
Application of Phytoplankton Taxonomic α-Diversity Indices to Assess Trophic States in Barrier Lake: A Case of Jingpo Lake
 and
and
Abstract
:1. Introduction
2. Materials and Methods
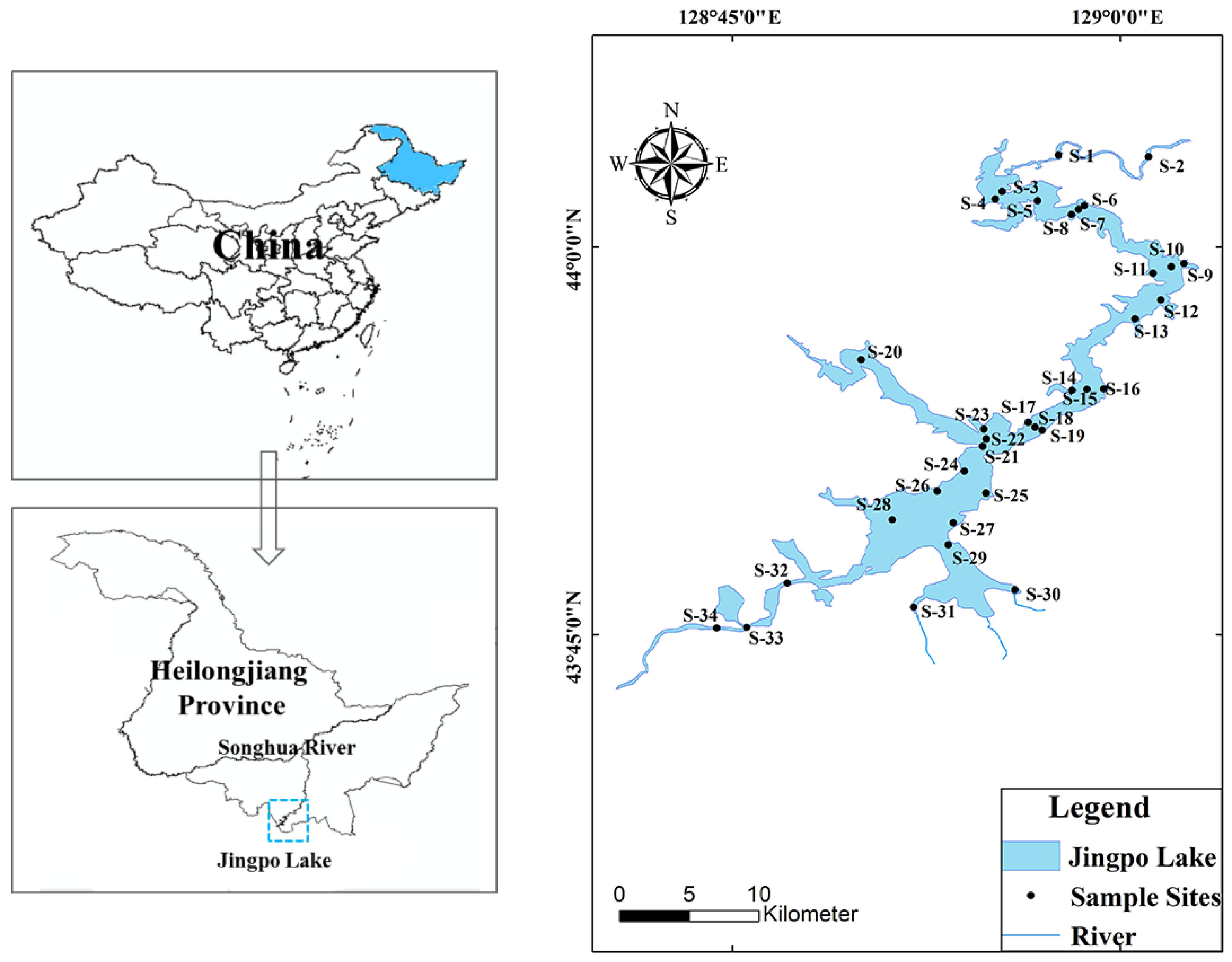
2.1. Study Area, Sample Collection, and Laboratory Analyses
2.2. Taxonomic α-Diversity Indices
- Shannon-Wiener index:
- Margalef index:
- Pielou evenness index:where ni and N are the numbers of individuals of species i and the total number of individuals of all species within site; S is the total species in the sample; fi is the occurrence frequency of the species i; Pi = ni/N. Evaluation standard: Margalef > 5 cleanness, >4 oligotrophic, >3 β-mesotrophic, <3 α-mesotrophic; Shannon-Wiener index > 3 oligotrophic, 2–3 β-mesotrophic, 1–2 α-mesotrophic, 0–1 eutrophic.
