
Transcriptomic Signature of Horseshoe Crab Carcinoscorpius rotundicauda Hemocytes’ Response to Lipopolysaccharides

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. LPS Challenge
2.3. RNA Extraction and Sequencing
2.4. RNA-Seq Analysis
3. Results
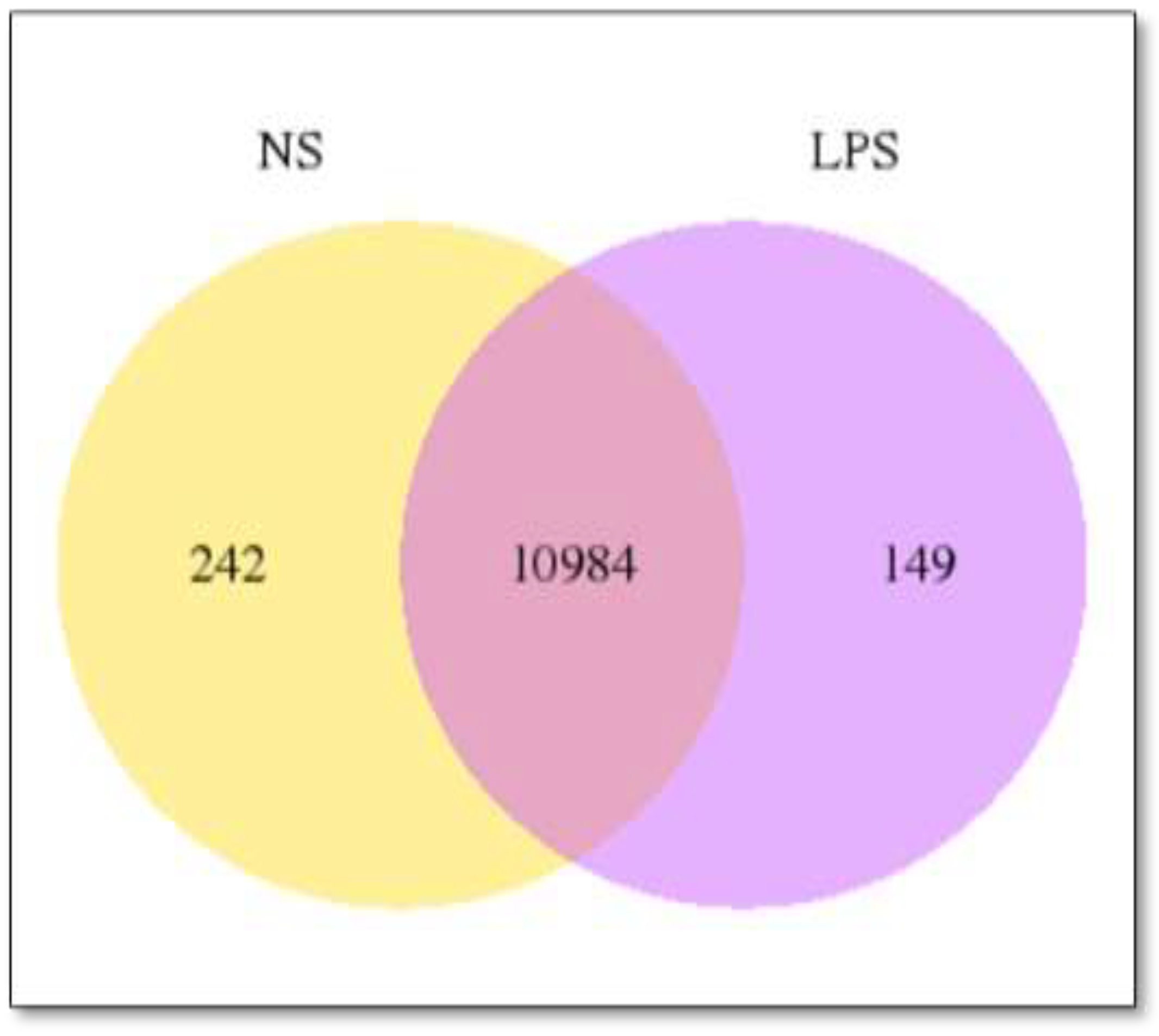
3.1. Transcriptomics Data
3.2. Differentially Expressed Genes (DEGs)
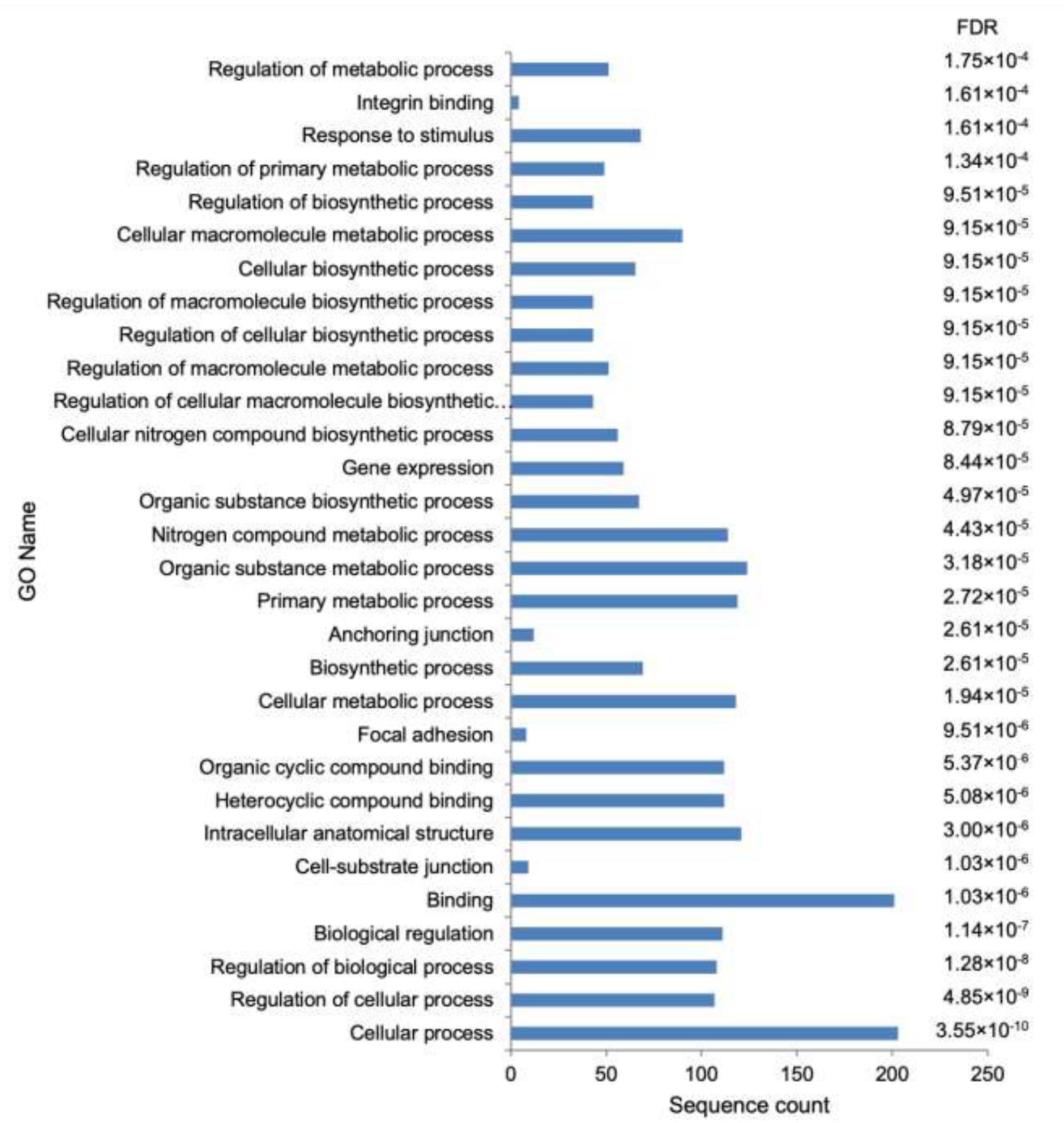
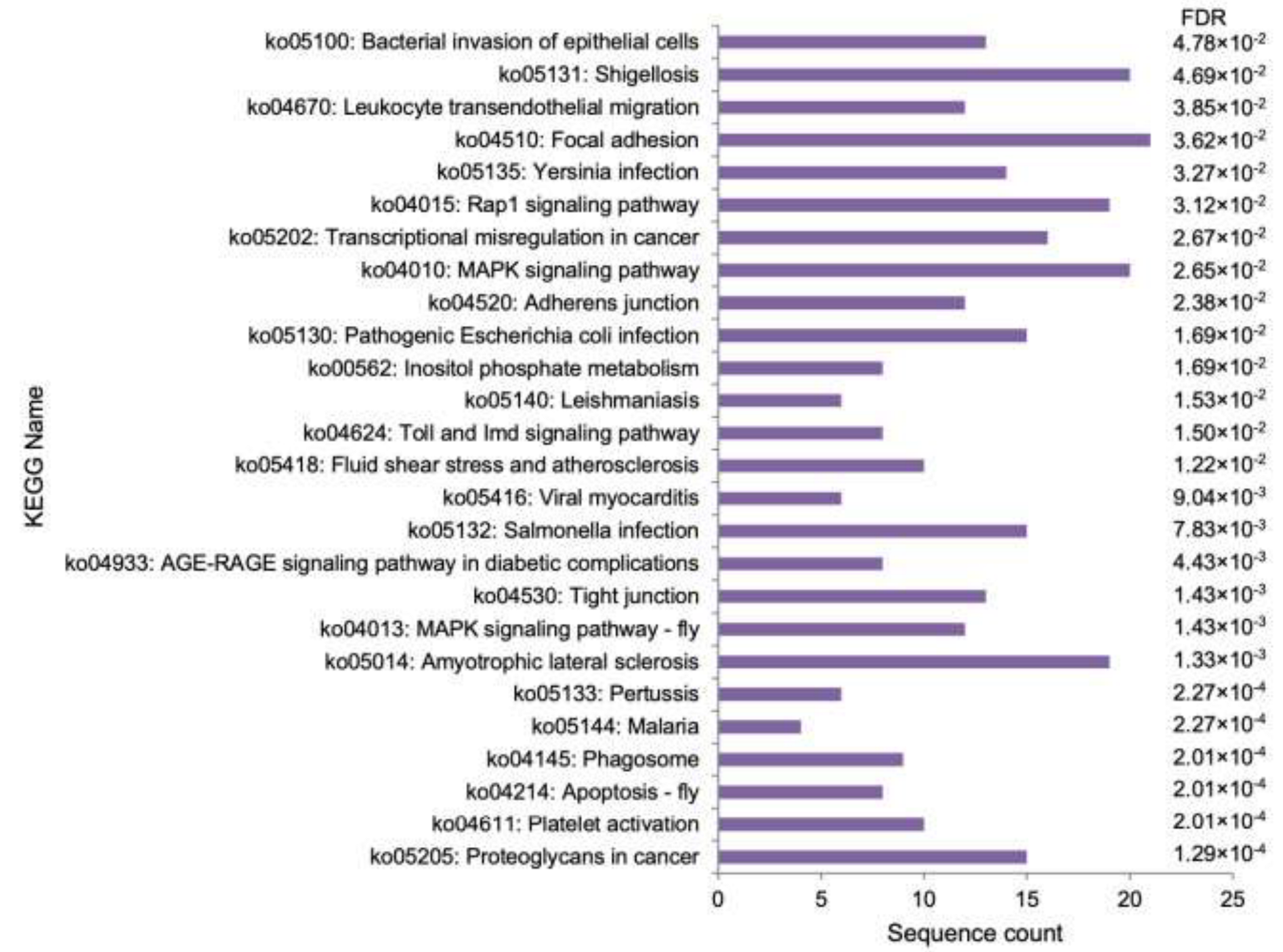
3.3. Functional Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sekiguchi, K.; Sugita, H. Systematics and hybridization in the four living species of horseshoe crabs. Evolution 1980, 34, 712–718. [Google Scholar] [CrossRef]
- Kenny, N.J.; Chan, K.W.; Nong, W.; Qu, Z.; Maeso, I.; Yip, H.Y.; Chan, T.F.; Kwan, H.S.; Holland, P.W.H.; Chu, K.H.; et al. Ancestral whole-genome duplication in the marine chelicerate horseshoe crabs. Heredity 2016, 116, 190–199. [Google Scholar] [CrossRef]
- Tinker-Kulberg, R.; Dellinger, K.; Brady, T.E.; Robertson, L.; Levy, J.H.; Abood, S.K.; LaDuca, F.M.; Kepley, C.L.; Dellinger, A.L. Horseshoe crab aquaculture as a sustainable endotoxin testing source. Front. Mar. Sci. 2020, 7, 153. [Google Scholar] [CrossRef] [Green Version]
- Krisfalusi-Gannon, J.; Ali, W.; Dellinger, K.; Robertson, L.; Brady, T.E.; Goddard, M.K.M.; Tinker-Kulberg, R.; Kepley, C.L.; Dellinger, A.L. The role of horseshoe crabs in the biomedical industry and recent trends impacting species sustainability. Front. Mar. Sci. 2018, 5, 185. [Google Scholar] [CrossRef]
- Ding, J.L.; Tan, K.C.; Thangamani, S.; Kusuma, N.; Seow, W.K.; Bui, T.H.H.; Wang, J.; Ho, B. Spatial and temporal coordination of expression of immune response genes during Pseudomonas infection of horseshoe crab, Carcinoscorpius rotundicauda. Genes Immun. 2005, 6, 557–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, C.M.; Chiu, S.F.; Akbarpour, M.; Bharat, A.; Ridge, K.M.; Bartom, E.T.; Winter, D.R. A beginner’s guide to analysis of RNA sequencing data. Am. J. Respir. Cell Mol. Biol. 2018, 59, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Sarmiento, M.E.; Chin, K.L.; Lau, N.S.; Aziah, I.; Ismail, N.; Norazmi, M.N.; Acosta, A.; Yaacob, N.S. Comparative transcriptome profiling of horseshoe crab Tachypleus gigas hemocytes in response to lipopolysaccharides. Fish Shellfish. Immunol. 