
Surveying Bat-Hosted Adenoviruses and Herpesviruses: A Comprehensive Analysis

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
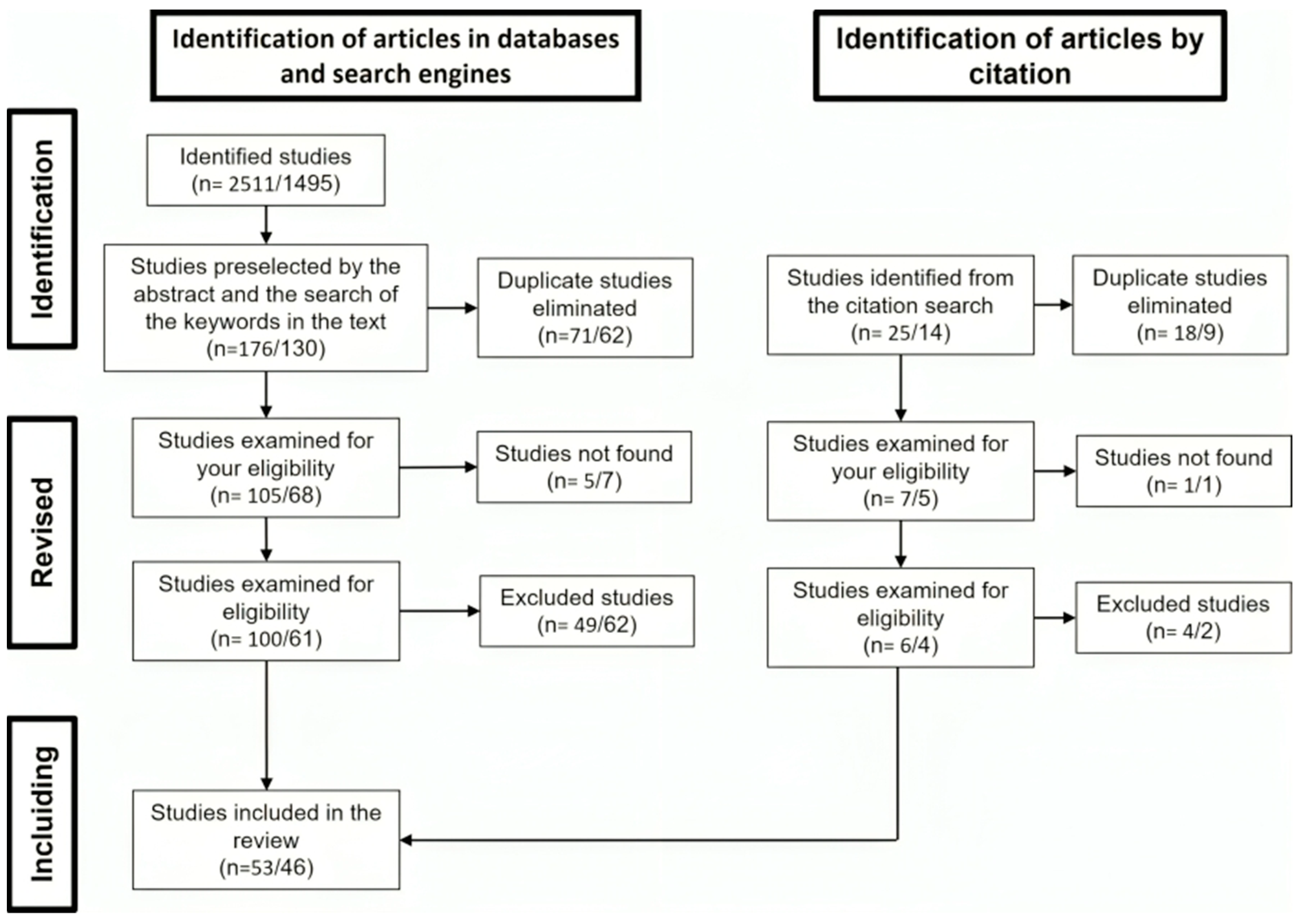
2.1. Literature Review and Bibliometric Analyses
2.2. Salience Analysis
3. Results
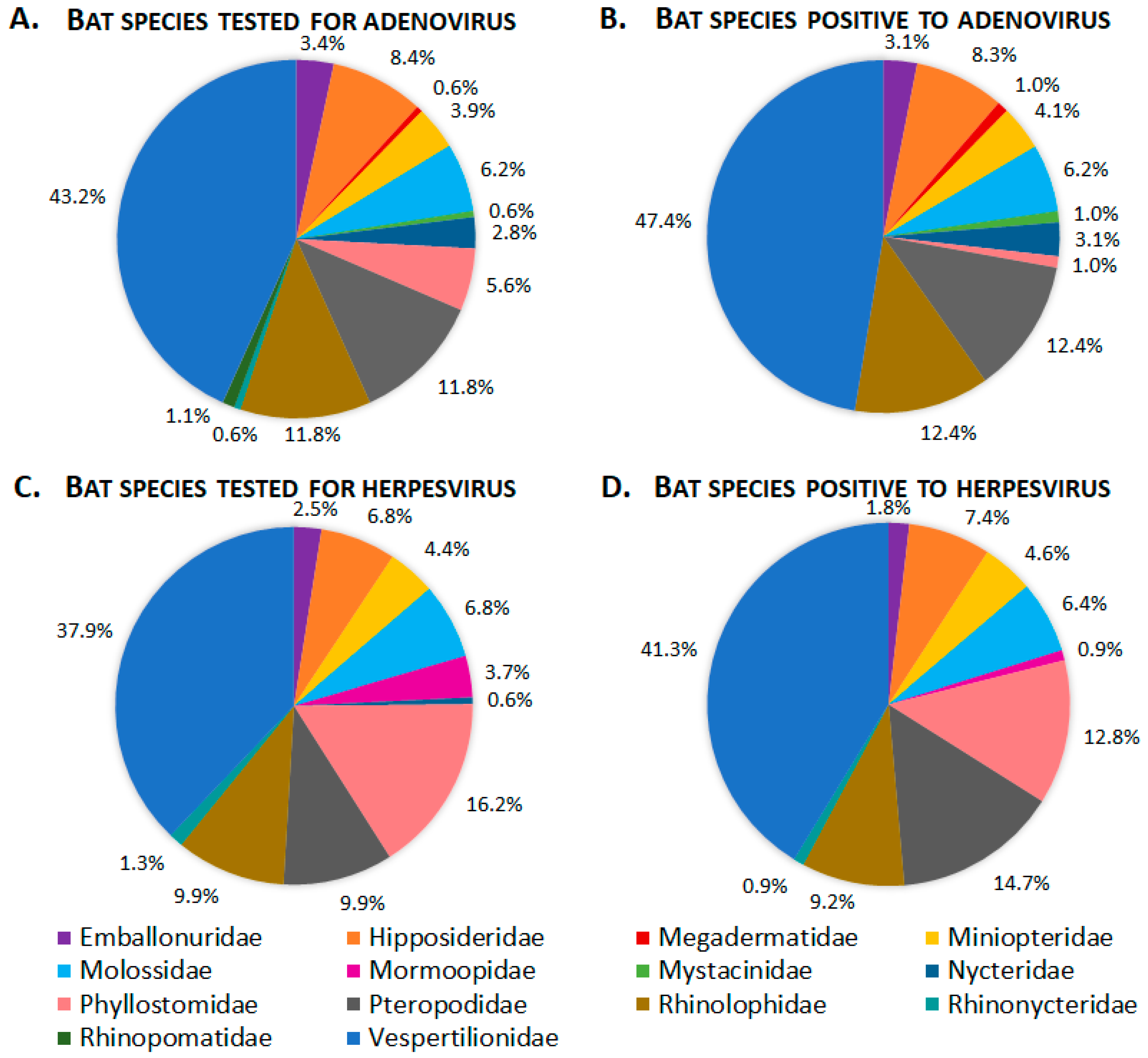
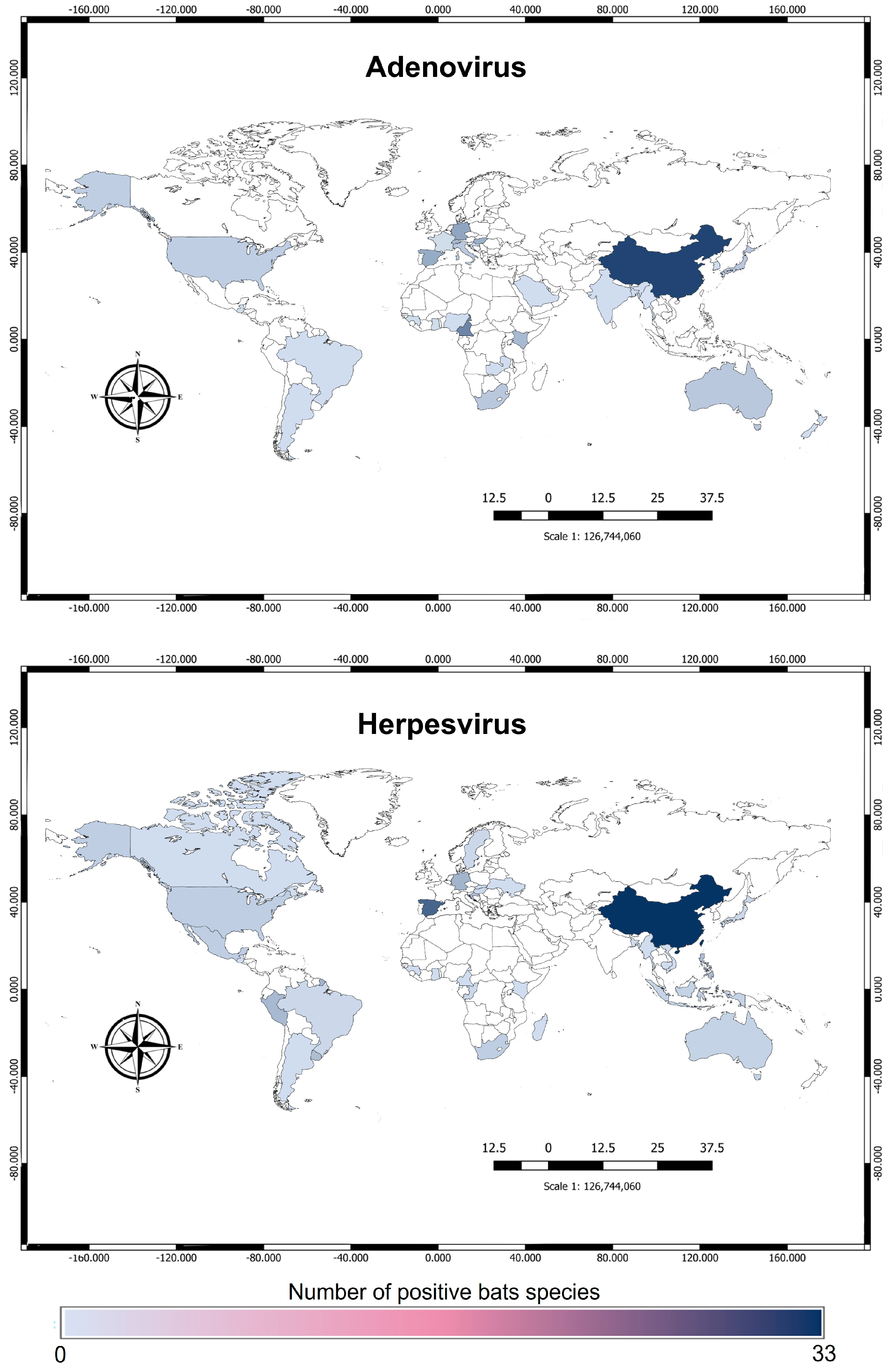
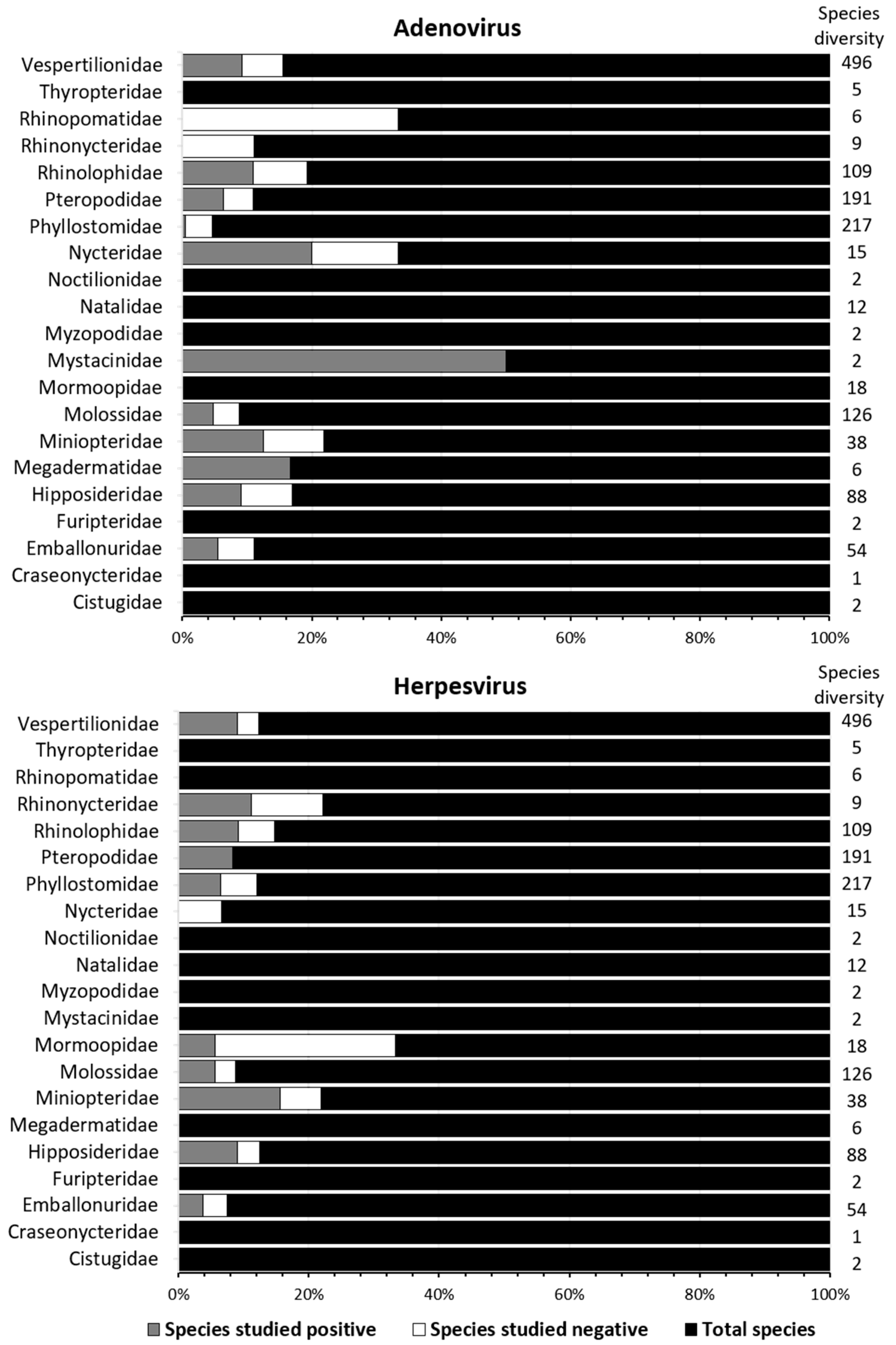
3.1. Adenoviridae
3.2. Herpesvirus
3.3. Published Research of Adenoviridae and Herpesviridae in Bats
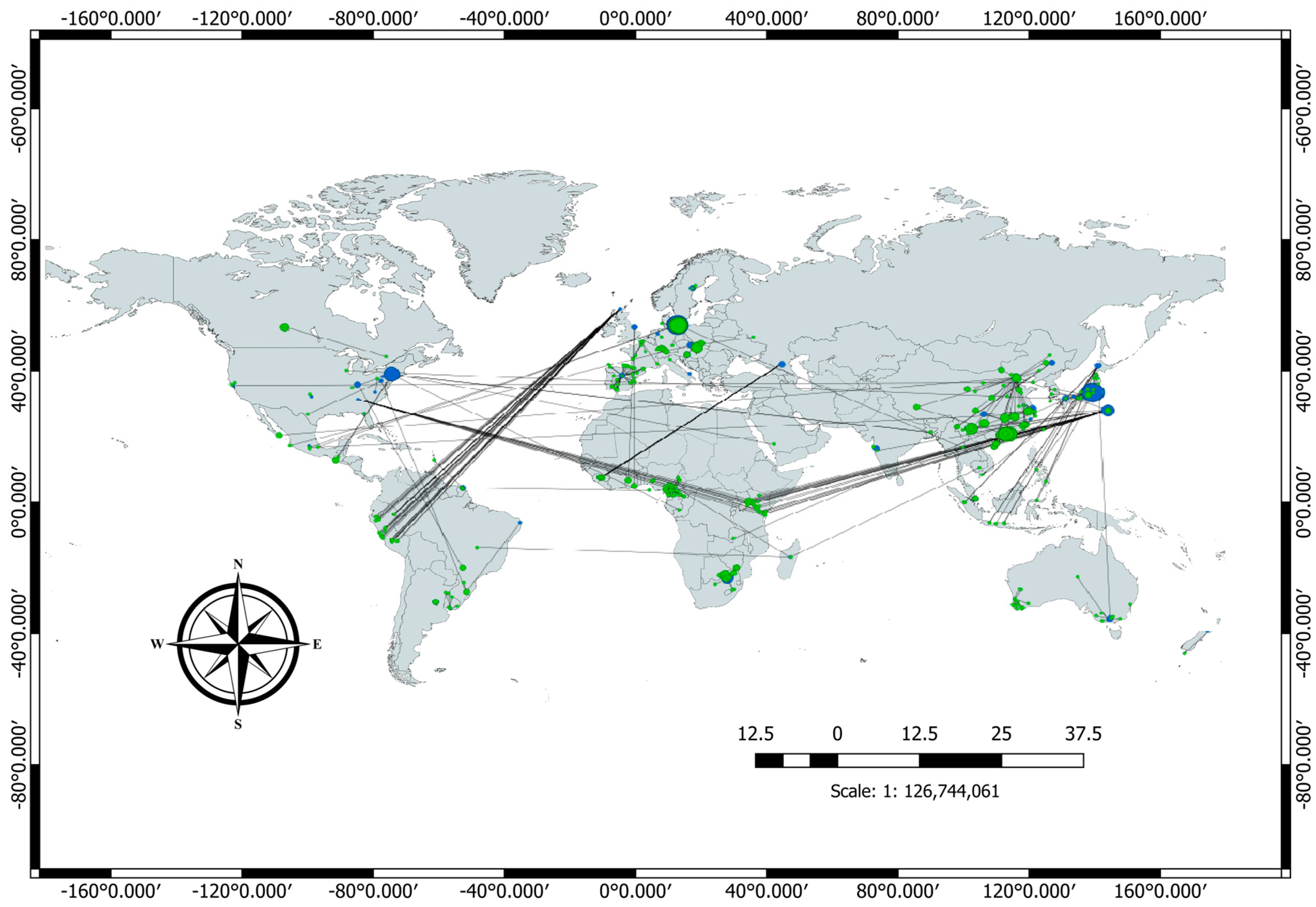
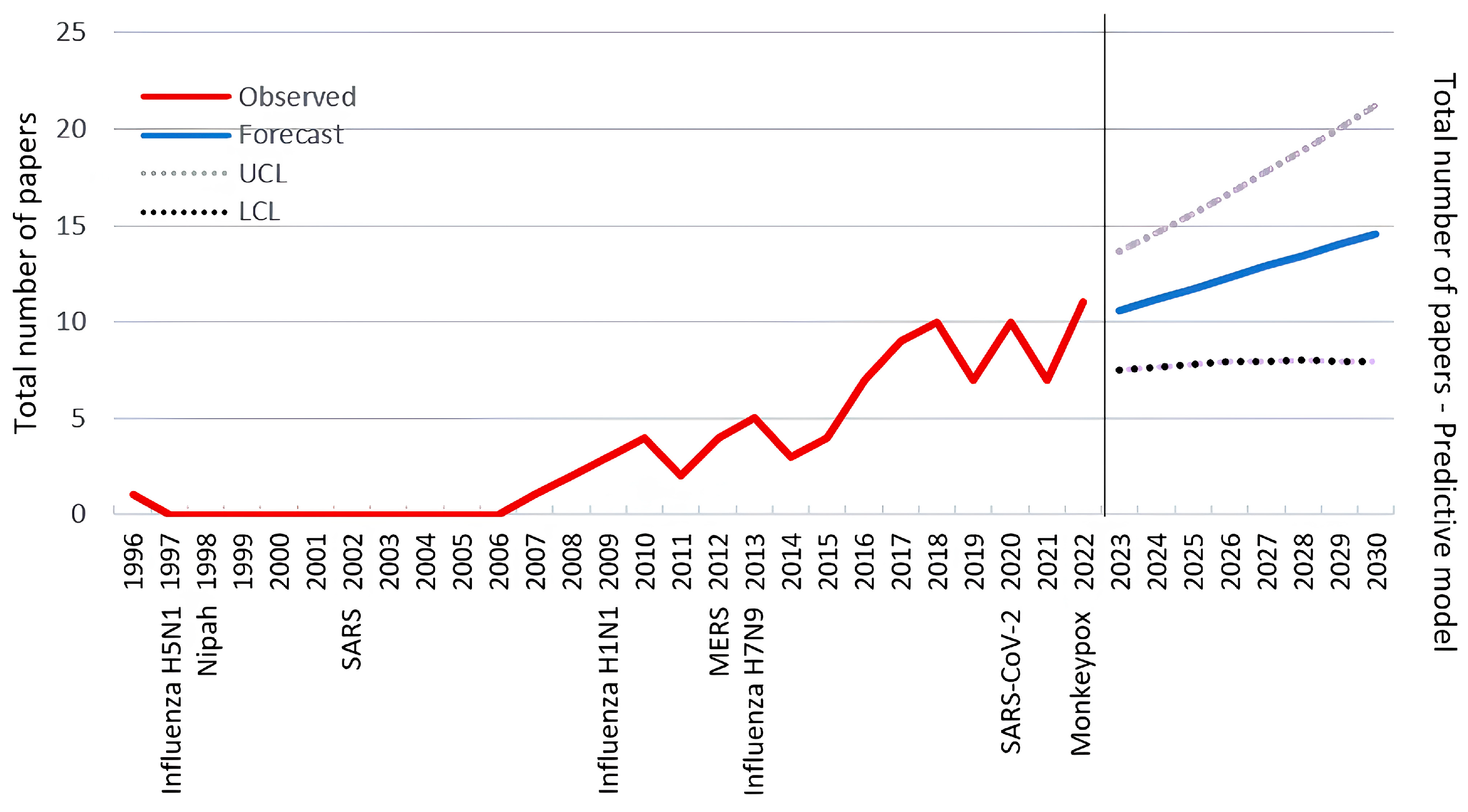
3.4. Bibliometric Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oliva, K.J.; Epstein, J.H.; Wang, L.-F.; Field, H.E.; Daszak, P. Are Bats Exceptional Viral Reservoirs? In New Directions in Conservaion Medicine: Applied Cases of Ecological Health; Aguirre, A.A., Ostfeld, R., Daszak, P., Eds.; Oxford Publishing: Oxford, UK, 2012; pp. 195–212. [Google Scholar]
- O’Shea, T.J.; Cryan, P.M.; Cunningham, A.A.; Fooks, A.R.; Hayman, D.T.S.; Luis, A.D.; Peel, A.J.; Plowright, R.K.; Wood, J.L.N. Bat Flight and Zoonotic Viruses. Emerg. Infect. Dis. 2014, 20, 741–745. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Baker, M.L.; Kulcsar, K.; Misra, V.; Plowright, R.; Mossman, K. Novel Insights into Immune Systems of Bats. Front. Immunol. 2020, 11, 26. [Google Scholar] [CrossRef] [PubMed]
