
Six Express Sequence Tag–Simple Sequence Repeat Primers Reveal Genetic Diversity in the Cultivars of Three Zanthoxylum Species

Abstract
:1. Introduction
2. Materials and Methods
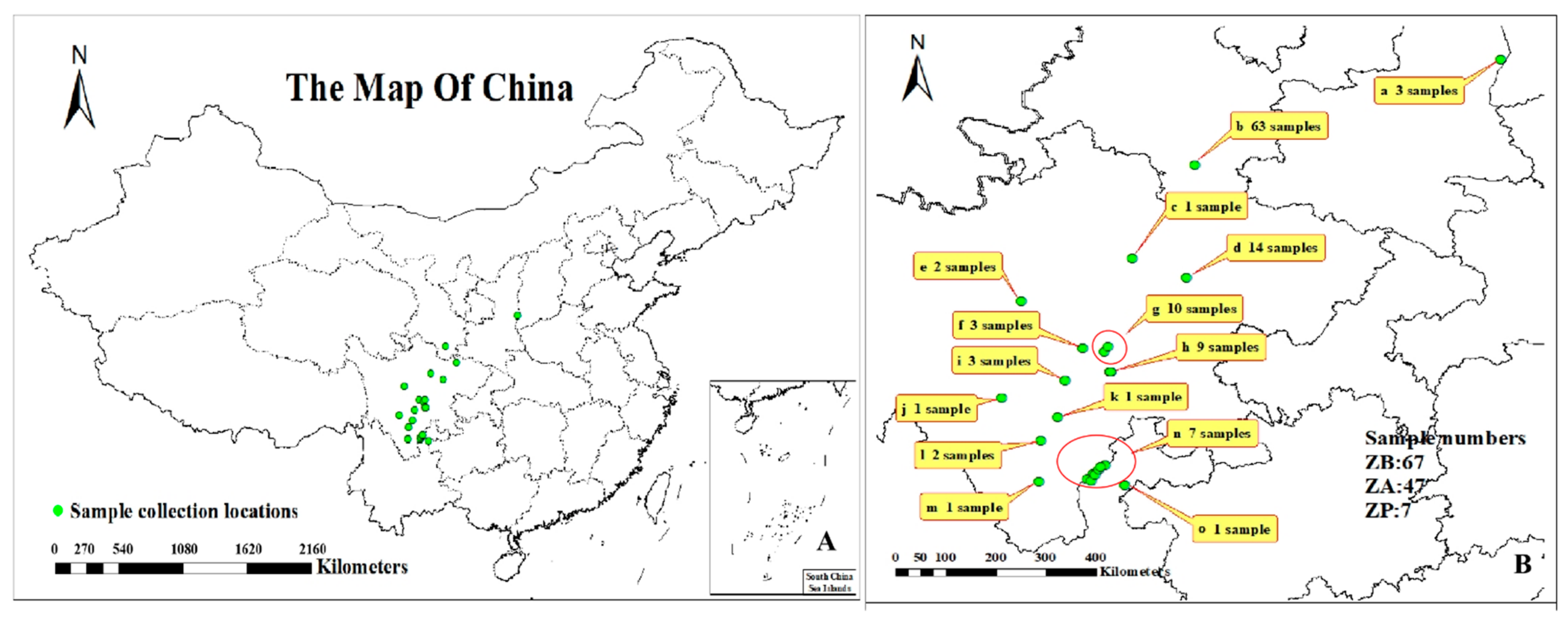
2.1. Materials
2.2. DNA Isolation, PCR Amplification and Capillary Electrophoresis Fluorescence
2.3. Polymorphism Assessment of Primers
2.4. Genetic Distance Calculation, UPGMA and PCA
2.5. Analysis of Molecular Variance (AMOVA) and Genetic Structure Analysis (GSA)
3. Results
3.1. Primers Showed Excellent Specificity and Polymorphism
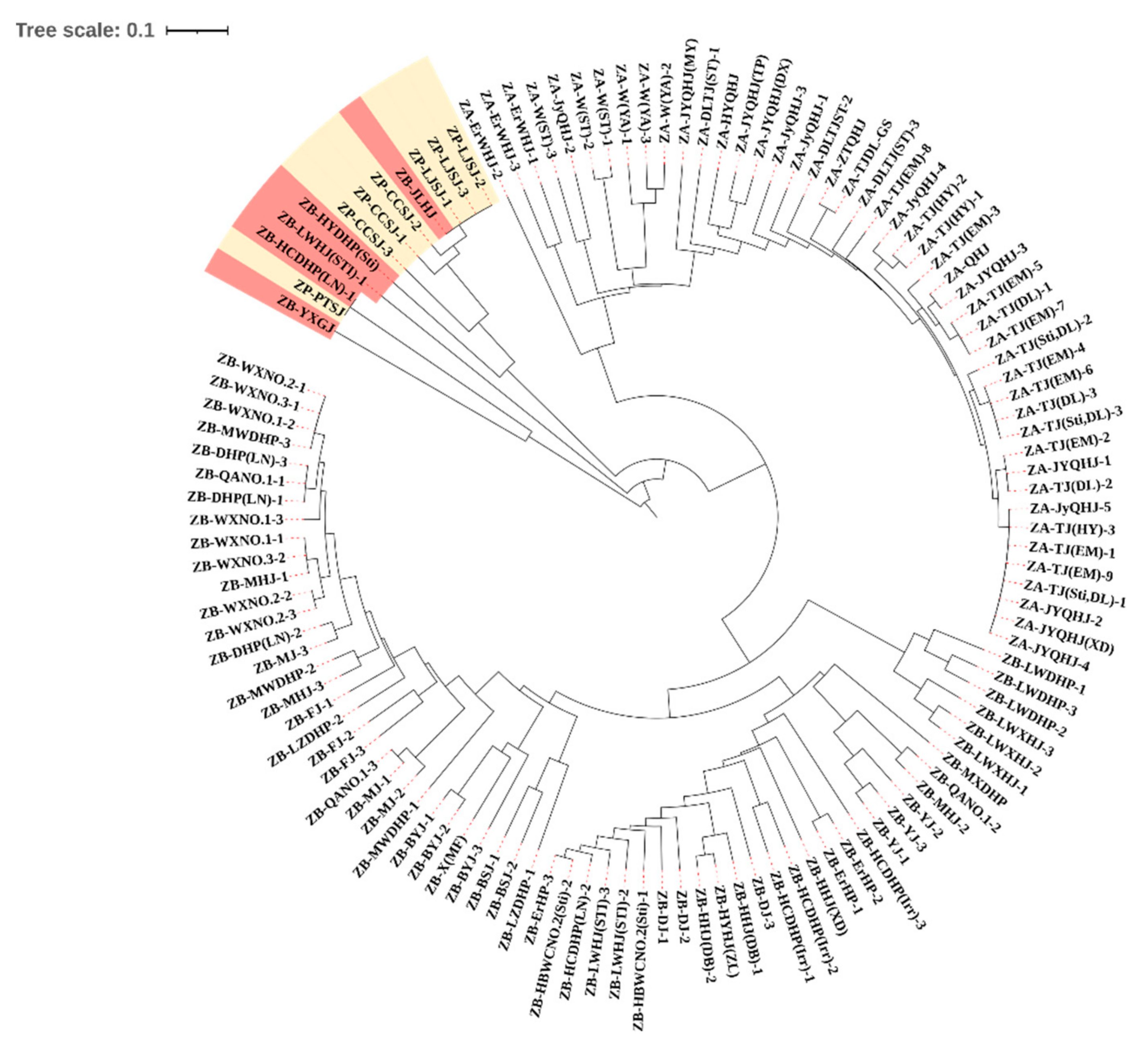
3.2. Most Samples Are Clustered in Correct Clade in the UPGMA Dendrogram
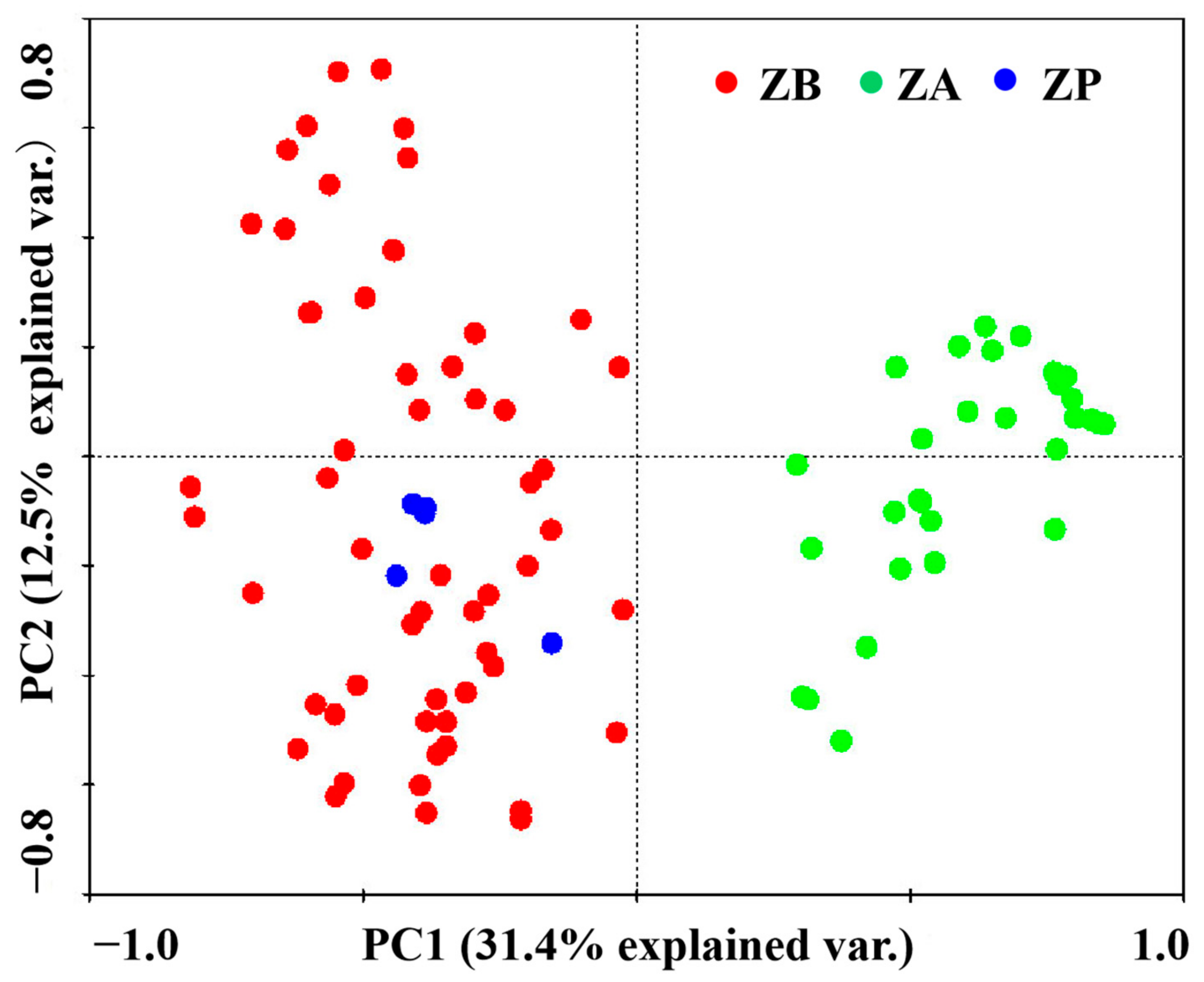
3.3. The Distribution Areas of ZA Were Separated while ZP and ZB Were Well-Overlapped Based on PCA
3.4. The Interspecific and Intraspecific Gene Flows Were Low
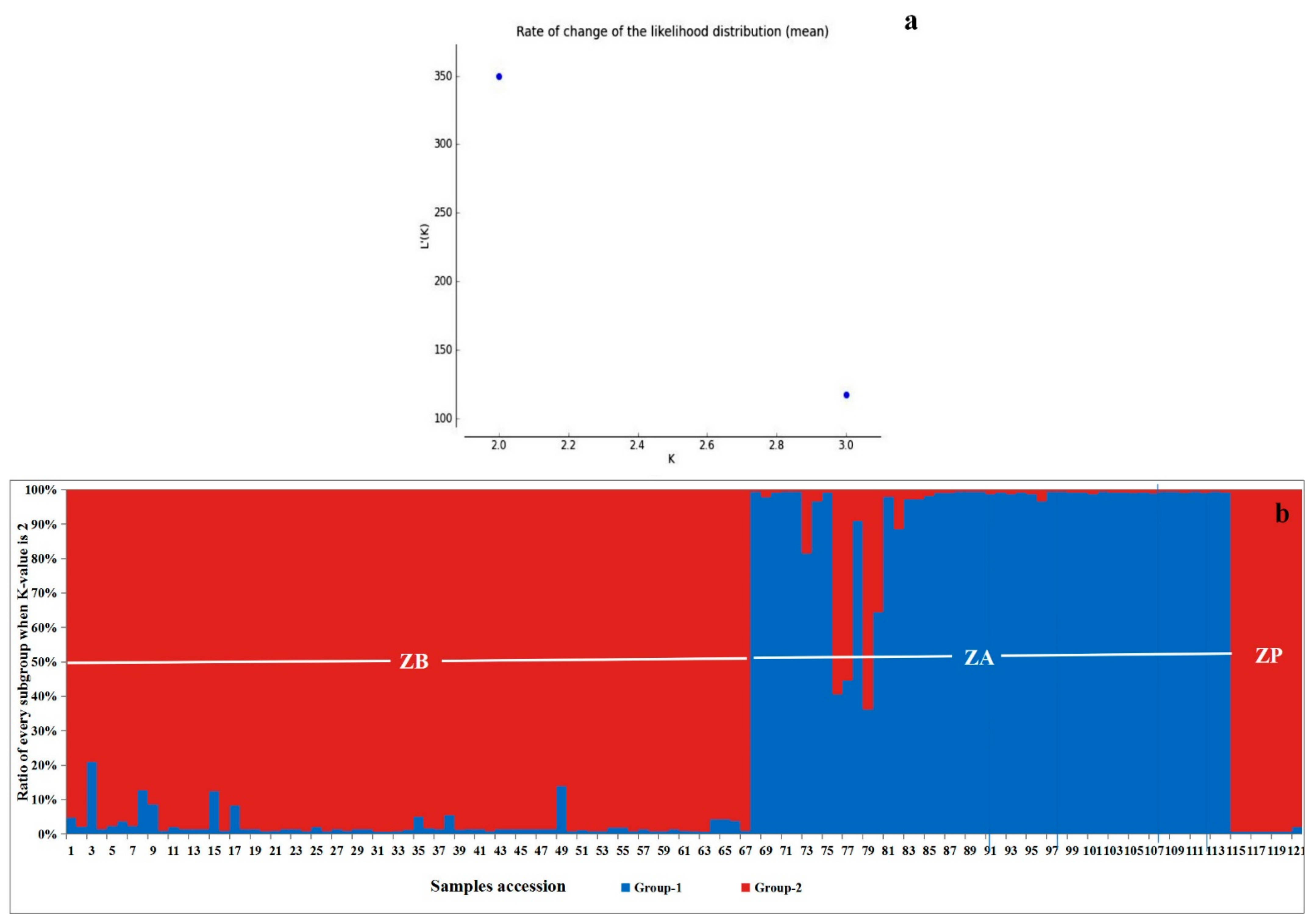
3.5. Three Zanthoxylum Species Divided into Two Genetic Structures
4. Discussion
4.1. EST-SSR Validation
4.2. Genetic Diversity Assessments
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| EST-SSR | Express sequence tag–simple sequence repeat |
| ZB | Zanthoxylum bungeanum |
| ZA | Zanthoxylum armatum |
| ZP | Zanthoxylum piperitum |
| RAPD | Random Amplified Polymorphic DNA |
| ISSR | Inter-Simple Sequence Repeat |
| MCMC | Markov Chain Monte Carlo |
| PCA | Principal Component Analysis |
| UPGMA | Unweighted Pair Group Method with Arithmetic Mean Analysis |
| GSA | Genetic Structure Analysis |
| NR | Non-Redundant Protein Sequence Database |
| COG | Clusters of Orthologous Group |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| GO | Gene Ontology |
References
- Asase, A.; Oppong-Mensah, G. Traditional antimalarial phytotherapy remedies in herbal markets in southern Ghana. J. Ethnopharmacol. 2009, 126, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Massoma, L.; Gasco, M.; Rubio, J.; Yucra, S.; Sock, E.N.; Gonzales, G.F. Effect of the ethanolic extract from Fagara tessmannii on testicular function, sex reproductive organs and hormonal level in adult male rats. J. Androl. 2011, 43, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Phuyal, N.; Jha, P.K.; Raturi, P.P.; Rajbhandary, S. Zanthoxylum armatum DC.: Current knowledge, gaps and opportunities in Nepal. J. Ethnopharmacol. 2019, 229, 326–341. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.X.; Hartley, T.G. Flora of China; Science Press: Beijing, China, 2008; Volume 11. [Google Scholar]