2021, 117, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, A.; Ariki, S.; Kawabata, S.-I. An antimicrobial peptide tachyplesin acts as a secondary secretagogue and amplifies lipopolysaccharide-induced hemocyte exocytosis. FEBS J. 2005, 272, 3863–3871. [Google Scholar] [CrossRef]
- Shingate, P.; Ravi, V.; Prasad, A.; Tay, B.-H.; Garg, K.M.; Chattopadhyay, B.; Yap, L.-M.; Rheindt, F.E.; Venkatesh, B. Chromosome-level assembly of the horseshoe crab genome provides insights into its genome evolution. Nat. Commun. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq--a Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwanaga, S. The molecular basis of innate immunity in the horseshoe crab. Curr. Opin. Immunol. 2002, 14, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Iwanaga, S.; Kawabata, S.-I. Evolution and phylogeny of defense molecules associated with innate immunity in horseshoe crab. Front. Biosci. 1998, 3, D973–D984. [Google Scholar] [PubMed] [Green Version]
- Wang, W.-F.; Xie, X.-Y.; Chen, K.; Chen, X.-L.; Zhu, W.-L.; Wang, H.-L. Immune responses to Gram-negative bacteria in hemolymph of the Chinese horseshoe crab, Tachypleus tridentatus. Front. Immunol. 2021, 11, 584808. [Google Scholar] [CrossRef]
- Arzeta-Pino, L.; Acosta, A.; Sarmiento, M.E.; Rojas-Contreras, M.; Rodríguez-Jaramillo, C.; Vázquez-Juárez, R. Herpes virus OsHV-1 and the protist Perkinsus marinus modify the expression of the Down syndrome cell adhesion molecule gene in gill and mantle of Crassostrea spp. Aquac. Res. 2018, 49, 3638–3646. [Google Scholar] [CrossRef]
- Ng, T.H.; Chiang, Y.-A.; Yeh, Y.-C.; Wang, H.-C. Review of Dscam-mediated immunity in shrimp and other arthropods. Dev. Comp. Immunol. 2014, 46, 129–138. [Google Scholar] [CrossRef]
- Ng, T.H.; Kurtz, J. Dscam in immunity: A question of diversity in insects and crustaceans. Dev. Comp. Immunol. 2020, 105, 103539. [Google Scholar] [CrossRef]
- Akira, S. Toll-like receptor signaling. J. Biol. Chem. 2003, 278, 38105–38108. [Google Scholar] [CrossRef] [Green Version]