- Calisher, C.H.; Childs, J.E.; Field, H.E.; Holmes, K.V.; Schountz, T. Bats: Important Reservoir Hosts of Emerging Viruses. Clin. Microbiol. Rev. 2006, 19, 531–545. [Google Scholar] [CrossRef] [PubMed]
- Fong, T.-T.; Lipp, E.K. Enteric Viruses of Humans and Animals in Aquatic Environments: Health Risks, Detection, and Potential Water Quality Assessment Tools. Microbiol. Mol. Biol. Rev. 2005, 69, 357–371. [Google Scholar] [CrossRef] [PubMed]
- Lynch, K.L.; Gooding, L.R.; Garnett-Benson, C.; Ornelles, D.A.; Avgousti, D.C. Epigenetics and the Dynamics of Chromatin during Adenovirus Infections. FEBS Lett. 2019, 593, 3551–3570. [Google Scholar] [CrossRef] [PubMed]
- Harrach, B.; Tarján, Z.L.; Benkő, M. Adenoviruses across the Animal Kingdom: A Walk in the Zoo. FEBS Lett. 2019, 593, 3660–3673. [Google Scholar] [CrossRef]
- Benko, M.; Aoki, K.; Arnberg, N.; Davison, A.J.; Echavarria, M.; Hess, M.; Jones, M.S.; Kajan, G.L.; Kajon, A.E.; Mittal, S.K.; et al. ICTV Virus Taxonomy Profile: Adenoviridae 2022. J. Gen. Virol. 2022, 103, 001721. [Google Scholar] [CrossRef]
- Whitley, R. Medical Microbiology; Baron, S., Ed.; University of Texas Medical Branch: Galveston, TX, USA, 1996. [Google Scholar]
- Molnár, V.; Jánoska, M.; Harrach, B.; Glávits, R.; Pálmai, N.; Rigó, D.; Sós, E.; Liptovszky, M. Detection of a Novel Bat Gammaherpesvirus in Hungary. Acta Vet. Hung. 2008, 56, 529–538. [Google Scholar] [CrossRef]
- Bascones-Martínez, A.; Pousa-Castro, X. Herpesvirus. Av. Odontoestomatol. 2011, 27, 11–24. [Google Scholar] [CrossRef]