- Nurain, I.O.; Bewaji, C.O.; Johnson, J.S.; Davenport, R.D.; Zhang, Y. Potential of Three Ethnomedicinal Plants as Antisickling Agents. Mol. Pharm. 2017, 14, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Zhang, G.; Jin, L.; Zhang, B.; Yan, D.; Yang, H.; Ye, Z.; Ma, T. Inhibition of PAI-1 activity by toddalolactone as a mechanism for promoting blood circulation and removing stasis by Chinese herb Zanthoxylum nitidum var. tomentosum. Front. Pharm. 2017, 8, 489. [Google Scholar] [CrossRef] [PubMed]
- Akakpo-Akue, J.; Kplé, T.K.M.; Coulibaly, K.; Ahon, G.M.; Fofié, Y.; Yapo-Crezoit, A.; Zirihi, G.N.; Kra, A.K.M. Ethnobotanical study of medicinal plants used against sickle cell anaemi-a in the eastern part of the Côte d’Ivoire. J. Anim. Plant Sci. 2020, 45, 7839–7852. [Google Scholar] [CrossRef]
- Tatiana, K.M.K.; Akakpo-Akue, J.; Yvette, F.; Marcel, A.G.; Julien, G.K.; Mathieu, K.A.K.; Ibrahime, S.; Chiayé, C.A.Y. Sickling cells inhibition and radical scavenging activities of Zanthoxylum leprieurii’s (GUILL) bark extracts: Comparative study. J. Pharm. Res. Int. 2020, 32, 42–50. [Google Scholar] [CrossRef]
- Zhang, H.; Ye, M. Research Status on the Taxonomic and Component of Green Zanthoxylum bungeanum Maxim. North Hortic. 2010, 14, 918–924. [Google Scholar]
- Peterson, S.W. Identification of common Aspergillus species. Mycologist 2002, 17, 128. [Google Scholar] [CrossRef]
- Feng, S.T.; Yang, Z.; Liu, L.; Chen, N.; Hou, Y.; Wei, A.Z. Genetic diversity and relationships of wild and cultivated Zanthoxylum germplasms based on sequence-related amplified polymorphism (SRAP) markers. Genet. Resour. Crop Evol. 2015, 62, 1193–1204. [Google Scholar] [CrossRef]
- Zhu, L.; Chen, X.; Li, J.H.; Jiang, W.D.; Zhong, C.P.; Zhou, J.; Wu, C.J. Textual research on origin of Zanthoxylum armatum and consideration of its definition. Chin. Tradit. Herb. Drugs 2018, 4, 987–992. [Google Scholar] [CrossRef]
- Fei, X.; Shi, Q.; Qi, Y.; Wang, S.; Lei, Y.; Hu, H.; Liu, Y.; Yang, T.; Wei, A.Z. ZbAGL11, a class D MADS-box transcription factor of Zanthoxylum bungeanum, is involved in sporophytic apomixis. Hortic. Res. 2021, 8, 23. [Google Scholar] [CrossRef] [PubMed]
