
Effects of Land Use and Pollution Loadings on Ecotoxicological Assays and Bacterial Taxonomical Diversity in Constructed Wetlands

Abstract
:1. Introduction
2. Materials and Methods
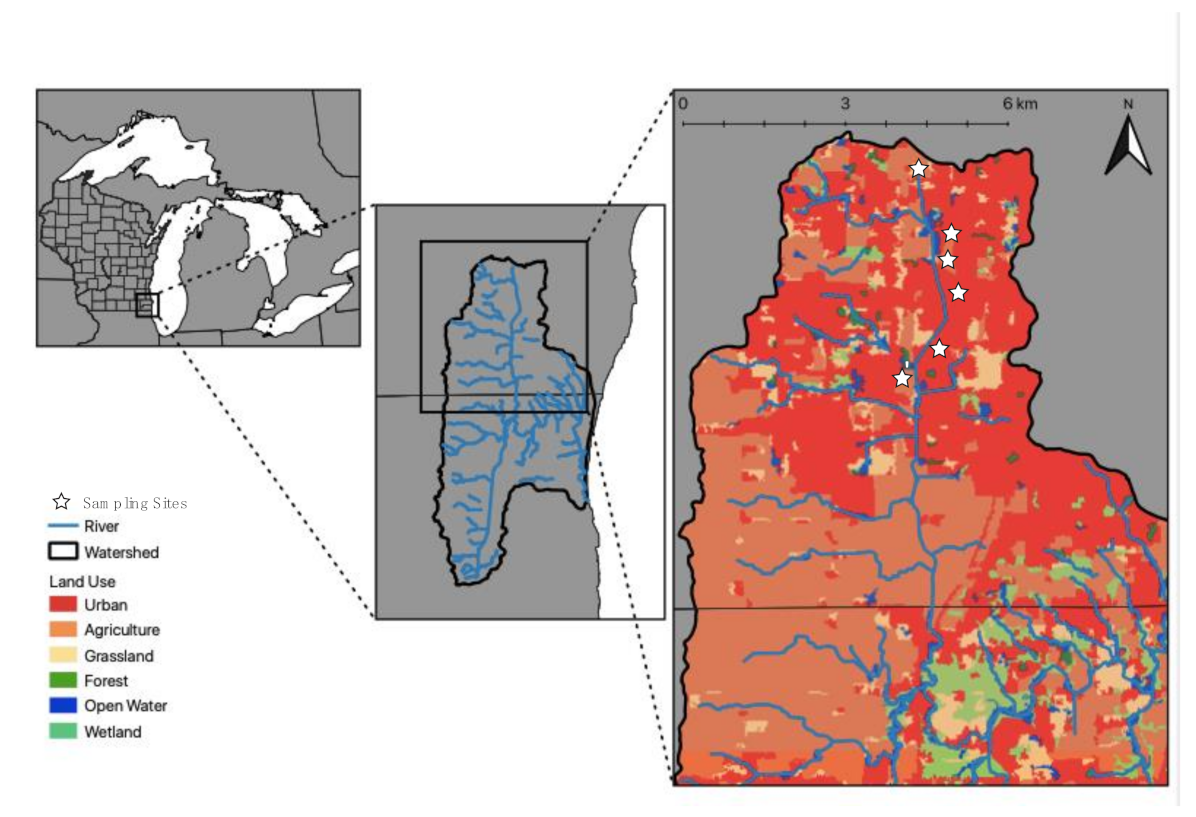
2.1. Study System, Land Use, and Site Characteristics
2.2. Sediment Sampling Collection and Water Quality Monitoring
2.3. Ecotoxicological Assays and Nutrient and Metal Measurements in Wetland Sites
2.4. Bacterial Community Structure
2.5. Data Analyses
3. Results
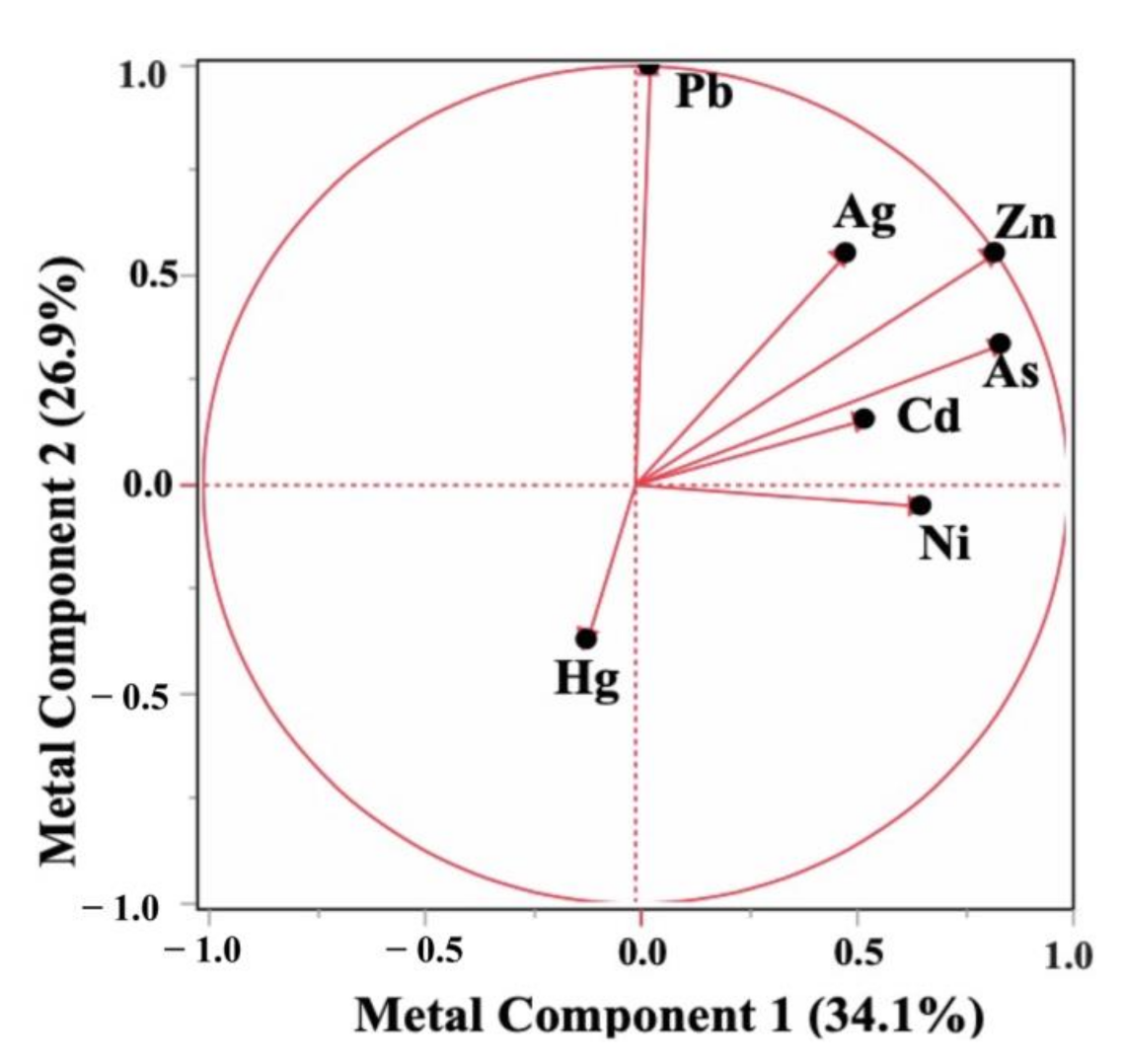
3.1. Distribution of Measured Nutrients, Metals and Factor Analysis of the Metal Concentrations
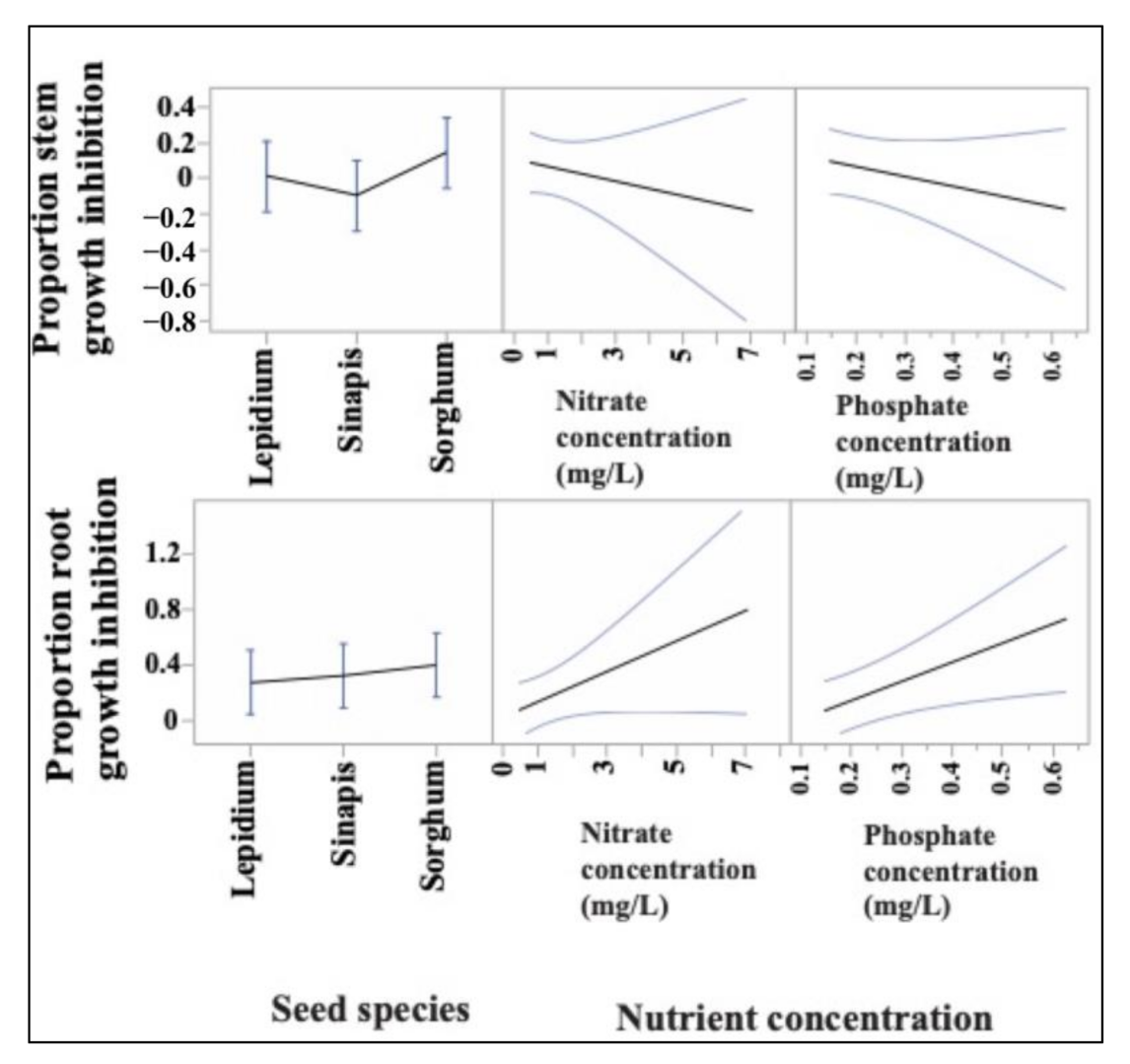
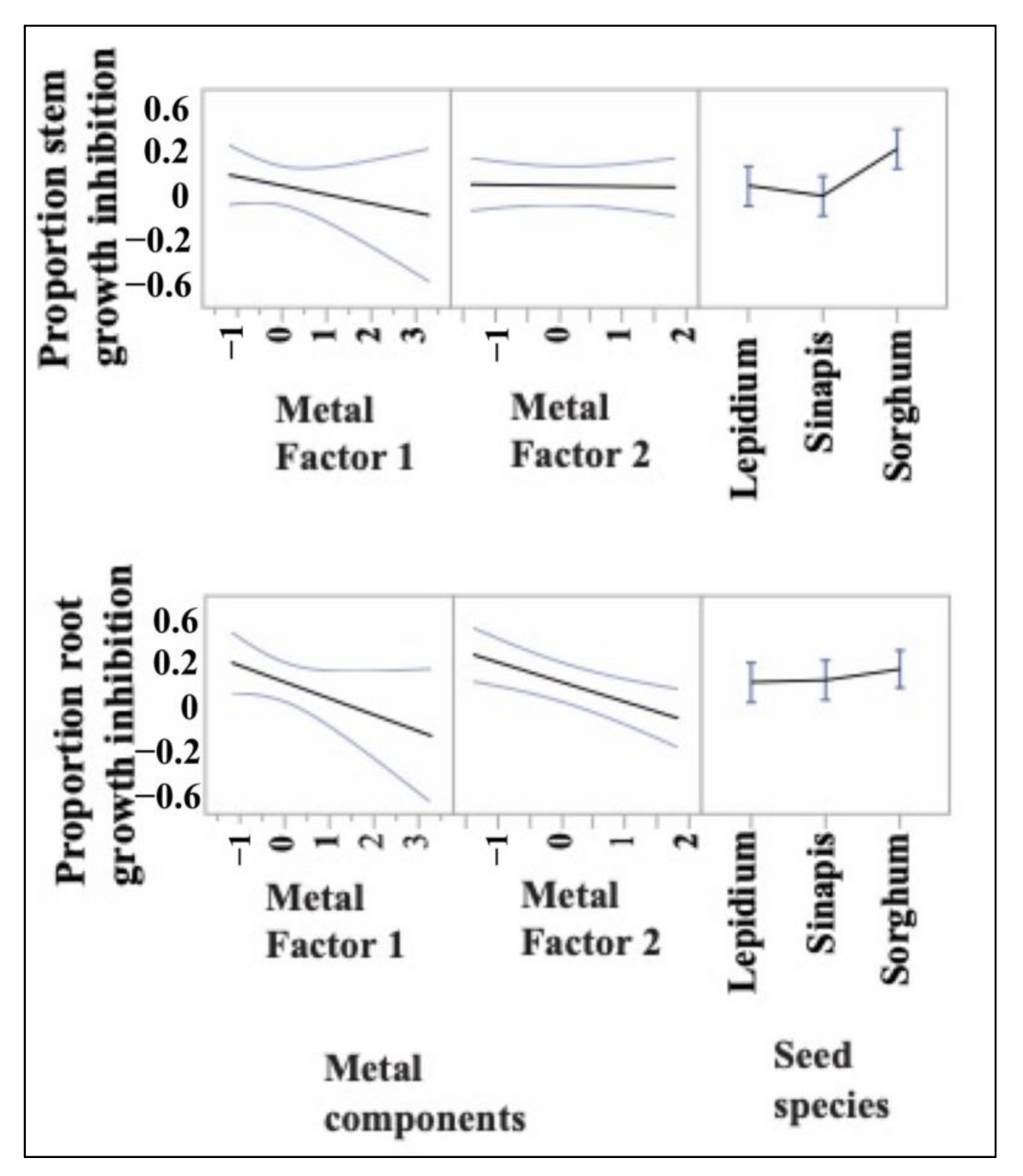
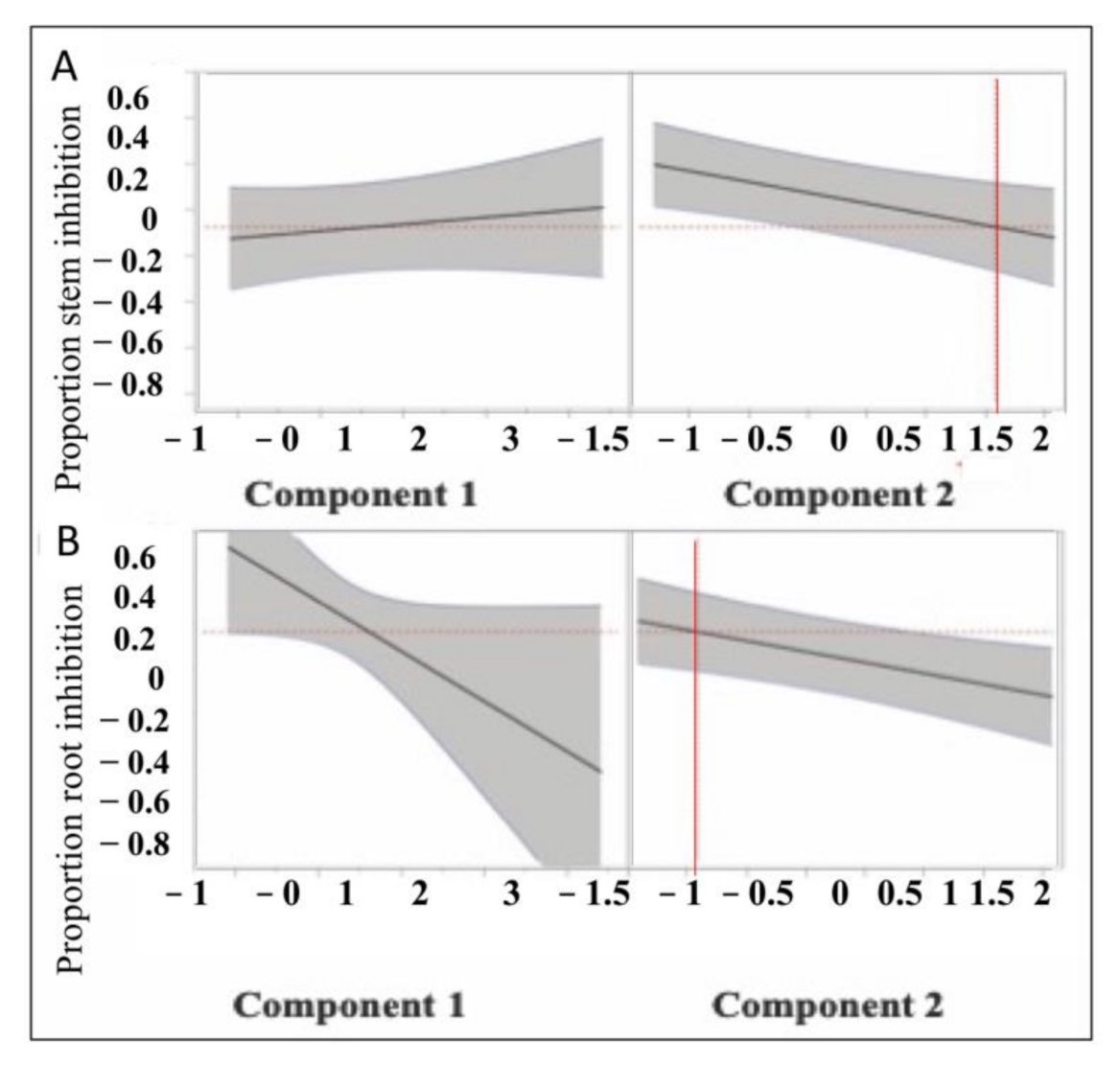
3.2. Ecotoxicological Indicators and Their Response to Nutrient and Metal Pollution Stress
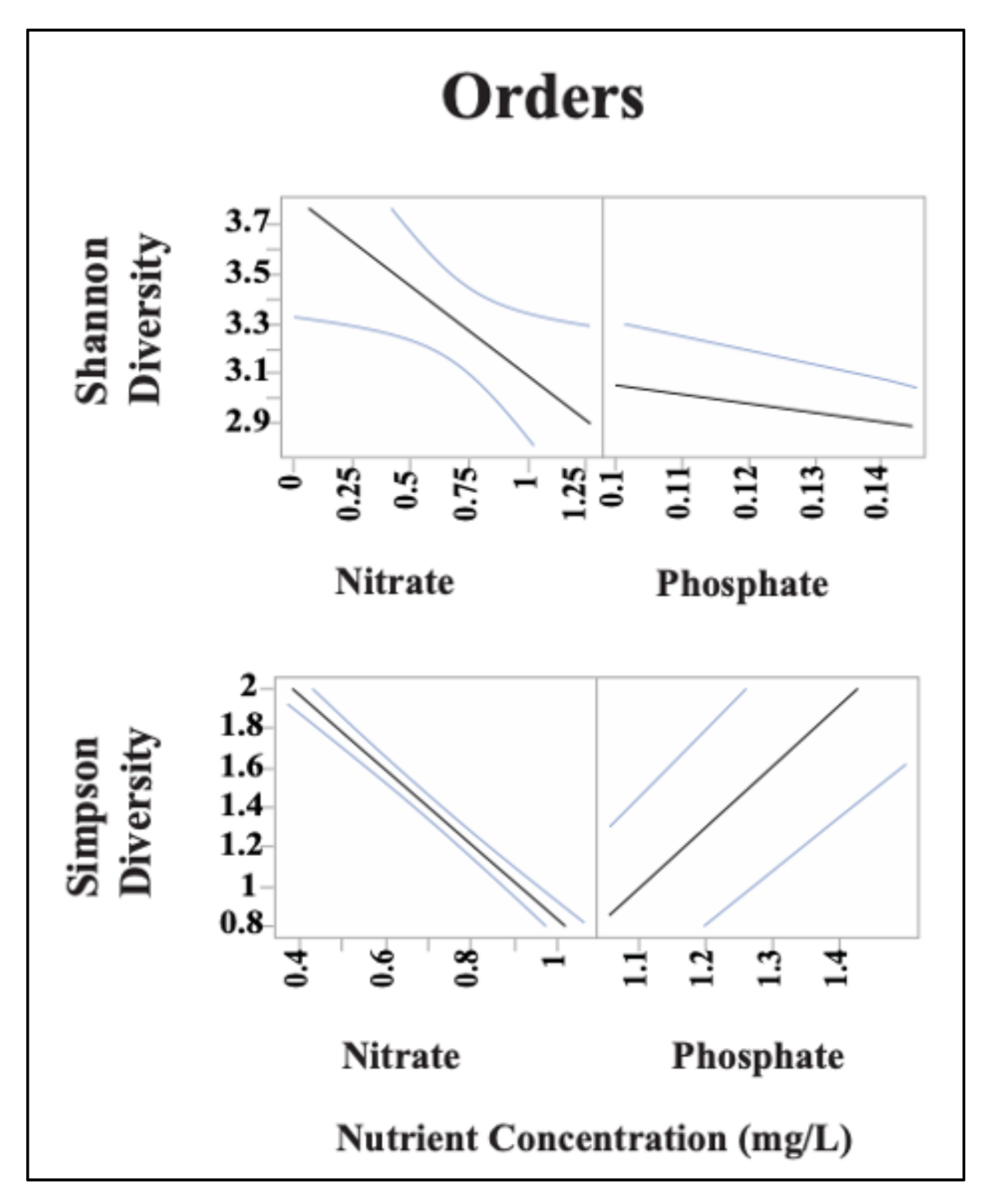
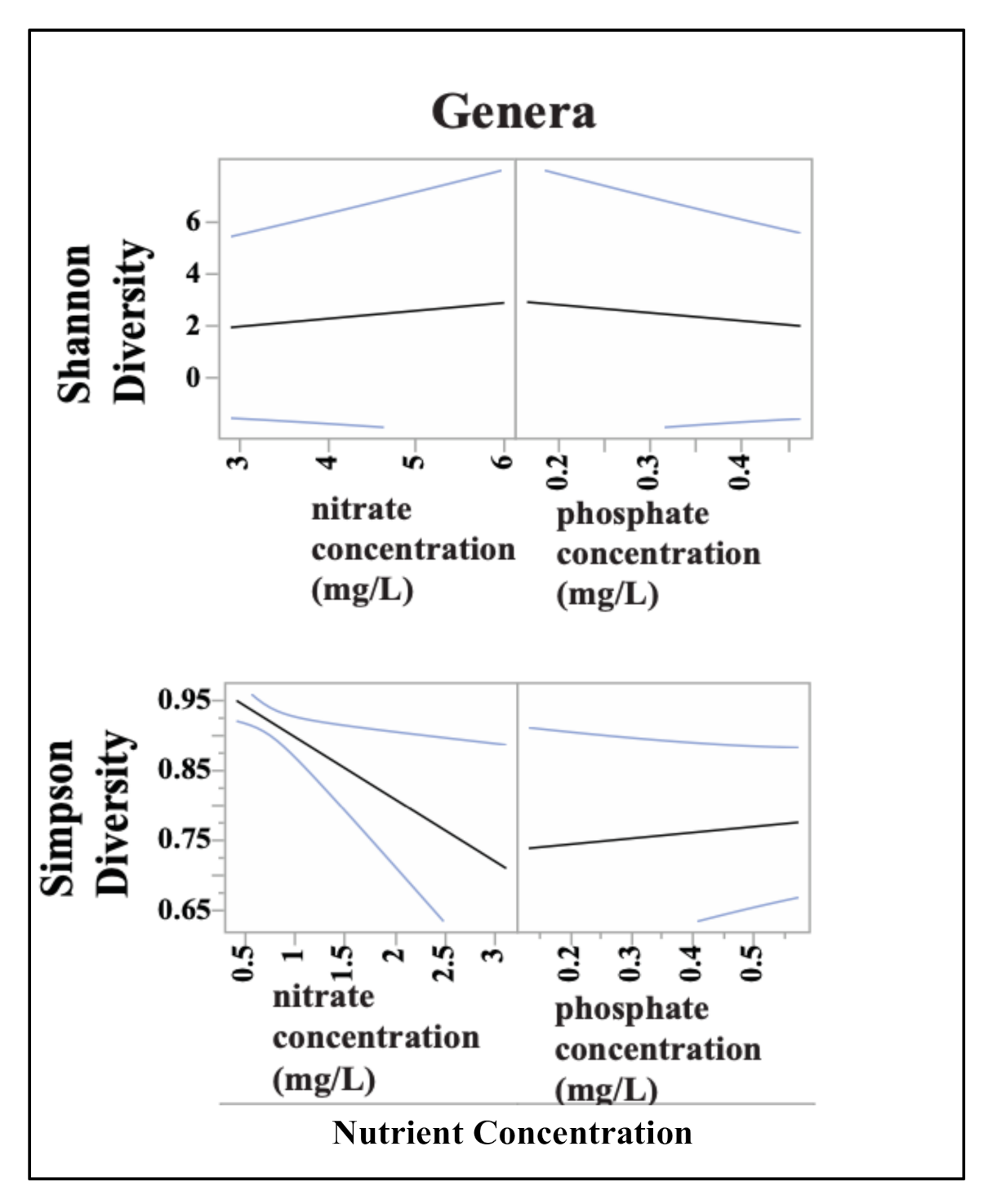
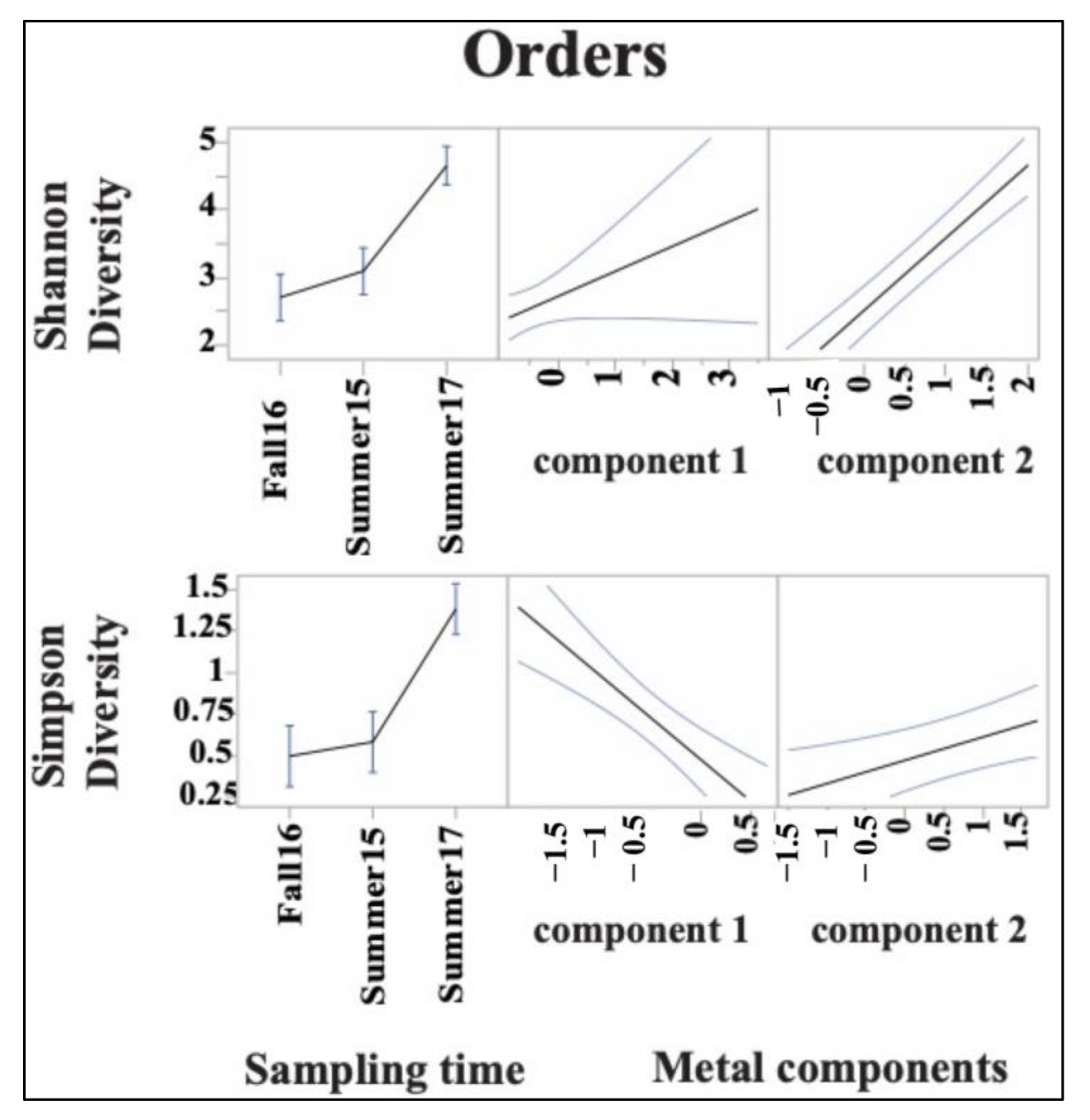