- Young, C.C.W.; Olival, K.J. Optimizing Viral Discovery in Bats. PLoS ONE 2016, 11, e0149237. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Preferred Reporting Items for Systematic Reviews and Meta-Analyses: The PRISMA Statement. PLOS Med. 2009, 151, e1000097. [Google Scholar]
- Page, M.J.; Moher, D.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. PRISMA 2020 Explanation and Elaboration: Updated Guidance and Exemplars for Reporting Systematic Reviews. BMJ 2021, 372, n160. [Google Scholar] [CrossRef]
- Wilson, D.E.; Mittermeirer, R.A. (Eds.) Bats, Handbook of the Mammals of the World, 1st ed.; Lynx Edicions: Barcelona, Spain, 2019; Volume 9, p. 1008. [Google Scholar]
- Hammer, Ø.; Harper, D.; Ryan, P. PAST: Paquete de Programas de Estadística Paleontológica Para Enseñanza y Análisis de Datos. Palaeontol. Electrón. 2001, 4, 4. [Google Scholar]
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An Open Source Software for Exploring and Manipulating Networks. Int. Conf. Web Soc. Media 2009, 3, 361–362. [Google Scholar] [CrossRef]
- Tahamtan, I.; Bornmann, L. Altmetrics and Societal Impact Measurements: Match or Mismatch? A Literature Review. Prof. Inf. 2020, 29, e290102. [Google Scholar] [CrossRef]
- Altmetric. Altmetric Database in JSON Format; 2023. Unpublished Raw Data. Available online: https://www.altmetric.com/about-us/our-data/ (accessed on 1 January 2023).
