
Characterising the Influence of First-Year Wheat Cultivar on Pseudomonas Selection and Function in a Take-All Infected Field
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Bacterial Isolation
2.2. Growth Conditions and Media
2.3. In Vitro Gt Inhibition Studies
2.4. DNA Extraction
2.5. PCR and DNA Sequencing
2.6. Sequence Analysis
2.7. Seed Sterilisation
2.8. In Planta Assays
2.9. In Planta Gt Antagonistic Assay
2.10. Statistical Analysis
3. Results
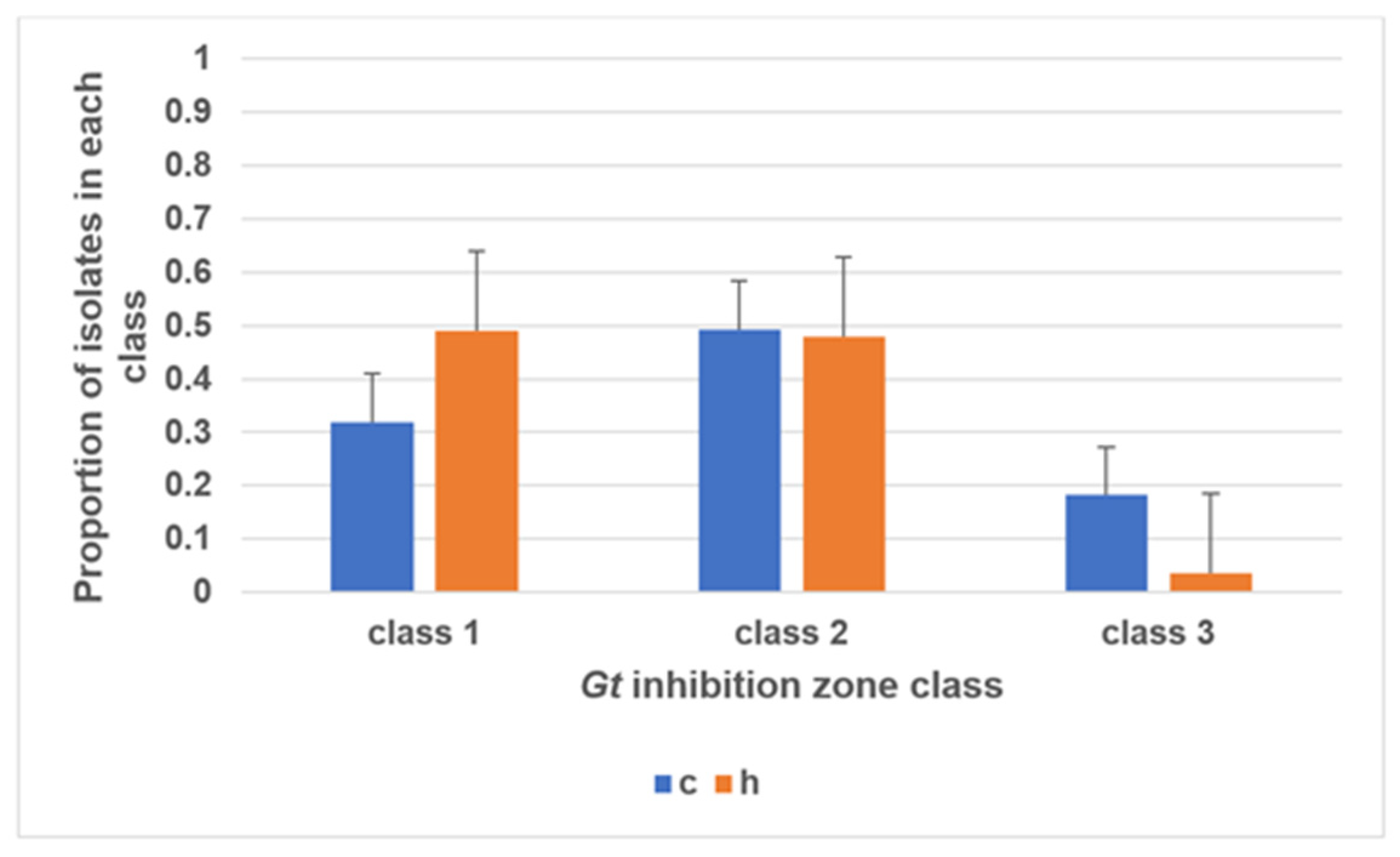
3.1. Screening of Isolates for Gt Antagonism and Classification of Potency
3.2. Biomarker Screening to Differentiate Antagonistic Bacteria
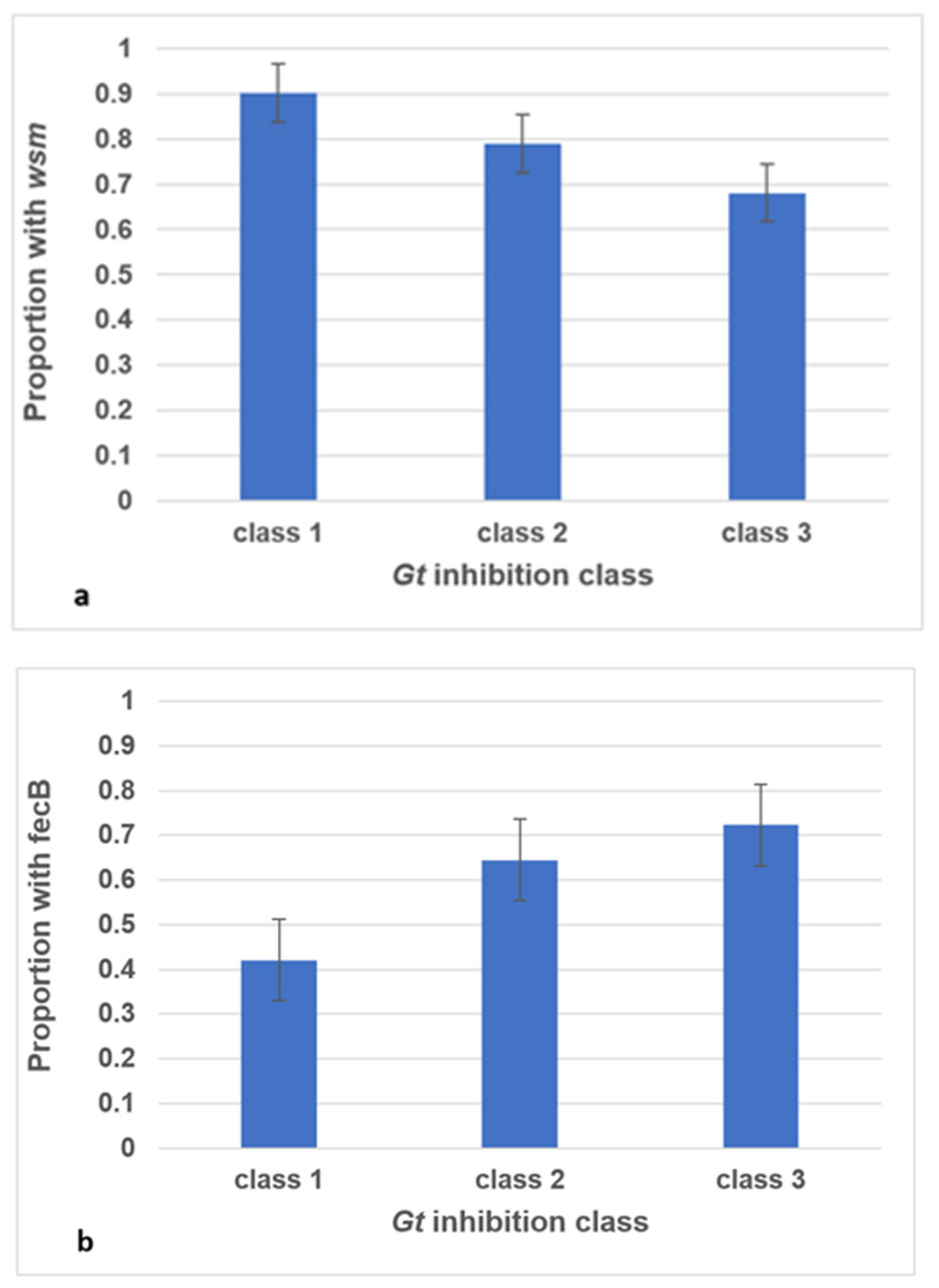
3.3. fecB and wsm Incidence Correlate with Fungal Inhibition
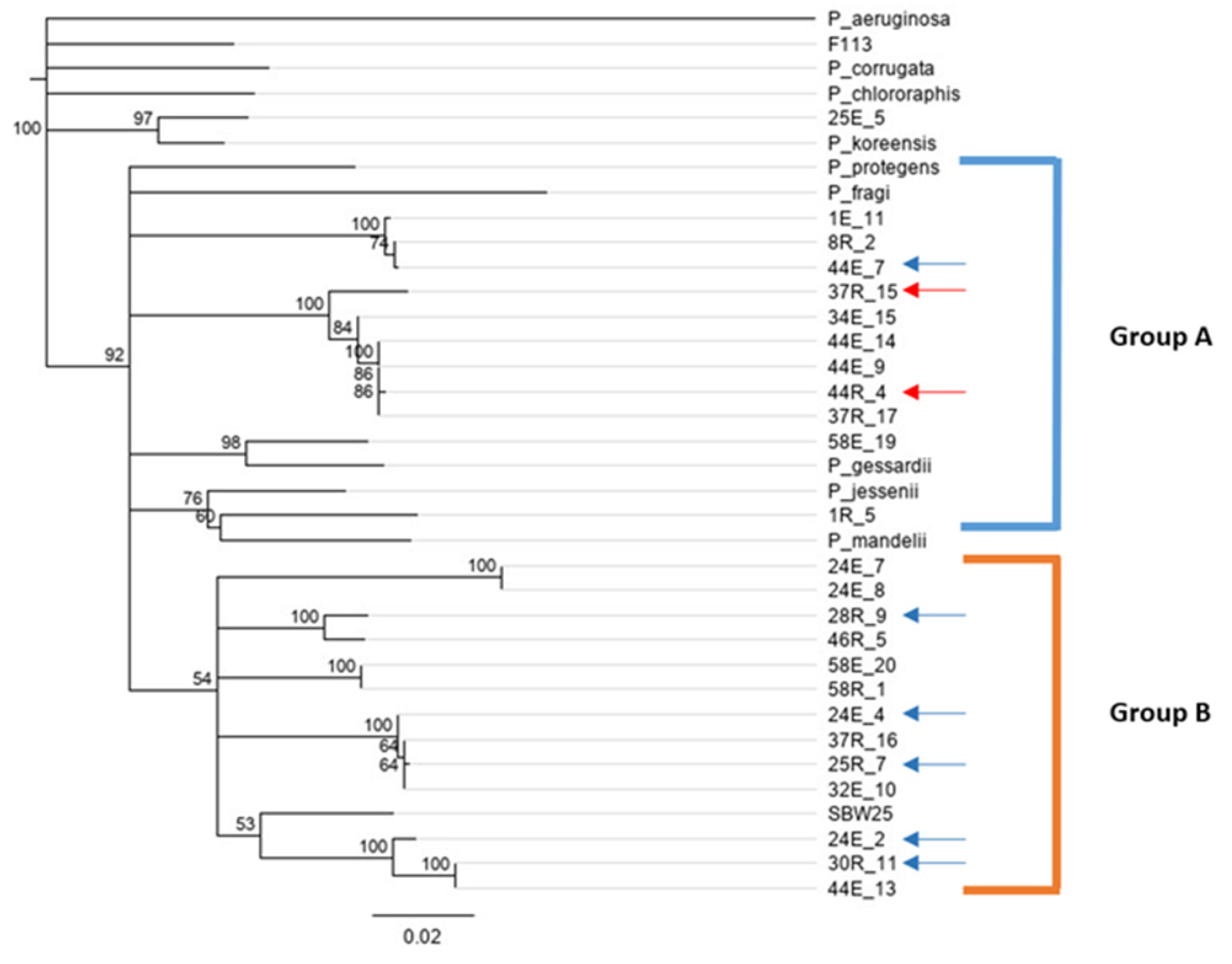
3.4. gyrB Phylogeny
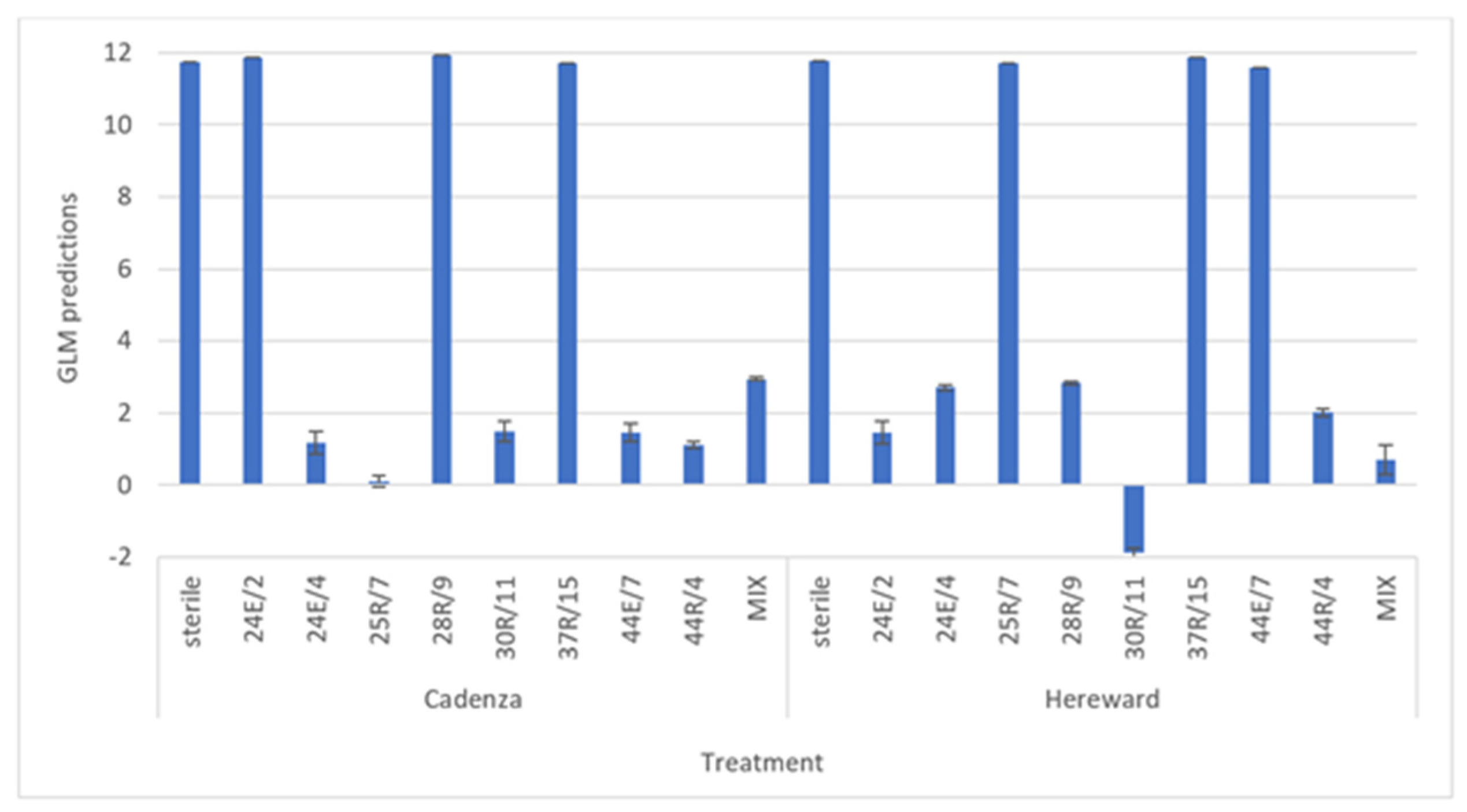
3.5. In Planta Take-All Disease Control Assays
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jenkyn, J.F.; Gutteridge, R.J.; White, R.P. Effects of break crops, and of wheat volunteers growing in break crops or in set-aside or conservation covers, all following crops of winter wheat, on the development of take-all (Gaeumannomyces graminis var. tritici) in succeeding crops of winter wheat. Ann. Appl. Biol. 2014, 165, 340–363. [Google Scholar] [CrossRef]
- McMillan, V.E.; Gutteridge, R.J.; Hammond-Kosack, K.E. Identifying variation in resistance to the take-all fungus, Gaeumannomyces graminis var. tritici, between different ancestral and modern wheat species. BMC Plant Biol. 2014, 14, 212. [Google Scholar] [CrossRef]
