
Diversity and Composition of Caddisflies (Insecta: Trichoptera) along an Elevation Gradient in Southeastern Mexico

Abstract
:1. Introduction
2. Materials and Methods
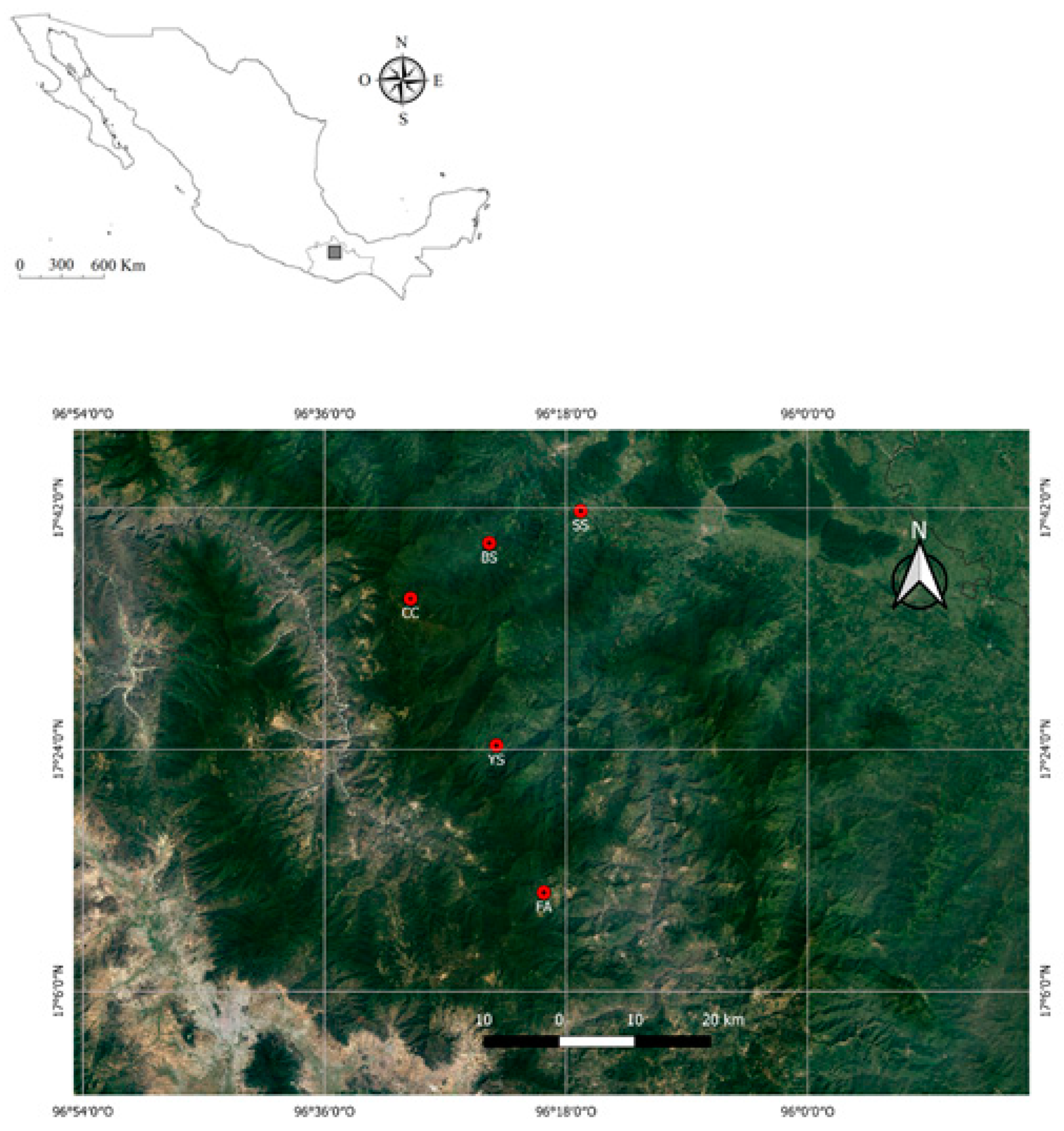
2.1. Study Area
2.2. Field Data Collection and Insect Identification
2.3. Data Analysis
3. Results
3.1. Environmental Factors
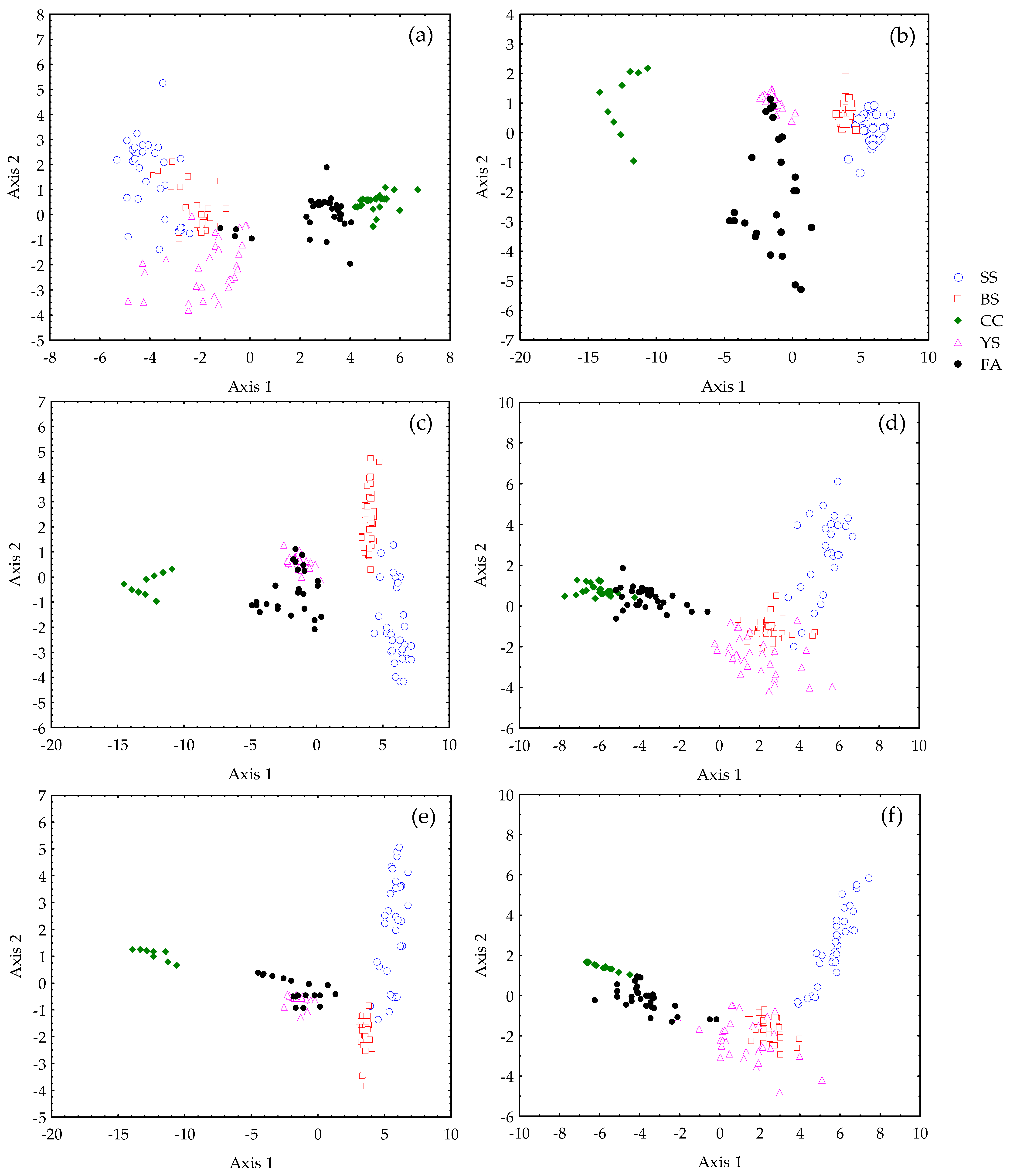
3.2. Richness and Composition
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wiggins, G.B. Larvae of the North America Caddisfly Genera (Trichoptera), 2nd ed.; University of Toronto Press: Toronto, ON, Canada, 1996; pp. 3–25. [Google Scholar]
- Holzenthal, R.W.; Blahnik, R.J.; Prather, A.L.; Kjer, K.M. Order Trichoptera Kirby, 1813 (Insecta), Caddisflies. Zootaxa 2007, 1668, 639–698. [Google Scholar] [CrossRef]
- Morse, J.C.; Frandsen, P.B.; Graf, W.; Thomas, J.A. Diversity and ecosystem services of Trichoptera. Insects 2019, 10, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, J.V. Aquatic Insect Ecology. 1. Biology and Habitat, 1st ed.; John Wiley and Sons, Inc.: New York, NY, USA, 1992; pp. 25–28. [Google Scholar]
- Halffter, G.; Moreno, C.E. Significado biológico de las diversidad alfa, beta y gamma. In Sobre Diversidad Biológica: El Significado de las Diversidades Alfa, Beta y Gamma; Halffter, G., Soberón, J., Koleff, P., Melic, A., Eds.; Zaragoza: m3m-Monografías Tercer Milenio; Sociedad Entomológica Aragonesa, Comisión Nacional para el uso y Conocimiento de la Biodiversidad, Grupo Diversitas y Consejo Nacional de Ciencia y Tecnología: Zaragoza, Spain, 2005; Volume 4, pp. 5–18. [Google Scholar]
- Maurer, B.A.; McGill, B.J. Measurement of species diversity. In Biological Diversity Frontiers in Measurement and Assessment, 1st ed.; Magurran, A.E., McGill, B.J., Eds.; Oxford University Press Inc.: New York, NY, USA, 2011; pp. 55–65. [Google Scholar]
- Whittaker, R.H. Vegetation of the Siskiyou Mountains, Oregon and California. Ecol. Monogr. 1960, 30, 279–338. [Google Scholar] [CrossRef]
- Breda, M.; Restello, R.M.; Giovenardi, R.; Vizzotto, A.P.; Soares, B.; Hepp, L.U. Alpha and beta diversities of Trichoptera (Insecta) assemblages in natural and rural subtropical streams. Acta Limnol. Bras. 2020, 32, e14. [Google Scholar] [CrossRef]
- Baselga, A.; Gómez-Rodríguez, C. Diversidad alfa, beta y gamma: ¿cómo medimos las diferencias entre comunidades biológicas? NACC 2019, 26, 39–45. [Google Scholar]
- Malicky, H.; Chantaramongkol, P. The altitudinal distribution of Trichoptera species in Mae Klang catchment on Doi Inthanon, northern Thailand: Stream zonation and cool- and warm-adapted groups. Rev. Hydrobiol. Trop. 1993, 26, 279–291. [Google Scholar]
- Ruiz-García, A.; Herrera-Grao, A.F.; Ferreras-Romero, M. Distribution of Trichoptera communities in the Hozgarganta catchment (Los Alcornocales Natural Park, SW Spain). Int. Rev. Hydrobiol. 2006, 91, 71–85. [Google Scholar] [CrossRef]
- Kučinić, M.; Vučković, I.; Kutnjak, H.; Jelaska, L.S.; Marguš, D. Diversity, distribution, ecology and biogeography of caddisflies (Insecta: Trichoptera) in the Krka river (National Park “Krka”, Croatia). Zoosymposia 2011, 5, 255–268. [Google Scholar] [CrossRef] [Green Version]
- Wigger, F.W.; Schmidlin, L.; Nagel, P.; von Fumetti, S. Macroinvertebrate assemblages of natural spring along an altitudinal gradient in the Bernese Alps, Switzerland. Ann. Limnol. 2015, 51, 237–247. [Google Scholar] [CrossRef] [Green Version]
- Miserendino, M.L. Distribución altitudinal de especies de Trichoptera en un sistema fluvial en Patagonia. Ecol. Austral 1999, 9, 28–34. [Google Scholar]
