
Reproductive Ecology and Nesting Site Characteristics of Four-Toed Salamanders (Hemidactylium scutatum) in Natural and Constructed Upland-Embedded Wetlands on the Appalachian Plateau, Kentucky


Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Analyses at the Wetland Level
3.2. Analyses at the Nest Level
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wright, A.H. Notes on the Muhlenberg’s turtle. Copeia 1918, 1918, 5–7. [Google Scholar] [CrossRef]
- Dahl, T.E. Status and Trends of Wetlands in the Conterminous United States 1986 to 1997; U.S. Department of the Interior, Fish and Wildlife Service: Washington, DC, USA, 2000. [Google Scholar]
- Dahl, T.E. Status and Trends of Wetlands in the Conterminous United States 2004 to 2009; U.S. Department of the Interior, Fish and Wildlife Service: Washington, DC, USA, 2011. [Google Scholar]
- Calhoun, A.J.K.; Arrigoni, J.; Brooks, R.P.; Hunter, M.L.; Richter, S.C. Creating successful vernal pools: A literature review and advice for practitioners. Wetlands 2014, 34, 1027–1038. [Google Scholar] [CrossRef] [Green Version]
- Pechmann, J.H.K.; Estes, R.A.; Scott, D.E.; Gibbons, J.W. Amphibian colonization and use of ponds created for trial mitigation of wetland loss. Wetlands 2001, 21, 93–111. [Google Scholar] [CrossRef]
- Denton, R.D.; Richter, S.C. Amphibian communities in natural and constructed ridge top wetlands with implication for wetland construction. J. Wildl. Manag. 2013, 77, 886–896. [Google Scholar] [CrossRef]
- Drayer, A.N.; Richter, S.C. Physical wetland characteristics influence amphibian community composition in constructed wetlands. Ecol. Eng. 2016, 93, 166–174. [Google Scholar] [CrossRef]
- Semlitsch, R.D.; Scott, D.E.; Pechmann, J.H.K. Time and size at metamorphosis related to adult fitness in Ambystoma talpoideum. Ecology 1988, 69, 184–192. [Google Scholar] [CrossRef]
- Rowe, C.L.; Dunson, W.A. Impacts of hydroperiod on growth and survival of larval amphibians in temporary ponds of central Pennsylvania, USA. Oecologia 1995, 102, 397–403. [Google Scholar] [CrossRef]
- Vasconcelos, D.; Calhoun, A.J.K. Monitoring created seasonal pools for functional success: A six-year case study of amphibian responses, Sears Island, Maine, USA. Wetlands 2006, 26, 992–1003. [Google Scholar] [CrossRef]
- Kross, C.S.; Richter, S.C. Species interactions in constructed wetlands result in population sinks for wood frogs (Lithobates sylvaticus) while benefitting eastern newts (Notophthalmus viridescens). Wetlands 2016, 36, 385–393. [Google Scholar] [CrossRef]
