1. Introduction
Tropical forests (TFs) cover 7% of the earth’s surface. Worldwide, 28% of dense forests are tropical mountain forests (TMFs). TFs contain 50% of the world’s forest biomass and are considered the most important natural carbon sinks, with a paramount importance in managing the global climate change [
1]. TMF are one of the most diverse and threatened ecosystems on earth; this is especially true for the eastern Andean forests [
2]. The last map of global diversity of vascular plant species [
3] emphasized the areas of TMFs as the most important hotspots of the world.
Furthermore, TFs generate 36% of the net primary terrestrial production, contributing to the regulation of carbon dioxide (CO
2) concentration in the atmosphere [
4,
5]. TMFs harbor hydrographic river basins and therefore, they are an essential component in the water regime regulation [
6]. Some other functions of these forests are producing wood and non-wood products, catching and storing precipitations and humidity, maintaining the quality of the water, and also reducing erosion and protecting against landslides and floods.
The so-called “Tropical Andes” hotspot includes various types of forests known worldwide for their high diversity [
2]. In this type of ecosystem, altitude plays a fundamental role in the distribution of diversity [
7]. Several authors recognized that altitude and its associated variables determine richness, floristic composition and structure of the forests in these montane environments [
8,
9]. In many cases, these variables have a synergic and direct effect on species richness, which generally reaches a peak at approximately 1000 m a.s.l.
In Ecuador, it is estimated that the total number of vascular plant species is between 18,000 and 22,000, one of the highest in the world. Additionally, along the Andes, there is a large number of endemic species restricted to just the middle elevations (900–3000 m) [
10]. Despite their global importance, the TMFs in Ecuador are the most threatened type of ecosystems, mainly due to change in land use [
11]. The latest reports indicate that already in 2005, 51% of the forest area was lost, with a deforestation rate of 1.7%, which is equivalent to 198,000 ha.
The TMFs distributed along the Andes Real Cordillera in southeastern Ecuador, also termed “montane cloud forest” [
12] or “evergreen montane forest” [
13], can be partitioned into “mountain rainforest” located between 1800–2800 m a.s.l. and “high mountain evergreen forest” located between 2800 and 3100 m a.s.l. [
14]. In both cases, the classification was based on widely used physiognomic patterns of the vegetation in response to macroscale geographic regions. Generally, hierarchical vegetation classification models only work at large scales, and elevation data and species distribution models are too broad to be useful. A classification based on structural parameters, diversity and functional traits of the species is still incipient; this is due to the fact that, at a geographic microscale, the response of the community to the high environmental heterogeneity is much more complicated, and the attempts to classify vegetation into physiognomic distinct categories are not straightforward. In this context, our study is based on the hypothesis that by using functional traits of the species, along with environmental variables, we would be able to delimit the different types of forests present in our study area.
3. Results
3.1. Grouping Plots
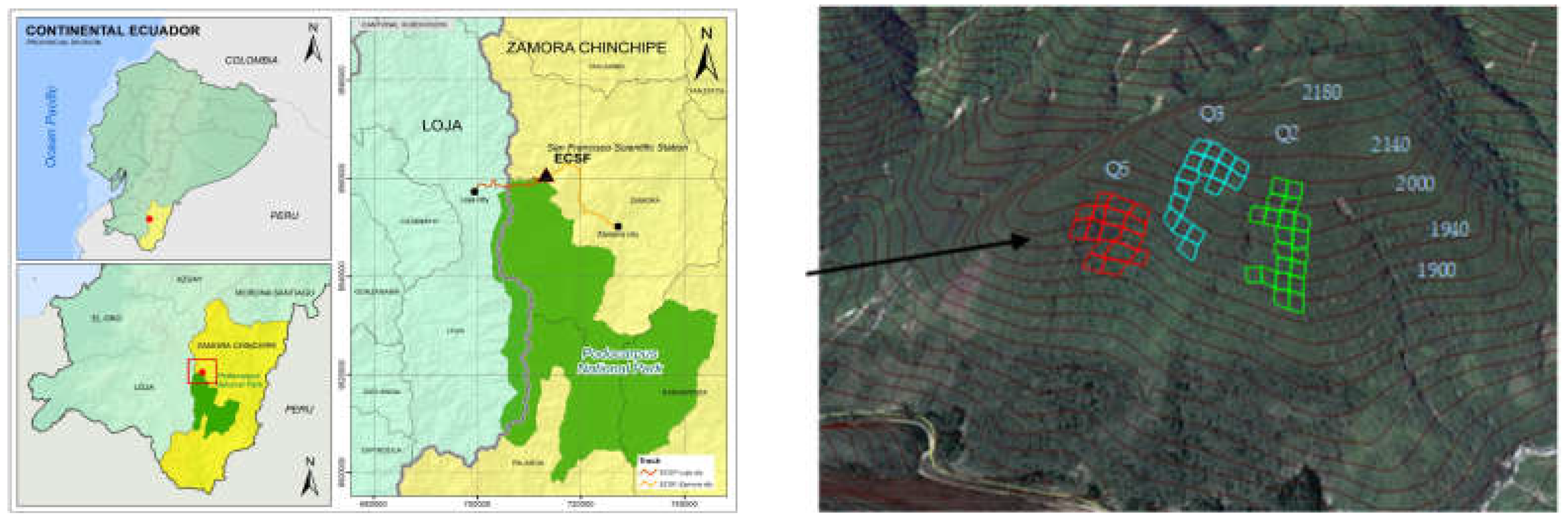
Based on plots of the three sampling sites with floristic similarity and a strong correlation with various attributes of the forest (basal area ha−1, canopy openness, trees ha−1 and alpha diversity), two types of forest could be determined, clearly different in structure and species composition. The spatial distribution of sample plots over the altitudinal gradient implies a change in the structure and diversity of each of the plots. Structural groups were defined based on a correlation between altitude and different attributes of each of the study plots.