- Henriques-Oliveira, A.L.; Nessimian, J.L. Aquatic macroinvertebrate diversity and composition in streams along an altitudinal gradient in Southeastern Brazil. Biota Neotrop. 2010, 10, 115–128. [Google Scholar] [CrossRef] [Green Version]
- Scheibler, E.E.; Claps, M.C.; Roig-Juñent, S.A. Temporal and altitudinal variations in benthic macroinvertebrates assemblages in an Andean river basin of Argentina. J. Limnol. 2014, 73, 76–92. [Google Scholar] [CrossRef] [Green Version]
- Henriques-Oliveira, A.L.; Nessimian, J.L.; Baptista, D. Diversity and composition of Trichoptera (Insecta) larvae assemblages in streams with different environmental conditions at Serra da Bocaina, Southeastern Brazil. Acta Limnol. Bras. 2015, 27, 394–410. [Google Scholar] [CrossRef]
- Castro, D.M.; Callisto, M.; Solar, R.R.C.; Macedo, D.R.; Fernades, G.W. Beta diversity of aquatic invertebrates increases along an altitudinal gradient in a Neotropical mountain. Biotropica 2019, 51, 399–411. [Google Scholar] [CrossRef]
- Janzen, D.H. Why mountain passes are higher in the tropics. Am. Nat. 1967, 101, 233–249. [Google Scholar] [CrossRef]
- Graham, C.H.; Carnaval, A.C.; Cadena, C.D.; Zamudio, K.R.; Roberts, T.E.; Parra, J.L.; McCain, C.M.; Bowie, R.C.K.; Moritz, C.; Baines, S.B.; et al. The origin and maintenance of montane diversity: Integrating evolutionary and ecological processes. Ecography 2014, 37, 711–719. [Google Scholar] [CrossRef]
- Polato, N.R.; Gill, B.A.; Shah, A.A.; Gray, M.M.; Casner, K.L.; Barthelet, A.; Messer, P.W.; Simmons, M.P.; Guayasamin, J.M.; Encalada, A.C.; et al. Narrow thermal tolerance and low dispersal drive higher speciation in tropical mountains. Proc. Natl. Acad. Sci. USA 2018, 115, 12471–12476. [Google Scholar] [CrossRef] [Green Version]
- Jacobsen, D. Contrasting patters in local and zonal family richness of stream invertebrates along an Andean altitudinal gradient. Freshw. Biol. 2004, 49, 1293–1305. [Google Scholar] [CrossRef]
- Jacobsen, D.; Schultz, R.; Encalada, A. Structure and diversity of stream invertebrate assemblages: The influence of temperature with altitude and latitude. Freshw. Biol. 1997, 38, 247–261. [Google Scholar] [CrossRef] [Green Version]
- Magliozzi, C.; Meyer, A.; Usseglio, P.; Robertson, A.; Grabowski, R.C. Investigating invertebrate biodiversity around large Wood: Taxonomic vs functional metrics. Aquat. Sci. 2020, 82, 69. [Google Scholar] [CrossRef]
- Rzedowski, J.; Palacios, R. El bosque de Engelhardtia (Oreomunnea) mexicana en la región de la Chinantla, Oaxaca, México, una reliquia del Cenozoico. Bol. Soc. Bot. México 1977, 36, 93–123. [Google Scholar] [CrossRef] [Green Version]
- Morrone, J.J. Biogeographic regionalization of the Sierra Madre del Sur province, Mexico. Rev. Mex. Biodivers. 2017, 88, 710–714. [Google Scholar] [CrossRef]
- Bueno-Soria, J.; Barba-Alvarez, R. New species of Plectropsyche Ross 1947 (Trichoptera: Hydropsychidae: Hydropsychinae). Zootaxa 2015, 4040, 421–432. [Google Scholar] [CrossRef]
- Trejo, I. Clima. In Biodiversidad de Oaxaca; García-Mendoza, A.J., Ordoñez, M.J., Briones-Salas, M., Eds.; Instituto de Biología, UNAM-Fondo Oaxaqueño para la Conservación de la Naturaleza-World Wildlife Fund: México City, México, 2004; Volume 1, pp. 67–85. [Google Scholar]
- Arriaga, L.; Espinoza-Rodríguez, J.M.; Aguilar-Zúñiga, C.; Martínez-Romero, E.; Gómez-Mendoza, L.; Loa, E. (Eds.) Regiones Terrestres Prioritarias de México; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: México City, México, 2000. [Google Scholar]
- Instituto Nacional de Estadística, Geografía e Informática (INEGI). Hidrología. 2020. Available online: https://www.inegi.org.mx/temas/hidrologia/ (accessed on 10 October 2021).