- Iwanaga, S.; Kawabata, S.-I.; Muta, T. New types of clotting factors and defense molecules found in horseshoe crab hemolymph: Their structures and functions. J. Biochem. 1998, 123, 1–15. [Google Scholar] [CrossRef]
- Diks, S.H.; van Deventer, S.J.; Peppelenbosch, M.P. Invited review: Lipopolysaccharide recognition, internalisation, signalling and other cellular effects. J. Endotoxin Res. 2001, 7, 335–348. [Google Scholar] [CrossRef]
- Raetz, C.R.; Whitfield, C. Lipopolysaccharide endotoxins. Annu. Rev. Biochem. 2002, 71, 635–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawabata, S.; Koshiba, T.; Shibata, T. The lipopolysaccharide-activated innate immune response network of the horseshoe crab. Invertebr. Surviv. J. 2009, 6, 59–77. [Google Scholar]
- Conrad, M.; Denobile, J.; Chaikhoutdinov, I.; Escribano, D.; Lee, K.-G.; Cohen, W.D. Cytoskeletal organization of limulus amebocytes pre-and post-activation: Comparative aspects. Biol. Bull. 2004, 207, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, K.A.; Kagan, J.C. Toll-like receptors and the control of immunity. Cell 2020, 180, 1044–1066. [Google Scholar] [CrossRef]
- Anderson, K.V. Toll signaling pathways in the innate immune response. Curr. Opin. Immunol. 2000, 12, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Bertheloot, D.; Latz, E.; Franklin, B.S. Necroptosis, pyroptosis and apoptosis: An intricate game of cell death. Cell. Mol. Immunol. 2021, 18, 1106–1121. [Google Scholar] [CrossRef]
- Häcker, G. Apoptosis in infection. Microbes Infect. 2018, 20, 552–559. [Google Scholar] [CrossRef]
- Arthur, J.S.C.; Ley, S.C. Mitogen-activated protein kinases in innate immunity. Nat. Rev. Immunol. 2013, 13, 679–692. [Google Scholar] [CrossRef]
- Zhang, Y.L.; Dong, C. MAP kinases in immune responses. Cell. Mol. Immunol. 2005, 2, 20–27. [Google Scholar]
- Dong, C.; Davis, R.J.; Flavell, R.A. MAP kinases in the immune response. Annu. Rev. Immunol. 2002, 20, 55–72. [Google Scholar] [CrossRef] [PubMed]
- Hammer, M.; Mages, J.R.; Dietrich, H.; Servatius, A.; Howells, N.; Cato, A.C.; Lang, R. Dual specificity phosphatase 1 (DUSP1) regulates a subset of LPS-induced genes and protects mice from lethal endotoxin shock. J. Exp. Med. 2006, 203, 15–20. [Google Scholar] [CrossRef]