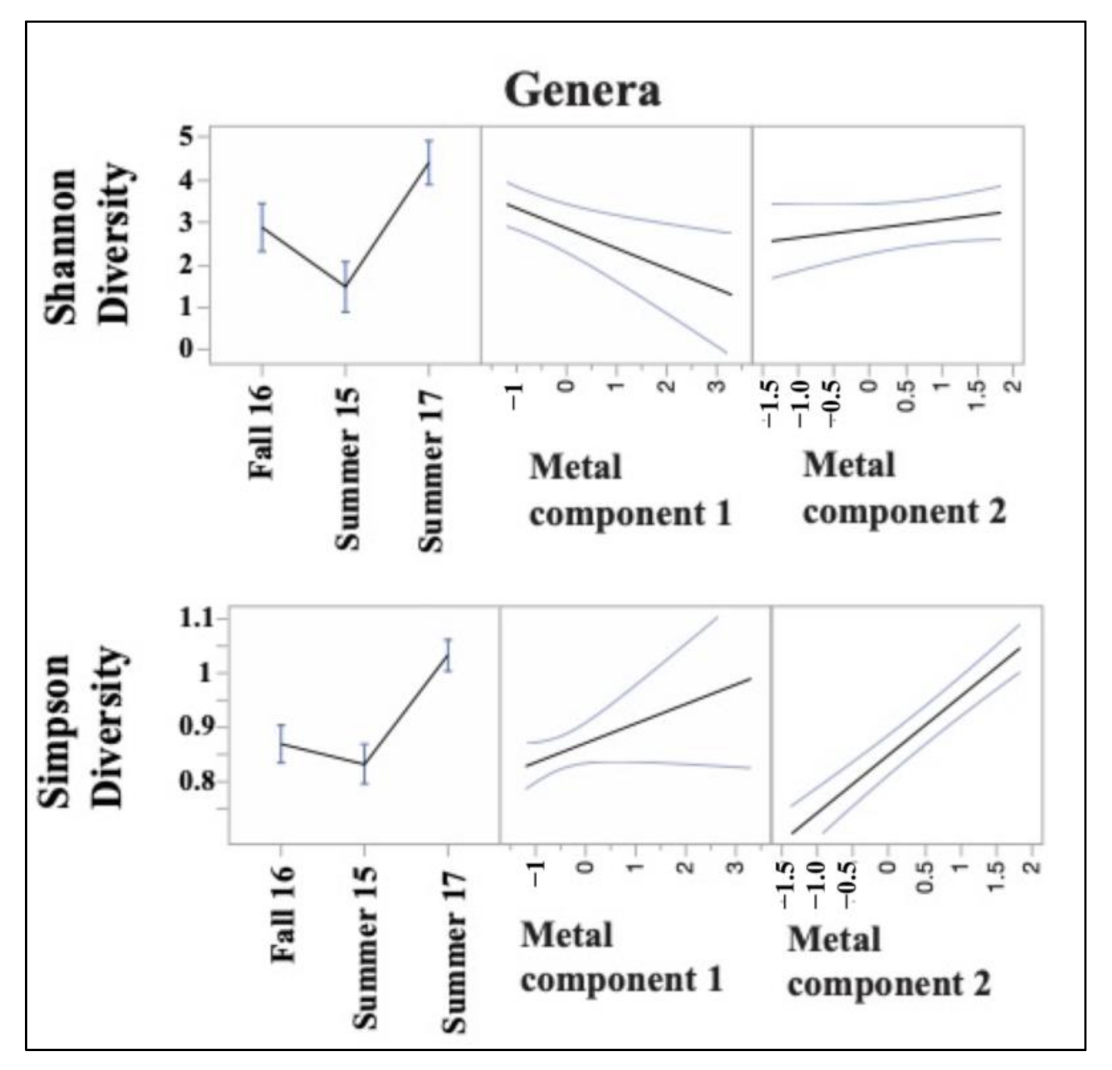
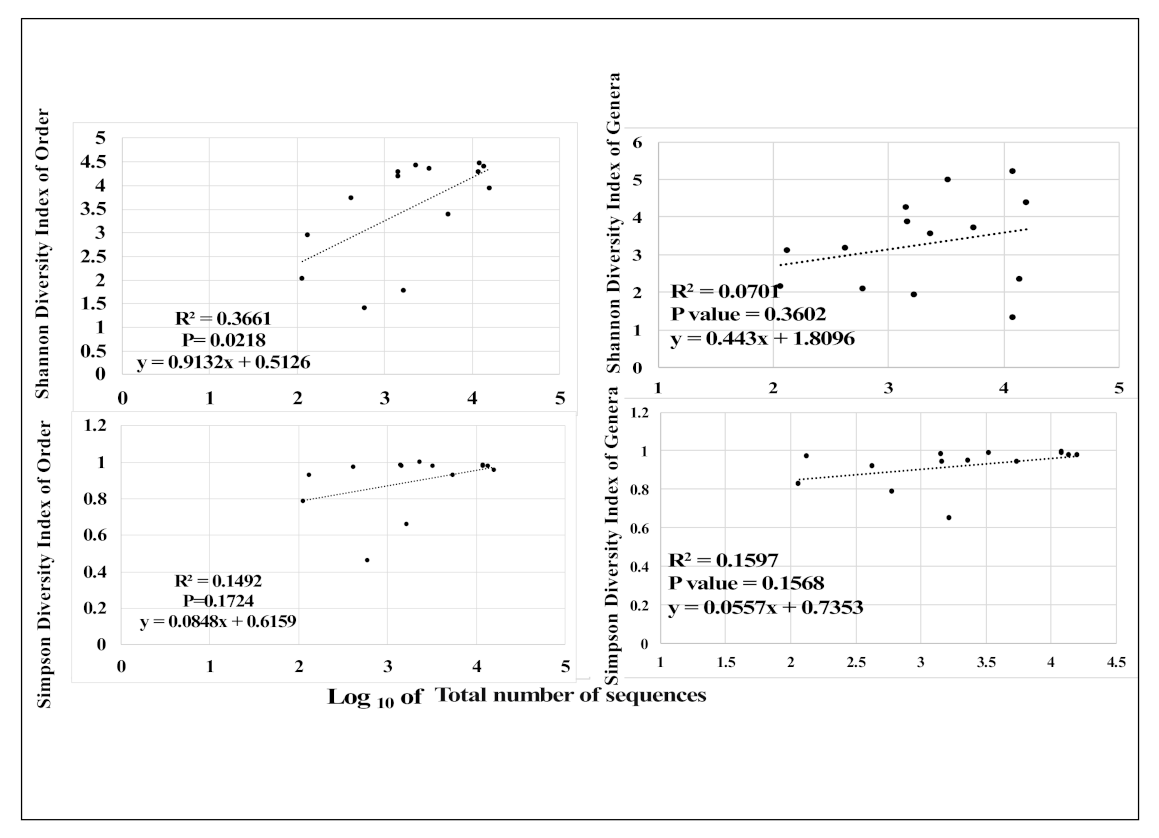
3.3. Sediment Bacterial Bioindicators and Response to Nutrient and Metal Pollution
4. Discussion
4.1. Specific Response Patterns of Ecotoxicological Bioindicators to Nutrient and Metal Pollution Stress
4.2. Bacterial Bioindicators
Specific Response Patterns of Bacterial Bioindicators to Nutrient and Metal Pollution Stress
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genera | Summer 15 | Fall 16 | Summer 17 | Order | Summer 15 | Fall 16 | Summer 17 |
|---|---|---|---|---|---|---|---|
| Bacillus | 60.30 | 3.85 | 0.04 | Rhodospirillales | 3.24 | 1.68 | 4.56 |
| Clostridium | 16.86 | 18.62 | 18.11 | Vibrionales | 9.60 | 0.00 | 0.00 |
| Pseudomonas | 3.19 | 26.24 | 41.67 | Saprospirales | 6.64 | 3.22 | 12.03 |
| Streptococcus | 2.20 | 0.71 | 2.00 | Bacillales | 59.49 | 7.33 | 3.39 |
| Bdellovibrio | 4.70 | 0.00 | 25.90 | Bacteroidales | 5.74 | 2.46 | 15.80 |