- García, E. Modificaciones al Sistema Climático de Köppen, 5th ed.; Instituto de Geografía-Universidad Nacional Autónoma de México: Ciudad de México, Mexico, 2004; pp. 19–74. [Google Scholar]
- Blahnik, R.J.; Holzenthal, R.W. Collection and curation of Trichoptera, with emphasis on pinned material. Nectopsyche Neotrop. Trichoptera Newsl. 2004, 1, 8–22. [Google Scholar]
- Triplehorn, C.A.; Johnson, N.F. Borror and DeLong’s Introduction to the Study of Insects, 7th ed.; Thomson Brooks/Cole: Belmont, MA, USA, 2005; pp. 745–775. [Google Scholar]
- Schmid, F. La famille des Xiphocentronides (Trichoptera: Annulipalpia). Mem. Entomol. Soc. Can. 1982, 121, 3–127. [Google Scholar] [CrossRef]
- StatSoft, Inc. Statistical User Guide; Complete Statistical System Statsoft: Oklahoma, OK, USA, 1995. [Google Scholar]
- Colwell, R.K. Estimates: Statistical Estimation of Species Richness and Shared Species from Samples (Version 9.1.0) User’s Guide and Application. 2013. Available online: http://viceroy.eeb.uconn.edu/estimates/EstimateSPages/EstSUsersGuide/EstimateSUsersGuide.htm (accessed on 1 September 2020).
- Jiménez-Valverde, A.; Hortal, J. Las curvas de acumulación de especies y la necesidad de evaluarla la calidad de los inventarios biológicos. Rev. Iber. Aracnol. 2003, 8, 151–161. [Google Scholar]
- González-Oreja, J.A.; De la Fuente-Díaz, A.A.; Hernández-Santín, L.D.; Buzo-Franco, D.; Bonache-Regidor, C. Evaluación de los estimadores no paramétricos de la riqueza de especies. Un ejemplo con las aves en áreas verdes de la ciudad de Puebla, México. Anim. Biodivers. Conserv. 2010, 33, 31–45. [Google Scholar] [CrossRef]
- Clarke, K.R.; Green, R.H. Statistical design and analysis for a ‘biological effects’ study. Mar. Ecol. Prog. Ser. 1988, 4, 213–226. [Google Scholar] [CrossRef]
- McAleece, N.; Gage, J.D.G.; Lambshead, P.J.D.; Paterson, G.L.J. BioDiversity Professional Statistics Analysis Software. Jointly Developed by the Scottish Association for Marine Science and the Natural History Museum London. 1997. Available online: https://www.sams.ac.uk/science/outputs/ (accessed on 26 November 2020).
- Jost, L. Entropy and Diversity. Oikos 2006, 113, 363–375. [Google Scholar] [CrossRef]
- Baselga, A.; Orme, C.D.L.; Villéger, S.; De Bortoli, J.; Leprieur, F. Betapart: Partitioning Beta Diversity into Turnover and Nestedness Components. R Package Version 1.4. 2017. Available online: http://CRAN.R-project.org/package=betapart (accessed on 26 August 2021).
- Chao, A.; Shen, T.J. Program SPADE (Species Prediction and Diversity Estimation). Program and User Guide. 2010. Available online: http://chao.stat.nthu.edu.tw/wordpress/software-download/ (accessed on 26 August 2021).
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatystical Analysis, 2nd ed.; Prentice-Hall: Upper Saddle River, NJ, USA, 1984; pp. 286–296. [Google Scholar]
- Bueno-Soria, J. Guía de Identificación Ilustrada de los Géneros de Larvas de Insectos del Orden Trichoptera de México, 1st ed.; Instituto de Biología-Universidad Nacional Autónoma de México: México City, Mexico, 2010; pp. 49–234. [Google Scholar]
- Razo-González, M. Caddisflies (Insecta: Trichoptera) from Santa Catarina Lachatao, Oaxaca, México: New species, new geographical records, and checklist, México. Zootaxa 2018, 438, 22–40. [Google Scholar] [CrossRef] [PubMed]
- Razo-González, M.; Castaño-Meneses, G.; Novelo-Gutiérrez, R.; Márquez, J. Preliminary evaluation of the nocturnal flight of caddisflies (Insecta: Trichoptera) in a temperate forest in Oaxaca, Mexico. Aquat. Insects 2020, 41, 339–355. [Google Scholar] [CrossRef]