- Cook, J.R. Take-all of wheat. Physiol. Mol. Plant Pathol. 2003, 62, 73–86. [Google Scholar] [CrossRef]
- Kwak, Y.-S.; Weller, D.M. Take-all of wheat and natural disease suppression: A review. Plant Pathol. J. 2013, 29, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.-M.; Wen, S.-S.; Mavrodi, D.V.; Mavrodi, O.V.; von Wettstein, D.; Thomashow, L.S.; Guo, J.-H.; Weller, D.M. Biological control of wheat root diseases by the CLP-producing strain Pseudomonas fluorescens HC1-07. Phytopathology 2014, 104, 248–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardener, B.B.M.; Weller, D.M. Changes in populations of rhizosphere bacteria associated with take-all disease of wheat. Appl. Environ. Microbiol. 2001, 67, 4414–4425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Qiao, H.; Huang, L.; Buchenauer, H.; Han, Q.; Kang, Z.; Gong, Y. Biological control of take-all in wheat by endophytic Bacillus subtilis E1R-j and potential mode of action. Biol. Control 2009, 49, 277–285. [Google Scholar] [CrossRef]
- Rainey, P.B. Adaptation of Pseudomonas fluorescens to the plant rhizosphere. Environ. Microbiol. 1999, 1, 243–257. [Google Scholar] [CrossRef]
- Xu, W.; Xu, L.; Deng, X.; Goodwin, P.H.; Xia, M.; Zhang, J.; Wang, Q.; Sun, R.; Pan, Y.; Wu, C.; et al. Biological control of Take-All and growth promotion in wheat by Pseudomonas chlororaphis YB-10. Pathogens 2021, 10, 903. [Google Scholar] [CrossRef]