- Kondoh, K.; Nishida, E. Regulation of MAP kinases by MAP kinase phosphatases. Biochim. Biophys. Acta Mol. Cell Res. 2007, 1773, 1227–1237. [Google Scholar] [CrossRef] [Green Version]
- Lang, R.; Raffi, F.A. Dual-specificity phosphatases in immunity and infection: An update. Int. J. Mol. Sci. 2019, 20, 2710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciesielska, A.; Matyjek, M.; Kwiatkowska, K. TLR4 and CD14 trafficking and its influence on LPS-induced pro-inflammatory signaling. Cell. Mol. Life Sci. 2021, 78, 1233–1261. [Google Scholar] [CrossRef] [PubMed]
- Płóciennikowska, A.; Zdioruk, M.I.; Traczyk, G.; Świątkowska, A.; Kwiatkowska, K. LPS-induced clustering of CD14 triggers generation of PI (4, 5) P 2. J. Cell Sci. 2015, 128, 4096–4111. [Google Scholar] [CrossRef] [Green Version]
- Shorning, B.Y.; Dass, M.S.; Smalley, M.J.; Pearson, H.B. The PI3K-AKT-mTOR pathway and prostate cancer: At the crossroads of AR, MAPK, and WNT signaling. Int. J. Mol. Sci. 2020, 21, 4507. [Google Scholar] [CrossRef]
- Li, S.; Sullivan, N.L.; Rouphael, N.; Yu, T.; Banton, S.; Maddur, M.S.; McCausland, M.; Chiu, C.; Canniff, J.; Dubey, S. Metabolic phenotypes of response to vaccination in humans. Cell 2017, 169, 862–877. [Google Scholar] [CrossRef] [Green Version]
- Liao, X.; Wang, C.; Wang, B.; Qin, H.; Hu, S.; Wang, P.; Sun, C.; Zhang, S. Comparative transcriptome analysis of Litopenaeus vannamei reveals that triosephosphate isomerase-like genes play an important role during decapod iridescent virus 1 infection. Front. Immunol. 2020, 11, 1904. [Google Scholar] [CrossRef]
- Wu, F.; Xie, Z.; Yan, M.; Li, Q.; Song, J.; Hu, M.; Wang, Y. Classification and characterization of hemocytes from two Asian horseshoe crab species Tachypleus tridentatus and Carcinoscorpius rotundicauda. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Roy, S.; Sahoo, A.; Kumar, V. Horseshoe crabs: Biomedical importance and its potential use in developing health-care products. Indian J. Geo Mar. Sci. 2016, 45, 1234–1244. [Google Scholar]
- Coates, C.J.; Whalley, T.; Nairn, J. Phagocytic activity of Limulus polyphemus amebocytes in vitro. J. Invertebr. Pathol. 2012, 111, 205–210. [Google Scholar] [CrossRef] [PubMed]