2.3. Statistical Analysis
2.4. Comprehensive Trophic Level Index (TLI)
3. Results
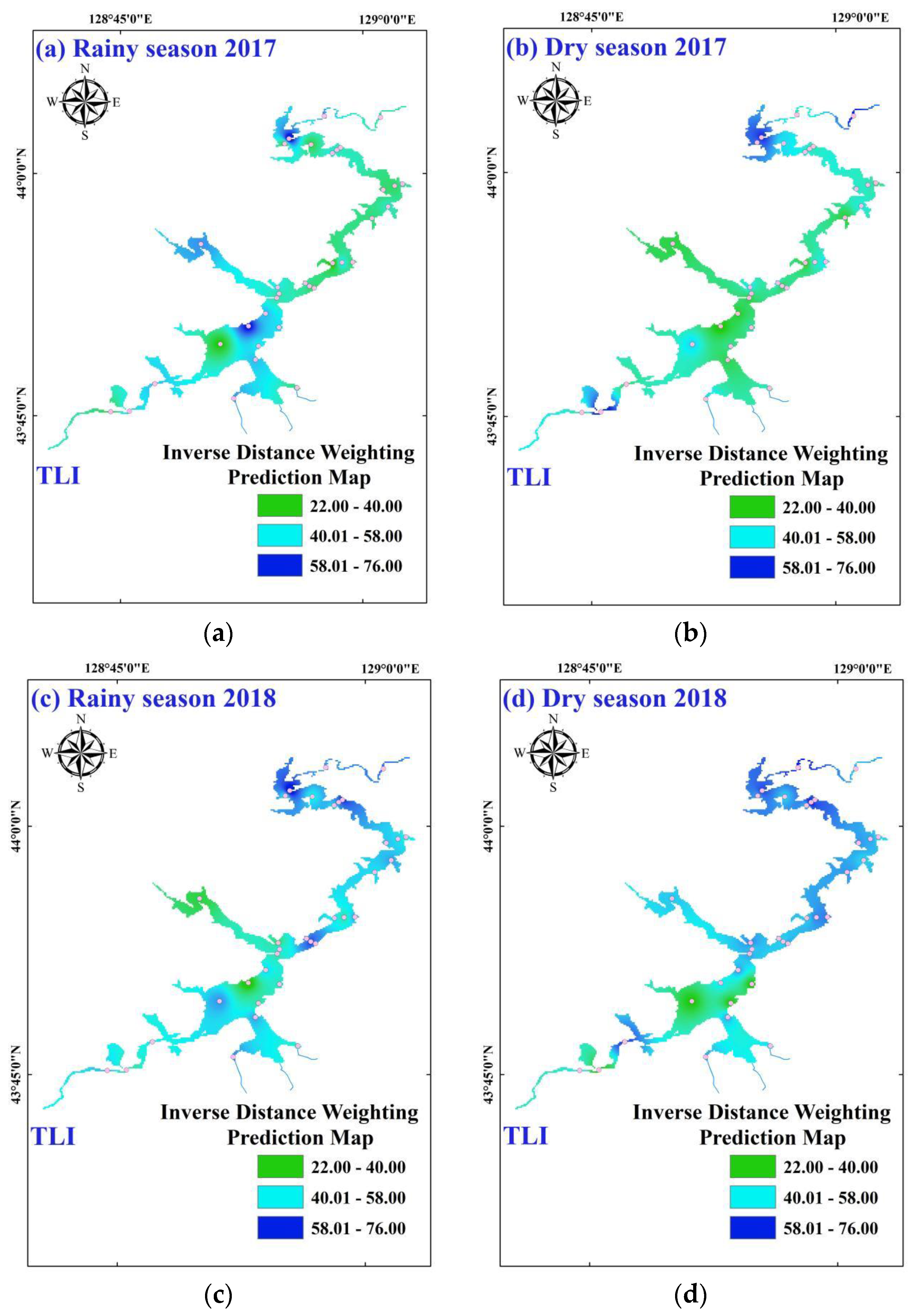
3.1. Environmental Characteristics and Trophic States
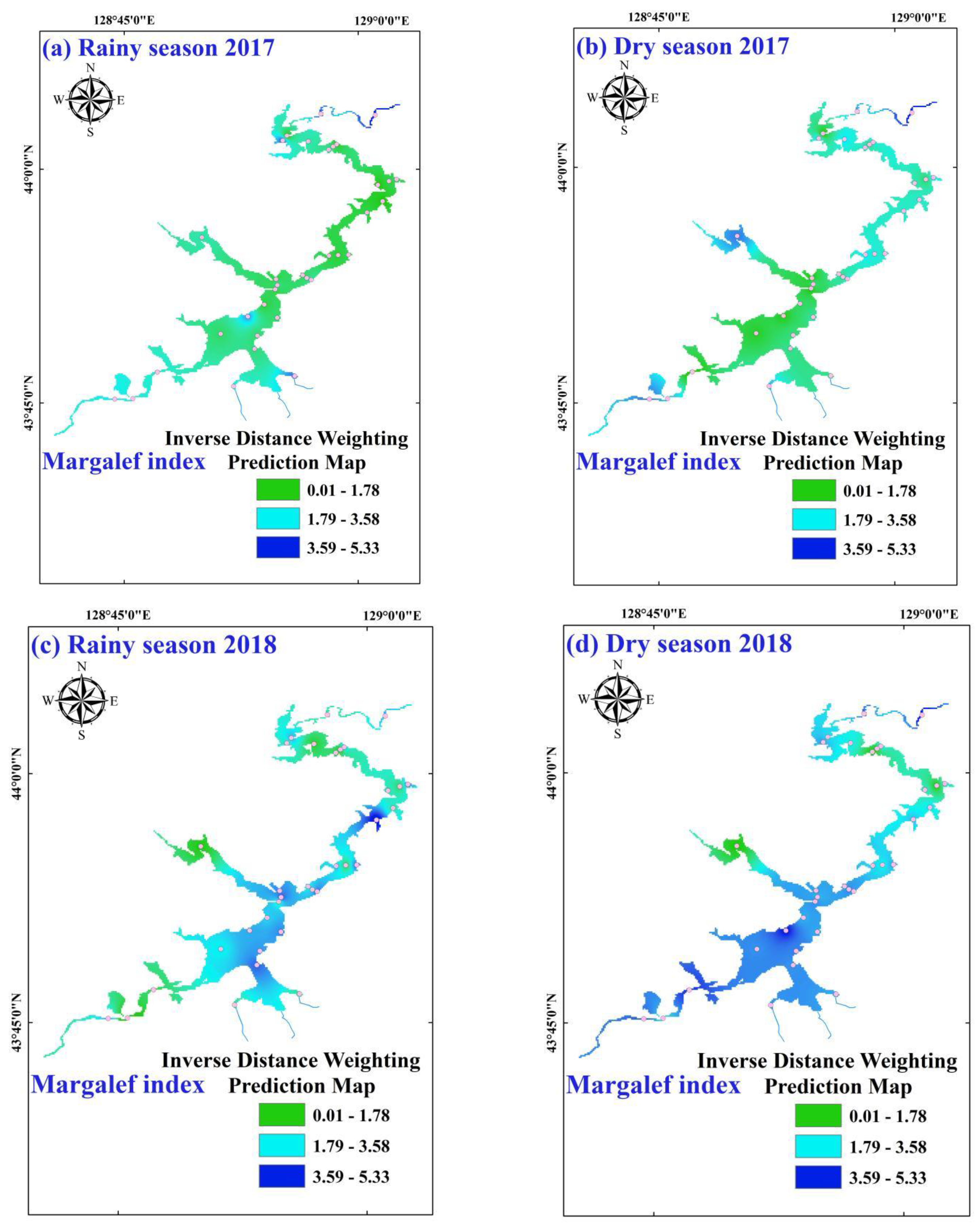
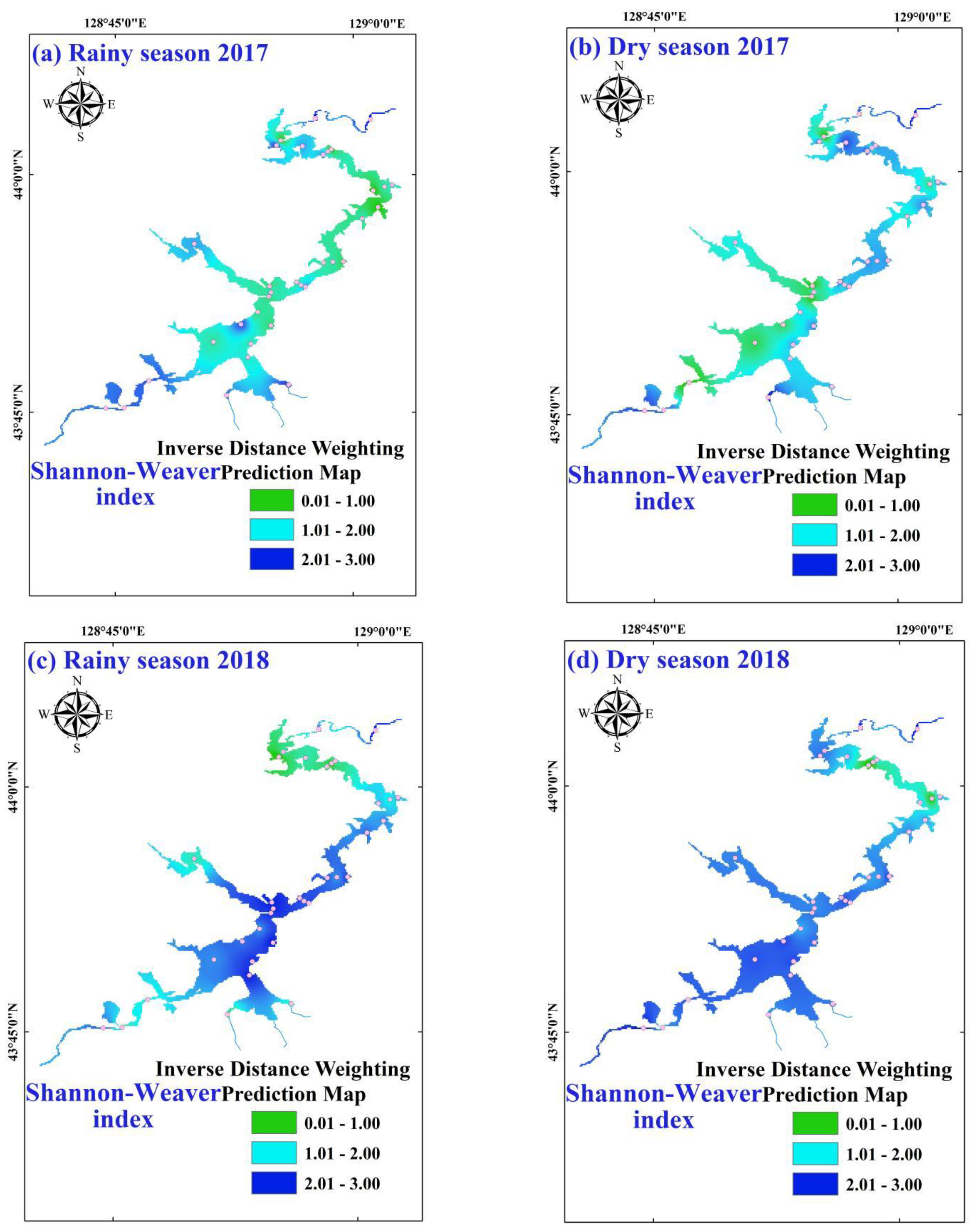
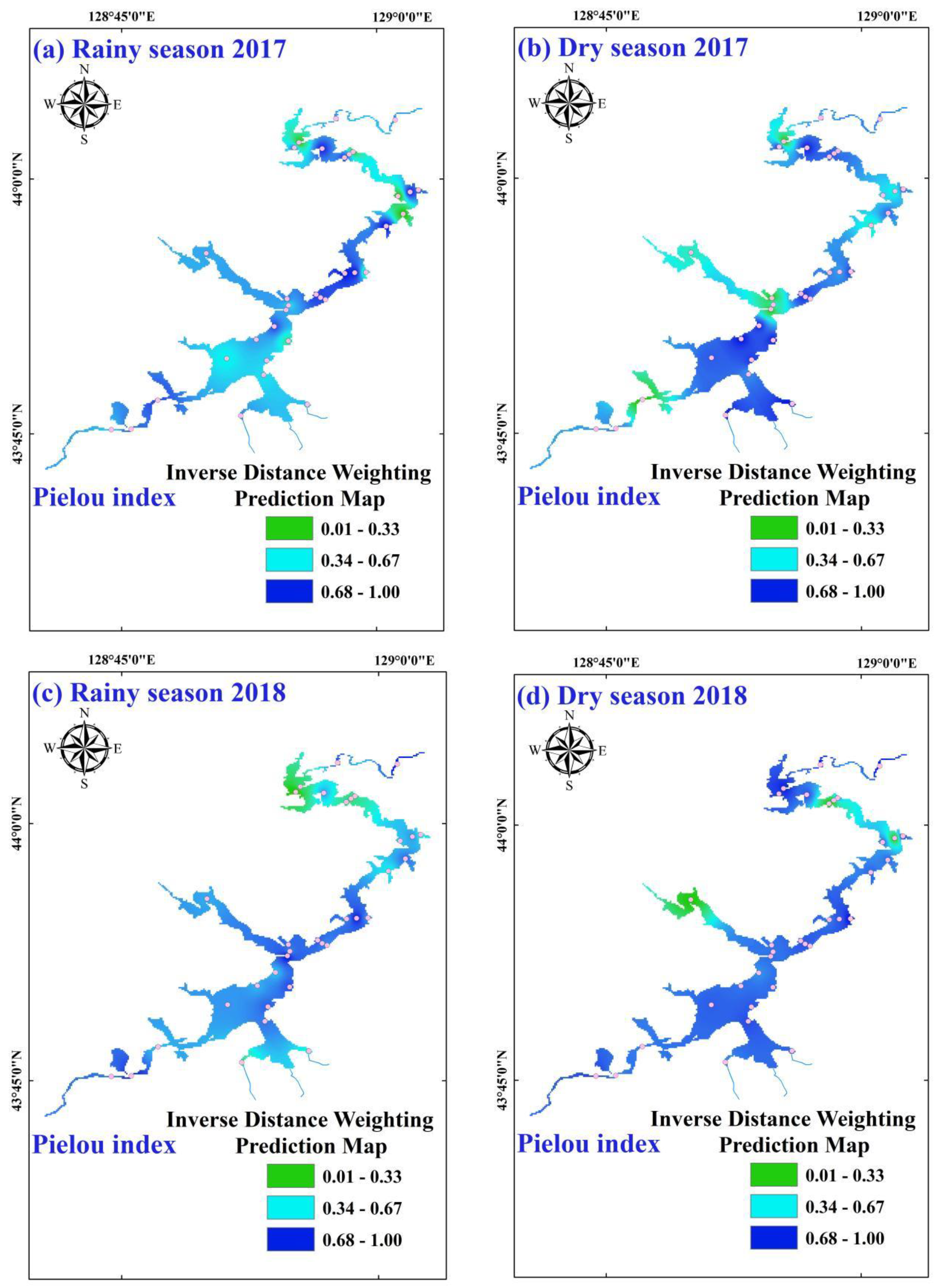
3.2. Temporal-Spatial Variations of Phytoplankton Diversity Indices
3.3. The Correlation between Phytoplankton Taxonomic α-Diversity Indices and TLI
4. Discussion
4.1. Effects of Environmental Factors on Phytoplankton Community Structure
4.2. Response of Phytoplankton α-Diversity Indices to Nutrient State Changes in Different Hydrological Periods
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Amorim, C.A.; Valenca, C.R.; Moura-Falcao, R.H.; Moura, A.d.N. Seasonal variations of morpho-functional phytoplankton groups influence the top-down control of a cladoceran in a tropical hypereutrophic lake. Aquat. Ecol. 2019, 53, 453–464. [Google Scholar] [CrossRef]
- Bolgovics, A.; B-Beres, V.; Varbiro, G.; Krasznai-K, E.A.; Acs, E.; Kiss, K.T.; Borics, G. Groups of small lakes maintain larger microalgal diversity than large ones. Sci. Total Environ. 2019, 678, 162–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacquemin, C.; Bertrand, C.; Franquet, E.; Mounier, S.; Misson, B.; Oursel, B.; Cavalli, L. Effects of catchment area and nutrient deposition regime on phytoplankton functionality in alpine lakes. Sci. Total Environ. 2019, 674, 114–127. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Chen, S.; Liu, H.; Guan, Y. Blue-green algae enhanced performance of diatom-based multimetric index on de fining lake condition under high level of human disturbance. Sci. Total Environ. 2020, 730. [Google Scholar] [CrossRef] [PubMed]
- Demir, A.N.; Fakioglu, O.; Dural, B. Phytoplankton functional groups provide a quality assessment method by the Q assemblage index in Lake Mogan (Turkey). Turk. J. Bot. 2014, 38, 169–179. [Google Scholar] [CrossRef]
- Korneva, L.G.; Solovyeva, V.V. Dynamics of Morphofuncitonal Groups of Phytoplankton in the Rybinsk Resrevoir and Assessment of the Resevoir Water Quality by the Community Index. Water Resour. 2021, 48, 65–72. [Google Scholar] [CrossRef]
- Liu, L.; Zhu, Y.; Li, B.; Chen, X. Assessment of phytoplankton biological integrity in Jiangsu section of Yangtze River based on MBFG. Water Res. Protein 2020, 36, 13–20. [Google Scholar]
- Ward, B.B.; Van Oostende, N. Phytoplankton assemblage during the North Atlantic spring bloom assessed from functional gene analysis. J. Plankton Res. 2016, 38, 1135–1150. [Google Scholar] [CrossRef] [Green Version]
- Marra, R.C.; Algarte, V.M.; Ludwig, T.A.V.; Padial, A.A. Diatom diversity at multiple scales in urban reservoirs in Southern Brazil reveals the likely role of trophic state. Limnologica 2018, 70, 49–57. [Google Scholar] [CrossRef]
- Behl, S.; Donval, A.; Stibor, H. The relative importance of species diversity and functional group diversity on carbon uptake in phytoplankton communities. Limnol. Oceanogr. 2011, 56, 683–694. [Google Scholar] [CrossRef]