- Fei, X.; Lei, Y.; Qi, Y.; Wang, S.; Hu, H.; Wei, A. Small RNA sequencing provides candidate miRNA-target pairs for revealing the mechanism of apomixis in Zanthoxylum bungeanum. BMC Plant Biol. 2021, 21, 178. [Google Scholar] [CrossRef] [PubMed]
- Kigen, G.; Kipkore, W.; Wanjohi, B.; Haruki, B.; Kemboi, J. Medicinal plants used by traditional healers in Sangurur, Elgeyo Marakwet County, Kenya. Pharmacogn. Res. 2017, 9, 333–347. [Google Scholar] [CrossRef] [PubMed]
- Ameh, S.J.; Tarfa, F.D.; Ebeshi, B.U. Traditional herbal management of sickle cell anemia: Lessons from Nigeria. Anemia 2012, 2012, 607436. [Google Scholar] [CrossRef] [PubMed]
- Andima, M.; Coghi, P.; Yang, L.J.; Wong, V.K.W.; Ngule, C.M.; Heydenreich, M.; Ndakala, A.J.; Yenesew, A.; Derese, S. Antiproliferative activity of secondary metabolites from Zanthoxylum zanthoxyloides Lam: In vitro and in silico studies. Pharmacogn. Commun. 2019, 10, 44–51. [Google Scholar] [CrossRef]
- Tine, Y.; Yang, Y.; Renucci, F.; Costa, J.; Wélé, A.; Paolini, J. LC-MS/MS analysis of flavonoid compounds from Zanthoxylum zanthoxyloides extracts and their antioxidant activities. Nat. Prod. Commun. 2017, 12, 1865–1868. [Google Scholar] [CrossRef]
- Zeng, Q.; Wang, Z.J.; Chen, S.; Wang, H.; Xie, T.Z.; Xu, X.J.; Xiang, M.L.; Chen, Y.C.; Luo, X.D. Phytochemical and anti-MRSA constituents of Zanthoxylum nitidum. Biomed. Pharmacother. 2022, 148, 112758. [Google Scholar] [CrossRef]
- Ogunbolude, Y.; Ibrahim, M.; Elekofehinti, O.O.; Adeniran, A.; Abolaji, A.O.; Rocha, J.B.T.; Kamdem, J.P. Effects of Tapinanthus globiferus and Zanthoxylum zanthoxyloides extracts on human leukocytes in vitro. J. Intercult. Ethnopharmacol. 2014, 3, 167–172. [Google Scholar] [CrossRef]
- Djordjevic, V.B. Free Radicals in Cell Biology. Int. Rev. Cytol. 2004, 237, 57–89. [Google Scholar]
- Tan, B.L.; Norhaizan, M.E.; Liew, W.P.P.; Sulaiman, R.H. Antioxidant and oxidative stress: A mutual interplay in age-related diseases. Front. Pharmacol. 2018, 9, 1162. [Google Scholar] [CrossRef] [PubMed]
- Willcox, J.K.; Ash, S.L.; Catignani, G.L. Antioxidants and prevention of chronic disease. Crit. Rev. Food Sci. Nutr. 2004, 44, 275–295. [Google Scholar] [CrossRef] [PubMed]