- Razo-González, G.; Novelo-Gutiérrez, R.; Castaño-Meneses, G.; Márquez, J. Atopsyche Banks (Trichoptera: Hydrobiosidae) from Mexico: New species, redescription, and identification key. Stud. Neotrop. Fauna Environ. 2021. [Google Scholar] [CrossRef]
- Luis-Martínez, A.; Vargas-Fernández, I.; Llorente-Bousquets, J. Lepidopterofauna de Oaxaca I: Distribución y fenología de los Papilionoidea de la Sierra de Juárez; Publicaciones Especiales del Museo de Zoología: México City, Mexico, 1991; Volume 3, pp. 1–119. [Google Scholar]
- Rodríguez, A.; Castro-Castro, A.; Vargas-Amado, G.; Vargas-Ponce, O.; Zamora-Tavares, P.; González-Gallegos, J.; Carrillo-Reyes, P.; Anguiano-Constante, M.; Carrasco-Ortiz, M.; García-Martínez, M.; et al. Richness, geographic distribution patterns, and areas of endemism of selected angiosperm groups in Mexico. J. Syst. Evol. 2018, 56, 537–549. [Google Scholar] [CrossRef] [Green Version]
- Aragón-Parada, J.; Carrillo-Reyes, P.; Rodríguez, A.; Munguía-Lino, G. Diversidad y distribución del género Sedum (Crassulaceae) en la Sierra Madre del Sur, México. Rev. Mex. Biodivers. 2019, 90, 1–17. [Google Scholar] [CrossRef]
- Hernández-Rodríguez, E.; Aguirre-Hidalgo, V. Diversidad de musgos del bosque nublado de la Sierra de Juárez, Oaxaca, México. Acta Bot. Mex. 2020, 127, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Flint, O.S.; Holzenthal, R.W.; Harris, S.C. Catalog of the Neotropical Caddisflies (Insecta: Trichoptera); Ohio Biological Survey: Columbus, OH, USA, 1999; pp. 12–14. [Google Scholar]
- Kimura, G.; Inoue, E.; Hirabayashi, K. Seasonal abundance of adult caddisfly (Trichoptera) in the middle reaches of the Shinano River in Central Japan. In Proceedings of the 6th International Conference on Urban Pests, Budapest, Hungary, 13–16 July 2008; National Education Centre-Press Ltd.: Veszprem, Hungary, 2008; pp. 259–266. [Google Scholar]
- Thompson, R.; Townsend, C. A truce with neutral theory: Local deterministic factors, species traits and dispersal limitation together determine patterns of diversity in stream invertebrates. J. Anim. Ecol. 2006, 75, 476–484. [Google Scholar] [CrossRef]
- Clarke, A.; Mac Nally, R.; Bond, N.; Lake, P.S. Macroinvertebrate diversity in headwater streams: A review. Freshw. Biol. 2008, 53, 1707–1721. [Google Scholar] [CrossRef]
- Monaghan, M.T.; Robinson, C.T.; Spaak, P.; Ward, J.V. Macroinvertebrate diversity in fragmented Alpine streams: Implications for freshwater conservation. Aquat. Sci. 2005, 67, 454–464. [Google Scholar] [CrossRef]
- Danehy, R.J.; Chan, S.S.; Lester, G.T.; Langshaw, R.B.; Turner, T.R. Periphyton and macroinvertebrate assemblage structure in headwaters bordered by mature, thinned, and clearcut douglas-fir stands. For. Sci. 2007, 53, 294–307. [Google Scholar] [CrossRef]
- Frady, C.; Johnson, S.; Li, J. Stream macroinvertebrate community responses as legacies of forest harvest at the H.J. Andrews experimental forest, Oregon. For. Sci. 2007, 53, 281–293. [Google Scholar] [CrossRef]
- Moldenke, A.R.; Linden, C.V. Effects of clearcutting and riparian buffers on the yield of adult aquatic macroinvertebrates from headwater streams. For. Sci. 2007, 53, 308–319. [Google Scholar] [CrossRef]
- Jacobsen, D. Altitudinal changes in diversity of macroinvertebrates from small streams in the Ecuadorian Andes. Arch. Hydrobiol. 2003, 158, 145–167. [Google Scholar] [CrossRef]
- Zacarías-Eslava, Y.; del Castillo, R.F. Comunidades vegetales templadas de la Sierra Juárez Oaxaca: Pisos altitudinales y sus posibles implicaciones ante el cambio climático. Bol. Soc. Bot. México 2010, 87, 13–28. [Google Scholar] [CrossRef] [Green Version]