- Beisner, B.E.; Longhi, M.L. Spatial overlap in lake phytoplankton: Relations with environmental factors and consequences for diversity. Limnol. Oceanogr. 2013, 58, 1419–1430. [Google Scholar] [CrossRef]
- Terseleer, N.; Bruggeman, J.; Lancelot, C.; Gypens, N. Trait-based representation of diatom functional diversity in a plankton functional type model of the eutrophied southern North Sea. Limnol. Oceanogr. 2014, 59, 1958–1972. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, L.C.; Simoes, N.R.; Bovo-Scomparin, V.M.; Jati, S.; Santana, N.F.; Roberto, M.C.; Train, S. Phytoplankton alpha diversity as an indicator of environmental changes in a neotropical floodplain. Ecol. Indic. 2015, 48, 334–341. [Google Scholar] [CrossRef]
- Schuster, K.F.; Tremarin, P.I.; de Souza-Franco, G.M. Alpha and beta diversity of phytoplankton in two subtropical eutrophic streams in southern Brazil. Acta Bot. Bras. 2015, 29, 597–607. [Google Scholar] [CrossRef] [Green Version]
- Nabout, J.C.; Nogueira, I.d.S.; de Oliveira, L.G.; Morais, R.R. Phytoplankton diversity (alpha, beta, and gamma) from the Araguaia River tropical floodplain lakes (central Brazil). Hydrobiologia 2007, 575, 455–461. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Aber, J.D.; Howarth, R.W.; Likens, G.E.; Matson, P.A.; Schindler, D.W.; Schlesinger, W.H.; Tilman, D. Human alteration of the global nitrogen cycle: Sources and consequences. Ecol. Appl. 1997, 7, 737–750. [Google Scholar] [CrossRef] [Green Version]
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.-I.; Knowler, D.J.; Leveque, C.; Naiman, R.J.; Prieur-Richard, A.-H.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef]
- Paerl, H.W.; Otten, T.G. Harmful Cyanobacterial Blooms: Causes, Consequences, and Controls. Microb. Ecol. 2013, 65, 995–1010. [Google Scholar] [CrossRef]
- Strayer, D.L.; Dudgeon, D. Freshwater biodiversity conservation: Recent progress and future challenges. J. N. Am. Benthol. Soc. 2010, 29, 344–358. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.J.; He, W.; Liu, W.X.; Qin, N.; Ouyang, H.L.; Wang, Q.M.; Kong, X.Z.; He, Q.S.; Yang, C.; Yang, B.; et al. The seasonal and spatial variations of phytoplankton community and their correlation with environmental factors in a large eutrophic Chinese lake (Lake Chaohu). Ecol. Indic. 2014, 40, 58–67. [Google Scholar] [CrossRef]
- Chen, R.; Shen, J.; Li, C.H.; Zhang, E.L.; Sun, W.W.; Ji, M. Mid- to late-Holocene East Asian summer monsoon variability recorded in lacustrine sediments from Jingpo Lake, Northeastern China. Holocene 2015, 25, 454–468. [Google Scholar] [CrossRef]
- Li, Y.L.; Liao, M.N.; Chen, R.; Shen, J. Diatom-based inference of variations in the strength of East Asian spring wind speeds since mid-Holocene. Palaeogeogr. Palaeocl. 2015, 418, 65–74. [Google Scholar] [CrossRef]
- Lin, K.; Wang, X.; Wang, Y.; Zhou, J.; Liu, L.; Zhu, Y.; Xia, Y. Composition and Influencing Factors of Algal Functional Groups in Jingpo Lake. Res. Environ. Sci. 2017, 30, 520–528. [Google Scholar]