- Okubo, Y. Bibliometric Indicators and Analysis of Research Systems: Methods and Examples; OECD Science, Technology and Industry Working Papers No. 1997/01; OECD Publishing: Paris, France, 1997. [Google Scholar]
- Hintze, J. NCSS Statistical Software, version 12; NCSS, LLC: New Canaan, CT, USA, 2018.
- Waruhiu, C.; Ommeh, S.; Obanda, V.; Agwanda, B.; Gakuya, F.; Ge, X.-Y.; Yang, X.-L.; Wu, L.-J.; Zohaib, A.; Hu, B.; et al. Molecular Detection of Viruses in Kenyan Bats and Discovery of Novel Astroviruses, Caliciviruses and Rotaviruses. Virol. Sin. 2017, 32, 101–114. [Google Scholar] [CrossRef]
- Zheng, X.Y.; Qiu, M.; Chen, H.F.; Chen, S.W.; Xiao, J.P.; Jiang, L.N.; Huo, S.-T.; Shi, T.-L.; Ma, L.-Z.; Liu, S.; et al. Molecular Detection and Phylogenetic Characterization of Bat and Human Adenoviruses in Southern China. Vector-Borne Zoonotic Dis. 2016, 16, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Mishra, N.; Fagbo, S.F.; Alagaili, A.N.; Nitido, A.; Williams, S.H.; Ng, J.; Lee, B.; Durosinlorun, A.; Garcia, J.A.; Jain, K.; et al. A Viral Metagenomic Survey Identifies Known and Novel Mammalian Viruses in Bats from Saudi Arabia. PLoS ONE 2019, 14, e0214227. [Google Scholar] [CrossRef]
- Griffiths, M.E.; Bergner, L.M.; Broos, A.; Meza, D.K.; Filipe, A.D.S.; Davison, A.; Tello, C.; Becker, D.J.; Streicker, D.G. Epidemiology and Biology of a Herpesvirus in Rabies Endemic Vampire Bat Populations. Nat. Commun. 2020, 11, 5951. [Google Scholar] [CrossRef]
- Wu, Z.; Yang, L.; Ren, X.; He, G.; Zhang, J.; Yang, J.; Qian, Z.; Dong, J.; Sun, L.; Zhu, Y.; et al. Deciphering the Bat Virome Catalog to Better Understand the Ecological Diversity of Bat Viruses and the Bat Origin of Emerging Infectious Diseases. ISME J. 2016, 10, 609–620. [Google Scholar] [CrossRef]
- Ntumvi, N.F.; Diffo, J.L.D.; Tamoufe, U.; Ndze, V.N.; Takuo, J.M.; Mouiche, M.M.M.; Nwobegahay, J.; Lebreton, M.; Gillis, A.; Rimoin, A.W.; et al. Evaluation of Bat Adenoviruses Suggests Co-Evolution and Host Roosting Behaviour as Drivers for Diversity. Microb. Genomics 2021, 7, 000561. [Google Scholar] [CrossRef]
- Li, Y.; Ge, X.; Zhang, H.; Zhou, P.; Zhu, Y.; Zhang, Y.; Yuan, J.; Wang, L.-F.; Shi, Z. Host Range, Prevalence, and Genetic Diversity of Adenoviruses in Bats. J. Virol. 2010, 84, 3889–3897. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Yang, X.-L.; Li, B.; Liu, Q.; Zhang, Q.; Liu, H.; Kan, H.P.; Wong, K.C.; Chek, S.N.; He, X.; et al. Detection of Diverse Viruses in Alimentary Specimens of Bats in Macau. Virol. Sin. 2017, 32, 226–234. [Google Scholar] [CrossRef]
- Watanabe, S.; Ueda, N.; Iha, K.; Masangkay, J.S.; Fujii, H.; Alviola, P.; Mizutani, T.; Maeda, K.; Yamane, D.; Walid, A.; et al. Detection of a New Bat Gammaherpesvirus in the Philippines. Virus Genes 2009, 39, 90–93. [Google Scholar] [CrossRef]
- Hermida, L.R.J.; Cadar, D.; Koundouno, F.R.; Juste, J.; Bialonski, A.; Baum, H.; García-Mudarra, J.L.; Hakamaki, H.; Bencsik, A.; Nelson, E.V.; et al. Metagenomic Snapshots of Viral Components in Guinean Bats. Microorganisms 2021, 9, 599. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.-Y.; Qiu, M.; Guan, W.-J.; Li, J.-M.; Chen, S.-W.; Cheng, M.-J.; Huo, S.-T.; Chen, Z.; Wu, Y.; Jiang, L.-N.; et al. Viral Metagenomics of Six Bat Species in Close Contact with Humans in Southern China. Arch. Virol. 2018, 163, 73–88. [Google Scholar] [CrossRef] [PubMed]
- Kamau, J.; Ergunay, K.; Webala, P.W.; Justi, S.A.; Bourke, B.P.; Kamau, M.W.; Hassell, J.; Chege, M.N.; Mwaura, D.K.; Simiyu, C.; et al. A Novel Coronavirus and a Broad Range of Viruses in Kenyan Cave Bats. Viruses 2022, 14, 2820. [Google Scholar] [CrossRef]
- Watanabe, S.; Maeda, K.; Suzuki, K.; Ueda, N.; Iha, K.; Taniguchi, S.; Shimoda, H.; Kato, K.; Yoshikawa, Y.; Morikawa, S.; et al. Novel Betaherpesvirus in Bats. Emerg. Infect. Dis. 2010, 16, 986–988. [Google Scholar] [CrossRef]
- He, B.; Li, Z.; Yang, F.; Zheng, J.; Feng, Y.; Guo, H.; Li, Y.; Wang, Y.; Su, N.; Zhang, F.; et al. Virome Profiling of Bats from Myanmar by Metagenomic Analysis of Tissue Samples Reveals More Novel Mammalian Viruses. PLoS ONE 2013, 8, e61950. [Google Scholar] [CrossRef]
- Kimprasit, T.; Nunome, M.; Iida, K.; Murakami, Y.; Wong, M.L.; Wu, C.H.; Kobayashi, R.; Hengjan, Y.; Takemae, H.; Yonemitsu, K.; et al. Dispersal History of Miniopterus fuliginosus Bats and Their Associated Viruses in East Asia. PLoS ONE 2021, 16, e0244006. [Google Scholar] [CrossRef]
- Dietrich, M.; Kearney, T.; Seamark, E.C.J.; Paweska, J.T.; Markotter, W. Synchronized Shift of Oral, Faecal and Urinary Microbiotas in Bats and Natural Infection Dynamics during Seasonal Reproduction. R. Soc. Open Sci. 2018, 5, 180041. [Google Scholar] [CrossRef] [PubMed]
- Holz, P.H.; Lumsden, L.F.; Druce, J.; Legione, A.R.; Vaz, P.; Devlin, J.M.; Hufschmid, J. Virus Survey in Populations of Two Subspecies of Bent-Winged Bats (Miniopterus orianae bassanii and oceanensis) in South-Eastern Australia Reveals a High Prevalence of Diverse Herpesviruses. PLoS ONE 2018, 13, e0197625. [Google Scholar] [CrossRef] [PubMed]
- Pozo, F.; Juste, J.; Vázquez-Morón, S.; Aznar-López, C.; Ibáñez, C.; Garin, I.; Aihartza, J.; Casas, I.; Tenorio, A.; Echevarría, J.E. Identification of Novel Betaherpesviruses in Iberian Bats Reveals Parallel Evolution. PLoS ONE 2016, 11, e0169153. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.; Yang, X.L.; Ge, X.Y.; Peng, C.; Liu, H.Z.; Zhang, Y.Z.; Zhang, L.B.; Shi, Z.L. Novel Bat Adenoviruses with Low G+C Content Shed New Light on the Evolution of Adenoviruses. J. Gen. Virol. 2017, 98, 739–748. [Google Scholar] [CrossRef]
- Šimić, I.; Zorec, T.M.; Lojkić, I.; Krešić, N.; Poljak, M.; Cliquet, F.; Picard-Meyer, E.; Wasniewski, M.; Zrnčić, V.; Ćukušić, A.; et al. Viral Metagenomic Profiling of Croatian Bat Population Reveals Sample and Habitat Dependent Diversity. Viruses 2020, 12, 891. [Google Scholar] [CrossRef]
- Zheng, X.; Qiu, M.; Chen, S.; Xiao, J.; Ma, L.; Liu, S.; Zhou, J.; Zhang, Q.; Li, X.; Chen, Z.; et al. High Prevalence and Diversity of Viruses of the Subfamily Gammaherpesvirinae, Family Herpesviridae, in Fecal Specimens from Bats of Different Species in Southern China. Arch. Virol. 2016, 161, 135–140. [Google Scholar] [CrossRef]
- Conrardy, C.; Tao, Y.; Kuzmin, I.V.; Niezgoda, M.; Agwanda, B.; Breiman, R.F.; Anderson, L.J.; Rupprecht, C.E.; Tong, S. Short Report: Molecular Detection of Adenoviruses, Rhabdoviruses, and Paramyxoviruses in Bats from Kenya. Am. J. Trop. Med. Hyg. 2014, 91, 258–266. [Google Scholar] [CrossRef]
- Moreira, M.L.; Botto, N.G.; Malta, L.; Delfraro, A.; Frabasile, S. Ecological and Conservation Significance of Herpesvirus Infection in Neotropical Bats. Ecohealth 2021, 18, 123–133. [Google Scholar] [CrossRef]
- James, S.; Donato, D.; de Thoisy, B.; Lavergne, A.; Lacoste, V. Novel Herpesviruses in Neotropical Bats and Their Relationship with Other Members of the Herpesviridae Family. Infect. Genet. Evol. 2020, 84, 104367. [Google Scholar] [CrossRef]