- Medhi, K.; Sarmah, D.K.; Deka, M.; Bhau, B.S. High gene flow and genetic diversity in three economically important Zanthoxylum Spp. of Upper Brahmaputra Valley Zone of NE India using molecular markers. Meta Gene 2014, 2, 706–721. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.L.; Park, W.G.; Kwon, O.W.; Hong, S.W. Ribosomal DNA internal transcribed spacer 1, and internal transcribed spacer 2 regions as targets for molecular identification of medically important Zanthoxylum schinifolium. Afr. J. Biotechnol. 2010, 9, 4661–4673. [Google Scholar]
- Feng, S.; Liu, Z.; Hu, Y.; Tian, J.; Yang, T.; Wei, A. Genomic analysis reveals the genetic diversity, population structure, evolutionary history and relationships of Chinese pepper. Hortic. Res. 2020, 7, 158. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.M.; Jo, A.; Jeong, J.H.; Kwon, Y.R.; Kim, H.B. Development and characterization of microsatellite primers for Zanthoxylum schinifolium (Rutaceae). Appl. Plant Sci. 2017, 5, 1600145. [Google Scholar] [CrossRef] [PubMed]
- Su, K.; Sun, J.; Han, J.; Zheng, T.; Sun, B.; Liu, S. Combined morphological and multi-omics analyses to reveal the developmental mechanism of Zanthoxylum bungeanum prickles. Front. Plant Sci. 2022, 13, 950084. [Google Scholar] [CrossRef]
- Tangphatsornruang, S.; Sraphet, S.; Singh, R.; Okogbenin, E.; Triwitayakorn, K. Development of polymorphic markers from expressed sequence tags of Manihot esculenta Crantz. Mol. Ecol. Resour. 2008, 8, 682–685. [Google Scholar] [CrossRef]
- Holton, T.A.; Christopher, J.T.; McClure, L.; Harker, N.; Henry, R.J. Identification and mapping of polymorphic SSR markers from expressed gene sequences of barley and wheat. Mol. Breed. 2002, 9, 63–71. [Google Scholar] [CrossRef]
- Scott, K.D.; Eggler, P.; Seaton, G.; Rossetto, M.; Ablett, E.M.; Lee, L.S.; Henry, R.J. Analysis of SSRs derived from grape ESTs. Theor. Appl. Genet. 2000, 100, 723–726. [Google Scholar] [CrossRef]
- Kantety, R.V.; La Rota, M.; Matthews, D.E.; Sorrells, M.E. Data mining for simple sequence repeats in expressed sequence tags from barley, maize, rice, sorghum and wheat. Plant Mol. Biol. 2002, 48, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.J.; Marek, L.F.; Shoemaker, R.C.; Lark, K.G.; Concibido, V.C.; Delannay, X.; Specht, J.E.; Cregan, P.B. A new integrated genetic linkage map of the soybean. Theor. Appl. Genet. 2004, 109, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.C.; Ping, Z.; Young, A.C.; Shu, H.; Fred, G. Mining and characterizing microsatellites from citrus ESTs. Theor. Appl. Genet. 2006, 112, 1248–1257. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.X.; Wei, A.Z.; Wang, L.H.; Liu, Y.L. Analysis of SSR Loci and Development of Molecular Markers in Zanthoxylum bungeanum Transcriptome. Chin. J. Agric. Biotech. 2018, 26, 1226–1236. [Google Scholar]