- Song, C.; Yu, H. Phytoplankton Diversity in Jingpo Lake and Water Quality Evaluation. J. North-East For. Univ. 2009, 37, 40–42. [Google Scholar]
- Yuan, Y.; Jiang, M.; Liu, X.; Yu, H.; Otte, M.L.; Ma, C.; Her, Y.G. Environmental variables influencing phytoplankton communities in hydrologically connected aquatic habitats in the Lake Xingkai basin. Ecol. Indic. 2018, 91, 1–12. [Google Scholar] [CrossRef]
- Bellinger, E.G.; Sigee, D.C. Freshwater Algae Identification, Enumeration and Use as Bioindicators; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2015. [Google Scholar]
- Guo, B.Q.; Wang, Q.C.; Hong-Li, Y.U.; Mou, P.P. Stream water quality and its influencing factor in lower order streams in upriver sections of Ashihe River. J. For. Res. 2005, 16, 181–186. [Google Scholar]
- Jin, X.; Tu, Q. Guidelines for Lake Eutrophication Investigation, 2nd ed.; China Environmental Science Press: Beijing, China, 1990. (In Chinese) [Google Scholar]
- Tao, F.; Chao, W.; Hou, J.; Wang, P.; Yao, L.; Dai, Q.; Yang, Y.; You, G. Effect of inter-basin water transfer on water quality in an urban lake: A combined water quality index algorithm and biophysical modelling approach. Ecol. Indic. 2017, 92, 61–71. [Google Scholar]
- Hu, J.C.; Song, Z.B.; Zhou, J.C.; Soininen, J.; Tan, L.; Cai, Q.H.; Tang, T. Differences in diversity and community assembly processes between planktonic and benthic diatoms in the upper reach of the Jinsha River, China. Hydrobiologia 2022, 849, 1577–1591. [Google Scholar] [CrossRef]
- Meng, F.; Li, Z.; Li, L.; Lu, F.; Liu, Y.; Lu, X.; Fan, Y. Phytoplankton alpha diversity indices response the trophic state variation in hydrologically connected aquatic habitats in the Harbin Section of the Songhua River. Sci. Rep. 2020, 10, 21337. [Google Scholar] [CrossRef]
- Motwani, G.; Raman, M.; Matondkar, P.; Parab, S.; Pednekar, S.; Solanki, H. Comparison between phytoplankton bio-diversity and various indices for winter monsoon and inter monsoon periods in north-eastern Arabian Sea. Indian J. Geo-Mar. Sci. 2014, 43, 1513–1518. [Google Scholar]
- Ding, Y.; Pan, B.; Zhao, G.; Sun, C.; Han, X.; Li, M. Geo-climatic factors weaken the effectiveness of phytoplankton diversity as a water quality indicator in a large sediment-laden river. Sci. Total Environ. 2021, 792, 148346. [Google Scholar] [CrossRef] [PubMed]
- Doan-Nhu, H.; Nguyen-Ngoc, L.; Nguyen, C.T. ENSO and anthropogenic impacts on phytoplankton diversity in tropical coastal waters. Prog. Oceanogr. 2016, 140, 1–13. [Google Scholar] [CrossRef]
- Tõnno, I.; Talas, L.; Freiberg, R.; Kisand, A.; Belle, S.; Stivrins, N.; Alliksaar, T.; Heinsalu, A.; Veski, S.; Kisand, V. Environmental drivers and abrupt changes of phytoplankton community in temperate lake Lielais Svētiņu, Eastern Latvia, over the last Post-Glacial period from 14.5 kyr. Quat. Sci. Rev. 2021, 263, 107006. [Google Scholar] [CrossRef]