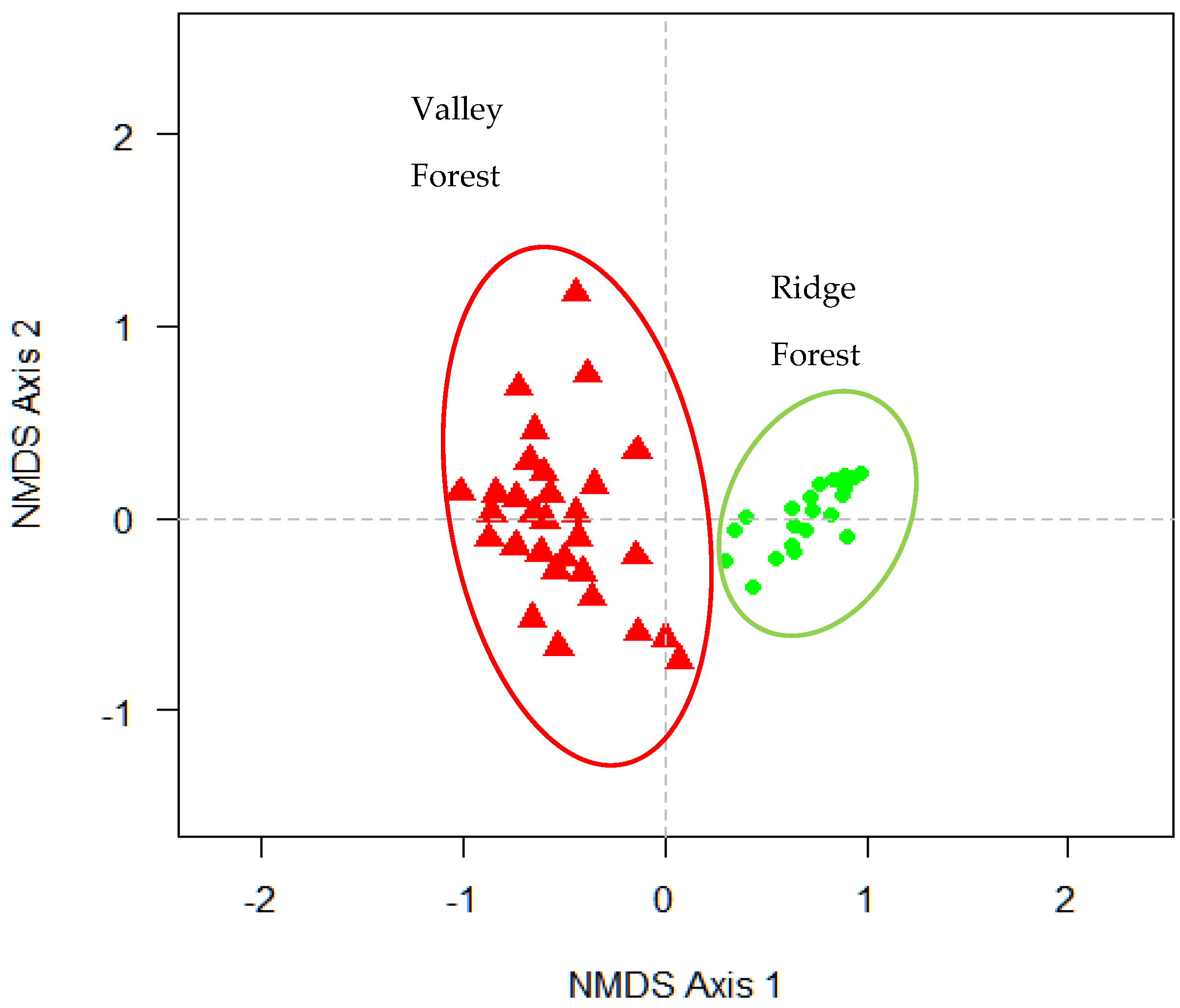
The results of NMDS analysis showed that the sampled plots were divided into two clearly defined groups. The floristic and abundance data used in the matrix formed two groups that we called “forest types”. The first group, named “Valley Forest” (VF), consisted of 15 plots from the Q2 site, and all the plots from the Q5 site. The other group, named “Ridge Forest” (RF), was made up of all the plots of the Q3 site and five plots of the Q2 site (
Figure 2).