- Cava, M.B.; Corronca, J.A.; Echeverría, A.J. Diversidad alfa y beta de los artrópodos en diferentes ambientes del Parque Nacional Los Cardones, Salta (Argentina). Rev. Biol. Trop. 2013, 61, 1785–1798. [Google Scholar] [CrossRef]
- Hepp, L.U.; Melo, A.S. Dissimilarity of stream insect assemblages: Effects of multiple scales and spatial distances. Hydrobiologia 2013, 703, 239–246. [Google Scholar] [CrossRef]
- Curry, C.J.; Baird, D.J. Habitat type and dispersal ability influence spatial structuring of larval Odonata and Trichoptera assemblages. Freshw. Biol. 2015, 60, 2142–2155. [Google Scholar] [CrossRef]
- Rohde, K. Latitudinal gradients in species diversity: The search for the primary cause. Oikos 1992, 65, 514–527. [Google Scholar] [CrossRef] [Green Version]
- Mora, A.; Szivák, I.; Deák, C.; Boda, R.; Csabai, Z.; Sály, P.; Takács, P.; Erős, T.; Bíró, P. Environmental factors influencing distribution of EPT assemblages in streams of Lake Balaton’s catchment area, Hungary. Zoosymposia 2011, 5, 360–371. [Google Scholar] [CrossRef]
- De Mendoza, G.; Ventura, M.; Catalan, J. Environmental factors prevail over dispersal constraints in determining the distribution and assembly of Trichoptera species in mountain lakes. Ecol. Evol. 2015, 5, 2518–2532. [Google Scholar] [CrossRef] [Green Version]
- Morrone, J.J. Regionalización biogeográfica y evolución biótica de México: Encrucijada de la biodiversidad del Nuevo Mundo. Rev. Mex. Biodiver. 2019, 9, 1–68. [Google Scholar] [CrossRef] [Green Version]
- Razo-González, M.; Márquez, J.; Castaño-Meneses, G.; Novelo-Gutiérrez, R. La complejidad biogeográfica de la Sierra de Juárez, Oaxaca, México, revelada a través del análisis de parsimonia de endemismos de especies de tricópteros (Insecta: Trichoptera). Rev. Mex. Biodivers. 2021, 92, e923808. [Google Scholar] [CrossRef]
- Halffter, G. La zona de transición mexicana y la megadiversidad de México: Del marco histórico a la riqueza actual. Dugesiana 2017, 24, 77–89. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
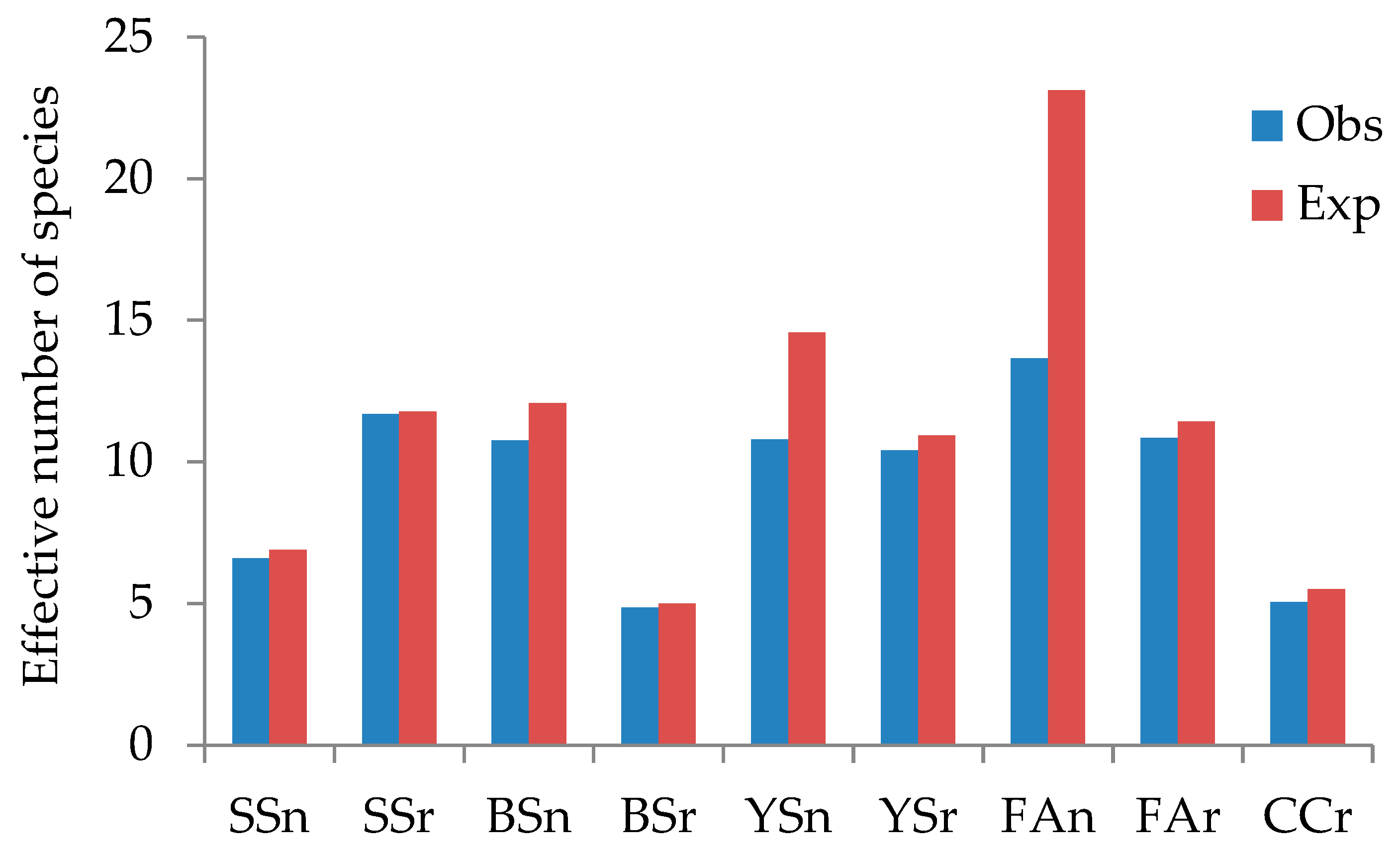{kind=link}
{kind=link}
| Stream | Geographic Location | Altitude (msnm) | MAP (mm) | MAT (°C) | Vegetation Type | Climate |
|---|---|---|---|---|---|---|
| Soyolapam (SS) | 17°41′47″ N 96°16′54″ W | 136 | 3000–3500 | 24.9 | Evergreen rainforest | Aw |
| Bobo (BS) | 17°39′23″ N 96°23′43″ W | 460 | 2500–3000 | 30 | Evergreen rainforest | Af |
| Yegu-Iag-Lagu (YS) | 17°24′18″ N 96°23′11″ W | 1320 | 1200–2300 | 21 | Cloud forest | (A)c(m) |
| Flor de Azahar (FA) | 17°13′21″ N 96°19′39″ W | 1967 | 800–1200 | 22.5 | Oak-pine forest | Cw |
| Cascadas Comaltepec (CC) | 17°35′15″ N 96°29′35″ W | 2432 | 2000–2500 | 9 | Pine forest | Cw |
| Stream | SS | BS | YS | FA | CC | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Season | n | r | n | r | n | r | n | r | n | r |
| T °C (±SE) | 16.74 (0.13) | 21.90 (0.24) | 15.73 (0.04) | 19.71 (0.14) | 10.31 (0.07) | 19.42 (0.33) | 9.43 (0.26) | 14.89 (0.37) | 2.26 (0.13) | 11.77 (0.11) |
| Minimum | 15.61 | 20.22 | 15.41 | 18.72 | 9.46 | 17.08 | 6.08 | 12.94 | 1.40 | 10.88 |
| Maximum | 18.18 | 29.24 | 16.39 | 23.62 | 11.66 | 24.77 | 14.19 | 20.89 | 3.49 | 14.65 |
| RH% (±SE) | 90.87 (0.15) | 88.60 (0.85) | 91.62 (0.10) | 91.49 (0.49) | 91.35 (0.09) | 92.13 (0.26) | 80.71 (0.95) | 93.19 (0.36) | 84.84 (0.87) | 94.45 (0.53) |
| Minimum | 88.33 | 68.74 | 89.87 | 80.87 | 89.65 | 87.21 | 69.25 | 81.89 | 72.32 | 87.33 |
| Maximum | 92.53 | 92.74 | 92.72 | 93.75 | 92.26 | 94.30 | 91.47 | 96.73 | 90.39 | 98.00 |
| Species/Assemblage | SS | BS | YS | FA | CC | Freq |
|---|---|---|---|---|---|---|
| Calamoceratidae | ||||||
| Phylloicus aeneus (Hagen, 1861) | * | * | * | 30 | ||
| Glossosomatidae | ||||||