- Yang, M.-M.; Mavrodi, D.V.; Mavrodi, O.V.; Bonsall, R.F.; Parejko, J.A.; Paulitz, T.C.; Thomashow, L.S.; Yang, H.-T.; Weller, D.M.; Guo, J.-H. Biological control of Take-all by fluorescent Pseudomonas spp. from Chinese wheat fields. Phytopathology 2011, 101, 1481–1491. [Google Scholar]
- Raaijmakers, J.M.; Weller, D.M.; Thomashow, L.S. Frequency of antibiotic-producing Pseudomonas spp. in natural environments. Appl. Environ. Microbiol. 1997, 63, 881–887. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Mavrodi, D.V.; Thomashow, L.S.; Weller, D.M. Differential response of wheat cultivars to Pseudomonas brassicacearum and Take-All decline soil. Phytopathology 2018, 108, 1363–1372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMillan, V.E.; Hammond-Kosack, K.E.; Gutteridge, R.J. Evidence that wheat cultivars differ in their ability to build up inoculum of the take-all fungus, Gaeumannomyces graminis var. tritici, under a first wheat crop. Plant Pathol. 2010, 60, 200–206. [Google Scholar]
- McMillan, V.E.; Canning, G.; Moughan, J.; White, R.P.; Gutteridge, R.J.; Hammond-Kosack, K.E. Exploring the resilience of wheat crops grown in short rotations through minimising the build-up of an important soil-borne fungal pathogen. Sci. Rep. 2018, 8, 9550. [Google Scholar] [CrossRef] [PubMed]
- Mauchline, T.H.; Chedom-Fotso, D.; Chandra, G.; Samuels, T.; Greenaway, N.; Backhaus, A.; McMillan, V.; Canning, G.; Powers, S.J.; Hammond-Kosack, K.E.; et al. An analysis of Pseudomonas genomic diversity in take-all infected wheat fields reveals the lasting impact of wheat cultivars on the soil microbiota. Environ. Microbiol. 2015, 17, 4764–4778. [Google Scholar] [CrossRef] [Green Version]