3.2. Influence of Altitudinal and Structural Parameters on Grouping Communities
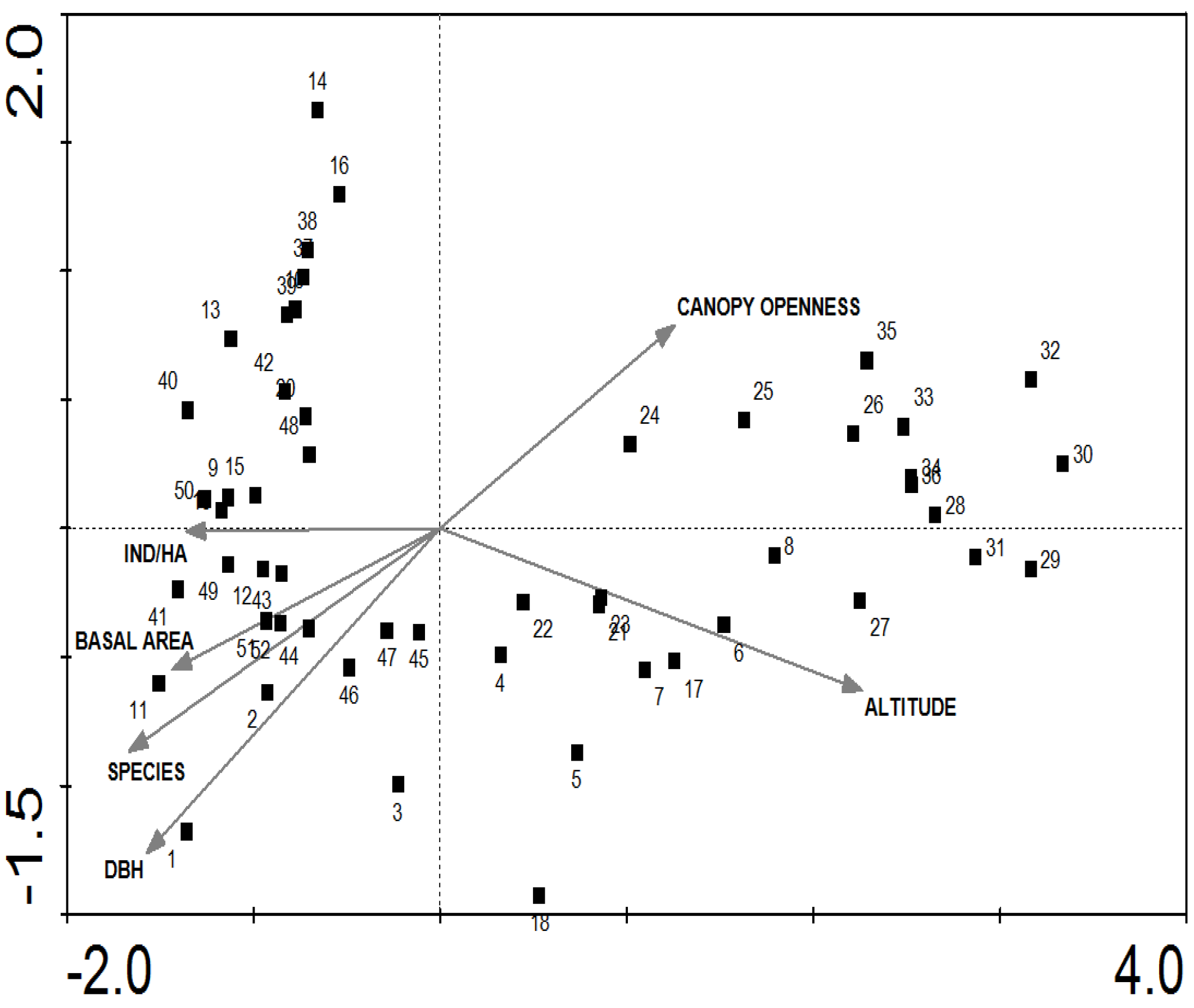
After canonical correspondence Analyses (CCA), altitude was a significative variable in the grouping of the plots, along with basal area and trees per hectare (
Figure 3). To a smaller degree, species diversity (expressed as Shannon index) and canopy openness were also factors that determined the grouping. The analyses of the values of the canonical axes explained 18.1% of variance in species data and 84% of its relationship with environmental variables (
Table 2). This indicates that the structural and floristic characteristics were the result of the influence of the altitudinal gradient and all environmental variables correlated to this (temperature environmental, precipitation, wind, soil, etc.).