- Bolatti, E.M.; Zorec, T.M.; Montani, M.E.; Hošnjak, L.; Chouhy, D.; Viarengo, G.; Casal, P.E.; Barquez, R.M.; Poljak, M.; Giri, A.A. A Preliminary Study of the Virome of the South American Free-Tailed Bats (Tadarida brasiliensis) and Identification of Two Novel Mammalian Viruses. Viruses 2020, 12, 422. [Google Scholar] [CrossRef]
- Li, Y.; Altan, E.; Reyes, G.; Halstead, B.; Deng, X.; Delwart, E. Virome of Bat Guano from Nine Northern California Roosts. J. Virol. 2021, 95, e02185-18. [Google Scholar] [CrossRef] [PubMed]
- Diakoudi, G.; Lanave, G.; Moreno, A.; Chiapponi, C.; Sozzi, E.; Prosperi, A.; Larocca, V.; Losurdo, M.; Decaro, N.; Martella, V.; et al. Surveillance for Adenoviruses in Bats in Italy. Viruses 2019, 11, 523. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Moore, N.E.; Murray, Z.L.; McInnes, K.; White, D.J.; Tompkins, D.M.; Hall, R.J. Discovery of Novel Virus Sequences in an Isolated and Threatened Bat Species, the New Ze Aland Lesser Short-Tailed Bat (Mystacina tuberculata). J. Gen. Virol. 2015, 96, 2442–2452. [Google Scholar] [CrossRef]
- Lima, F.E.D.S.; Cibulski, S.P.; Elesbao, F.; Carnieli Junior, P.; Batista, H.B.D.C.R.; Roehe, P.M.; Franco, A.C. First Detection of Adenovirus in the Vampire Bat (Desmodus rotundus) in Brazil. Virus Genes 2013, 47, 378–381. [Google Scholar] [CrossRef]
- Wray, A.K.; Olival, K.J.; Morán, D.; Lopez, M.R.; Alvarez, D.; Navarrete-Macias, I.; Liang, E.; Simmons, N.B.; Lipkin, W.I.; Daszak, P.; et al. Viral Diversity, Prey Preference, and Bartonella Prevalence in Desmodus rotundus in Guatemala. Ecohealth 2016, 13, 761–774. [Google Scholar] [CrossRef] [PubMed]
- Escalera-Zamudio, M.; Rojas-Anaya, E.; Sergios-Orestis, K.; Taboada, B.; Loza-Rubio, E.; Méndez-Ojeda, M.L.; Arias, C.F.; Osterrieder, N.; Greenwood, A.D. Bats, Primates, and the Evolutionary Origins and Diversification of Mammalian Gammaherpesviruses. MBio 2016, 7, e01425-16. [Google Scholar] [CrossRef]
- Salmier, A.; Tirera, S.; De Thoisy, B.; Franc, A.; Darcissac, E.; Donato, D.; Bouchier, C.; Lacoste, V.; Lavergne, A. Virome Analysis of Two Sympatric Bat Species (Desmodus rotundus and Molossus molossus) in French Guiana. PLoS ONE 2017, 12, e0186943. [Google Scholar] [CrossRef] [PubMed]
- Janíková, M.; Briestenská, K.; Salinas-Ramos, V.B.; Mistríková, J.; Kabát, P. Molecular Detection of Murine Gammaherpesvirus 68 (MHV-68) in Bats from Mexico. Acta Virol. 2020, 64, 509–511. [Google Scholar] [CrossRef]
- Razafindratsimandresy, R.; Jeanmaire, E.M.; Counor, D.; Vasconcelos, P.F.; Sall, A.A.; Reynes, J.M. Partial Molecular Characterization of Alphaherpesviruses Isolated from Tropical Bats. J. Gen. Virol. 2009, 90, 44–47. [Google Scholar] [CrossRef]
- Barrón-Rodríguez, R.J.; Parra-Laca, R.; Rojas-Anaya, E.; Romero-Espinoza, J.Á.I.; Ayala-Sumuano, J.T.; Vázquez-Pérez, J.A.; García-Espinosa, G.; Loza-Rubio, E. Evidence of Viral Communities in Three Species of Bats from Rural Environment in Mexico. Acta Chiropterologica 2022, 24, 249–264. [Google Scholar] [CrossRef]
- Sasaki, M.; Setiyono, A.; Handharyani, E.; Kobayashi, S.; Rahmadani, I.; Taha, S.; Adiani, S.; Subangkit, M.; Nakamura, I.; Sawa, H.; et al. Isolation and Characterization of a Novel Alphaherpesvirus in Fruit Bats. J. Virol. 2014, 88, 9819–9829. [Google Scholar] [CrossRef] [PubMed]
- Wada, Y.; Sasaki, M.; Setiyono, A.; Handharyani, E.; Rahmadani, I.; Taha, S.; Adiani, S.; Latief, M.; Kholilullah, Z.A.; Subangkit, M.; et al. Detection of Novel Gammaherpesviruses from Fruit Bats in Indonesia. J. Med. Microbiol. 2018, 67, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Sano, K.; Okazaki, S.; Taniguchi, S.; Masangkay, J.S.; Puentespina, R.; Eres, E.; Cosico, E.; Quibod, N.; Kondo, T.; Shimoda, H.; et al. Detection of a Novel Herpesvirus from Bats in the Philippines. Virus Genes 2015, 51, 136–139. [Google Scholar] [CrossRef] [PubMed]
- Baker, K.S.; Leggett, R.M.; Bexfield, N.H.; Alston, M.; Daly, G.; Todd, S.; Tachedjian, M.; Holmes, C.E.G.; Crameri, S.; Wang, L.F.; et al. Metagenomic Study of the Viruses of African Straw-Coloured Fruit Bats: Detection of a Chiropteran Poxvirus and Isolation of a Novel Adenovirus. Virology 2013, 441, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, H.; Kajihara, M.; Nao, N.; Shigeno, A.; Fujikura, D.; Hang’Ombe, B.M.; Mweene, A.S.; Mutemwa, A.; Squarre, D.; Yamada, M.; et al. Characterization of a Novel Bat Adenovirus Isolated from Straw-Colored Fruit Bat (Eidolon helvum). Viruses 2017, 9, 371. [Google Scholar] [CrossRef]
- Mendenhall, I.H.; Wen, D.L.H.; Jayakumar, J.; Gunalan, V.; Wang, L.; Mauer-Stroh, S.; Su, Y.C.F.; Smith, G.J.D. Diversity and Evolution of Viral Pathogen Community in Cave Nectar Bats (Eonycteris spelaea). Viruses 2019, 11, 250. [Google Scholar] [CrossRef]
- Maeda, K.; Hondo, E.; Terakawa, J.; Kiso, Y.; Nakaichi, N.; Endoh, D.; Sakai, K.; Morikawa, S.; Mizutani, T. Isolation of Novel Adenovirus from Fruit Bat (Pteropus dasymallus yayeyamae). Emerg. Infect. Dis. 2008, 14, 347–349. [Google Scholar] [CrossRef]
- Inagaki, T.; Yamada, S.; Fujii, H.; Yoshikawa, T.; Shibamura, M.; Harada, S.; Fukushi, S.; Le, M.Q.; Nguyen, C.T.; Nguyen, T.T.T.; et al. Characterization of a Novel Alphaherpesvirus Isolated from the Fruit Bat Pteropus lylei in Vietnam. J. Virol. 2020, 94, e00673-20. [Google Scholar] [CrossRef]
- Anthony, S.J.; Epstein, J.H.; Murray, K.A.; Navarrete-Macias, I.; Zambrana-Torrelio, C.M.; Solovyov, A.; Ojeda-Flores, R.; Arrigo, N.C.; Islam, A.; Ali, K.S.; et al. A Strategy to Estimate Unknown Viral Diversity in Mammals. MBio 2013, 4, e00598-13. [Google Scholar] [CrossRef]
- Van Brussel, K.; Mahar, J.E.; Ortiz-Baez, A.S.; Carrai, M.; Spielman, D.; Boardman, W.S.J.; Baker, M.L.; Beatty, J.A.; Geoghegan, J.L.; Barrs, V.R.; et al. Faecal Virome of the Australian Grey-Headed Flying Fox from Urban/Suburban Environments Contains Novel Coronaviruses, Retroviruses and Sapoviruses. Virology 2022, 576, 42–51. [Google Scholar] [CrossRef]
- Brock, A.P.; Cortés-Hinojosa, G.; Plummer, C.E.; Conway, J.A.; Roff, S.R.; Childress, A.L.; Wellehan, J.F.X. A Novel Gammaherpesvirus in a Large Flying Fox (Pteropus vampyrus) with Blepharitis. J. Vet. Diagnostic Investig. 2013, 25, 433–437. [Google Scholar] [CrossRef] [PubMed]
- Jansen Van Vuren, P.; Allam, M.; Wiley, M.R.; Ismail, A.; Storm, N.; Birkhead, M.; Markotter, W.; Palacios, G.; Paweska, J.T. A Novel Adenovirus Isolated from the Egyptian Fruit Bat in South Africa Is Closely Related to Recent Isolates from China. Sci. Rep. 2018, 8, 9584. [Google Scholar] [CrossRef] [PubMed]
- Jánoska, M.; Vidovszky, M.; Molnár, V.; Liptovszky, M.; Harrach, B.; Benko, M. Novel Adenoviruses and Herpesviruses Detected in Bats. Vet. J. 2011, 189, 118–121. [Google Scholar] [CrossRef]
- Raut, C.G.; Yadav, P.D.; Towner, J.S.; Amman, B.R.; Erickson, B.R.; Cannon, D.L.; Sivaram, A.; Basu, A.; Nichol, S.T.; Mishra, A.C.; et al. Isolation of a Novel Adenovirus from Rousettus leschenaultii Bats from India. Intervirology 2012, 55, 488–490. [Google Scholar] [CrossRef] [PubMed]