- King, E.O.; Ward, M.K.; Raney, D.E. Two simple media for the demonstration of pyocyanin and fluorescin. J. Lab. Clin. Med. 1954, 44, 301–307. [Google Scholar] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Yamamoto, S.; Kasai, H.; Arnold, D.L.; Jackson, R.W.; Vivian, A.; Harayama, S. Phylogeny of the genus Pseudomonas: Intrageneric structure reconstructed from the nucleotide sequences of gyrB and rpoD genes The GenBank accession numbers for the sequences determined in this work are: gyrB, D37926, D37297, D86005–D86019 and AB039381–AB039492; rpoD, D86020–D86036 and AB039493–AB039624. Microbiology 2000, 146, 2385–2394. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Weller, D.M. Suppression of Take-all of wheat by seed treatments with fluorescent Pseudomonads. Phytopathology 1983, 73, 463. [Google Scholar] [CrossRef] [Green Version]
- Mavrodi, D.V.; Mavrodi, O.V.; Parejko, J.A.; Bonsall, R.F.; Kwak, Y.-S.; Paulitz, T.C.; Thomashow, L.S.; Weller, D.M. Accumulation of the antibiotic phenazine-1-carboxylic acid in the rhizosphere of dryland cereals. Appl. Environ. Microbiol. 2012, 78, 804–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Mavrodi, D.V.; Yang, M.; Thomashow, L.S.; Mavrodi, O.V.; Kelton, J.; Weller, D.M. Pseudomonas synxantha 2-79 transformed with pyrrolnitrin biosynthesis genes has improved biocontrol activity against soilborne pathogens of wheat and canola. Phytopathology 2020, 110, 1010–1017. [Google Scholar] [CrossRef] [PubMed]