- MacPherson, J.C.; Pavlovich, J.G.; Jacobs, R.S. Phospholipid composition of the granular amebocyte from the horseshoe crab, Limulus polyphemus. Lipids 1998, 33, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Mazgaeen, L.; Gurung, P. Recent advances in lipopolysaccharide recognition systems. Int. J. Mol. Sci. 2020, 21, 379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, M.J.; Kell, D.B.; Pretorius, E. The role of lipopolysaccharide-induced cell signalling in chronic inflammation. Chronic Stress 2022, 6, 24705470221076390. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, A.E.; Rotjan, R.D.; Kagan, J.C. Lipopolysaccharide detection by the innate immune system may be an uncommon defence strategy used in nature. Open Biol. 2022, 12, 220146. [Google Scholar] [CrossRef]
- Adebayo, I.A.; Habib, M.A.H.; Sarmiento, M.E.; Acosta, A.; Yaacob, N.S.; Ismail, M.N. Proteomic analysis of Malaysian Horseshoe crab (Tachypleus gigas) hemocytes gives insights into its innate immunity host defence system and other biological processes. PLoS ONE 2022, 17, e0272799. [Google Scholar] [CrossRef]
- Amiss, A.S.; von Pein, J.B.; Webb, J.R.; Condon, N.D.; Harvey, P.J.; Phan, M.-D.; Schembri, M.A.; Currie, B.J.; Sweet, M.J.; Craik, D.J.; et al. Modified horseshoe crab peptides target and kill bacteria inside host cells. Cell. Mol. Life Sci. 2021, 79, 38. [Google Scholar] [CrossRef]
- Wang, W.-F.; Xie, X.-Y.; Huang, Y.; Li, Y.-K.; Liu, H.; Chen, X.-L.; Wang, H.-L. Identification of a novel antimicrobial peptide from the ancient marine arthropod Chinese horseshoe crab, Tachypleus tridentatus. Front. Immunol. 2022, 13, 794779. [Google Scholar] [CrossRef]
- Saucedo-Vázquez, J.P.; Gushque, F.; Vispo, N.S.; Rodriguez, J.; Gudiño-Gomezjurado, M.E.; Albericio, F.; Tellkamp, M.P.; Alexis, F. Marine Arthropods as a source of antimicrobial peptides. Mar. Drugs 2022, 20, 501. [Google Scholar] [CrossRef]
- Qu, Z.; Leung, T.C.N.; Nong, W.; Yip, H.Y.; Lee, I.H.T.; Cheung, S.G.; Ming, N.S.; So, W.L.; Bendena, W.G.; Tobe, S.S.; et al. Hemolymph proteomics and gut microbiota of horseshoe crabs Tachypleus tridentatus and Carcinoscorpius rotundicauda. Front. Mar. Sci. 2020, 7, 579706. [Google Scholar] [CrossRef]