- Petranka, J.W. Salamanders of the United States and Canada; Smithsonian Institution Press: Washington, DC, USA, 1998. [Google Scholar]
- Beazley, K.; Cardinal, N. A systematic approach for selecting focal species for conservation in the forests of Nova Scotia and Maine. Environ. Conserv. 2004, 31, 91–101. [Google Scholar] [CrossRef]
- Harris, R.N. Four-toed salamander, Hemidactylium scutatum. In Amphibian Declines: The Conservation Status of United States Species; Lannoo, M.J., Ed.; University of California Press: Berkeley, CA, USA, 2005; pp. 780–781. [Google Scholar]
- Harris, R.N.; Gill, D.E. Communal nesting, brooding behavior, and embryonic survival of the four-toed salamander Hemidactylium scutatum. Herpetologica 1980, 36, 141–144. [Google Scholar]
- Harris, R.N.; Hames, W.W.; Knight, I.T.; Carreño, C.A.; Vess, T.J. Experimental analysis of joint nesting in the salamander Hemidactylium scutatum (Caudata: Plethodontidae), the effects of population density. Anim. Behav. 1995, 50, 1309–1316. [Google Scholar] [CrossRef]
- Harris, R.N. Body condition and order of arrival affect cooperative nesting behaviour in four-toed salamanders, Hemidactylium scutatum. Anim. Behav. 2008, 75, 229–233. [Google Scholar] [CrossRef]
- Chalmers, R.J.; Loftin, C.S. Wetland and microhabitat use by nesting four-toed salamanders in Maine. J. Herpetol. 2006, 40, 478–485. [Google Scholar] [CrossRef]
- Wahl, G.W.; Harris, R.N.; Nelms, T. Nest site selection and embryonic survival in four-toed salamanders, Hemidactylium scutatum (Caudata, Plethodontidae). Herpetologica 2008, 64, 12–19. [Google Scholar] [CrossRef]
- Resetarits, W.J.; Wilbur, H.M. Choice of oviposition site by Hyla chrysoscelis: Role of predators and competitors. Ecology 1989, 70, 220–228. [Google Scholar] [CrossRef]
- Brown, D.R.; Richter, S.C. Meeting the challenges to preserving Kentucky’s biodiversity. Sustain 2012, 25, 22–33. [Google Scholar]
- Woods, A.J.; Omernik, J.M.; Martin, W.H.; Pond, G.J.; Andrews, W.M.; Call, S.M.; Comstock, J.A.; Taylor, D.D. Ecoregions of Kentucky (Color Poster with Map, Descriptive Text, Summary Tables and Photographs); Map Scale 1:1,000,000; Geological Survey: Reston, VA, USA, 2002. [Google Scholar]
- Skelly, D.K.; Freidenburgh, L.K.; Kiesecker, J.M. Forest canopy and the performance of larval amphibians. Ecology 2002, 83, 983–992. [Google Scholar] [CrossRef]