- Mazzola, M.; Gu, Y.-H. Wheat genotype-specific induction of soil microbial communities suppressive to disease incited by Rhizoctonia solani anastomosis group (AG)-5 and AG-8. Phytopathology 2002, 92, 1300–1307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehrabi, Z.; McMillan, V.E.; Clark, I.M.; Canning, G.; Hammond-Kosack, K.E.; Preston, G.; Hirsch, P.R.; Mauchline, T.H. Pseudomonas spp. diversity is negatively associated with suppression of the wheat take-all pathogen. Sci. Rep. 2016, 6, 29905. [Google Scholar] [CrossRef] [Green Version]
- Jousset, A.; Rochat, L.; Lanoue, A.; Bonkowski, M.; Keel, C.; Scheu, S. Plants respond to pathogen infection by enhancing the antifungal gene expression of root-associated bacteria. Mol. Plant-Microbe Interact. 2011, 24, 352–358. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.M.; Wang, D.; Ries, T.R.; Pierson, L.S., III; Pierson, E.A. An upstream sequence modulates phenazine production at the level of transcription and translation in the biological control strain Pseudomonas chlororaphis 30-84. PLoS ONE 2018, 13, e0193063. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus | Primer Name | Primer Sequence | Tm (°C) | Product Size (bp) |
|---|---|---|---|---|
| wsm | wsmF | 5′ GGCAAYGCCGARHTSATCC 3′ | 63 | 603 |
| wsmR | 5′ GCACCARCGSTCYTTRTAYTCRCGGTC 3′ | |||
| fecB | fecBF | 5′ TGATCGTSGCCGACCTCAAYCG 3′ | 70 | 455 |
| fecBR | 5′ CCACARCGGCTGCTTGCTCCAG 3′ | |||
| gyrB | gyrBF | 5′ CAGGAAACAGCTATGACCAYGSNGGNGGNAARTTYRA 3′ | 63 | 888–891 |
| gyrBR | 5′ TGTAAAACGACGGCCAGTGCNGGRTCYTTYTCYTGRCA 3′ | |||
| gyrB | gyrBSF | 5′ CAGGAAACAGCTATGACC 3′ | n/a | n/a |