| Flavobacterium | 5.97 | 5.29 | 11.84 | Caldilineales | 3.36 | 1.84 | 3.10 |
| Treponema | 3.60 | 2.17 | 12.22 | Clostridiales | 15.24 | 10.91 | 17.97 |
| Thiobacillus | 5.54 | 8.52 | 16.04 | Myxococcales | 8.37 | 2.97 | 9.84 |
| Paenibacillus | 2.81 | 0.00 | 0.00 | Pirellulales | 5.01 | 2.60 | 6.20 |
| Gemmata | 2.86 | 0.00 | 0.00 | Actinomycetales | 20.39 | 5.90 | 12.09 |
| Vibrio | 13.04 | 0.00 | 0.00 | Burkholderiales | 14.25 | 101.86 | 52.31 |
| Pelomonas | 0.00 | 47.62 | 5.31 | Rhizobiales | 9.86 | 6.92 | 17.27 |
| Herbaspirillum | 0.00 | 20.66 | 2.39 | Xanthomonadales | 6.81 | 7.19 | 10.53 |
| Geobacter | 1.89 | 5.61 | 3.50 | Desulfuromonadales | 0.79 | 8.74 | 1.41 |
| Gaiella | 1.41 | 4.35 | 0.00 | Gaiellales | 2.64 | 11.71 | 2.84 |
| Sphingomonas | 0.00 | 11.33 | 10.13 | Rhodocyclales | 2.67 | 4.38 | 9.12 |
| Ralstonia | 0.00 | 3.49 | 0.34 | Pseudomonadales | 3.69 | 9.62 | 18.59 |
| SJA-88 | 0.00 | 2.01 | 11.42 | Rhizobiales | 9.86 | 6.92 | 17.27 |