- Brasil, J.; Santos, J.B.O.; Sousa, W.; Menezes, R.F.; Huszar, V.L.M.; Attayde, J.L. Rainfall leads to habitat homogenization and facilitates plankton dispersal in tropical semiarid lakes. Aquat. Ecol. 2020, 54, 225–241. [Google Scholar] [CrossRef]
- Li, X.; Huang, T.; Ma, W.; Sun, X.; Zhang, H. Effects of rainfall patterns on water quality in a stratified reservoir subject to eutrophication: Implications for management. Sci. Total Environ. 2015, 521, 27–36. [Google Scholar] [CrossRef]
- Rooney, G.G.; van Lipzig, N.; Thiery, W. Estimating the effect of rainfall on the surface temperature of a tropical lake. Hydrol. Earth Syst. Sci. 2018, 22, 6357–6369. [Google Scholar] [CrossRef]
- Onyango, A.O.; Xu, H.; Lin, Z. Diurnal cycle of rainfall over Lake Victoria Basin during the long-rain season based on TRMM satellite estimate. Int. J. Climatol. 2020, 40, 4622–4637. [Google Scholar] [CrossRef]
- Meng, P.J.; Tew, K.S.; Hsieh, H.Y.; Chen, C.C. Relationship between magnitude of phytoplankton blooms and rainfall in a hyper-eutrophic lagoon: A continuous monitoring approach. Mar. Pollut. Bull. 2017, 124, 897–902. [Google Scholar] [CrossRef]
- Pandey, L.K.; Lavoie, I.; Morin, S.; Park, J.; Lyu, J.; Choi, S.; Lee, H.; Han, T. River water quality assessment based on a multi-descriptor approach including chemistry, diatom assemblage structure, and non-taxonomical diatom metrics. Ecol. Indic. 2018, 84, 140–151. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.l.; Lu, Y.l.; He, G.z.; Han, J.y.; Wang, T.y. Exploration of relationships between phytoplankton biomass and related environmental variables using multivariate statistic analysis in a eutrophic shallow lake: A 5-year study. J. Environ. Sci. 2007, 19, 920–927. [Google Scholar] [CrossRef]
- Shi, X.; Sun, Z.; Liu, G.; Xu, H. Insights into community-based discrimination of water quality status using an annual pool of phytoplankton in mid-subtropical canal systems. Environ. Sci. Pollut. R. 2015, 22, 1199–1206. [Google Scholar] [CrossRef] [PubMed]
- Arab, S.; Hamil, S.; Rezzaz, M.A.; Chaffai, A.; Arab, A. Seasonal variation of water quality and phytoplankton dynamics and diversity in the surface water of Boukourdane Lake, Algeria. Arab. J. Geosci. 2019, 12, 29. [Google Scholar] [CrossRef]
- de Souza, D.G.; Bueno, N.C.; Bortolini, J.C.; Rodrigues, L.C.; Bovo-Scomparin, V.M.; de Souza Franco, G.M. Phytoplankton functional groups in a subtropical Brazilian reservoir: Responses to impoundment. Hydrobiologia 2016, 779, 47–57. [Google Scholar] [CrossRef]
- de Oliveira, F.H.P.C.; Capela e Ara, A.L.S.; Moreira, C.H.P.; Lira, O.O.; Padilha, M.D.R.F.; Shinohara, N.K.S. Seasonal changes of water quality in a tropical shallow and eutrophic reservoir in the metropolitan region of Recife (Pernambuco-Brazil). An. Acad. Bras. Cienc. 2014, 86, 1863–1872. [Google Scholar] [CrossRef] [Green Version]
- Hou, L.; Zhou, Z.; Wang, R.; Li, J.; Dong, F.; Liu, J. Research on the Non-Point Source Pollution Characteristics of Important Drinking Water Sources. Water 2022, 14, 211. [Google Scholar] [CrossRef]