| gyrBSR | 5′ TGTAAAACGACGGCCAGT 3′ | |||
| phl | Phl2a | 5′ GAGGACGTCGAAGACCACCA 3′ | 67 | 745 |
| Phl2b | 5′ ACCGCAGCATCGTGTATGAG 3′ | |||
| phzCD | PCA2a | 5′ TTGCCAAGCCTCGCTCCAAC 3′ | 67 | 1150 |
| PCA3b | 5′ CCGCGTTGTTCCTCGTTCAT 3′ |
| No. | Isolate * | Planting Combination | Mean Inhibition Zone (cm) | Gt 1:1 Antagonistic Assay |
|---|---|---|---|---|
| 1 | 24E-2 | (C, Xi-19) | 1.24 |  |
| 2 | 24E-4 | (C, Xi-19) | 1.05 |  |
| 3 | 25R-7 | (C, Xi-19) | 1.10 |  |
| 4 | 28R-9 | (H, Xi-19) | 1.08 |  |
| 5 | 30R-11 | (C, Xi-19) | 1.23 |  |
| 6 | 44E-7 | (H, H) | 1.44 |  |
| No | Isolate | Class | Y1 | Y2 | wsm | fecB | Identified as |
|---|---|---|---|---|---|---|---|
| 25E/5 | 1 | C | Xi | 0 | 1 | Pseudomonas R-42020 | |
| 1E/11 | 2 | H | H | 1 | 1 | Pseudomonas fluorescens | |
| 8R/2 | 1 | C | H | 1 | 1 | Pseudomonas orientalis | |
| 44E/7 | 3 | H | H | 1 | 1 | Pseudomonas orientalis | |
| 37R/15 | 1 | H | H | 1 | 0 | Pseudomonas sp. GH1-PS70 | |
| 34E/15 | 2 | H | Xi | 1 | 1 | Pseudomonas sp. RZ109 | |
| 44E/14 | 1 | H | H | 1 | 0 | Pseudomonas marginalis | |
| 44R/4 | 1 | H | H | 1 | 0 | Pseudomonas sp. Ra3 | |
| 44E/9 | 3 | H | H | 1 | 0 | Pseudomonas marginalis | |
| 37R/17 | 1 | H | H | 1 | 0 | Pseudomonas sp. RZ109 | |
| 58E/19 | 1 | C | H | 0 | 0 | Pseudomonas sp. GH1-PS43 | |
| 1R/5 | 1 | H | H | 0 | 0 | Pseudomonas sp. GH1-PS83 | |
| 24E/7 | 3 | C | Xi | 1 | 1 | Pseudomonas poae | |
| 24E/8 | 1 | C | Xi | 1 | 1 | Pseudomonas poae | |
| 28R/9 | 3 | H | Xi | 1 | 1 | Pseudomonas orientalis | |