- Shete, A.M.; Yadav, P.; Kumar, V.; Nikam, T.; Mehershahi, K.; Kokate, P.; Patil, D.; Mourya, D.T. Development of Polymerase Chain Reaction-Based Diagnostic Tests for Detection of Malsoor Virus & Adenovirus Isolated from Rousettus Species of Bats in Maharashtra, India Development of Polymerase Chain Reaction-Based Diagnostic Tests for Detection of Malso. Indian J. Med. Res. 2017, 145, 90–96. [Google Scholar] [CrossRef]
- Katayama, M.; Murakami, S.; Matsugo, H.; Kamiki, H.; Fujii, M.; Takenaka-Uema, A.; Horimoto, T. Complete Genome Sequence of a Novel Bat Mastadenovirus C Strain Isolated from Rhinolophus Cornutus in Japan. Arch. Virol. 2022, 167, 979–982. [Google Scholar] [CrossRef]
- Vidovszky, M.Z.; Kohl, C.; Boldogh, S.; Görföl, T.; Wibbelt, G.; Kurth, A.; Harrach, B. Random Sampling of the Central European Bat Fauna Reveals the Existence of Numerous Hitherto Unknown Adenoviruses. Acta Vet. Hung. 2015, 63, 508–525. [Google Scholar] [CrossRef] [PubMed]
- Iglesias-Caballero, M.; Juste, J.; Vázquez-Morón, S.; Falcon, A.; Aznar-Lopez, C.; Ibáñez, C.; Pozo, F.; Ruiz, G.; Berciano, J.M.; Garin, I.; et al. New Adenovirus Groups in Western Palaearctic Bats. Viruses 2018, 10, 443. [Google Scholar] [CrossRef]
- Wu, Z.; Ren, X.; Yang, L.; Hu, Y.; Yang, J.; He, G.; Zhang, J.; Dong, J.; Sun, L.; Du, J.; et al. Virome Analysis for Identification of Novel Mammalian Viruses in Bat Species from Chinese Provinces. J. Virol. 2012, 86, 10999–11012. [Google Scholar] [CrossRef]
- Noguchi, K.; Kuwata, R.; Shimoda, H.; Mizutani, T.; Hondo, E.; Maeda, K. The Complete Genomic Sequence of Rhinolophus Gammaherpesvirus 1 Isolated from a Greater Horseshoe Bat. Arch. Virol. 2019, 164, 317–319. [Google Scholar] [CrossRef]
- Wiederkehr, M.A.; Qi, W.; Schoenbaechler, K.; Fraefel, C.; Kubacki, J. Virus Diversity, Abundance, and Evolution in Three Different Bat Colonies in Switzerland. Viruses 2022, 14, 1911. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Zhu, C.; Wang, Y.; Ai, L.; Yang, L.; Ye, F.; Ding, C.; Chen, J.; He, B.; Zhu, J.; et al. Virome Analysis for Identification of Novel Mammalian Viruses in Bats from Southeast China. Sci. Rep. 2017, 7, 10917. [Google Scholar] [CrossRef] [PubMed]
- Ai, L.; Zhu, C.; Zhang, W.; He, T.; Ke, Y.; Wu, J.; Yin, W.; Zou, X.; Ding, C.; Luo, Y.; et al. Genomic Characteristics and Pathogenicity of a New Bat Adenoviruses Strains That Was Isolated in at Sites along the Southeastern Coasts of the P. R. of China from 2015 to 2019. Virus Res. 2022, 308, 198653. [Google Scholar] [CrossRef] [PubMed]
- Cameron, K.; Hayes, B.; Olson, S.H.; Smith, B.R.; Pante, J.; Laudisoit, A.; Goldstein, T.; Joly, D.O.; Bagamboula MPassi, R.; Lange, C.E. Detection of First Gammaherpesvirus Sequences in Central African Bats. New Microbes New Infect. 2020, 36, 100705. [Google Scholar] [CrossRef] [PubMed]
- Briestenská, K.; Janíková, M.; Kabát, P.; Csepányiová, D.; Zukal, J.; Pikula, J.; Kováčová, V.; Linhart, P.; Banďouchová, H.; Mistríková, J. Bats as Another Potential Source of Murine Gammaherpesvirus 68 (MHV-68) in Nature. Acta Virol. 2018, 62, 337–339. [Google Scholar] [CrossRef]
- Prada, D.; Boyd, V.; Baker, M.L.; O’Dea, M.; Jackson, B. Viral Diversity of Microbats within the South West Botanical Province of Western Australia. Viruses 2019, 11, 1157. [Google Scholar] [CrossRef]
- Hackenbrack, N.; Rogers, M.B.; Ashley, R.E.; Keel, M.K.; Kubiski, S.V.; Bryan, J.A.; Ghedin, E.; Holmes, E.C.; Hafenstein, S.L.; Allison, A.B. Evolution and Cryo-Electron Microscopy Capsid Structure of a North American Bat Adenovirus and Its Relationship to Other Mastadenoviruses. J. Virol. 2017, 91, e01504-16. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, E.F.; Haskew, A.N.; Gates, J.E.; Huynh, J.; Moore, C.J.; Frieman, M.B. Metagenomic Analysis of the Viromes of Three North American Bat Species: Viral Diversity among Different Bat Species That Share a Common Habitat. J. Virol. 2010, 84, 13004–13018. [Google Scholar] [CrossRef]
- Subudhi, S.; Rapin, N.; Dorville, N.; Hill, J.E.; Town, J.; Willis, C.K.R.; Bollinger, T.K.; Misra, V. Isolation, Characterization and Prevalence of a Novel Gammaherpesvirus in Eptesicus fuscus, the North American Big Brown Bat. Virology 2018, 516, 227–238. [Google Scholar] [CrossRef]
- Gerow, C.M.; Rapin, N.; Voordouw, M.J.; Elliot, M.; Misra, V.; Subudhi, S. Arousal from Hibernation and Reactivation of Eptesicus fuscus Gammaherpesvirus (EfHV) in Big Brown Bats. Transbound. Emerg. Dis. 2018, 66, 1054–1062. [Google Scholar] [CrossRef]
- Mühldorfer, K.; Speck, S.; Kurth, A.; Lesnik, R.; Freuling, C.; Müller, T.; Kramer-Schadt, S.; Wibbelt, G. Diseases and Causes of Death in European Bats: Dynamics in Disease Susceptibility and Infection Rates. PLoS ONE 2011, 6, e29773. [Google Scholar] [CrossRef]
- Han, H.-J.; Wen, H.-L.; Zhao, L.; Liu, J.-W.; Luo, L.-M.; Zhou, C.-M.; Qin, X.-R.; Zhu, Y.-L.; Liu, M.-M.; Qi, R.; et al. Novel Coronaviruses, Astroviruses, Adenoviruses and Circoviruses in Insectivorous Bats from Northern China. Zoonoses Public Health 2017, 64, 636–646. [Google Scholar] [CrossRef]
- Cholleti, H.; De Jong, J.; Blomström, A.-L.; Berg, M. Characterization of Pipistrellus pygmaeus Bat Virome from Sweden. Palliat. Med. 2022, 14, 1654. [Google Scholar] [CrossRef]
- Kohl, C.; Brinkmann, A.; Radonić, A.; Dabrowski, P.W.; Mühldorfer, K.; Nitsche, A.; Wibbelt, G.; Kurth, A. The Virome of German Bats: Comparing Virus Discovery Approaches. Sci. Rep. 2021, 11, 7430. [Google Scholar] [CrossRef] [PubMed]
- Armero, A.; Li, R.; Bienes, K.M.; Chen, X.; Li, J.; Xu, S.; Chen, Y.; Hughes, A.C.; Berthet, N.; Wong, G. Myotis fimbriatus Virome, a Window to Virus Diversity and Evolution in the Genus Myotis. Viruses 2022, 14, 1899. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Matsugo, H.; Maruyama, J.; Kamiki, H.; Takada, A.; Maeda, K.; Takenaka-Uema, A.; Tohya, Y.; Murakami, S.; Horimoto, T. Characterization of a Novel Species of Adenovirus from Japanese Microbat and Role of CXADR as Its Entry Factor. Sci. Rep. 2019, 9, 573. [Google Scholar] [CrossRef]
- Tandler, B. Cytomegalovirus in the Principal Submandibular Gland of the Little Brown Bat, Myotis lucifugus. J. Comp. Pathol. 1996, 114, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wibbelt, G.; Kurth, A.; Yasmum, N.; Bannert, M.; Nagel, S.; Nitsche, A.; Ehlers, B. Discovery of Herpesviruses in Bats. J. Gen. Virol. 2007, 88, 2651–2655. [Google Scholar] [CrossRef]
- Drexler, J.F.; Corman, V.M.; Wegner, T.; Tateno, A.F.; Zerbinati, R.M.; Gloza-Rausch, F.; Seebens, A.; Müller, M.A.; Drosten, C. Amplification of Emerging Viruses in a Bat Colony. Emerg. Infect. Dis. 2011, 17, 449–456. [Google Scholar] [CrossRef]
- Hardmeier, I.; Aeberhard, N.; Qi, W.; Schoenbaechler, K.; Kraettli, H.; Hatt, J.M.; Fraefel, C.; Kubacki, J. Metagenomic Analysis of Fecal and Tissue Samples from 18 Endemic Bat Species in Switzerland Revealed a Diverse Virus Composition Including Potentially Zoonotic Viruses. PLoS ONE 2021, 16, e0252534. [Google Scholar] [CrossRef]