- Kim, J.A.; Roy, N.S.; Lee, I.H.; Choi, A.Y.; Choi, B.S.; Yu, Y.S.; Park, N.I.; Park, K.C.; Kim, S.; Yang, H.S.; et al. Genome-wide transcriptome profiling of the medicinal plant, Zanthoxylum planispinum, using a single-molecule direct RNA sequencing approach. Genomics 2019, 111, 973–979. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Zhao, L.; Liu, Z.; Liu, Y.; Yang, T.; Wei, A. De novo transcriptome assembly of Zanthoxylum bungeanum using Illumina sequencing for evolutionary analysis and simple sequence repeat marker development. Sci. Rep. 2017, 7, 16754. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.C.; Xiang, L.; Su, Y.Y.; Yang, L.; Li, H.Y.; Luo, X.M.; Sun, W.; Ye, M. Development of EST-SSR markers of Zanthoxylum species based on RNA-sequencing. J. Northwest A F Univ. 2019, 4, 16–24. [Google Scholar] [CrossRef]
- Guo, W.Z.; Zhao, J.X.; Cao, J.H. Cultivation and management techniques of Zanthoxylum piperitum. North. Hortic 2012, 20, 36–37. [Google Scholar]
- Nei, M. Genetic Distance between Populations. Am. Nat. 1972, 106, 283–292. [Google Scholar] [CrossRef]
- Lewontin, R.C. Testing the Theory of Natural Selection. Nature 1972, 236, 181–182. [Google Scholar] [CrossRef]
- Peakall, R.; Peter, E.S. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of Population Structure Using Multilocus Genotype Data. Genetics 2000, 155, 9197–9201. [Google Scholar] [CrossRef] [PubMed]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Linked loci and correlated allele frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [CrossRef] [PubMed]
- Evanno, G.S.; Regnaut, S.J.; Goudet, J. Detecting the number of clusters of individuals using the software Structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Anuradha, U.; Kadam, U.S.; Priya, C.M.; Karibasappa, G.S. Microsatellite and RAPD analysis of grape (Vitis spp.) accessions and identification of duplicates/misnomers in germplasm collection. Indian J. Hortic. 2010, 67, 8–15. [Google Scholar] [CrossRef]
- Upadhyay, A.; Kadam, U.S.; Chacko, P.; Aher, L.; Karibasappa, G.M. Microsatellite analysis to differentiate clones of Thompson Seedless grapevine. Indian J. Hortic. 2010, 67, 260–263. [Google Scholar] [CrossRef]