| 46R/5 | 3 | C | H | 1 | 1 | Pseudomonas orientalis | |
| 58E/20 | 2 | C | H | 1 | 1 | Pseudomonas sp. GH1-PS43 | |
| 58R/1 | 1 | C | H | 1 | 1 | Pseudomonas sp. GH1-PS43 | |
| 24E/4 | 3 | C | Xi | 1 | 1 | Pseudomonas salmonii | |
| 37R/16 | 2 | H | H | 1 | 0 | Pseudomonas fluorescens | |
| 25R/7 | 3 | C | Xi | 1 | 1 | Pseudomonas sp. | |
| 32E/10 | 3 | H | H | 1 | 1 | Pseudomonas poae 36C8 | |
| 24E/2 | 3 | C | Xi | 1 | 1 | Pseudomonas sp. R-41739 | |
| 30R/11 | 3 | C | Xi | 0 | 0 | Pseudomonas orientalis | |
| 44E/13 | 2 | H | H | 1 | 1 | Pseudomonas fluorescens |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al Zadjali, M.; Rabiey, M.; McMillan, V.; Shaw, L.J.; Hammond-Kosack, K.; Malone, J.G.; Mauchline, T.H.; Jackson, R.W. Characterising the Influence of First-Year Wheat Cultivar on Pseudomonas Selection and Function in a Take-All Infected Field. Crops 2023, 3, 195-208. https://doi.org/10.3390/crops3030019
Al Zadjali M, Rabiey M, McMillan V, Shaw LJ, Hammond-Kosack K, Malone JG, Mauchline TH, Jackson RW. Characterising the Influence of First-Year Wheat Cultivar on Pseudomonas Selection and Function in a Take-All Infected Field. Crops. 2023; 3(3):195-208. https://doi.org/10.3390/crops3030019
Chicago/Turabian StyleAl Zadjali, Mahira, Mojgan Rabiey, Vanessa McMillan, Liz J. Shaw, Kim Hammond-Kosack, Jacob G. Malone, Tim H. Mauchline, and Robert W. Jackson. 2023. "Characterising the Influence of First-Year Wheat Cultivar on Pseudomonas Selection and Function in a Take-All Infected Field" Crops 3, no. 3: 195-208. https://doi.org/10.3390/crops3030019