- Dacheux, L.; Cervantes-Gonzalez, M.; Guigon, G.; Thiberge, J.M.; Vandenbogaert, M.; Maufrais, C.; Caro, V.; Bourhy, H. A Preliminary Study of Viral Metagenomics of French Bat Species in Contact with Humans: Identification of New Mammalian Viruses. PLoS ONE 2014, 9, e87194. [Google Scholar] [CrossRef]
- Shabman, R.S.; Shrivastava, S.; Tsibane, T.; Attie, O.; Jayaprakash, A.; Mire, C.E.; Dilley, K.E.; Puri, V.; Stockwell, T.B.; Geisbert, T.W.; et al. Isolation and Characterization of a Novel Gammaherpesvirus from a Microbat Cell Line. mSphere 2016, 1, e00070-15. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.N.; Angiel, M. Two Novel Adenoviruses Found in Cave Myotis Bats (Myotis velifer) in Oklahoma. Virus Genes 2020, 56, 99–103. [Google Scholar] [CrossRef]
- Geldenhuys, M.; Mortlock, M.; Weyer, J.; Bezuidt, O.; Seamark, E.C.J.; Kearney, T.; Gleasner, C.; Erkkila, T.H.; Cui, H.; Markotter, W. A Metagenomic Viral Discovery Approach Identifies Potential Zoonotic and Novel Mammalian Viruses in Neoromicia Bats within South Africa. PLoS ONE 2018, 13, e0194527. [Google Scholar] [CrossRef] [PubMed]
- Feijó, A.; Wang, Y.; Sun, J.; Li, F.; Wen, Z.; Ge, D.; Xia, L.; Yang, Q. Research Trends on Bats in China: A Twenty-First Century Review. Mamm. Biol. 2019, 98, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Aellen, V. Contribution à l’étude Des Chiroptères Du Cameroun. Mémoires Société Sci. Nat. Neuchâtel 1952, 8, 1–121. [Google Scholar]
- Waghiiwimbom, M.D.; Eric-Moise, B.F.; Jules, A.P.; Aimé, T.K.J.; Tamesse, J.L. Diversity and Community Structure of Bats (Chiroptera) in the Centre Region of Cameroon. Afr. J. Ecol. 2020, 58, 211–226. [Google Scholar] [CrossRef]
- Sonntag, M.; Mühldorfer, K.; Speck, S.; Wibbelt, G.; Kurth, A. New Adenovirus in Bats, Germany. Emerg. Infect. Dis. 2009, 15, 2052–2055. [Google Scholar] [CrossRef]
- Mitchell-Jones, A.J.; Amori, G.; Bogdanowicz, W.; Krystufek, B.; Reijnders, P.J.H.; Spitzenberger, F.; Stubbe, M.; Thissen, J.B.M.; Vohralík, V.; Zima, J. The Atlas of European Mammals; Academic Press: London, UK, 1999; Volume 3. [Google Scholar]
- Secemu Listado de Quiropteros de España, Portugal y Gibraltar. Available online: https://secemu.org/wp-content/uploads/2023/04/LISTADO-ESPANA-GIBRALTAR-PORTUGAL-DE-QUIROPTEROS__todas-las-lenguas_actualizada.pdf (accessed on 18 September 2023).
- Loh, E.H.; Nava, A.; Murray, K.A.; Olival, K.J.; Guimarães, M.; Shimabukuro, J.; Zambrana-Torrelio, C.; Fonseca, F.R.; de Oliveira, D.B.L.; de Almeida Campos, A.C.; et al. Prevalence of Bat Viruses Associated with Land-Use Change in the Atlantic Forest, Brazil. Front. Cell. Infect. Microbiol. 2022, 12, 921950. [Google Scholar] [CrossRef]
- Peng, C.; Zhang, D.; Li, C.; Li, Y.; Zhang, H.; Li, N.; Xiao, P. Rhinolophus Sinicus Virome Revealed Multiple Novel Mosquito-Borne Zoonotic Viruses. Front. Cell. Infect. Microbiol. 2022, 12, 960507. [Google Scholar] [CrossRef]
- Froidevaux, J.S.P.; Roemer, C.; Lemarchand, C.; Martí-Carreras, J.; Maes, P.; Rufray, V.; Uriot, Q.; Uriot, S.; López-Baucells, A. Second Capture of Promops centralis (Chiroptera) in French Guiana after 28 Years of Mist-Netting and Description of Its Echolocation and Distress Calls. Acta Amaz. 2020, 50, 327–334. [Google Scholar] [CrossRef]
- Simonin, Y. Emerging Diseases/Viruses Prevention Control, Surveillance, and One Health. Trop. Med. Infect. Dis. 2023, 8, 257. [Google Scholar] [CrossRef]
- Peel, A.J.; Field, H.E.; Aravena, M.R.; Edson, D.; McCallum, H.; Plowright, R.K.; Prada, D. Coronaviruses and Australian Bats: A Review in the Midst of a Pandemic. Aust. J. Zool. 2020, 67, 346–360. [Google Scholar] [CrossRef]
- Hernández-Aguilar, I.; Lorenzo, C.; Santos-Moreno, A.; Naranjo, E.J.; Navarrete-Gutiérrez, D. Coronaviruses in Bats: A Review for the Americas. Viruses 2021, 13, 1226. [Google Scholar] [CrossRef] [PubMed]
- Geldenhuys, M.; Mortlock, M.; Epstein, J.H.; Pawęska, J.T.; Weyer, J.; Markotter, W. Overview of Bat and Wildlife Coronavirus Surveillance in Africa: A Framework for Global Investigations. Viruses 2021, 13, 936. [Google Scholar] [CrossRef] [PubMed]
- Kohl, C.; Nitsche, A.; Kurth, A. Update on Potentially Zoonotic Viruses of European Bats. Vaccines 2021, 9, 690. [Google Scholar] [CrossRef] [PubMed]
- Cohen, L.E.; Fagre, A.C.; Chen, B.; Carlson, C.J.; Becker, D.J. Coronavirus Sampling and Surveillance in Bats from 1996–2019: A Systematic Review and Meta-Analysis. Nat. Microbiol. 2023, 8, 1176–1186. [Google Scholar] [CrossRef] [PubMed]
- Hayward, J.A.; Tachedjian, G. Retroviruses of Bats: A Threat Waiting in the Wings? MBio 2021, 12, e0194121. [Google Scholar] [CrossRef]
- Mbu’U, C.M.; Mbacham, W.F.; Gontao, P.; Sado Kamdem, S.L.; Nlôga, A.M.N.; Groschup, M.H.; Wade, A.; Fischer, K.; Balkema-Buschmann, A. Henipaviruses at the Interface between Bats, Livestock and Human Population in Africa. Vector-Borne Zoonotic Dis. 2019, 19, 455–465. [Google Scholar] [CrossRef]
- Rossetto, F.; Iglesias-Caballero, M.; Liedtke, H.C.; Gomez-Mestre, I.; Berciano, J.M.; Pérez-Suárez, G.; De Paz, O.; Ibáñez, C.; Echevarría, J.E.; Casas, I.; et al. Mating Strategy Is Determinant of Adenovirus Prevalence in European Bats. PLoS ONE 2020, 15, e0226203. [Google Scholar] [CrossRef]
- Finoketti, F.; dos Santos, R.N.; Campos, A.A.S.; Zani, A.L.D.S.; Barboza, C.M.; Fernandes, M.E.S.; de Cassia, P.S.T.; dos Santos, D.D.; Werneck, B.G.; Filho, H.O.; et al. Detection of Adenovirus, Papillomavirus and Parvovirus in Brazilian Bats of the Species Artibeus lituratus and Sturnira lilium. Arch. Virol. 2019, 164, 1015–1025. [Google Scholar] [CrossRef]
- Borkenhagen, L.K.; Fieldhouse, J.K.; Seto, D.; Gray, G.C. Are Adenoviruses Zoonotic? A Systematic Review of the Evidence. Emerg. Microbes Infect. 2019, 8, 1679–1687. [Google Scholar] [CrossRef] [PubMed]
- Benkő, M.; Harrach, B.; Kremer, E.J. Do Nonhuman Primate or Bat Adenoviruses Pose a Risk for Human Health? Futur. Microbiol 2014, 9, 269–272. [Google Scholar] [CrossRef]
- Markotter, W.; Coertse, J.; De Vries, L.; Geldenhuys, M.; Mortlock, M. Bat-Borne Viruses in Africa: A Critical Review. J. Zool. 2020, 311, 77–98. [Google Scholar] [CrossRef] [PubMed]
- Wallau, G.L.; Barbier, E.; Tomazatos, A.; Schmidt-Chanasit, J.; Bernard, E. The Virome of Bats Inhabiting Brazilian Biomes: Knowledge Gaps and Biases towards Zoonotic Viruses. Microbiol. Spectr. 2023, 11, e0407722. [Google Scholar] [CrossRef]
- Hayman, D.T. Bats as Viral Reservoirs. Annu. Rev. Virol. 2016, 3, 77–99. [Google Scholar] [CrossRef] [PubMed]
- Van Brussel, K.; Holmes, E.C. Zoonotic Disease and Virome Diversity in Bats. Curr. Opin. Virol. 2022, 52, 192–202. [Google Scholar] [CrossRef]
- Fagre, A.C.; Kading, R.C. Can Bats Serve as Reservoirs for Arboviruses? Viruses 2019, 11, 215. [Google Scholar] [CrossRef]
- Turmelle, A.S.; Olival, K.J. Correlates of Viral Richness in Bats (Order Chiroptera). Ecohealth 2009, 6, 522–539. [Google Scholar] [CrossRef]
- Simmons, N.B.; Cirranello, A.L. Bat Species of the World: A Taxonomic and Geographic Database. Version 1.4. Available online: https://batnames.org (accessed on 12 February 2023).