- Tinker-Kulberg, R.; Dellinger, A.; Brady, T.E.; Robertson, L.; Goddard, M.K.M.; Bowzer, J.; Abood, S.K.; Kepley, C.; Dellinger, K. Effects of diet on the biochemical properties of limulus amebocyte lysate from horseshoe crabs in an aquaculture setting. Front. Mar. Sci. 2020, 7, 541604. [Google Scholar] [CrossRef]
- Liu, X.; Liu, J.; Xiong, K.; Zhang, C.; Fang, J.K.-H.; Song, J.; Tai, Z.; Zhu, Q.; Hu, M.; Wang, Y. Effects of ocean acidification on molting, oxidative stress, and gut microbiota in juvenile horseshoe crab Tachypleus tridentatus. Front. Physiol. 2022, 12, 813582. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kwan, K.Y.; Li, S.; Liu, Y.; Weng, B.; Huang, W.; Weng, Z. Transcriptome analysis of the digestive tract of Tachypleus tridentatus and Carcinoscorpius rotundicauda. J. Ocean. Univ. China 2022, 21, 591–600. [Google Scholar] [CrossRef]
- Sarmiento, M.E.; Chin, K.L.; Lau, N.S.; Aziah, I.; Norazmi, M.N.; Acosta, A.; Ismail, N.; Yaacob, N.S. Mitochondrial DNA sequence of the horseshoe crab Tachypleus gigas. Mitochondrial DNA B Resour. 2021, 6, 1710–1714. [Google Scholar] [CrossRef]
- Gygi, S.P.; Rochon, Y.; Franza, B.R.; Aebersold, R. Correlation between protein and mRNA abundance in yeast. Mol. Cell. Biol. 1999, 19, 1720–1730. [Google Scholar] [CrossRef] [Green Version]
- Koussounadis, A.; Langdon, S.P.; Um, I.H.; Harrison, D.J.; Smith, V.A. Relationship between differentially expressed mRNA and mRNA-protein correlations in a xenograft model system. Sci. Rep. 2015, 5, 10775. [Google Scholar] [CrossRef] [Green Version]
- Maier, T.; Güell, M.; Serrano, L. Correlation of mRNA and protein in complex biological samples. FEBS Lett. 2009, 583, 3966–3973. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sample | Number of Raw Reads | Number of Clean Reads | Clean Bases (Gb) | Mapped Reads | Mapped Ratio (%) |
|---|---|---|---|---|---|
| CrNS1 | 73,148,414 | 73,144,174 | 10.92 | 60,165,568 | 82.3 |
| CrNS2 | 76,737,552 | 76,732,692 | 11.46 | 62,257,906 | 81.1 |
| CrNS3 | 82,742,120 | 82,736,822 | 12.36 | 67,902,779 | 82.1 |
| Total | 232,628,086 | 232,613,688 | 34.74 | 190,326,253 | Average: 81.83 |