- Schiesari, L. Pond canopy cover: A resource gradient for anuran larvae. Freshw. Biol. 2006, 51, 412–423. [Google Scholar] [CrossRef] [Green Version]
- Guidugli-Cook, M.; Richter, S.C.; Scott, B.J.; Brown, D.R. Field-based assessment of wetland condition, wetland extent, and the National Wetlands Inventory in Kentucky, USA. Wetl. Ecol. Manag. 2017, 25, 517–532. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis, 5th ed.; Prentice Hall: Hoboken, NJ, USA, 2010. [Google Scholar]
- Mazerolle, M.J. AICcmodavg: Model Selection and Multimodel Inference Based on (Q)AIC(c). R Package Version 2.1-0. 2016. Available online: https://cran.r-project.org/package=AICcmodavg (accessed on 11 October 2022).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Arnold, T.W. Uninformative parameters and model selection using Akaike’s Information Criterion. J. Wildl. Manag. 2010, 74, 1175–1178. [Google Scholar] [CrossRef]
- Blanchard, F.N. The life history of the four-toed salamander. Am. Nat. 1923, 57, 262–268. [Google Scholar] [CrossRef]
- Porej, D.; Hetherington, T.E. Designing wetlands for amphibians: The importance of predatory fish and shallow littoral zones in structuring of amphibian communities. Wetl. Ecol. Manag. 2005, 13, 445–455. [Google Scholar] [CrossRef]
- Breitenbach, G.L. The frequency of communal nesting and solitary brooding in the salamander, Hemidactylium scutatum. J. Herpetol. 1982, 16, 341–346. [Google Scholar] [CrossRef]
- Biebighauser, T.R. Wetland Restoration and Construction, A Technical Guide; Upper Susquehanna Coalition: Owego, NY, USA, 2011. [Google Scholar]
- Fenton, N.J.; Frego, K.A. Bryophyte (moss and liverwort) conservation under remnant canopy in managed forests. Biol. Conserv. 2005, 122, 41–7430. [Google Scholar] [CrossRef]
- Botting, R.S.; Fredeen, A.L. Contrasting terrestrial lichen, liverwort, and moss diversity between old-growth and young second-growth forest on two soil textures in central British Columbia. Botany 2006, 84, 120–132. [Google Scholar] [CrossRef] [Green Version]
- Stewart, K.J.; Mallik, A.U. Bryophyte responses to microclimatic edge effects across riparian buffers. Ecol. Appl. 2006, 16, 1474–1486. [Google Scholar] [CrossRef]
- Fedders, R.B. Amphibian and Plant Communities of Natural and Constructed Upland-Embedded Wetlands in the Daniel Boone National Forest. Master’s Thesis, Eastern Kentucky University, Richmond, KY, USA, 2018. [Google Scholar]