| Culoptila jamapa Bueno-Soria and Santiago-Fragoso, 1996 | x * | x * | 40 | |||
| Culoptila pararusia Blahnik and Holzenthal, 2006 | x * | 20 | ||||
| Mortoniella falcicula Blahnik and Holzenthal, 2008 | * | 10 | ||||
| Mortoniella meralda (Mosely, 1954) | x * | 20 | ||||
| Mortoniella mexicana Blahnik and Holzenthal, 2008 | * | x | 20 | |||
| Protoptila bicornuta Flint, 1963 | * | 10 | ||||
| Protoptila ixtala Mosely, 1937 | x * | 20 | ||||
| Protoptila liqua Mosely, 1954 | x * | 20 | ||||
| Protoptila resolda Mosely, 1937 | * | 10 | ||||
| Protoptila rota Mosely, 1937 | * | 10 | ||||
| Protoptila spangleri Flint, 1967 | x * | 20 | ||||
| Helicopsychidae | ||||||
| Cochliopsyche vazquezae Flint, 1986 | * | 10 | ||||
| Helicopsyche borealis (Hagen, 1861) | x * | * | * | 40 | ||
| Helicopsyche dampfi Ross, 1956 | * | 10 | ||||
| Helicopsyche planata Ross, 1956 | x | x | * | 30 | ||
| Helicopsyche tuxtlensis Bueno-Soria, 1983 | x | * | * | 30 | ||
| Helicopsyche sp.+ | * | 10 | ||||
| Hydrobiosidae | ||||||
| Atopsyche bifurcata Razo-González and Novelo-Gutiérrez, 2021 | * | 10 | ||||
| Atopsyche calopta Ross and King, 1952 | x | x | 20 | |||
| Atopsyche dampfi Ross and King, 1952 | x | x | x * | x * | 60 | |
| Atopsyche erigia Ross, 1947 | * | 10 | ||||
| Atopsyche huenga Flint, 1974 | * | 10 | ||||
| Atopsyche japoda Ross and King, 1952 | x | x | 20 | |||
| Atopsyche jujmi Razo-González and Novelo-Gutiérrez, 2021 | * | 10 | ||||
| Atopsyche majada Ross, 1947 | x * | * | 30 | |||
| Atopsyche pilcomayo Schmid, 1989 | * | 10 | ||||
| Hydropsychidae | ||||||
| Cheumatopsyche gelita Denning, 1952 | x | 10 | ||||
| Diplectrona chiapensis Flint, 1967 | x | x | x * | * | 50 | |
| Hydropsyche toschiae (Denning, 1965) | x | x * | x * | x * | 70 | |
| Hydropsyche sp. 1+ | x * | x * | * | 50 | ||
| Hydropsyche sp. 2+ | x * | * | 30 | |||
| Leptonema albovirens (Walker, 1852) | x * | 20 | ||||
| Leptonema championi Mosely, 1933 | x | 10 | ||||
| Leptonema crassum Ulmer, 1905 | * | 10 | ||||
| Leptonema pinotepa Bueno-Soria et al., 2001 | x * | 20 | ||||
| Leptonema simulans mayanum Flint et al., 1987 | x * | 20 | ||||
| Plectropsyche hoogstraali Ross, 1947 | x * | x * | 40 | |||
| Smicridea bulara Flint and Denning, 1989 | * | 10 | ||||
| Smicridea dampfi Flint, 1974 | * | 10 | ||||
| Smicridea dithyra Flint, 1974 | x * | x * | * | 50 | ||
| Smicridea lachanha Bueno-Soria and Hamilton, 1986 | x * | 20 | ||||
| Smicridea radula Flint, 1974 | x * | x * | * | 50 | ||
| Smicridea signata (Banks, 1903) | * | 10 | ||||
| Hydroptilidae | ||||||
| Anchitrichia spangleri Flint, 1970 | x * | 20 | ||||
| Costatrichia lodora Mosely, 1937 | x * | 20 | ||||
| Hydroptila denza Ross, 1948 | * | 10 | ||||
| Hydroptila mexicana Mosely, 1937 | x * | 20 | ||||
| Hydroptila misolha Bueno-Soria, 1984 | * | 10 | ||||
| Ithytrichia mexicana Harris and Contreras-Ramos, 1989 | * | * | 20 | |||
| Leucotrichia imitator Flint, 1970 | x | 10 | ||||
| Mayatrichia rualda Mosely, 1937 | x * | * | 30 | |||
| Metrichia cf. circulatrix Bueno-Soria 2002 | * | 10 | ||||
| Metrichia crenula Bueno-Soria 2002 | x | 10 | ||||
| Metrichia yavesia Bueno-Soria, 2002 | x * | 20 | ||||
| Metrichia sp. 1+ | * | 10 | ||||
| Metrichia sp. 2+ | * | 10 | ||||
| Neotrichia exicoma (Mosely, 1937) | * | 10 | ||||
| Neotrichia xicana (Mosely, 1937) | * | x | 20 | |||
| Neotrichia cf. maria Bueno and Hamilton, 1986 | x | 10 | ||||
| Ochrotrichia pacifica Flint, 1972 | x * | 20 | ||||
| Ochrotrichia yavesia Bueno and Holzenthal, 2004 | * | x | 20 | |||
| Ochrotrichia yetla Bueno-Soria, 2009 | * | x * | 30 | |||
| Ochrotrichia sp. 1+ | * | 10 | ||||
| Ochrotrichia sp. 2+ | * | * | 20 | |||
| Oxyethira azteca (Mosely, 1937) | x | x | 20 | |||