- Han, H.J.; Wen, H.L.; Zhou, C.M.; Chen, F.F.; Luo, L.M.; Liu, J.W.; Yu, X.J. Bats as Reservoirs of Severe Emerging Infectious Diseases. Virus Res. 2015, 205, 1–6. [Google Scholar] [CrossRef]
- Halpin, K.; Young, P.L.; Field, H.E.; Mackenzie, J.S. Isolation of Hendra Virus from Pteropid Bats: A Natural Reservoir of Hendra Virus. J. Gen. Virol. 2000, 81, 1927–1932. [Google Scholar] [CrossRef]
- Phelps, K.L.; Hamel, L.; Alhmoud, N.; Ali, S.; Bilgin, R.; Sidamonidze, K.; Urushadze, L.; Karesh, W.; Olival, K.J. Bat Research Networks and Viral Surveillance: Gaps and Opportunities in Western Asia. Viruses 2019, 11, 240. [Google Scholar] [CrossRef] [PubMed]
- Calisher, C.H. Viruses in Bats: A Historic Review. In Bats and Viruses: A New Frontier of Emerging Infectious Diseases; Wang, L.-F., Cowled, C., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 23–45. [Google Scholar]
- Wang, L.F.; Crameri, G. Emerging Zoonotic Viral Diseases. Rev. Sci. Tech. 2014, 33, 569–581. [Google Scholar] [CrossRef] [PubMed]
- Letko, M.; Seifert, S.N.; Olival, K.J.; Plowright, R.K.; Munster, V.J. Bat-Borne Virus Diversity, Spillover and Emergence. Nat. Rev. Microbiol. 2020, 18, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Weber, N.; Nagy, M.; Markotter, W.; Schaer, J.; Puechmaille, S.J.; Sutton, J.; Dávalos, L.M.; Dusabe, M.-C.; Ejotre, I.; Fenton, M.B.; et al. Robust Evidence for Bats as Reservoir Hosts Is Lacking in Most African Virus Studies: A Review and Call to Optimize Sampling and Conserve Bats. Biol. Lett. 2023, 19, 20230358. [Google Scholar] [CrossRef]
- Puechmaille, S.J.; Ar Gouilh, M.; Dechmann, D.; Fenton, B.; Geiselman, C.; Medellin, R.; Mittermeier, R.; Racey, P.; Reeder, D.A.M.; Schaer, J.; et al. Misconceptions and Misinformation about Bats and Viruses. Int. J. Infect. Dis. 2021, 105, 606–607. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AdVs | Bat Family | Bat Species | Country | Positive | Amplification PCR | Reference |
|---|---|---|---|---|---|---|
| Mastadenovirus | Vespertilionidae | Hypsugo savii | Spain | 3 | DNA Polymerase gene | [74] |
| Vespertilionidae | Hypsugo savii | Italy | 2 | DNA Polymerase gene | [48] | |
| Vespertilionidae | Pipistrellus kuhlii | Italy | 1 | DNA Polymerase gene | [48] | |
| Mastadenovirus Vespertilionid AdV-1 | Vespertilionidae | Nyctalus noctula | Germany | 4 | DNA Polymerase gene | [73] |
| Vespertilionidae | Myotis mystacinus | Germany | 1 | DNA Polymerase gene | [73] | |
| Vespertilionidae | Pipistrellus nathusii | Germany | 1 | DNA Polymerase gene | [73] |
| HSVs | Bat Family | Bat Species | Country | Positive | Identification Technique | References |
|---|---|---|---|---|---|---|
| Gammaherpesvirinae Rhadinovirus BatGHV-1 | Vespertilionidae | Eptesicus serotinus | Germany | 2 | PCR (Glycoprotein B and DPOL gene) | [87,94] |
| Vespertilionidae | Myotis nattereri | Germany | 1 | PCR (Glycoprotein B and DPOL gene) | [94] | |
| Vespertilionidae | Pipistrellus nathusii | Germany | 1 | PCR (Glycoprotein B and DPOL gene) | [94] | |
| Vespertilionidae | Pipistrellus pipistrellus | Germany | 2 | PCR (Glycoprotein B and DPOL gene) | [94] | |
| Gammaherpesvirinae Rhadinovirus BatGHV-2 | Vespertilionidae | Myotis nattereri | Germany | 2 | PCR (DPOL gene) | [94] |
| Vespertilionidae | Myotis myotis | Germany | 1 | PCR (DPOL gene) | [94] | |
| Gammaherpesvirinae Rhadinovirus BatGHV-3 | Vespertilionidae | Nyctalus noctula | Germany | 8 | PCR (DPOL gene) | [87,94] |
| Vespertilionidae | Myotis nattereri | Germany | 1 | PCR (DPOL gene) | [94] | |
| Vespertilionidae | Myotis myotis | Germany | 1 | PCR (DPOL gene) | [94] | |
| Gammaherpesvirinae Rhadinovirus BatGHV-4 | Vespertilionidae | Nyctalus noctula | Germany | 2 | PCR (Glycoprotein B and DPOL gene) | [94] |
| Vespertilionidae | Myotis nattereri | Germany | 1 | PCR (Glycoprotein B and DPOL gene) | [94] | |
| Gammaherpesvirinae Rhadinovirus BatGHV-5 | Vespertilionidae | Pipistrellus nathusii | Germany | 9 | PCR (Glycoprotein B and DPOL gene) | [87,94] |
| Vespertilionidae | Nyctalus noctula | Germany | 1 | PCR (DPOL gene) | [87] | |
| Vespertilionidae | Myotis myotis | Germany | 1 | PCR (DPOL gene) | [87] | |
| Vespertilionidae | Myotis mystacinus | Germany | 1 | PCR (DPOL gene) | [87] | |
| Betaherpesvirinae BatBHV-1 | Vespertilionidae | Myotis nattereri | Germany | 3 | PCR (DPOL gene) | [94] |
| Vespertilionidae | Pipistrellus pipistrellus | Germany | 2 | PCR (DPOL gene) | [94] | |
| Gammaherpesvirinae Rhadinovirus MHV-68 | Vespertilionidae | Myotis blythii | Slovakia | 4 | Virus-neutralizing antibodies | [81] |
| Vespertilionidae | Barbastella barbastella | Slovakia | 1 | Virus-neutralizing antibodies | [81] | |
| Vespertilionidae | Nyctalus noctula | Ukraine | 1 | PCR (ORF50 gene) | [81] | |
| Phyllostomidae | Leptonycteris yerbabuenae * | Mexico | 1 | PCR (ORF50 gene) | [54] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Méndez-Rodríguez, A.; Horta, P.; Zarza, H.; Constante-Pérez, L.G.; Salgado-Mejia, F.; López-Wilchis, R.; Juste, J. Surveying Bat-Hosted Adenoviruses and Herpesviruses: A Comprehensive Analysis. Diversity 2024, 16, 46. https://doi.org/10.3390/d16010046
Méndez-Rodríguez A, Horta P, Zarza H, Constante-Pérez LG, Salgado-Mejia F, López-Wilchis R, Juste J. Surveying Bat-Hosted Adenoviruses and Herpesviruses: A Comprehensive Analysis. Diversity. 2024; 16(1):46. https://doi.org/10.3390/d16010046
Chicago/Turabian StyleMéndez-Rodríguez, Aline, Pedro Horta, Heliot Zarza, Luis Guillermo Constante-Pérez, Fernando Salgado-Mejia, Ricardo López-Wilchis, and Javier Juste. 2024. "Surveying Bat-Hosted Adenoviruses and Herpesviruses: A Comprehensive Analysis" Diversity 16, no. 1: 46. https://doi.org/10.3390/d16010046