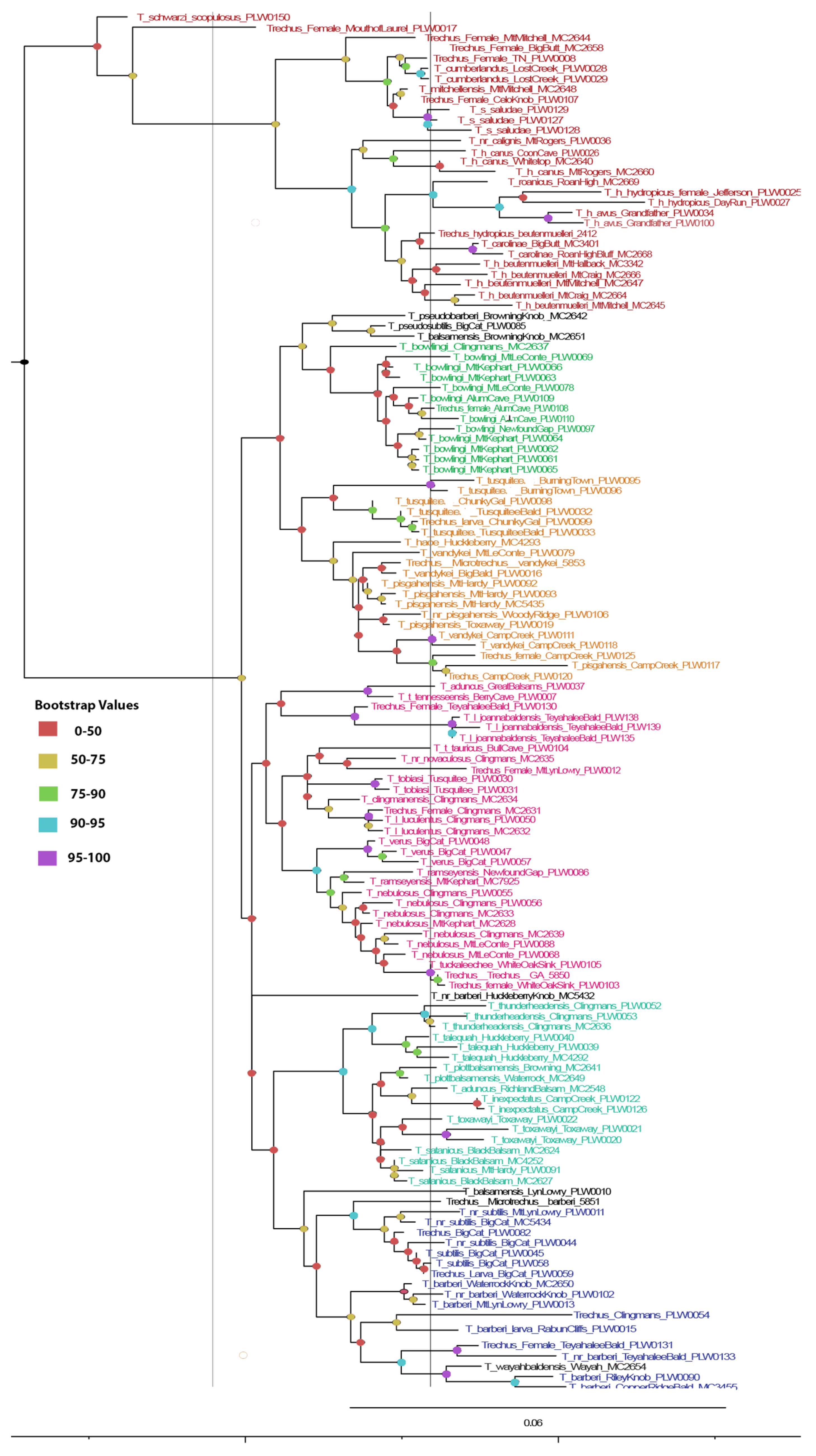
Maximum likelihood phylogram of Appalachian Trechus. Clades are coded to their species groups: red—T. hydropicus group, teal—T. uncifer group, pink—T. nebulosus group, orange—T. vandykei subgroup, green—T. bowlingi subgroup, and blue—T. barberi subgroup. Black-coded individuals fell outside their historical group. Bootstrap values are indicated at nodes. The tree is rooted at T. obtusus (not shown).
Figure 2.
Maximum likelihood phylogram of Appalachian Trechus. Clades are coded to their species groups: red—T. hydropicus group, teal—T. uncifer group, pink—T. nebulosus group, orange—T. vandykei subgroup, green—T. bowlingi subgroup, and blue—T. barberi subgroup. Black-coded individuals fell outside their historical group. Bootstrap values are indicated at nodes. The tree is rooted at T. obtusus (not shown).