3.3. Correlation between Elevation and Characteristic of Forest
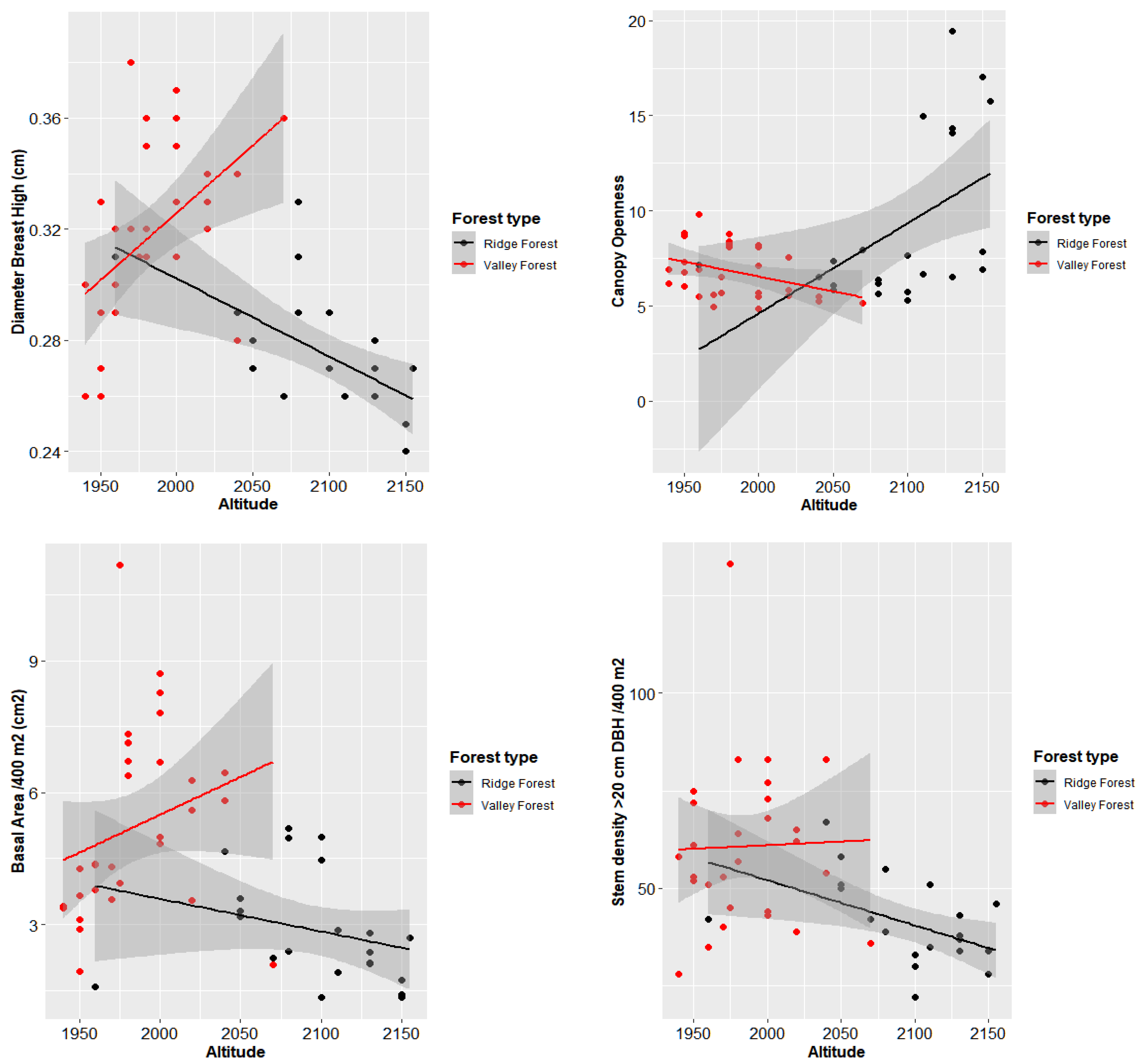
The basal area decreases ad altitude increased; at 1900 m a.s.l., the average basal area can reach 44 m
2 per hectare (includes only the trees >20 cm DBH). In the plots at 2100 m a.s.l., the average basal area had values of 5 to 14 m
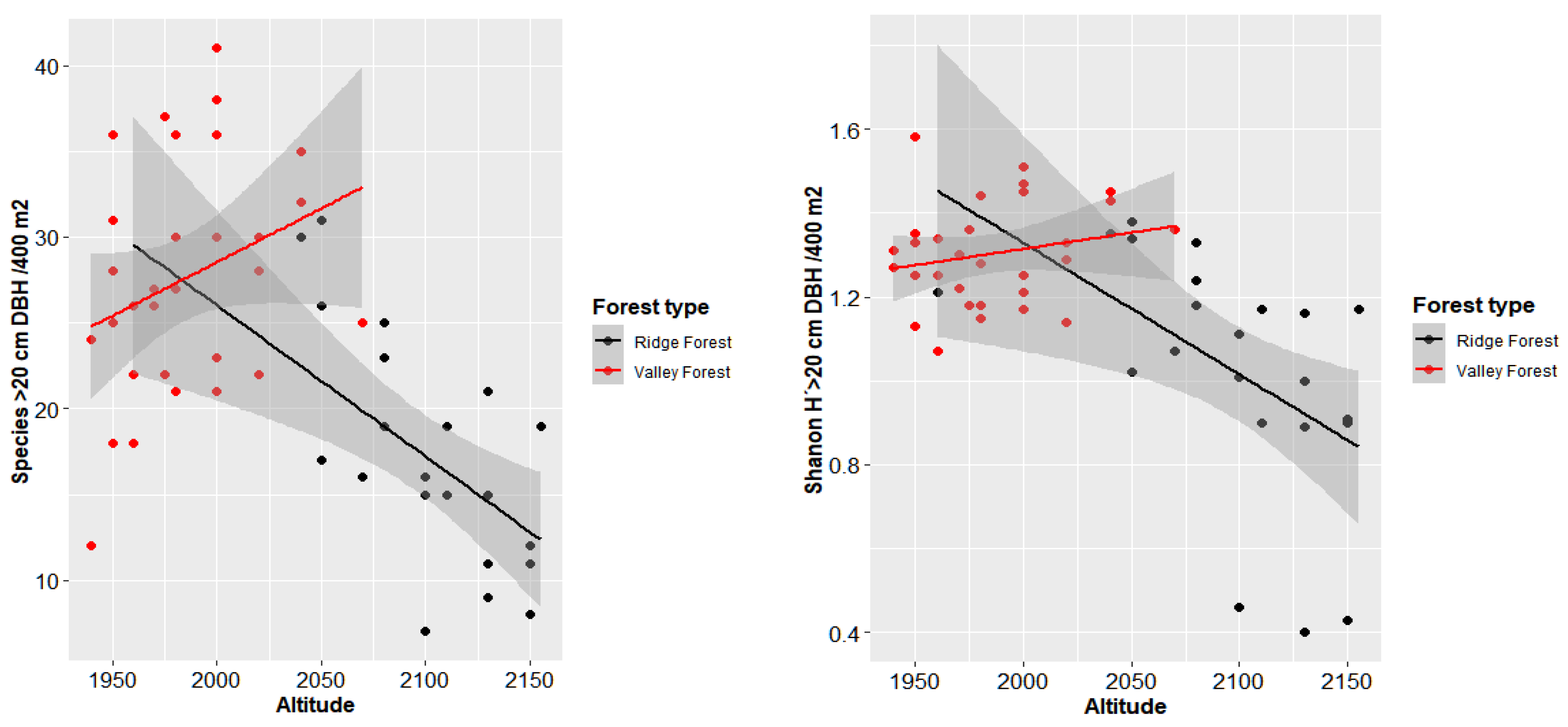
2 per hectare. The same applied to the number of individuals and to the diversity of the sampled sites. This confirmed the trends found in tropical mountain forests, which means that as altitude increases, the diversity of tree species decreases. Canopy openness was higher with the increasing altitude of the plots. The graphs showed a strong correlation between altitude and variable characteristics of each plot (
Figure 4).
The VF is characterized by the presence of Tabebuia chrysantha (Jacq.) G. Nicholson, Cedrela montana Moritz ex Turcz., Inga acreana Harms. and Ficus citrifolia Mill. There are also other species such as Cecropia montana Warb. ex Snethl., Guarea pterorhachis Harms and Heliocarpus americanus L. that are unique to this group.
The RF is characterized by the presence of Podocarpus oleifolius D. Don ex Lamb., Hyeronima moritziana Mull. Arg, Clusia ducuoides Engl., which were species selected as PCT. Other species that characterize the group are Purdiaea nutans Planch., Graffenrieda emarginata (Ruiz and Pav.) and Alchornea grandifolia Triana and Mull Arg.
3.4. Structural Parameters of Floristic Groups
In VF, we encountered 141 species, which belonged to 51 families, while in RF, the diversity was represented by 86 species belonging to 51 families.
Table 3 shows the relative diversity values of the most important families in each of the determined forest types.
Table 4 shows the values of density and relative dominance of the most important VF species.
Table 5 shows the values of density and relative dominance of the most important RF species.