| Rhodobacter | 0.00 | 0.00 | 5.78 | Sphingomonadales | 2.43 | 6.29 | 15.34 |
| Hyphomicrobium | 1.76 | 0.00 | 4.81 | Bdellovibrionales | 1.69 | 0.68 | 10.97 |
| Crenothrix | 2.90 | 2.17 | 10.26 | Fusobacteriales | 0.62 | 0.00 | 6.12 |
| Methylotenera | 1.50 | 4.74 | 10.29 | Rhodocyclales | 2.67 | 4.38 | 9.12 |
| Flavobacteriales | 3.79 | 2.81 | 6.92 | ||||
| Rhodobacterales | 3.36 | 1.58 | 6.48 | ||||
| Pirellulales | 5.01 | 2.60 | 6.20 | ||||
| Myxococcales | 8.37 | 2.97 | 9.84 |
Appendix B

References
- Rathbun, J.E.; Huellmantel, L.L.; Tracy, M.; Smith, V.E.; Ahlgren, K. Rapid Sediment Assessment: Indicator Analysis and Screening Analysis Approaches. J. Great Lakes Res. 1996, 22, 523–533. [Google Scholar] [CrossRef]
- Steinman, A.D.; Scott, J.; Green, L.; Partridge, C.; Oudsema, M.; Hassett, M.; Kindervater, E.; Rediske, R.R. Persistent organic pollutants, metals, and the bacterial community composition associated with microplastics in Muskego Lake (MI). J. Great Lakes Res. 2020, 46, 1444–1458. [Google Scholar] [CrossRef]
- Weigelhofer, G.; Hein, T.E.; Bondar-Kunze, E. Phosphorus and Nitrogen Dynamics in Riverine Systems: Human Impacts and Management Options. In Riverine Ecosystem Management: Science for Governing towards a Sustainable Future; Schmutz, S., Sendzimir, J., Eds.; Springer International Publishing: Cham, Germany, 2018; pp. 187–202. [Google Scholar]
- Ali, H.; Khan, E.; Ilahi., I. Environmental Chemistry and Ecotoxicology of Hazardous Heavy Metals: Environmental Persistence, Toxicity, and Bioaccumulation. J. Chem. 2019, 6730305. [Google Scholar] [CrossRef] [Green Version]
- Crispell-Synder, I. Executive Summary of WDNR Facilitation Presentation for Pike River Improvements by Mount Pleasant Storm Water Drainage District No 1. Pike River Improvement Project; 1997. Available online: http://www.mtpleasantwi.gov/2325/Pike-River-Improvements-Project (accessed on 19 March 2016).