- Lott, R.B.; Hunt, R.J. Estimating evapotranspiration in natural and constructed wetlands. Wetlands 2001, 21, 614–628. [Google Scholar] [CrossRef]
- Corser, J.D.; Dodd, C.K., Jr. Fluctuations in a metapopulation of nesting four-toed salamanders, Hemidactylium scutatum, in the Great Smoky Mountains National Park, USA, 1999–2003. Nat. Areas J. 2004, 24, 135–140. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nest and Wetland Factors | Natural Wetlands | Constructed Wetlands | Statistics |
|---|---|---|---|
| Wetland-level factors | |||
| Nests Eggs Perimeter (m) | 22.2 ± 13.5 808.3 ± 367.1 94.0 ± 26.6 | 12.3 ± 3.4 756.2 ± 187.7 46.1 ± 5.9 | U = 16.5, p = 0.81 U = 14.0, p = 0.52 U = 7.0, p = 0.078 |
| Area (m2) | 383.0 ± 194.0 | 138.6 ± 49.3 | U = 11.0, p = 0.262 |
| Total moss (m) | 41.7 ± 29.0 | 10.2 ± 5.6 | U = 13.0, p = 0.423 |
| Canopy-closure (%) | 96.1 ± 1.5 | 84.7 ± 4.6 | U = 3.0, p = 0.016 |
| KY-WRAM score | 72.5 ± 3.6 | 47.7 ± 4.3 | U = 3.0, p = 0.016 |
| Nest-level factors | |||
| Slope (degrees) | 53.41 ± 1.86 | 34.86 ± 2.06 | F = 0.9, P1,12.9 = 0.351 |
| Distance to water (cm) | 9.17 ± 0.51 | 15.58 ± 1.26 | F = 0.9, P1,12.6 = 0.368 |
| Moisture (%) | 28.9 ± 0.76 | 25.9 ± 0.53 | F = 2.7, P1,11.5 = 0.126 |
| pH | 5.78 ± 0.05 | 6.27 ± 0.03 | F = 8.7, P1,15.8 = 0.010 |
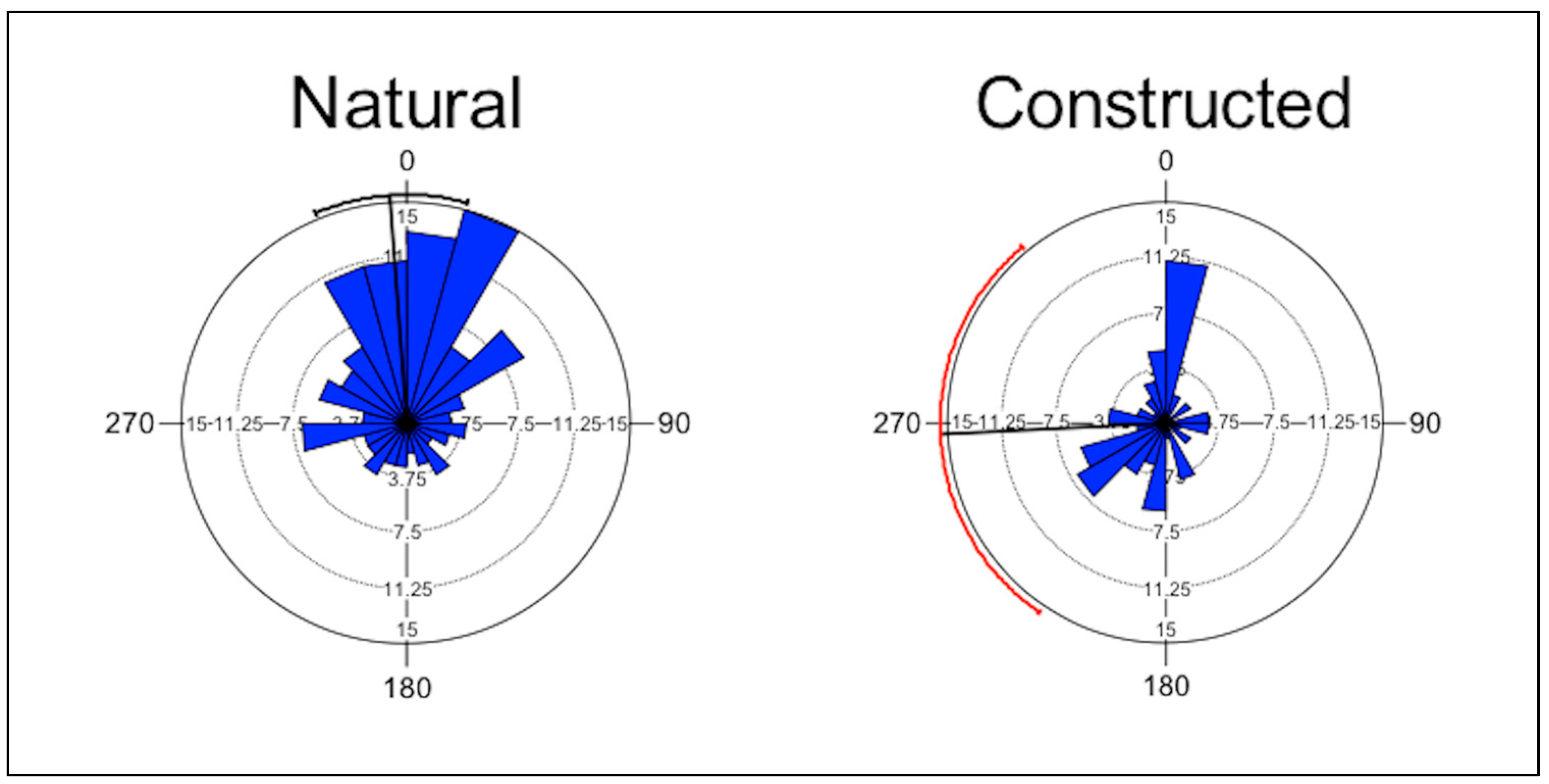
| Wetland | Type | Rayleigh Test (Z) | Rayleigh Test (p) | Mean Vector (µ) | Circular Standard Deviation |
|---|---|---|---|---|---|
| Booth | Natural | 7.85 | <0.001 | 348.45 | 89.27 |
| DC2 | Natural | 2.48 | 0.082 | 16.67 | 76.90 |
| DC6 | Natural | 1.89 | 0.152 | 31.88 | 73.97 |
| ELN | Natural | 3.26 | 0.033 | 14.87 | 54.30 |
| HEN | Natural | 0.78 | 0.488 | 317.89 | 73.21 |
| JRN | Natural | 3.36 | 0.028 | 334.80 | 49.08 |
| 35–97 | Constructed | 4.42 | 0.011 | 337.09 | 74.53 |
| ELA | Constructed | 0.06 | 0.948 | 344.43 | 120.69 |
| HEA | Constructed | 2.60 | 0.073 | 173.35 | 78.55 |
| JRA | Constructed | 0.13 | 0.901 | 80.00 | 94.20 |
| LRA | Constructed | 1.62 | 0.202 | 215.03 | 77.32 |
| P5 | Constructed | 3.57 | 0.025 | 263.92 | 70.16 |
| Parameter | K | Log-Likelihood | AICC | ΔAICC | Weight |
|---|---|---|---|---|---|
| Number of eggs per wetland | |||||
| Total moss | 3 | −9.17 | 27.34 | 0 | 0.60 |
| Wetland-type, total moss | 4 | −8.17 | 30.06 | 2.73 | 0.15 |
| Wetland-type, total moss, canopy | 5 | −5.33 | 30.66 | 3.32 | 0.11 |
| Intercept-only | 2 | −12.90 | 31.12 | 3.79 | 0.09 |