Figure 3.
Bayesian-calibrated dated (MY) phylogram of Appalachian Trechus with a structure modelled from Figure 2. Circles indicate nodes of speciation colored by species group: red—T. hydropicus group, teal—T. uncifer group, pink—T. nebulosus group, orange—T. vandykei subgroup, green—T. bowlingi subgroup, and blue—T. barberi subgroup. Black-coded individuals fell outside their historical group.
Figure 3.
Bayesian-calibrated dated (MY) phylogram of Appalachian Trechus with a structure modelled from Figure 2. Circles indicate nodes of speciation colored by species group: red—T. hydropicus group, teal—T. uncifer group, pink—T. nebulosus group, orange—T. vandykei subgroup, green—T. bowlingi subgroup, and blue—T. barberi subgroup. Black-coded individuals fell outside their historical group.

Figure 4.
Lineage-through-time plot of Appalachian Trechus divergence. Error bars accommodate the uncertainty of dating reflected in the BEAST2 analysis.
Figure 4.
Lineage-through-time plot of Appalachian Trechus divergence. Error bars accommodate the uncertainty of dating reflected in the BEAST2 analysis.

Figure 5.
Map of southern Appalachia coded by species groups present. Dots indicate localities of previous collections. This includes those we collected, museum and databased specimens, and the previous literature on the group. Groups and subgroups are divided as denoted in Table 1. Squares are Trechus s.str. and circles Trechus (Microtrechus); the color code is as follows: red—T. hydropicus group, teal—T. uncifer group, pink—T. nebulosus group, orange—T. vandykei subgroup, green—T. bowlingi subgroup, and blue—T. barberi subgroup.
Figure 5.
Map of southern Appalachia coded by species groups present. Dots indicate localities of previous collections. This includes those we collected, museum and databased specimens, and the previous literature on the group. Groups and subgroups are divided as denoted in Table 1. Squares are Trechus s.str. and circles Trechus (Microtrechus); the color code is as follows: red—T. hydropicus group, teal—T. uncifer group, pink—T. nebulosus group, orange—T. vandykei subgroup, green—T. bowlingi subgroup, and blue—T. barberi subgroup.

Table 2.
Protocol for polymerase chain reactions (PCRs) conducted in-house. * Sequences for COI did not always render useable data, but the PCR was attempted for each individual.
Table 2.
Protocol for polymerase chain reactions (PCRs) conducted in-house. * Sequences for COI did not always render useable data, but the PCR was attempted for each individual.
| Gene | Forward Primer | Reverse Primer | Reference | Denaturing | Annealing | Cycle # | Sample Frequency |
|---|---|---|---|---|---|---|---|
| COI | LCO1490 | HCO2198 | [41] | 94 °C 0:30 | 50 °C 0:30 | 35 | Every Individual * |
| BF2 | BR2 | [42] | |||||
| Topo | TP643F | TP932R | [43] | 94 °C 0:30 | 57°/52°/45° 0:30 | 6/6/36 | Species Group |
| TP675F | TP932R | [43] | |||||
| CAD4 | CD806F | CD1098R2 | [43] | 94 °C 0:20 | 60 °C 0:20 | 39 | Species Group |
| CD821F | CD1098R2 | [43] | 94 °C 0:20 | 55 °C 0:20 | 37 | ||
| 18S | 18S5′ | 18Sb5.0 | [44] | 94 °C 0:30 | 50 °C 0:30 | 35 | Species Group |
| 28S | NLF184/21 | LS1041R | [45] | 94 °C 0:30 | 50 °C 0:30 | 35 | Species Group |
| Wingless | wg550F | wgAbRZ | [43] | 94 °C 0:20 | 52 °C 0:20 | 37 | Species |
| wg578F | wgAbR | [43] | 94 °C 0:20 | 54 °C 0:20 | 35 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wooden, P.L.S.; Caterino, M.S. Trechus (Coleoptera: Carabidae) of Appalachia: A Phylogenetic Insight into the History of High Elevation Leaf Litter Communities. Diversity 2024, 16, 212. https://doi.org/10.3390/d16040212
AMA Style
Wooden PLS, Caterino MS. Trechus (Coleoptera: Carabidae) of Appalachia: A Phylogenetic Insight into the History of High Elevation Leaf Litter Communities. Diversity. 2024; 16(4):212. https://doi.org/10.3390/d16040212
Chicago/Turabian StyleWooden, Patricia L. S., and Michael S. Caterino. 2024. "Trechus (Coleoptera: Carabidae) of Appalachia: A Phylogenetic Insight into the History of High Elevation Leaf Litter Communities" Diversity 16, no. 4: 212. https://doi.org/10.3390/d16040212
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.