- Mir, A.H.; Bhat, M.A.; Fayaz, H.; Wani, A.A.; Dar, S.A.; Maqbool, S.; Yasin, M.; Mir, J.I.; Khan, M.A.; Sofi, P.A.; et al. SSR markers in revealing extent of genetic diversity and phylogenetic relationships among chickpea core collection accessions for Western Himalayas. Mol. Biol. Rep. 2022, 49, 11469–11479. [Google Scholar] [CrossRef]
- Chen, J.; Liu, Q.; Lu, C.; Liu, Q.; Pan, J.; Zhang, J.; Dong, S. Genetic diversity of Prunus armeniaca L. var. ansu Maxim. germplasm revealed by simple sequence repeat (SSR) markers. PLoS ONE 2022, 17, e0269424. [Google Scholar] [CrossRef]
- Kazemeini, F.; Asri, Y.; Mostafavi, G.; Kalvandi, R.; Mehregan, I. RAPD-based evaluation of genetic diversity among populations of the Iranian endemic species Rhabdosciadium aucheri Boiss. (Apiaceae). Mol. Biol. Rep. 2020, 47, 9345–9352. [Google Scholar] [CrossRef]
- Sen, S.; Skaria, R.; Muneer, P.M.A. Genetic diversity analysis in Piper species (Piperaceae) using RAPD markers. Mol. Biotechnol. 2010, 46, 72–79. [Google Scholar] [CrossRef]
- Shahnazari, N.; Noormohammadi, Z.; Sheidai, M.; Koohdar, F. A new insight on genetic diversity of sweet oranges: CAPs-SSR and SSR markers. J. Genet. Eng. Biotechnol. 2022, 20, 105. [Google Scholar] [CrossRef] [PubMed]
- Chavhan, R.L.; Sable, S.; Narwade, A.V.; Hinge, V.R.; Kalbande, B.B.; Mukherjee, A.K.; Chakrabarty, P.K.; Kadam, U.S. Multiplex molecular marker-assisted analysis of significant pathogens of cotton (Gossypium sp.). Biocatal. Agric. Biotechnol. 2023, 47, 102557. [Google Scholar] [CrossRef]
- Hu, C.Y.; Tsai, Y.Z.; Lin, S.F. Development of STS and CAPS markers for variety identification and genetic diversity analysis of tea germplasm in Taiwan. Bot. Stud. 2014, 55, 12. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.J. Evaluation of Antiviral Activity of Zanthoxylum Species against Picornaviruses. Osong Public Health Res. Perspect. 2016, 7, 400–403. [Google Scholar] [CrossRef] [PubMed]
- Alam, F.; Us Saqib, Q.N. Evaluation of Zanthoxylum armatum Roxb for in vitro biological activities. J. Tradit. Chin. Med. 2017, 7, 515–518. [Google Scholar] [CrossRef] [PubMed]
- Varshney, R.K.; Thiel, T.; Stein, N.; Langridge, P.; Graner, A. In silico analysis on frequency and distribution of microsatellites in ESTs of some cereal species. Cell. Mol. Biol. Lett. 2002, 7, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.C.; Liu, X.W.; Li, Y.; Zhao, D.G. Transcriptome Data Assembly and Gene Function Annotation of Zanthoxylum piperitum DC var. inerme Makino. Genom. Appl. Biol. 2016, 35, 1805–1819. [Google Scholar] [CrossRef]