| CrLPS1 | 82,252,646 | 82,246,900 | 12.27 | 67,152,864 | 81.6 |
| CrLPS2 | 74,186,052 | 74,180,514 | 11.07 | 60,499,561 | 81.6 |
| CrLPS3 | 89,009,478 | 89,003,364 | 13.29 | 73,527,595 | 82.6 |
| Total | 245,448,176 | 245,430,778 | 36.63 | 201,180,020 | Average: 81.93 |
| No. | Annotation | C. rotundicauda (Current Study) | T. gigas | T. tridentatus |
|---|---|---|---|---|
| 1 | GO:0009987 | cellular process | X | X |
| 2 | GO:0050794 | regulation of cellular process | X | |
| 3 | GO:0050789 | regulation of biological process | X | |
| 4 | GO:0065007 | biological regulation | X | |
| 5 | GO:0005488 | binding | X | |
| 6 | GO:0030055 | cell–substrate junction | ||
| 7 | GO:0005622 | intracellular anatomical structure | X | |
| 8 | GO:1901363 | heterocyclic compound binding | ||
| 9 | GO:0097159 | organic cyclic compound binding | ||
| 10 | GO:0005925 | focal adhesion | X | |
| 11 | GO:0044237 | cellular metabolic process | X | |
| 12 | GO:0009058 | biosynthetic process | ||
| 13 | GO:0070161 | anchoring junction | ||
| 14 | GO:0044238 | primary metabolic process | ||
| 15 | GO:0071704 | organic substance metabolic process | ||
| 16 | GO:0006807 | nitrogen compound metabolic process | X | |
| 17 | GO:1901576 | organic substance biosynthetic process | ||
| 18 | GO:0010467 | gene expression | ||
| 19 | GO:0044271 | cellular nitrogen compound biosynthetic process | ||
| 20 | GO:2000112 | regulation of cellular macromolecule biosynthetic process | ||
| 21 | GO:0060255 | regulation of macromolecule metabolic process | ||
| 22 | GO:0031326 | regulation of cellular biosynthetic process | ||
| 23 | GO:0010556 | regulation of macromolecule biosynthetic process | ||
| 24 | GO:0044249 | cellular biosynthetic process | ||
| 25 | GO:0044260 | cellular macromolecule metabolic process | ||
| 26 | GO:0009889 | regulation of biosynthetic process | ||
| 27 | GO:0080090 | regulation of primary metabolic process | ||
| 28 | GO:0050896 | response to stimulus | X | |
| 29 | GO:0005178 | integrin binding | ||
| 30 | GO:0019222 | regulation of metabolic process |
| Annotation | C. rotundicauda (Current Study) | T. gigas | T. tridentatus | |
|---|---|---|---|---|