- Ehlinger, T.; DeThorne, L.; Berner, B. Monitoring of Stream Habitat and Aquatic Biotic Integrity—Pike River North and South Branches, Racine and Kenosha Counties, Wisconsin; University of Wisconsin Milwaukee: Milwaukee, WI, USA, 2009. [Google Scholar]
- Ehlinger, T.; DeThorne, L. Monitoring of Stream Habitat and Aquatic Biotic Integrity—Pike River North and South Branches, Racine and Kenosha Counties, Wisconsin, Interim Report; University of Wisconsin Milwaukee: Milwaukee, WI, USA, 2004. [Google Scholar]
- Tixier, G.; Lafont, M.; Grapentine, L.; Rochfort, Q.; Marsalek, J. Ecological risk assessment of urban stormwater ponds: Literature review and proposal of a new conceptual approach providing ecological quality goals and the associated bioassessment tools. Ecol. Indic. 2011, 11, 1497–1506. [Google Scholar] [CrossRef]
- Marsalek, J.; Rochfort, Q.; Grapentine, L. Aquatic habitat issues in urban stormwater management: Challenges and potential solutions. Ecohydrol. Hydrobiol. 2005, 5, 269. [Google Scholar]
- Moreno-Mateos, D.; Power, M.E.; Comín, F.A.; Yockteng, R. Structural and functional loss in restored wetland ecosystems. PLoS Biol. 2012, 10, e1001247. [Google Scholar] [CrossRef] [Green Version]
- Bodelier, P.; Dedysh, S. Microbiology of wetlands. Front. Microbiol. 2013, 4, 79. [Google Scholar] [CrossRef] [Green Version]
- Knox, A.; Nelson, E.; Halverson, N.; Gladden, J. Long-Term Performance of a Constructed Wetland for Metal Removal. Soil Sediment Contam. 2010, 19, 667–685. [Google Scholar] [CrossRef]
- Banach, A.M.; Kuźniar, A.; Grządziel, J.; Wolińska, A. Azolla filiculoides L. as a source of metal-tolerant microorganisms. PLoS ONE 2020, 15, e0232699. [Google Scholar] [CrossRef] [PubMed]
- Groffman, P.; Hanson, G.; Kiviat, E.; Stevens, G. Variation in Microbial Biomass and Activity in Four Different Wetland Types. Soil Sci. Soc. Am. J. 1996, 60, 622. [Google Scholar] [CrossRef]
- Xie, Y.; Fan, J.; Zhu, W.; Amombo, E.; Lou, Y.; Chen, L.; Fu, J. Effect of heavy metals pollution on soil microbial diversity and Bermudagrass genetic variation. Front. Plant Sci. 2016, 7, 755. [Google Scholar] [CrossRef] [PubMed]
- Sobolev, D.; Begonia, M. Effects of heavy metal contamination upon soil microbes: Lead-induced changes in general and Denitrifying microbial communities as evidenced by molecular markers. Int. J. Environ. Res. Public Health 2008, 5, 450–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Zhao, X.; Chao, L.; Zhang, W.; You, T.; Zhang, J. Diversity change of microbial communities responding to zinc and arsenic pollution in a river of northeastern china. J. Zhejiang Univ. Sci. B 2014, 15, 670–680. [Google Scholar] [CrossRef] [Green Version]
- Müller, A.K.; Westergaard, K.; Christensen, S.; Sørensen, S.J. The effect of long-term mercury pollution on the soil microbial community. FEMS Microbiol. Ecol. 2001, 36, 11–19. [Google Scholar] [CrossRef]
- Frossard, A.; Donhauser, J.; Mestrot, A.; Gygax, S.; Bååth, E.; Frey, B. Long- and short-term effects of mercury pollution on the soil microbiome. Soil Biol. Biochem. 2018, 120, 191–199. [Google Scholar] [CrossRef]
- An, F.; Diao, Z.; Lv, J. Microbial diversity and community structure in agricultural soils suffering from 4 years of Pb contamination. Can. J. Microbiol. 2018, 64, 305–316. [Google Scholar] [CrossRef] [Green Version]
- Gummersheimer, B.S.; Giblin, T. Identification of Lead Resistant Bacteria from a Heavily Contaminated Site. Beta Beta Beta Biol. Soc. Stable JSTOR 2003, 74, 48–54. [Google Scholar]
- Aylagas, E.; Borja, Á.; Tangherlini, M.; Dell’Anno, A.; Corinaldesi, C.; Michell, C.T. A bacterial community-based index to assess the ecological status of estuarine and coastal environments. Mar. Pollut. Bull. 2017, 114, 679–688. [Google Scholar] [CrossRef]
- Olsen, G.J.; Lane, D.J.; Giovannoni, S.J.; Pace, N.R.; Stahl, D.A. Microbial ecology and evolution: A ribosomal RNA approach. Annu. Rev. Microbiol. 1986, 40, 337–365. [Google Scholar] [CrossRef]
- Janda, J.; Abbott, S. 16S rRNA Gene Sequencing for Bacterial Identification in the Diagnostic Laboratory: Pluses, Perils, and Pitfalls. J. Clin. Microbiol. 2007, 45, 2761–2764. [Google Scholar] [CrossRef] [Green Version]
- Pace, N.R.; Stahl, D.A.; Lane, D.J.; Olsen, G.J. The Analysis of Natural Microbial Populations by Ribosomal RNA Sequences. In Advances in Microbial Ecology; Marshall, K.C., Ed.; Springer: Boston, MA, USA, 1986; Volume 9, pp. 1–55. [Google Scholar]
- Winters, A.D.; Marsh, T.L.; Brenden, T.O.; Faisal, M. Analysis of bacterial communities associated with the benthic amphipod Diporeia in the Laurentian great lakes basin. Can. J. Microbiol. 2014, 61, 72–81. [Google Scholar] [CrossRef]
- Bouzat, J.L.; Hoostal, M.J.; Looft, T. Spatial patterns of bacterial community composition within Lake Erie sediments. J. Great Lakes Res. 2013, 39, 344–351. [Google Scholar] [CrossRef]
- Shange, R.; Haugabrooks, E.; Ankumah, R.; Ibekwe, A.; Smith, R.; Dowd, S. Assessing the Diversity and Composition of Bacterial Communities across a Wetland. Transition, Upland Gradient in Macon County Alabama. Diversity 2013, 5, 461–478. [Google Scholar] [CrossRef] [Green Version]
- Bucci, J.P.; Szempruch, A.J.; Caldwell, J.M.; Ellis, J.C.; Levine, J.F. Seasonal changes in microbial community structure in freshwater stream sediment in a North Carolina river basin. Diversity 2014, 6, 18–32. [Google Scholar] [CrossRef] [Green Version]
- Parmar, T.K.; Rawtani, D.; Agrawal, Y.K. Bioindicators: The natural indicator of environmental pollution. Front. Life Sci. 2016, 9, 110–118. [Google Scholar] [CrossRef] [Green Version]
- Microbiotest Inc. Standard Operating Procedure. Phytotoxkit. Seed Germination and Early Growth Microbiotest with Higher Plants. 2019. Available online: https://www.microbiotests.com/wp-content/uploads/2019/05/Phytotoxicity-test_Phytotoxkit-solid-samples_Standard-Operating-Procedure.pdf (accessed on 10 January 2019).
- Czerniawska-Kusza, I.; Ciesielczuk, T.; Kusza, G.; Cichoń, A. Comparison of the Phytotoxkit microbiotest and chemical variables for toxicity evaluation of sediments. Environ. Toxicol. 2006, 21, 367–372. [Google Scholar] [CrossRef]
- Czerniawska-Kusza, I.; Kusza, G. The potential of the Phytotoxkit microbiotest for hazard evaluation of sediments in eutrophic freshwater ecosystems. Environ. Monit. Assess. 2011, 179, 113–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Persoone, G.; Vangheluwe, M. Toxicity determination of the sediments of the river Seine in France by application of a battery of microbiotests. In New Microbiotests for Routine Toxicity Screening and Biomonitoring; Kluwer Academic: New York, NY, USA, 2000; pp. 427–436. [Google Scholar]
- Ghosh Roy, S.; Ehlinger, T.J.; Jablonski, M.R. Effect of metals on growth inhibitions of Sinapis, Sorghum and Lepidium in textile dye waste contaminated soil, India. JEPEG 2019, 20, 608–619. [Google Scholar]
- Murray, K.S.; Rogers, D.T.; Kaufman, M.M. Heavy metals in an urban watershed in southeastern Michigan. J. Environ. Qual. 2004, 33, 163–172. [Google Scholar] [CrossRef]
- Hobbie, S.E.; Finlay, J.C.; Janke, B.D.; Nidzgorski, D.A.; Millet, D.B.; Baker, L.A. Contrasting nitrogen and phosphorus budgets in urban watersheds and implications for managing urban water pollution. Proc. Natl. Acad. Sci. USA 2017, 114, 4177–4182. [Google Scholar] [CrossRef] [Green Version]
- SEWRPC, Racine County Mapbook, 2010. Available online: https://www.sewrpc.org/SEWRPC.htm (accessed on 14 March 2015).
- Ghosh Roy, S.; Ehlinger, T.J. Relationships between land use, predicted pollution loadings and ecotoxicological assays in constructed wetlands. Rom. J. Ecol. Environ. Chem. 2020, 2, 118–129. [Google Scholar]
- Storer, D.A. A simple high sample volume ashing procedure for determination of soil organic matter. Commun. Soil Sci. Plant Anal. 1984, 15, 759–772. [Google Scholar] [CrossRef]
- YSI. YSI a Xylem Brand. 2020. Available online: https://www.ysi.com (accessed on 19 March 2016).
- Hach, Nitrate, Cadmium Reduction Method, 8039. USA. 2019. Available online: https://www.hach.com (accessed on 19 March 2016).