| Oxyethira tica Holzenthal and Harris, 1992 | * | 10 | ||||
| Rhyacopsyche mexicana (Flint, 1967) | * | 10 | ||||
| Rhyacopsyche sp.+ | * | 10 | ||||
| Zumatrichia filosa Mosely, 1937 | * | 10 | ||||
| Zumatrichia multisetosa Flint, 1970 | * | 10 | ||||
| Zumatrichia sp.+ | * | x | 20 | |||
| Lepidostomatidae | ||||||
| Lepidostoma bakeri Flint, 1965 | x * | 20 | ||||
| Lepidostoma frontale (Banks, 1901) | x | x * | 30 | |||
| Lepidostoma zapoteca Razo-González, 2018 | x | 10 | ||||
| Lepidostoma sp. 1+ | x * | x | 30 | |||
| Lepidostoma sp. 2+ | x * | 20 | ||||
| Lepidostoma sp. 3+ | * | * | 20 | |||
| Leptoceridae | ||||||
| Nectopsyche dorsalis (Banks, 1901) | x * | 20 | ||||
| Nectopsyche gemmoides Flint, 1981 | x * | x * | * | 50 | ||
| Nectopsyche pavida (Hagen, 1861) | x | 10 | ||||
| Oecetis disjuncta (Banks, 1920) | x * | x * | 40 | |||
| Oecetis metlacensis Bueno-Soria, 1981 | x * | x * | * | * | 60 | |
| Oecetis silviae Bueno-Soria, 1981 | * | * | 20 | |||
| Oecetis sordida Blahnik and Holzenthal, 2014 | x * | 20 | ||||
| Oecetis sp.+ | * | 10 | ||||
| Triaenodes sp.+ | * | 10 | ||||
| Odontoceridae | ||||||
| Marilia flexuosa Ulmer, 1905 | * | 10 | ||||
| Philopotamidae | ||||||
| Chimarra acuta Ross, 1959 | * | 10 | ||||
| Chimarra barrettae (Banks, 1900) | x * | * | 30 | |||
| Chimarra bicolor (Banks, 1901) | * | * | * | 30 | ||
| Chimarra dentosa Ross, 1948 | * | 10 | ||||
| Chimarra laguna Ross, 1951 | * | 10 | ||||
| Chimarra mexicana (Banks, 1900) | x | x * | * | 40 | ||
| Chimarra oaxaca Blahnik, 1998 | * | 10 | ||||
| Chimarra ortiziana Flint, 1967 | * | 10 | ||||
| Chimarra pelaezi Bueno-Soria, 1985 | * | x * | * | 40 | ||
| Chimarra ridleyi (Denning, 1941) | * | 10 | ||||
| Chimarra setosa Ross, 1959 | * | 10 | ||||
| Wormaldia barbai Muñoz-Quesada and Holzenthal, 2015 | * | x * | * | * | 50 | |
| Wormaldia noveloi Razo-González, 2018 | * | * | 20 | |||
| Wormaldia planae Ross and king, 1956 | x | 10 | ||||
| Wormaldia tarasca Bueno-Soria and Holzenthal, 1986 | * | 10 | ||||
| Polycentropodidae | ||||||
| Cernotina calcea Ross, 1938 | * | 10 | ||||
| Cernotina chiapaneca Bueno-Soria, 2010 | * | 10 | ||||
| Polycentropus aliciae Barba-Álvarez and Bueno-Soria, 2005 | * | 10 | ||||
| Polycentropus aztecus Flint, 1967 | x * | 20 | ||||
| Polycentropus casicus Denning and Sykora, 1966 | * | 10 | ||||
| Polycentropus mayanus Flint, 1981 | x * | * | 30 | |||
| Polycentropus mexicanus (Banks, 1901) | x * | 20 | ||||
| Polycentropus veracruzensis Flint, 1981 | * | * | 20 | |||
| Polyplectropus carolae Bueno-Soria, 1990 | * | 10 | ||||
| Polyplectropus charlesi (Ross, 1941) | * | 10 | ||||
| Psychomyiidae | ||||||
| Tinodes sp. | x * | 20 | ||||
| Xiphocentronidae | ||||||
| Cnodocentron sp.+ | * | 10 | ||||
| Xiphocentron aureum Flint, 1967 | * | 10 |
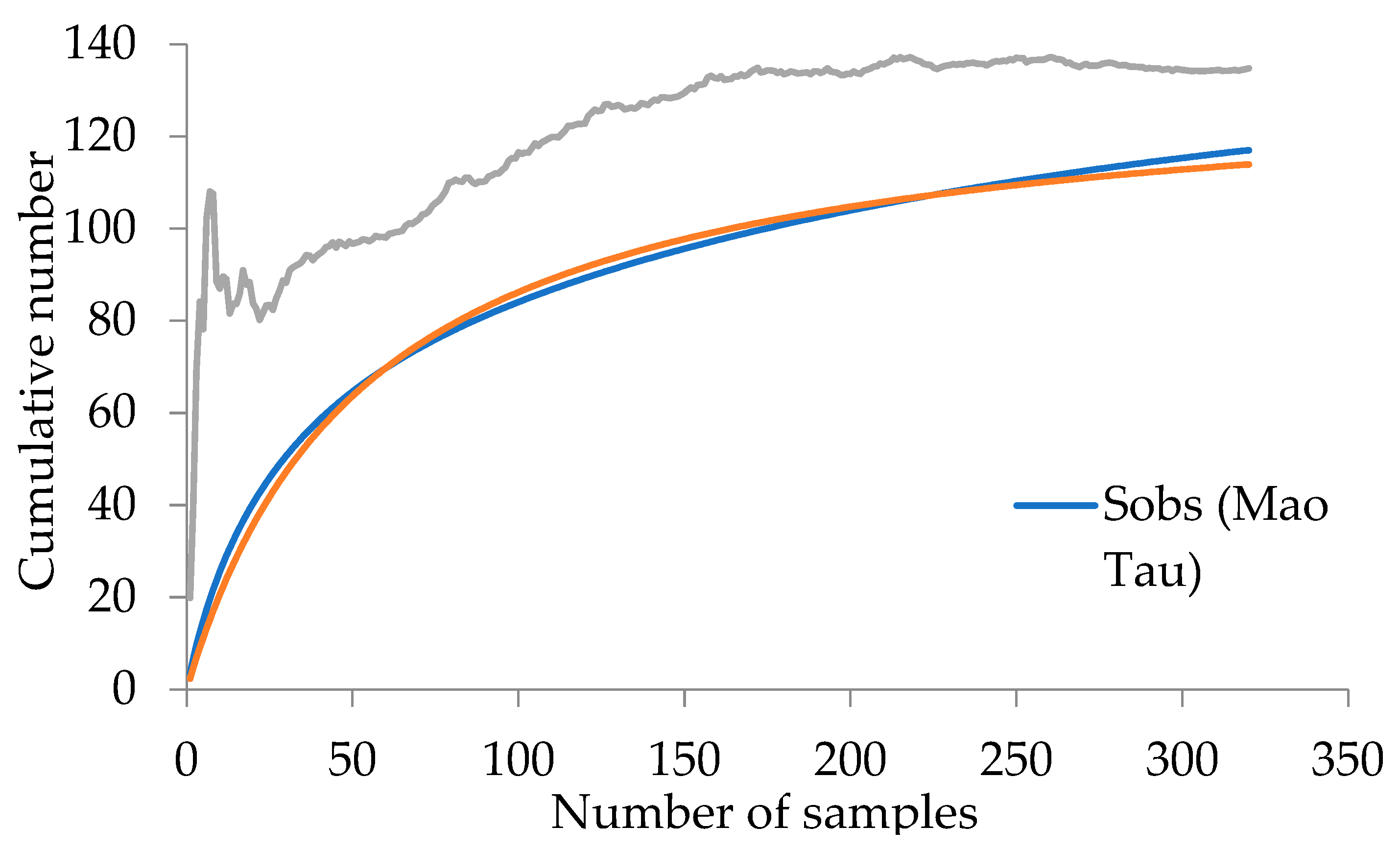
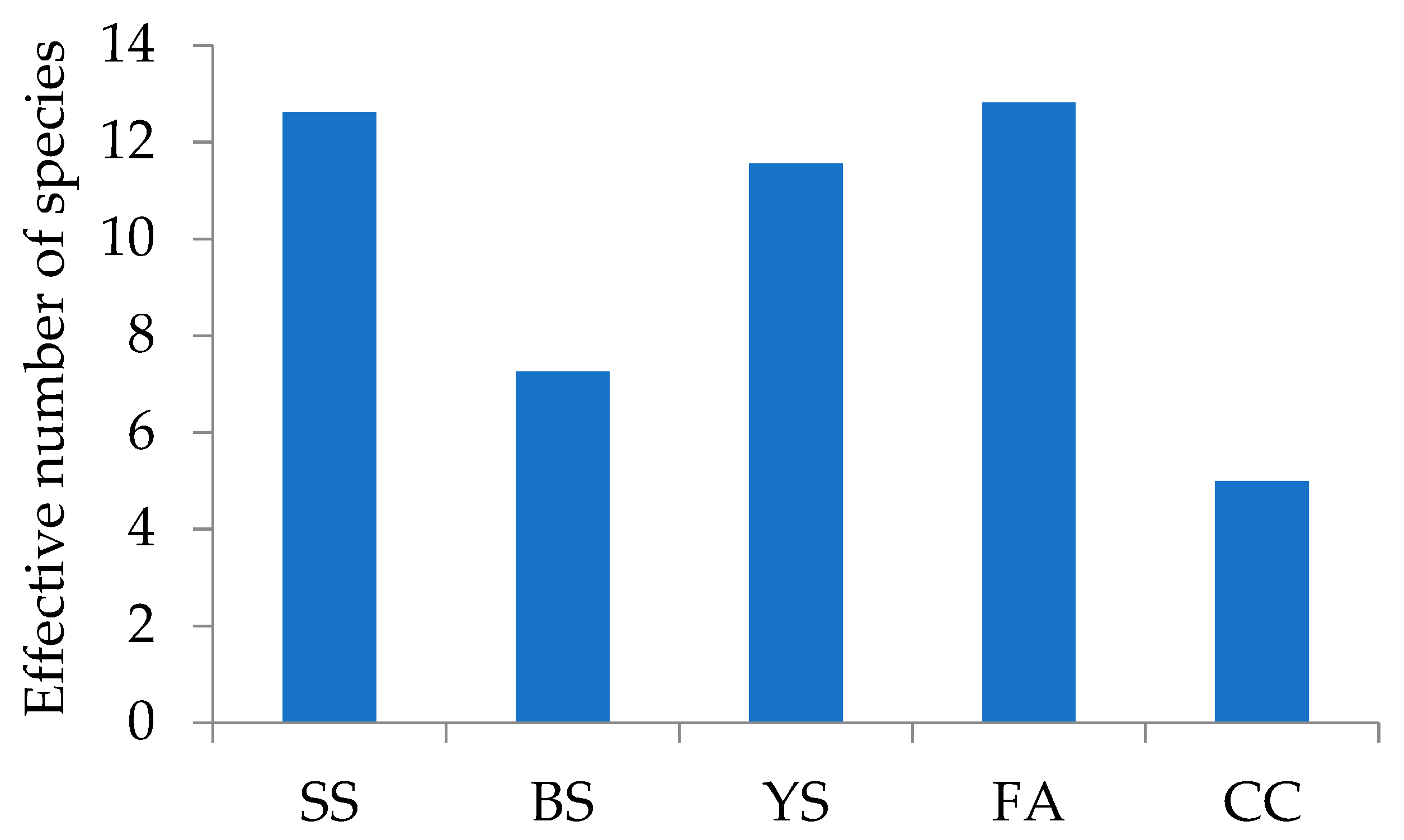
| Richness | True Diversity | ||||||
|---|---|---|---|---|---|---|---|
| Site | ♂ Abundance | Obs. | Clench | Chao2 | Excl. | Obs. | Est. |
| SSn | 504 | 34 | 47 (72.34) | 39 (87.18) | 13(11.02) | 6.60 | 6.90 (95.65) |
| BSn | 191 | 27 | 40 (67.50) | 69 (39.13) | 4(3.39) | 10.76 | 12.08 (89.07) |
| YSn | 25 | 12 | 24 (50.00) | 13 (92.31) | 1(0.85) | 10.80 | 14.57 (74.12) |
| Fan | 29 | 16 | 35 (45.71) | 30 (53.33) | 6(5.08) | 13.65 | 23.13 (59.01) |
| CCn | 1 | 1 | - | - | 1(0.85) | 1 | - |
| SSr | 5038 | 61 | 67 (91.04) | 66 (92.42) | 41(34.75) | 11.68 | 11.77 (99.24) |
| BSr | 1138 | 32 | 38 (94.12) | 62 (51.61) | 6(5.08) | 4.86 | 4.99 (97.39) |
| YSr | 527 | 37 | 55 (67.27) | 55 (67.27) | 8(6.78) | 10.41 | 10.94 (95.16) |