| 1 | ko05100 | Bacterial invasion of epithelial cells | ||
| 2 | ko04510 | Focal adhesion | X | |
| 3 | ko04670 | Leukocyte transendothelial migration | ||
| 4 | ko05131 | Shigellosis | ||
| 5 | ko04015 | Rap1 signaling pathway | ||
| 6 | ko05135 | Yersinia infection | ||
| 7 | ko05202 | Transcriptional misregulation in cancer | X | |
| 8 | ko04520 | Adherens junction | ||
| 9 | ko04010 | MAPK signaling pathway | X | |
| 10 | ko05130 | Pathogenic Escherichia coli infection | ||
| 11 | ko00562 | Inositol phosphate metabolism | ||
| 12 | ko05140 | Leishmaniasis | X | |
| 13 | ko04624 | Toll and Imd signaling pathway | X | X |
| 14 | ko05418 | Fluid shear stress and atherosclerosis | X | |
| 15 | ko05416 | Viral myocarditis | ||
| 16 | ko04933 | AGE-RAGE signaling pathway in diabetic complications | ||
| 17 | ko05132 | Salmonella infection | ||
| 18 | ko04530 | Tight junction | X | |
| 19 | ko04013 | MAPK signaling pathway-fly | ||
| 20 | ko05014 | Amyotrophic lateral sclerosis | ||
| 21 | ko05133 | Pertussis | X | |
| 22 | ko05144 | Malaria | ||
| 23 | ko04145 | Phagosome | ||
| 24 | ko04214 | Apoptosis-fly | ||
| 25 | ko04611 | Platelet activation | X | |
| 26 | ko05205 | Proteoglycans in cancer |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarmiento, M.E.; Chin, K.L.; Lau, N.-S.; Ismail, N.; Norazmi, M.N.; Acosta, A.; Yaacob, N.S. Transcriptomic Signature of Horseshoe Crab Carcinoscorpius rotundicauda Hemocytes’ Response to Lipopolysaccharides. Curr. Issues Mol. Biol. 2022, 44, 5866-5878. https://doi.org/10.3390/cimb44120399
Sarmiento ME, Chin KL, Lau N-S, Ismail N, Norazmi MN, Acosta A, Yaacob NS. Transcriptomic Signature of Horseshoe Crab Carcinoscorpius rotundicauda Hemocytes’ Response to Lipopolysaccharides. Current Issues in Molecular Biology. 2022; 44(12):5866-5878. https://doi.org/10.3390/cimb44120399
Chicago/Turabian StyleSarmiento, Maria E., Kai Ling Chin, Nyok-Sean Lau, Noraznawati Ismail, Mohd Nor Norazmi, Armando Acosta, and Nik Soriani Yaacob. 2022. "Transcriptomic Signature of Horseshoe Crab Carcinoscorpius rotundicauda Hemocytes’ Response to Lipopolysaccharides" Current Issues in Molecular Biology 44, no. 12: 5866-5878. https://doi.org/10.3390/cimb44120399