- Huang, C.; Li, Y.; Yang, H.; Sun, D.; Yu, Z.; Zhang, Z.; Chen, X.; Xu, L. Detection of algal bloom and factors influencing its formation in Taihu Lake from 2000 to 2011 by MODIS. Environ. Earth Sci. 2014, 71, 3705–3714. [Google Scholar] [CrossRef]
- Cai, Y.; Lu, x.; Ba, Q.; Fan, Y. Plankton community structure and its relationship with environmental factors in spring and summer in Jingpo Lake. Oceanogr. Limnol. 2019, 13, 84–92. (In Chinese) [Google Scholar]
- Arnan, X.; Cerda, X.; Retana, J. Partitioning the impact of environment and spatial structure on alpha and beta components of taxonomic, functional, and phylogenetic diversity in European ants. PEER J. 2015, 3, e1241. [Google Scholar] [CrossRef] [Green Version]
- Korneva, L.G. Changes in phytoplankton diversity in the Volga Basin waterbodies. Inland Water Biol. 2010, 3, 322–328. [Google Scholar] [CrossRef]
- Xiao, J.; Wang, B.; Qiu, X.L.; Yang, M.; Liu, C.Q. Interaction between carbon cycling and phytoplankton community succession in hydropower reservoirs: Evidence from stable carbon isotope analysis. Sci. Total Environ. 2021, 774, 145141. [Google Scholar] [CrossRef]
- Cho, I.H.; Kim, H.K.; Lee, M.H.; Kim, Y.J.; Lee, H.; Kim, B.H. The Effect of Monsoon Rainfall Patterns on Epilithic Diatom Communities in the Hantangang River, Korea. Water 2020, 12, 1471. [Google Scholar] [CrossRef]
- Zhang, Y.; Peng, C.; Huang, S.; Wang, J.; Xiong, X.; Li, D. The relative role of spatial and environmental processes on seasonal variations of phytoplankton beta diversity along different anthropogenic disturbances of subtropical rivers in China. Environ. Sci. Pollut. Res. 2019, 26, 1422–1434. [Google Scholar] [CrossRef] [PubMed]
- Hansson, L.A.; Annadotter, H.; Bergman, E.; Hamrin, S.F.; Jeppesen, E.; Kairesalo, T.; Luokkanen, E.; Nilsson, P.A.; Sondergaard, M.; Strand, J. Biomanipulation as an application of food-chain theory: Constraints, synthesis, and recommendations for temperate lakes. Ecosystems 1998, 6, 558–574. [Google Scholar] [CrossRef]
- Jeppesen, E.; Peder Jensen, J.; Søndergaard, M.; Lauridsen, T.; Junge Pedersen, L.; Jensen, L. Top-down control in freshwater lakes: The role of nutrient state, submerged macrophytes and water depth. Hydrobiologia 1997, 342/343, 151–164. [Google Scholar] [CrossRef]
- Salcher, M.M.; Posch, T.; Pernthaler, J. In situ substrate preferences of abundant bacterioplankton populations in a prealpine freshwater lake. ISME J. 2013, 5, 896–907. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Jiang, Y.J.; He, W.; Liu, W.X.; Kong, X.Z.; Jorgensen, S.E.; Xu, F.L. The tempo-spatial variations of phytoplankton diversities and their correlation with trophic state levels in a large eutrophic Chinese lake. Ecol. Indic. 2016, 66, 153–162. [Google Scholar] [CrossRef]
- Noirmain, F.; Baray, J.-L.; Tridon, F.; Cacault, P.; Billard, H.; Voyard, G.; Van Baelen, J.; Latour, D. Interdisciplinary strategy to survey phytoplankton dynamics of a eutrophic lake under rain forcing: Description of the instrumental set-up and first results. Biogeosciences 2022, 24, 1–30. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SpCond (μs/cm) | WT | DO | pH | TN | TP | CODMn | |