| Canopy | 3 | −12.49 | 33.98 | 6.64 | 0.02 |
| Wetland-type | 3 | −12.88 | 34.76 | 7.42 | 0.01 |
| Number of nests per wetland | |||||
| Total moss | 3 | −10.53 | 30.06 | 0 | 0.87 |
| Wetland-type, total moss | 4 | −10.32 | 34.35 | 4.29 | 0.10 |
| Intercept-only | 2 | −16.09 | 37.51 | 7.45 | 0.02 |
| Number of eggs per nest | |||||
| Global model | 13 | −543.13 | 1115.78 | 0 | 0.99 |
| Nest-level full model | 10 | −552.01 | 1126.10 | 10.32 | 0.01 |
| Dependent Variable | Predictor Variable | Model-Averaged Estimate (¦Â) | Unconditional SE | 85% CI |
|---|---|---|---|---|
| No. of eggs | Total moss | 0.010 | 0.003 | 0.005, 0.015 |
| Canopy | 0.036 | 0.020 | 0.007, 0.065 | |
| Wetland-type | ||||
| Natural | −0.593 | 0.431 | −1.214, 0.028 | |
| Constructed | 0 | 0 | 0, 0 | |
| No. of nests | Total moss | 0.015 | 0.004 | 0.009, 0.020 |
| Canopy | 0.010 | 0.029 | −0.032, 0.051 | |
| Wetland-type | ||||
| Natural | −0.217 | 0.428 | −0.833, 0.399 | |
| Constructed | 0 | 0 | 0, 0 | |
| Eggs per nest | All moss substrate | −0.001 | 0.002 | −0.003, 0.003 |
| Slope | −0.002 | 0.004 | −0.007, 0.003 | |
| Moisture | 0.038 | 0.012 | 0.020, 0.055 | |
| pH | −0.109 | 0.181 | −0.370, 0.152 | |
| Female presence | 0.315 | 0.072 | 0.211, 0.419 | |
| Distance to water | −0.007 | 0.003 | −0.122. −0.002 | |
| Total moss | −0.003 | 0.001 | −0.005, −0.002 | |
| Canopy | 0.031 | 0.012 | 0.013, 0.048 | |
| Wetland-type | ||||
| Natural | −0.690 | 0.207 | −0.988, −0.392 | |
| Constructed | 0 | 0 | 0, 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
King, S.K.; Richter, S.C. Reproductive Ecology and Nesting Site Characteristics of Four-Toed Salamanders (Hemidactylium scutatum) in Natural and Constructed Upland-Embedded Wetlands on the Appalachian Plateau, Kentucky. Diversity 2022, 14, 995. https://doi.org/10.3390/d14110995
King SK, Richter SC. Reproductive Ecology and Nesting Site Characteristics of Four-Toed Salamanders (Hemidactylium scutatum) in Natural and Constructed Upland-Embedded Wetlands on the Appalachian Plateau, Kentucky. Diversity. 2022; 14(11):995. https://doi.org/10.3390/d14110995
Chicago/Turabian StyleKing, Susan K., and Stephen C. Richter. 2022. "Reproductive Ecology and Nesting Site Characteristics of Four-Toed Salamanders (Hemidactylium scutatum) in Natural and Constructed Upland-Embedded Wetlands on the Appalachian Plateau, Kentucky" Diversity 14, no. 11: 995. https://doi.org/10.3390/d14110995