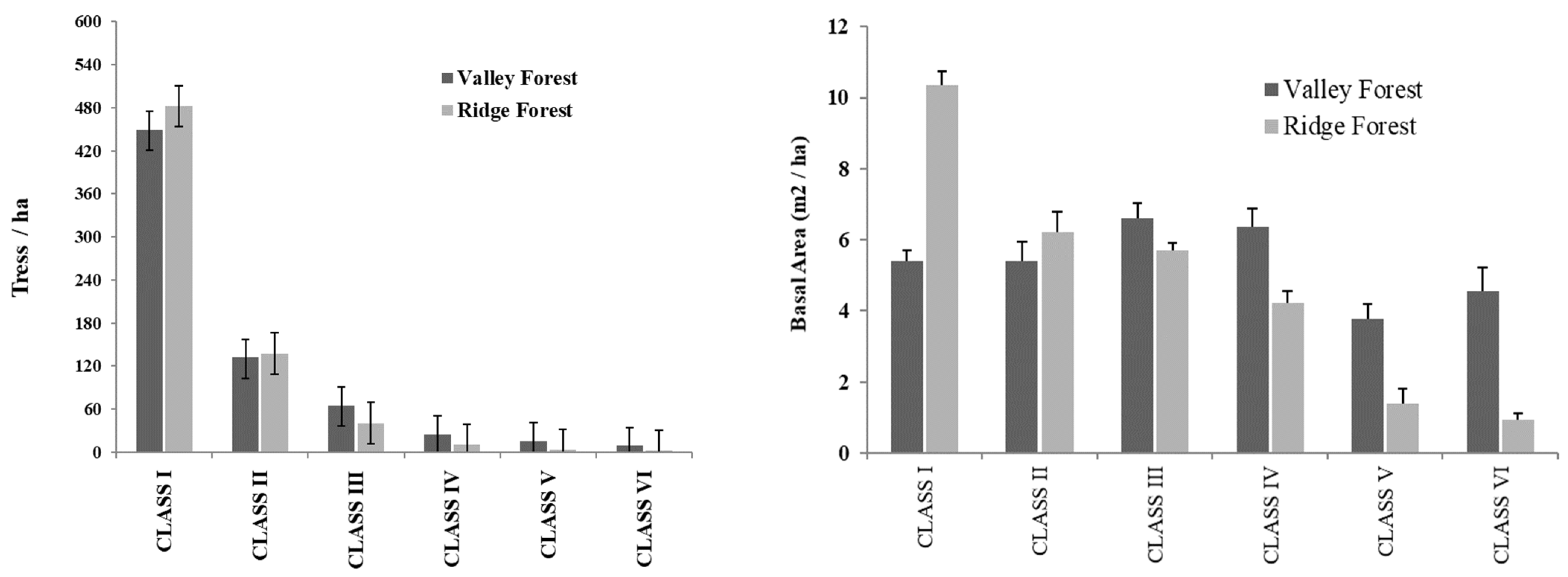
The RF had a total basal area of 54.3 m2, with an average of 10.3 ± 3.1 m2ha−1, while the VF had a total basal area of 168.7 m2, with an average of 21.8 ± 7.9 m2ha−1, considering only the trees >20 cm DBH.
In the RF, there was a total of 862 individuals > 20 cm DBH and an average of 164.2 ± 35.3 trees ha−1. In the Valley Forest, there was a total of 1933 individuals >20 cm DBH and an average of 248.8 ± 81.4 trees ha−1.
The total basal area of trees in the class 5.1–20 cm DBH in the RF was 3.47 m2, which averaged 11.5 ± 3.9 m2 ha−1. In VF, the total basal area was 3.7 m2 in the class 5.1–20 cm DBH, which means an average of 8.8 ± 3.8 m2 ha−1. In RF, there was a total of 443 individuals in the class 5.1–20 cm DBH, with an average of 1464 ± 461.8 trees ha−1. In VF, there was a total of 392 individuals in the class 5.1–20 cm DBH with an average of 906.8 ± 383 ind./ha.
The figures below show the distributions of individuals in all diametric classes in the two forest types (
Figure 5).
The structural difference between the two floristic groups was low, but most evident in the first two diametric classes. In RF, there were more individuals per hectare in the first two classes, while in the Valley Forest, there were more individuals per hectare in the higher diameter classes.
In VF, there were more trees in the larger diameter classes, allowing for harvest at a higher intensity. In RF, the small number of large trees suggests lower harvest intensity, the large number of small individuals and the structure of natural regeneration being a typical feature of these forests at this altitude. Regarding the higher number of individuals in lower diameter classes, the RF contained higher basal area values in these classes than the other group (
Figure 5).
The diameter distributions of trees in RBSF indicate that the trees were distributed in large quantities in smaller diameter classes and the numbers decreased in a negative exponential way for higher diameter classes. This inverse J-shape distribution type represents a good structure which is typical for natural forests (
Figure 5). Each group has its unique species. In the VF, there were 87 exclusive species, representing 47.5% of the total species identified in the study.