|---|---|---|---|---|---|---|---|
| (℃) | (mg/L) | (mg/L) | (mg/L) | (mg/L) | |||
| Rainy season | 106 ± 77 | 25.37 ± 7.73 | 9.1 ± 3.6 | 9.00 ± 2.715 | 0.20 ± 0.08 | 0.02 ± 0.02 | 1.25 ± 1.2 |
| Dry season | 126.75 ± 44.65 | 10.55 ± 6.45 | 8 ± 6 | 8.13 ± 1.415 | 0.14 ± 0.09 | 0.03 ± 0.023 | 3.16 ± 2.98 |
| 2017 | 123.5 ± 94.5 | 15.93 ± 7.53 | 8 ± 6 | 8.995 ± 2.715 | 0.15 ± 0.08 | 0.03 ± 0.0225 | 3.09 ± 3.04 |
| 2018 | 133.4 ± 48.4 | 14.33 ± 12.69 | 8.13 ± 3.88 | 8.73 ± 1.55 | 0.23 ± 0.17 | 0.03 ± 0.02 | 2.69 ± 2.51 |
| t-Test | |||||||
| Dry season × Rainy season | ns | p < 0.01 | ns | ns | p < 0.01 | p < 0.05 | p < 0.05 |
| 2017 × 2018 | ns | ns | ns | ns | ns | ns | ns |
| t-Test | |||||||
|---|---|---|---|---|---|---|---|
| Rainy Season | Dry Season | 2017 | 2018 | 2017–2018 | Rainy Season × Dry Season | 2017 × 2018 | |
| Average abundance (×104 ind./L) | 159.91 | 100.27 | 77.10 | 188.25 | 130.09 | p < 0.05 | p < 0.05 |
| Bacillariophyta (×104 ind./L) | 101.83 ± 165.52 | 79.03 ± 172.29 | 68.19 ± 182.57 | 114.77 ± 150.04 | 90.43 | p < 0.05 | p < 0.05 |
| Cyanobacteria (×104 ind./L) | 25.56 ± 54.56 | 1.82 ± 4.94 | 2.20 ± 5.90 | 26.64 ± 56.00 | 13.69 | p < 0.05 | p < 0.05 |
| Chlorophyta (×104 ind./L) | 19.79 ± 40.36 | 2.94 ± 5.56 | 1.19 ± 2.94 | 22.70 ± 40.75 | 11.37 | p < 0.05 | p < 0.05 |
| Cryptophyta (×104 ind./L) | 9.29 ± 14.74 | 13.67 ± 22.82 | 4.43 ± 9.72 | 18.83 ± 23.51 | 11.48 | ns | p < 0.05 |
| Euglenophyta (×104 ind./L) | 2.92 ± 6.60 | 1.71 ± 7.78 | 0.65 ± 1.61 | 4.12 ± 9.93 | 2.32 | ns | p < 0.05 |
| other (×104 ind./L) | 0.52 ± 1.38 | 1.10 ± 3.14 | 0.43 ± 2.27 | 1.20 ± 2.51 | 0.81 | ns | ns |
| Margalef index | 2.15 ± 2.58 | 2.27 ± 2.66 | 1.31 ± 2.66 | 3.15 ± 1.94 | 2.23 | p < 0.05 | p < 0.01 |
| Shannon-Wiener index | 1.69 ± 1.34 | 1.54 ± 1.42 | 1.21 ± 1.28 | 2.04 ± 1.88 | 1.63 | p < 0.05 | p < 0.01 |
| Pielou index | 0.70 ± 0.36 | 0.72 ± 0.28 | 0.74 ± 0.33 | 0.68 ± 0.33 | 0.71 | ns | ns |
| Rainy Season | Dry Season | 2017 | 2018 | ||
|---|---|---|---|---|---|
| Bacillariophyta | Fragilaria capucina | 19.08% | 2.76% | 3.65% | 17.10% |
| Ulnaria ulna | 8.49% | 3.75% | 4.42% | 7.74% | |
| Asterionella formosa | 0.14% | 0.32% | 0.14% | 0.17% | |
| Melosira granulata | 12.63% | 4.80% | 2.24% | 12.97% | |
| Melosira granulata var. angustissima | 0.76% | 0.22% | 1.56% | 0.12% | |
| Aulacoseira pusilla | 0.14% | 0.36% | 0.45% | 0.14% | |
| Melosira varians | 0.29% | 13.72% | 16.58% | 0.35% | |
| Nitzschia palea | 5.26% | 6.86% | 11.49% | 3.41% | |
| Cymbella turgidula | 3.78% | 4.86% | 8.48% | 2.31% | |
| Gomphonema parvulum | 0.15% | 0.97% | 1.08% | 0.18% | |
| Chlorophyta | Crucigenia rectangularis | 4.64% | 0.07% | 0.14% | 4.23% |
| Crucigenia apiculata | 1.52% | 0.07% | 0.14% | 1.40% | |
| Schroederia spiralis | 1.03% | 0.52% | 0.14% | 1.21% | |
| Schroederia robusta | 0.14% | 0.27% | 0.19% | 0.09% | |
| Ankistrodesmus angustus | 0.72% | 0.63% | 0.14% | 0.97% | |