- Hach, Phosphorus, Reactive (Orthophosphate), 8048. USA. 2019. Available online: https://www.hach.com (accessed on 19 March 2016).
- Baranowski, R.; Rybak, A.; Baranowska, I. Speciation Analysis of Elements in Soil Samples by XRF. Pol. J. Environ. Stud. 2002, 11, 473–482. [Google Scholar]
- Kenna, T.C.; Nitsche, F.O.; Herron, M.M.; Mailloux, B.J.; Peteet, D.; Sritrairat, S.; Sands, E.; Baumgarten, J. Evaluation and calibration of a Field Portable X-Ray Fluorescence spectrometer for quantitative analysis of siliciclastic soils and sediments. J. Anal. At. Spectrom. 2011, 26, 395–405. [Google Scholar] [CrossRef]
- Díaz Rizo, O.; Gelen Rudnikas, A.; Lavin Pérez, R.D.; Arencibia Caraballo, G.; Alessandro Rodríguez, K.D. XRF analysis of sediments from Nuevitas Bay (Cuba): Assessment of current heavy metal contamination. Nucleus 2014, 55, 11–14. [Google Scholar]
- Bruker. 2017. Available online: https://www.bruker.com/events/users-meetings/microct/2017.html (accessed on 9 February 2018).
- EPA. Field Portable X-ray Fluorescence Spectrometry for the Determination of Elemental Concentrations in Soil and Sediment. 2007. Available online: https://www.epa.gov/sites/production/files/2015-12/documents/6200.pdf (accessed on 24 April 2018).
- DiScenza, D.J.; Keimowitz, A.R.; Fitzgerald, N. Calibration and Evaluation of an X-ray Fluorescence Method for the Determination of Lead and Arsenic in Soils. J. Environ. Anal. Chem. 2014, 1, 1–3. [Google Scholar] [CrossRef]
- NIST. National Institute of Standards and Technology. 2013. Available online: https://www.nist.gov (accessed on 19 March 2018).
- Zięba-Palus, J.; Kunicki, M. Application of the micro-fTIR spectroscopy, Raman spectroscopy and XRF method examination of inks. Forensic Sci. Int. 2006, 158, 164–172. [Google Scholar] [CrossRef] [PubMed]
- MP Biomedicals, Fast DNA TM Spin Kit. 2017. Available online: https://www.mpbio.com (accessed on 16 March 2016).
- Li, J.; Li, B.; Zhou, Y.; Xu, J.; Zhao, J. A rapid DNA extraction method for PCR amplification from wetland soils. Lett. Appl. Microbiol. 2011, 52, 626–633. [Google Scholar] [CrossRef] [PubMed]
- Burbach, K.; Seifert, J.; Pieper, D.H.; Camarinha-Silva, A. Evaluation of DNA extraction kits and phylogenetic diversity of the porcine gastrointestinal tract based on Illumina sequencing of two hypervariable regions. Microbiologyopen 2016, 5, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.T.; Peng, X.; Deng, G.H.; Sheng, H.F.; Wang, Y.; Zhou, H.W.; Tam, N.F.Y. Illumina Sequencing of 16S rRNA Tag Revealed Spatial Variations of Bacterial Communities in a Mangrove Wetland. Environ. Microbiol. 2013, 66, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokal, R.; Rohlf, J. Biometry, 2nd ed.; W.H. Freeman and Company: San Fransisco, CA, USA, 1981. [Google Scholar]
- SAS. JMP®, Version <14>; SAS Institute Inc.: Cary, NC, USA, 2019; Available online: https://www.jmp.com/en_us/home.html (accessed on 10 January 2015).
- Manikandan, R.; Sahi, S.V.; Venkatachalam, P. Impact assessment of mercury accumulation and biochemical and molecular response of Mentha arvensis: A potential hyperaccumulator plant. Sci. World J. 2015, 2015, 715217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heidenreich, B.; Mayer, K.; Sandermann, H.J.; Ernst, D. Mercury-induced genes in Arabidopsis thaliana: Identification of induced genes upon long-term mercuric ion exposure. Plant Cell Environ. 2001, 24, 1227–1234. [Google Scholar] [CrossRef]
- Baker, A.J.M.; McGrath, S.P.; Reeves, R.D.; Smith, J.A.C. Metal hyperaccumulator plants: A review of the ecology and physiology of biochemical resource for phytoremediation of metal-polluted soils. In Phytoremediation Contaminated Soil Water; CRC Press: Boca Raton, FL, USA, 1999; pp. 85–107. [Google Scholar]
- Liu, C.W.; Sung, Y.; Chen, B.C.; Lai, H.Y. Effects of nitrogen fertilizers on the growth and nitrate content of lettuce (Lactuca sativa L.). Int. J. Environ. Res. Public Health 2014, 11, 4427–4440. [Google Scholar] [CrossRef] [Green Version]
- Pang, J.; Ryan, M.H.; Tibbett, M.; Cawthray, G.R.; Siddique, K.H.M.; Bolland, M.D.A.; Denton, M.D.; Lambers, H. Variation in morphological and physiological parameters in herbaceous perennial legumes in response to phosphorus supply. Plant Soil 2010, 331, 241–255. [Google Scholar] [CrossRef]
- López-Luna, J.; González-Chávez, M.; Esparza-García, F.; Rodríguez-Vázquez, R. Toxicity assessment of soil amended with tannery sludge, trivalent chromium and hexavalent chromium, using wheat, oat and sorghum plants. J. Hazard. Mater. 2009, 163, 829–834. [Google Scholar] [CrossRef]
- Gerik, T.; Bean, B.; Vanderlip, R. Sorghum Growth and Development; Texas A&M University System: College Station, TX, USA, 2010; Available online: http://glasscock.agrilife.org/files/2015/05/Sorghum-Growth-and-Development.pdf (accessed on 7 July 2016).
- Zhang, D.; Luo, J.; Lee, Z.; Maspolim, Y.; Gersberg, R.; Liu, Y. Characterization of bacterial communities in wetland mesocosms receiving pharmaceutical-enriched wastewater. Ecol. Eng. 2016, 90, 215–224. [Google Scholar] [CrossRef]
- Calheiros, C.S.C.; Teixeira, A.; Pires, C.; Franco, A.R.; Duque, A.F.; Crispim, L.F.C.; Moura, S.C.; Castro, P.M.L. Bacterial community dynamics in horizontal flow constructed wetlands with different plants for high salinity industrial wastewater polishing. Water Res. 2010, 44, 5032–5038. [Google Scholar] [CrossRef] [Green Version]
- Ligi, T.; Oopkaup, K.; Truu, M.; Preem, J.K.; Nõlvak, H.; Mitsch, W.J.; Truu, J. Characterization of bacterial communities in soil and sediment of a created riverine wetland complex using high-throughput 16SrRNA amplicon sequencing. Ecol. Eng. 2014, 72, 56–66. [Google Scholar] [CrossRef]
- Zhang, M.J.; Huang, F.K.; Wang, G.Y.; Liu, X.Y.; Wen, J.K.; Zhang, X.S.; Huang, Y.S.; Xia, Y. Geographic distribution of cadmium and its interaction with the microbial community in the Longjiang River: Risk evaluation after a shocking pollution accident. Sci. Rep. 2017, 7, 227. [Google Scholar] [CrossRef]
- Wang, Y.; Sheng, H.F.; He, Y.; Wu, J.Y.; Jiang, Y.X.; Tam, N.F.Y.; Zhou, H.W. Comparison of the levels of bacterial diversity in freshwater, intertidal wetland, and marine sediments by using millions of illumina tags. Appl. Environ. Microbiol. 2012, 78, 8264–8271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, X.; Peng, X.J.; Jin, B.S.; Xiao, M.; Chen, J.K.; Li, B.; Fang, C.M.; Nie, M. Spatial distribution of bacterial communities driven by multiple environmental factors in a beach wetland of the largest freshwater lake in China. Front. Microbiol. 2015, 6, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoeva, M.K.; Aris-Brosou, S.; Chételat, J.; Hintelmann, H.; Pelletier, P.; Poulain, A.J. Microbial community structure in lake and Wetland sediments from a high arctic polar desert revealed by targeted Transcriptomics. PLoS ONE 2014, e89531. [Google Scholar] [CrossRef] [Green Version]
- Hughes, J.; Bohannan, B. Application of ecological diversity statistics in microbial ecology. In Molecular Microbial Ecology Manual, 2nd ed.; Springer: Dordrecht, The Netherlands, 2004; pp. 1321–1344. [Google Scholar]
- Begon, M.; Harper, J.L.; Townsend, C.R.; Begon, M. Ecology: Individuals, Populations, and Communities, 3rd ed.; Blackwell Science: Oxford, UK, 1996. [Google Scholar]
- Karr, J.R.; Chu, E.W. Restoring Life in Running Waters; Island Press: Washington, DC, USA; Covelo, CA, USA, 1999. [Google Scholar]
- Magurran, A. Measuring Biological Diversity; Blackwell Publishing: Malden, MA, USA, 2004. [Google Scholar]
- Faoro, H.; Alves, A.C.; Souza, E.M.; Rigo, L.U.; Cruz, L.M.; Al-Janabi, S.M.; Monteiro, R.A.; Baura, V.A.; Pedrosa, F.O. Influence of soil characteristics on the diversity of bacteria in the southern brazilian atlantic forest. Appl. Environ. Microbiol. 2010, 76, 4744–4749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Lv, X.; Ma, B.; Qu, F.; Ning, K.; Wu, H. A meta-analysis of the bacterial and archaeal diversity observed in wetland soils. Salt Marshes Ecosyst. Veg. Restor. Strateg. 2014, 2014, 13–32. [Google Scholar]
- USEPA. The Sources and Solutions: Agriculture. 2016. Available online: https://www.epa.gov/nutrientpollution/sources-and-solutions-agriculture (accessed on 7 July 2016).