| Far | 283 | 25 | 30 (83.33) | 29 (86.21) | 10(8.47) | 10.85 | 11.43 (94.93) |
| CCr | 44 | 8 | 11 (72.73) | 8 (100) | 5(4.24) | 5.05 | 5.50 (91.82) |
| SJ | 7402 | 118 | 133 (88.72) | 134 (88.06) | 77(65.25) | 17.22 | 19.45 (88.53) |
| Clench | ||||||||
|---|---|---|---|---|---|---|---|---|
| Stream | S | n | a | b | a/b | R2 | Slope | % |
| SSn | 34 | 32 | 3.76 | 0.08 | 47.24 | 0.99 | 0.30 | 71.98 |
| BSn | 27 | 32 | 2.42 | 0.06 | 40.05 | 0.99 | 0.28 | 67.42 |
| YSn | 12 | 32 | 0.76 | 00.03 | 24.25 | 0.99 | 0.19 | 49.49 |
| FAn | 16 | 32 | 0.89 | 0.02 | 35.95 | 0.99 | 0.28 | 44.50 |
| CCn | 1 | 16 | - | - | - | - | - | - |
| SSr | 61 | 32 | 16.54 | 0.25 | 67.12 | 0.99 | 0.21 | 90.89 |
| BSr | 32 | 32 | 4.79 | 0.12 | 38.93 | 0.99 | 0.20 | 82.21 |
| YSr | 37 | 32 | 3.27 | 0.06 | 55.03 | 0.99 | 0.39 | 67.23 |
| FAr | 25 | 32 | 3.72 | 0.12 | 30.86 | 0.99 | 0.16 | 81.01 |
| CCr | 8 | 32 | 0.98 | 0.09 | 11.19 | 0.99 | 0.07 | 71.50 |
| SJtotal | 118 | 320 | 2.44 | 0.02 | 133.41 | 0.99 | 0.97 | 87.70 |
| BS | YS | FA | CC | Mean | ||
|---|---|---|---|---|---|---|
| SS | βcc | 0.72 | 0.85 | 0.92 | 1.00 | 0.87 |
| β-3 | 0.59 | 0.78 | 0.86 | 1.00 | 0.81 | |
| βrich | 0.13 | 0.07 | 0.07 | 0.00 | 0.07 | |
| BS | βcc | 0.65 | 0.87 | 0.98 | 0.83 | |
| β-3 | 0.63 | 0.83 | 0.93 | 0.80 | ||
| βrich | 0.01 | 0.04 | 0.05 | 0.03 | ||
| YS | βcc | 0.72 | 0.93 | 0.83 | ||
| β-3 | 0.63 | 0.77 | 0.70 | |||
| βrich | 0.09 | 0.17 | 0.13 | |||
| FA | βcc | 0.94 | 0.94 | |||
| β-3 | 0.86 | 0.86 | ||||
| βrich | 0.08 | 0.08 |
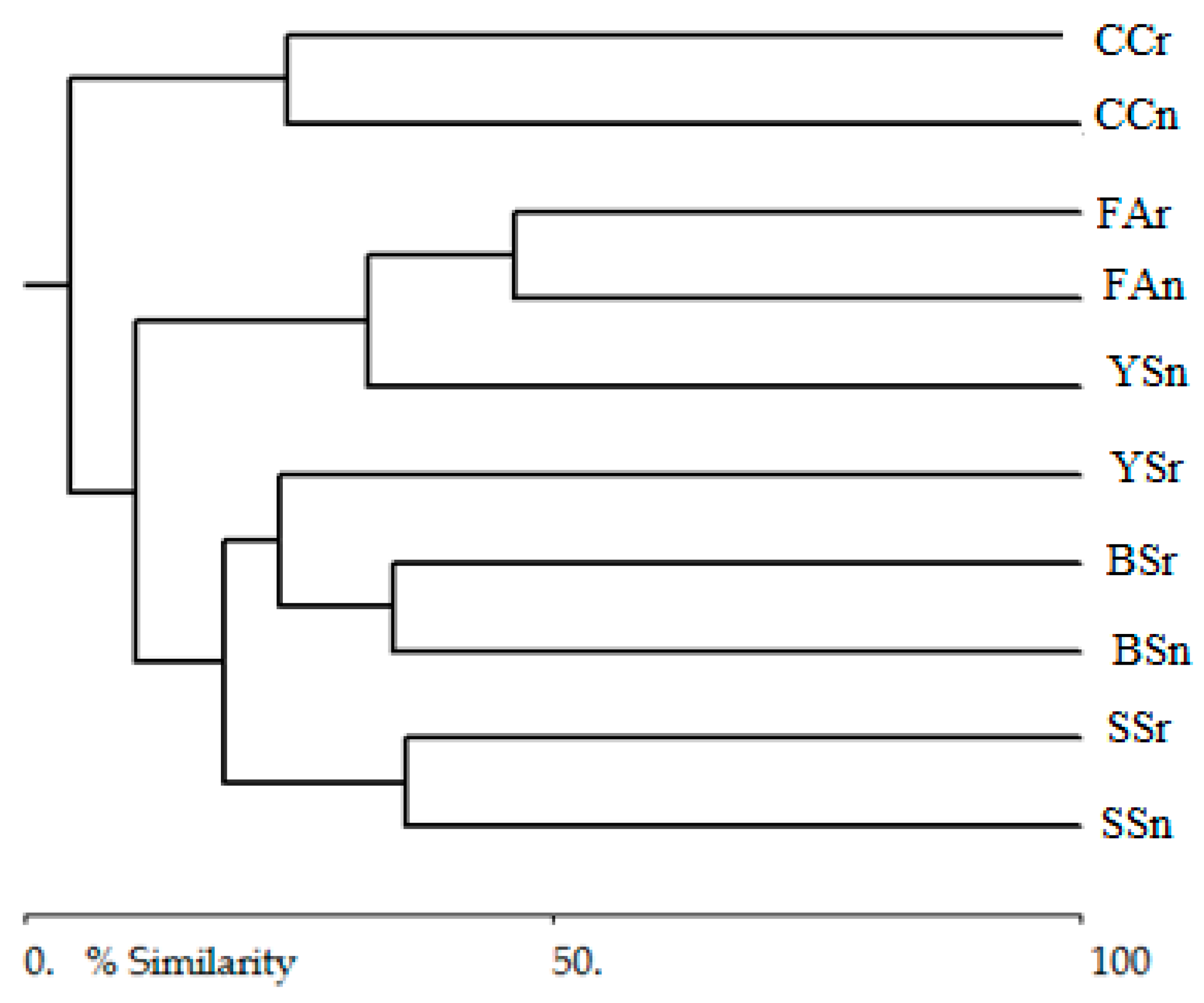
| SSn | BSn | YSn | FAn | CCn | SSr | BSr | YSr | FAr | CCr | |
|---|---|---|---|---|---|---|---|---|---|---|
| SSn | 0.00 | 0.93 | 0.99 | 0.99 | 1.00 | 0.72 | 0.93 | 0.98 | 0.99 | 1.00 |
| BSn | 0.00 | 0.93 | 0.89 | 1.00 | 0.91 | 0.93 | 0.78 | 0.87 | 0.99 | |
| YSn | 0.00 | 0.32 | 1.00 | 1.00 | 0.99 | 0.67 | 0.74 | 0.99 | ||
| FAn | 0.00 | 1.00 | 1.00 | 0.98 | 0.61 | 0.52 | 0.95 | |||
| CCn | 0.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | ||||
| SSr | 0.00 | 0.41 | 0.88 | 0.99 | 1.00 | |||||
| BSr | 0.00 | 0.90 | 0.97 | 1.00 | ||||||
| YSr | 0.00 | 0.22 | 0.99 | |||||||
| FAr | 0.00 | 0.99 | ||||||||
| CCr | 0.00 |
| Total Richness | Total Abundance | Nectopsyche gemmoides | Smicridea radula | |
|---|---|---|---|---|
| Season | 256.40 * | 147.65 * | 37.39 * | 70.74 * |
| Site | 127.55 * | 74.55 * | 132.08 * | 38.84 * |
| Interaction | 16.79 * | 24.11 * | 33.26 * | 27.97 * |
| Total Richness | Total Abundance | Nectopsyche gemmoides | Smicridea radula | |
|---|---|---|---|---|
| Season | r2 = 0.47 * gl = 2.32 p = 0.00 | r2 = 0.42 * gl = 2.32 p = 0.00 | r2 = 0.19 * gl = 2.32 p = 0.00 | r2 = 0.33 * gl = 2.32 p = 0.00 |
| Site | r2 = −0.43 * gl = 2.32 p = 0.00 | r2 = −0.43 * gl = 2.32 p = 0.00 | r2 = −0.52 * gl = 2.32 p = 0.00 | r2 = −0.44 * gl = 2.32 p = 0.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Razo-González, M.; Novelo-Gutiérrez, R.; Castaño-Meneses, G.; Márquez, J. Diversity and Composition of Caddisflies (Insecta: Trichoptera) along an Elevation Gradient in Southeastern Mexico. Diversity 2023, 15, 110. https://doi.org/10.3390/d15010110
Razo-González M, Novelo-Gutiérrez R, Castaño-Meneses G, Márquez J. Diversity and Composition of Caddisflies (Insecta: Trichoptera) along an Elevation Gradient in Southeastern Mexico. Diversity. 2023; 15(1):110. https://doi.org/10.3390/d15010110
Chicago/Turabian StyleRazo-González, María, Rodolfo Novelo-Gutiérrez, Gabriela Castaño-Meneses, and Juan Márquez. 2023. "Diversity and Composition of Caddisflies (Insecta: Trichoptera) along an Elevation Gradient in Southeastern Mexico" Diversity 15, no. 1: 110. https://doi.org/10.3390/d15010110