3.5. Functional Traits Influence Forest Separation
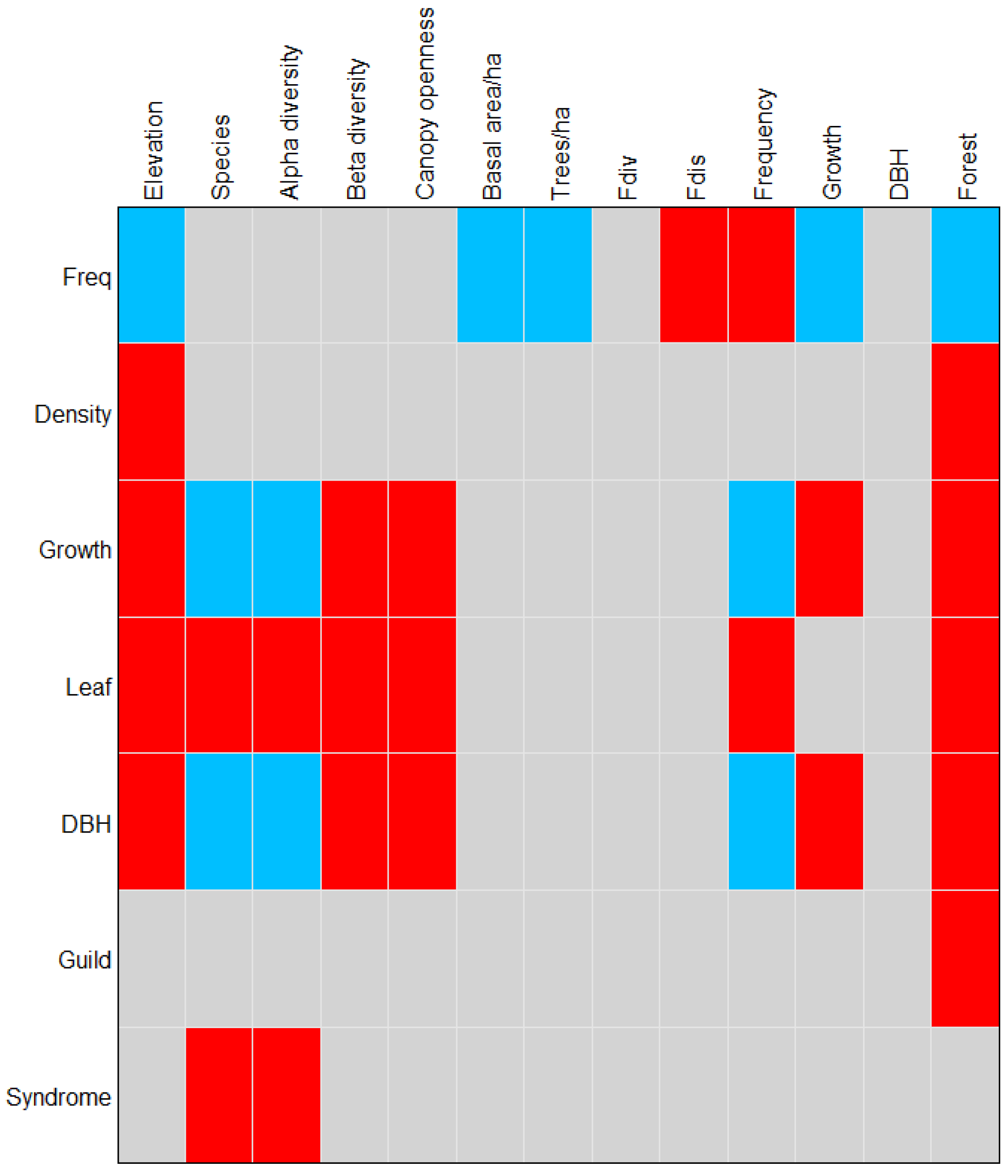
The previous results confirmed that the elevation and its associated variables were the main factors that drive the composition and structure of the determined forest types. Likewise, there was a significant correlation at the community level between environmental factors and the plant functional traits as a response to these environmental changes. When incorporating the functional species traits in the determination of the forest types, we found that the conservative traits, such as the dispersal syndrome, were significantly related to the species (p = 0.01) and the alpha diversity of the forest types (p = 0.04). The ecological guild to which the species belongs was strongly related to the elevation (p = 0.05) and the forest type (p = 0.01).
Species acquisitive traits also play a significant role in characterizing forest types. Diametric growth, as the main acquisitive trait, was significantly related to altitude (p = 0.004) and its related variables that, as we observed previously, play an important role in the separation of forest communities.