- Ward, M.H. Too much of a good thing? Nitrate from nitrogen fertilizers and cancer. Rev. Environ. Health 2009, 24, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Trajanovska, S.; Britz, M.L.; Bhave, M. Detection of heavy metal ion resistance genes in Gram-positive and Gram-negative bacteria isolated from a lead contaminated site. Biodegradation 1997, 8, 113–124. [Google Scholar] [CrossRef]
- Gadd, G.M. Heavy Metal Pollutants: Environments and Biotechnological Aspects, Encycloped; Academic Press Inc.: San Diego, CA, USA, 1992. [Google Scholar]
- Chen, B.; Liu, J.; Wang, Z.; Dong, L.; Fan, J.; Qu, J. Remediation of Pb-Resistant Bacteria to Pb Polluted Soil. J. Environ. Prot. 2011, 2, 30–141. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Huang, Z.; He, L.; Sheng, X. Assessment of bacterial communities and characterization of lead-resistant bacteria in the rhizosphere soils of metal-tolerant Chenopodium ambrosioides grown on lead-zinc mine tailings. Chemosphere 2012, 87, 1171–1178. [Google Scholar] [CrossRef] [PubMed]
- Jarosławiecka, A.; Piotrowska-Seget, Z. Lead resistance in micro-organisms. Microbiology 2014, 160, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Jebara, S.H.; Abdelkerim, S.; Fatnassi, I.C.; Chiboub, M.; Saadani, O.; Jebara, M. Identification of effective Pb resistant bacteria isolated from Lens culinaris growing in lead contaminated soils. J. Basic Microbiol. 2015, 55, 346–353. [Google Scholar] [CrossRef]
- Figueiredo, N.L.L.; Canário, J.; Duarte, A.; Serralheiro, M.L.; Carvalho, C. Isolation and characterization of mercury-resistant bacteria from sediments of tagus estuary (Portugal): Implications for environmental and human health risk assessment. J. Toxicol. Environ. Health Part A Curr. Issues 2014, 77, 155–168. [Google Scholar] [CrossRef]
- Naguib, M.; Khairalla, A.; El-Gendy, A.; Elkhatib, W. Isolation and Characterization of mercury-resistant bacteria from wastewater sources in Egypt. Can. J. Microbiol. 2019, 65, 308–321. [Google Scholar] [CrossRef]
- Irawati, W.; Patricia; Soraya, Y.; Hugo Baskoro, A. A Study on Mercury-Resistant Bacteria Isolated from a Gold Mine in Pongkor Village, Bogor, Indonesia. HAYATI J. Biosci. 2012, 19, 197–200. [Google Scholar] [CrossRef] [Green Version]
- Kowalczyk, A.; Wilińska, M.; Chyc, M.; Bojko, M.; Latowski, D. Isolation, screening and identification of mercury resistant bacteria from mercury contaminated soil. E3S Web Conf. 2016, 10, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Alia, N.; Sardar, K.; Said, M.; Salma, K.; Sadia, A.; Sadaf, S.; Toqeer, A.; Miklas, S. Toxicity and bioaccumulation of heavy metals in spinach (Spinacia oleracea) grown in a controlled environment. Int. J. Environ. Res. Public Health 2015, 12, 7400–7416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messer, T.; Burchell, M.; Birgand, F.; Broome, S.; Chescheir, G. Nitrate removal potential of restored wetlands loaded with agricultural drainage water: A mesocosm scale experimental approach. Ecol. Eng. 2017, 106, 541–554. [Google Scholar] [CrossRef]









| A. Landcover (Percent in Watershed) | ||||||
|---|---|---|---|---|---|---|
| Wetland Site | Watershed Area (ha) | Percent | Percent | Percent | Percent | Percent |
| Residential | Commercial | Industrial | Agricultural | Undeveloped | ||
| 1 | 104.45 | 11 | 15.1 | 12.1 | 61.6 | 0 |
| 2 | 334.18 | 42.3 | 0 | 0 | 57.5 | 0 |
| 3 | 267.46 | 41.8 | 0 | 0 | 58.2 | 0 |
| 4 | 2.88 | 58.9 | 6 | 0 | 35.2 | 0 |
| 5 | 493.72 | 15.7 | 14.2 | 20.8 | 0 | 49.3 |
| 6 | 720 | 0 | 72.2 | 20.2 | 0 | 7.2 |
| B. Water and Sediment Characteristics | ||||||
| Wetland Site | Median Temperature (°C) | Median pH | Median Specific Conductance (mS/cm) | Median Dissolved Oxygen (%) | Median Dissolved Oxygen (mg/L) | Median Organic Matter Percent |
| 1 | 21.5 | 7.6 | 870 | 105.9 | 9.2 | 8.1 |
| 2 | 21.2 | 7.7 | 634 | 101.8 | 8.7 | 13.3 |
| 3 | 20.2 | 7.2 | 701 | 79.2 | 6.9 | 17 |
| 4 | 19.9 | 7.2 | 907 | 48.1 | 3.8 | 8.4 |
| 5 | 21.4 | 7.7 | 974.5 | 88.6 | 7.8 | 3.8 |
| 6 | 21.8 | 7.1 | 1395.5 | 81.5 | 7.1 | 14 |
| Seed Species | Nitrate | Phosphate | Pb | Hg | R2 | |
|---|---|---|---|---|---|---|
| (A) Stem Inhibition | ||||||
| Lepidium | ||||||
| Sinapis | 0.31 (p = 0.0656) | −1.96 (p = 0.0286) | 0.84 | |||
| Sorghum | −1.77 (p = 0.1249) | 1.88 (p = 0.2417) | 998.83 (p = 0.1879) | 4.80 (p = 0.1548) | 0.96 | |
| (B) Root Inhibition | ||||||
| Seed Species | Nitrate | Phosphate | Pb | Hg | R2 | |
| Lepidium | 2.12 (p = 0.0998) | −1559.8 (p = 0.1216) | −6.75 (p = 0.1310) | 0.82 | ||
| Sinapis | −1011 (p = 0.1962) | −2.46 (p = 0.1196) | 0.55 | |||
| Sorghum | −234.54 (p = 0.0031) | 0.39 |
| As | Zn | Hg | Pb | R2 | |
|---|---|---|---|---|---|
| (A) Order Shannon | 3.76 (p = 0.0084) | 1794.42 (p = 0.00169) | 0.33 | ||
| diversity index | |||||
| (B) Order Simpson | 0 | ||||
| diversity index | |||||
| As | Zn | Hg | Pb | R2 | |
| (D) Genera Shannon diversity index | 5.53 (p = 0.00022) | 11.0024 (p = 0.00001) | −1845.24 (p = 0.0066) | 0.52 | |
| (E) Genera Simpson diversity index | 0.36 (p = 0.01618) | 161.35 (p = 0.00595) | 0.27 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghosh Roy, S.; Wimpee, C.F.; McGuire, S.A.; Ehlinger, T.J. Effects of Land Use and Pollution Loadings on Ecotoxicological Assays and Bacterial Taxonomical Diversity in Constructed Wetlands. Diversity 2021, 13, 149. https://doi.org/10.3390/d13040149
Ghosh Roy S, Wimpee CF, McGuire SA, Ehlinger TJ. Effects of Land Use and Pollution Loadings on Ecotoxicological Assays and Bacterial Taxonomical Diversity in Constructed Wetlands. Diversity. 2021; 13(4):149. https://doi.org/10.3390/d13040149
Chicago/Turabian StyleGhosh Roy, Subhomita, Charles F. Wimpee, Stephen A. McGuire, and Timothy J. Ehlinger. 2021. "Effects of Land Use and Pollution Loadings on Ecotoxicological Assays and Bacterial Taxonomical Diversity in Constructed Wetlands" Diversity 13, no. 4: 149. https://doi.org/10.3390/d13040149