- Eujayl, I.; Sorrells, M.; Baum, M.; Wolters, P.; Powell, W. Assessment of genotypic variation among cultivated durum wheat based on EST-SSRS and genomic SSRs. Euphytica 2001, 119, 39–43. [Google Scholar] [CrossRef]
- Thiel, T.; Michalek, W.; Varshney, R.; Graner, A. Exploiting EST databases for the development and characterization of gene-derived SSR-markers in barley (Hordeum vulgare L.). Theor. Appl. Genet. 2003, 106, 411–422. [Google Scholar] [CrossRef]
- Dong, J.; Guang, Y.Z. Analysis of Microsatellites in Citrus Unigenes. Act. Genet. Sin. 2006, 33, 345–353. [Google Scholar] [CrossRef]
- Zhao, L.L.; Feng, S.J.; Tian, J.Y.; Wei, A.Z.; Yang, T.X. Internal transcribed spacer 2 (ITS2) barcodes: A useful tool for identifying Chinese Zanthoxylum. Appl. Plant. Sci. 2018, 6, e01157. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.L.; Park, W.G.; Kwon, O.W.; Soon, K.H. Molecular identification of Zanthoxylum piperitum by the internal transcribed spacers as targets using newly designed species-specific primers. J. Biotechnol. 2010, 150, 505. [Google Scholar] [CrossRef]
- Wang, C.; Zhao, J.X.; Bi, J.; Wang, C.R.; Guo, W.Z.; Lin, Y. The Techniques of Fruitsetting Improvement of Zanthoxylu piperitum f. inerme Makino. North. Hortic. 2012, 26, 96–99. [Google Scholar] [CrossRef]
- Suprasanna, P.; Jain, S.M.; Ochatt, S.J.; Kulkarni, V.M.; Predieri, S. Applications of In Vitro Techniques in Mutation Breeding of Vegetatively Propagated Crops; CABI International: Wallingford, UK, 2012. [Google Scholar] [CrossRef]
- Beyaz, R.; Yildiz, M. The Use of Gamma Irradiation in Plant Mutation Breeding. In Plant Engineering; InTech: Tokyo, Japan, 2017. [Google Scholar] [CrossRef]
- Ahloowalia, B. Induced mutations-A new paradigm in plant breeding. Euphytica 2001, 118, 167–173. [Google Scholar] [CrossRef]
- Wright, S. Evolution in Mendelian populations. Bull. Math. Biol. 1990, 52, 241–295. [Google Scholar] [CrossRef] [PubMed]
- Ellstrand, N.C.; Roose, M.L. Patterns of genotypic diversity in clonal plant species. Am. J. Bot. 1987, 74, 123–131. [Google Scholar] [CrossRef]