Both the acquisitive and the conservative traits are important in the conformation of the forest communities; the Four Corner analysis allowed us to include the characteristics of the species that in other analyses went unnoticed, but were significant when classifying the vegetation.
Figure 6 shows the correlations between the traits and the environmental variables and
Table 6 shows the correlation values between the functional traits of the species and the environmental variables.
4. Discussion
In the proposed model for classification of continental vegetation of Ecuador, Sierra et al. [
14] used the regional division of the territory and included the concept of subregions. The hierarchical model divided the regions into sectors, i.e., each region had subregions, and these in turn were divided into sectors, with nesting vegetation physiognomy (Mangrove Forest, Shrubland, Espinar, Savannah, Paramo and Gelidofitia) and other hierarchical criteria such as environmental criteria (dry, wet, fog), phenological criteria (evergreen, deciduous, semi deciduous) and floristic altitudinal levels (lowland, premontane, lower montane, montane, upper montane), determining 62 cover types, 27 on the coast, 24 in the Andes and 11 in the Amazonia.
However, the classification at the plant formation level included large areas that were evidently heterogeneous with respect to key factors, mainly in the composition and structure of the forests. Other authors, such as Valencia et al. [
12], called this formation “montane cloud forest” in southeastern Ecuador. This is also known as “Evergreen Montane Forest” [
13], using climatic (“cloud”) and functional (“evergreen”) elements in the naming of the formations.
Other classifications proposed altitudinal limits as the main factor in the separation of forests, along with a geomorphological factor [
43], asserting that the forest in the southern region of Ecuador can be divided into evergreen lower montane forests (up to 2100–2200 m a.s.l.) and upper montane forests, up to the tree line. Above ~2700 m, a shrub-dominated sub-paramo prevails, where small patches of Elfin Forest, the so-called Ceja Andina, dominate the landscape. Both montane forest types can be subdivided into a lower slope (ravine) forest and an upper slope (ridge) forest [
44,
45].
A new approach of vegetation classification at smaller scales is based on using variables of structure, taxonomic and functional diversity (traits of the species) and environmental factors (including elevation); as a result, four types of forest can be distinguished by combining different types of classification [
8]. For example, [
46] studied structural parameters; [
47] the floristic trees composition; [
48] the bryophytes species composition and [
49] making a synopsis of previous studies.
All these studies that classified vegetation using environmental parameters associated with elevation were consistent with the categories of vegetation in the RBSF. Even the topographic variables at smaller scales play an important role in the classification of the vegetation, as demonstrated by the studies carried out by [
50] in the same study area and [
51] who combine topographic variables and functional features to determine small-scale species association in tropical forests of China.
In the Andean vegetation classifications, the elevation gradient and its associated variables play an essential role in separating forests. This is also true in our case, where elevation works at the microscale, that is, the elevational influence occurs in relatively small areas and over relatively short distances, influencing not only the structure and composition of forests, but also the variation in the composition of traits in vascular plants [
52], such as leaf size [
53], seed dispersal [
54] and wood density [
55], which are basically the same traits that we used for our analysis.
Although we did not include topographic variables in our analysis, there were reports that these correlated with vegetation classification, especially at a small scale, and were significant in identifying ecological units that included vegetation structure and composition [
56], delimiting forest lowland and montane formations [
57]. The topographic variables also affect the functional traits of the species, especially the mass of seeds and the density of the wood [
51]; in our case, species wood density was also correlated with altitude and forest type.
Regarding the scale of influence, it should be noted that, although we determined in our study that the influence of elevation and associated variables is also significant at microscales, there are cases in which elevation works at larger scales and its evaluation was also significant [
58]. This emphasizes the fact that, in addition to the environmental factors associated with elevation, there are other factors intrinsic to the species that help the differentiation of plant communities.
In a specific approach, altitude and its associated environmental factors are crucial when determining and differentiating forest types. In the VF up to 2050 m a.s.l., the number of trees tends to remain relatively stable, while in the RF, the decline in the number of trees at higher altitudes was evident. This pattern is not strict along the Cordillera de los Andes, but it may be subject to changes in temperature and humidity over relatively short intervals [
59].
Finally, when referring to the groups determined by our study, we can indicate that the VF were characterized by lower stem density, but with greater basal areas (tree diameters) and higher canopies compared to the RF, where less tree species were also present. The differences in forest structure are mainly due to the climatic conditions and prevailing soil types [
60,
61]. At the phytosociological level, the floristic structure and composition of the VF and the RF are coupled to more widely distributed floristic formations, such as the order
Alzateetalia verticillatae and
Purdiaeaetalia nutantis, respectively [
62], which validates the floristic analysis carried out in our study area, by the coincidence of indicator species of each floristic association.
For both floristic groups, the microclimatic and topographic conditions cause the species to find suitable sites and share habitat and topographic preferences of occurrence [
50], an argument that also reinforces the grouping of the species present in the forest of the RBSF.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}