| Ankistrodesmus falcatus | 0.08% | 0.63% | 0.14% | 0.35% | |
| Ankistrodesmus acicularis | 0.06% | 0.06% | 0.06% | 0.06% | |
| Eudorina echidna | 1.03% | 0.14% | 0.11% | 0.90% | |
| Pandorina morum | 0.24% | 0.14% | 0.46% | 0.14% | |
| Ulothrix subconstricta | 0.06% | 0.06% | 0.06% | 0.06% | |
| Cyanobacteria | Pseudanabaena limnetica | 6.10% | 0.19% | 0.06% | 5.58% |
| Anabaena azotica | 4.12% | 0.31% | 1.33% | 3.31% | |
| Anabaena catenula | 3.78% | 0.14% | 0.14% | 3.42% | |
| Aphanizomenon flosaquae | 0.35% | 0.14% | 0.14% | 0.32% | |
| Oscillatoria princes | 0.22% | 0.17% | 0.21% | 0.20% | |
| Aphanocapsa delicatissima | 0.14% | 0.14% | 0.14% | 0.14% | |
| Merismopedia tenuissima | 0.14% | 0.68% | 0.14% | 0.48% | |
| Chroococcus minutus | 0.07% | 0.07% | 0.07% | 0.07% | |
| Cryptophyta | Chroomonas caudata | 0.68% | 2.56% | 0.14% | 1.97% |
| Chroomonas acuta | 0.16% | 0.20% | 0.57% | 0.14% | |
| Cryptomonas erosa | 3.65% | 10.59% | 5.01% | 6.76% | |
| Cryptomonas ovata | 1.32% | 0.28% | 0.16% | 1.27% | |
| Euglenophyta | Euglena viridis | 0.65% | 0.46% | 0.22% | 0.74% |
| Trachelomonas oblonga | 0.49% | 0.66% | 0.42% | 0.61% |
| Margalef Index | Shannon-Wiener Index | Pielou Evenness Index | |
|---|---|---|---|
| All | |||
| Correlation with TLI | 0.066 | 0.074 | 0.074 |
| Rainy-season | |||
| Correlation with TLI | 0.495 ** | 0.494 ** | 0.043 |
| Dry-season | |||
| Correlation with TLI | −0.513 ** | −0.411 ** | 0.172 |
| 2017 | |||
| Correlation with TLI | 0.219 | 0.273 * | 0.241 |
| 2018 | |||
| Correlation with TLI | −0.005 | −0.062 | −0.125 |
| Margalef Index | Shannon-Wiener Index | Pielou Evenness Index | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Samples | All | Dry Season | Rainy Season | All | Dry Season | Rainy Season | All | Dry Season | Rainy Season |
| Margalef index | |||||||||
| All | 0.886 ** | −0.241 ** | |||||||
| Dry Season | 0.64 | 0.92 ** | 0.6 | 0.53 | 0.13 | ||||
| Rainy Season | 0.54 | 0.93 ** | 0.31 | 0.24 | |||||
| Shannon-Wiener index | |||||||||
| All | 0.082 | ||||||||
| Dry Season | 0.51 | 0.78 ** | 0.13 | ||||||
| Rainy Season | 0.29 | 0.53 | |||||||
| Pielou evenness index | |||||||||
| All | |||||||||
| Dry Season | 0.12 | ||||||||
| Rainy Season | |||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, Y.; Qi, L.; Shan, T.; Liu, Y.; Zhang, N.; Lu, X.; Fan, Y. Application of Phytoplankton Taxonomic α-Diversity Indices to Assess Trophic States in Barrier Lake: A Case of Jingpo Lake. Diversity 2022, 14, 1003. https://doi.org/10.3390/d14111003
Cai Y, Qi L, Shan T, Liu Y, Zhang N, Lu X, Fan Y. Application of Phytoplankton Taxonomic α-Diversity Indices to Assess Trophic States in Barrier Lake: A Case of Jingpo Lake. Diversity. 2022; 14(11):1003. https://doi.org/10.3390/d14111003
Chicago/Turabian StyleCai, Yang, Lin Qi, Tao Shan, Yan Liu, Nannan Zhang, Xinxin Lu, and Yawen Fan. 2022. "Application of Phytoplankton Taxonomic α-Diversity Indices to Assess Trophic States in Barrier Lake: A Case of Jingpo Lake" Diversity 14, no. 11: 1003. https://doi.org/10.3390/d14111003