- Buso, G.S.C.; Rangel, P.H.; Ferreira, M.E. Analysis of genetic variability of south American wild rice populations (Oryza glumaepatula) with isozymes and RAPD markers. Mol. Ecol. 1998, 7, 107–118. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | Degrees of Freedom (df) | Statistics (SS) | Mean Square (MS) | Est. Var. | Percentage of Variance Component | p-Value | |
|---|---|---|---|---|---|---|---|
| Among Populations (AP) | Interspecific populations of ZB, ZA and ZP | 2 | 149.543 | 74.771 | 2.224 | 43% | p > 0.001 |
| Intraspecific populations of ZB | 21 | 130.700 | 6.224 | 1.721 | 53% | p > 0.001 | |
| Intraspecific populations of ZA | 9 | 29.320 | 3.258 | 0.523 | 29% | p > 0.001 | |
| Within Populations (WP) | Interspecific populations of ZB, ZA and ZP | 118 | 349.854 | 2.965 | 2.965 | 57% | \ |
| Intraspecific populations of ZB | 38 | 58.333 | 1.535 | 1.535 | 47% | \ | |
| Intraspecific populations of ZA | 29 | 36.706 | 1.266 | 1.266 | 71% | \ | |
| Total | Interspecific populations of ZB, ZA and ZP | 120 | 499.397 | \ | 5.189 | 100% | \ |
| Intraspecific populations of ZB | 59 | 189.033 | \ | 3.256 | 100% | \ | |
| Intraspecific populations of ZA | 38 | 66.026 | \ | 1.789 | 100% | \ | |
| Genetic differentiation coefficient (PhiPT) | Interspecific populations of ZB, ZA and ZP | 0.429 | |||||
| Intraspecific populations of ZB | 0.529 | ||||||
| Intraspecific populations of ZA | 0.293 | ||||||
| Gene flow (Nm) | Interspecific populations of ZB, ZA and ZP | 0.333 | |||||
| Intraspecific populations of ZB | 0.223 | ||||||
| Intraspecific populations of ZA | 0.605 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, Y.; He, Z.; Li, Y.; Ye, M.; Xiang, L. Six Express Sequence Tag–Simple Sequence Repeat Primers Reveal Genetic Diversity in the Cultivars of Three Zanthoxylum Species. Curr. Issues Mol. Biol. 2023, 45, 7183-7196. https://doi.org/10.3390/cimb45090454
Deng Y, He Z, Li Y, Ye M, Xiang L. Six Express Sequence Tag–Simple Sequence Repeat Primers Reveal Genetic Diversity in the Cultivars of Three Zanthoxylum Species. Current Issues in Molecular Biology. 2023; 45(9):7183-7196. https://doi.org/10.3390/cimb45090454
Chicago/Turabian StyleDeng, Yangchuan, Zhoujian He, Yanlin Li, Meng Ye, and Li Xiang. 2023. "Six Express Sequence Tag–Simple Sequence Repeat Primers Reveal Genetic Diversity in the Cultivars of Three Zanthoxylum Species" Current Issues in Molecular Biology 45, no. 9: 7183-7196. https://doi.org/10.3390/cimb45090454