
Mesenchymal Stem Cells in the Treatment of Human Spinal Cord Injury: The Effect on Individual Values of pNF-H, GFAP, S100 Proteins and Selected Growth Factors, Cytokines and Chemokines

 , , , , ,
, , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
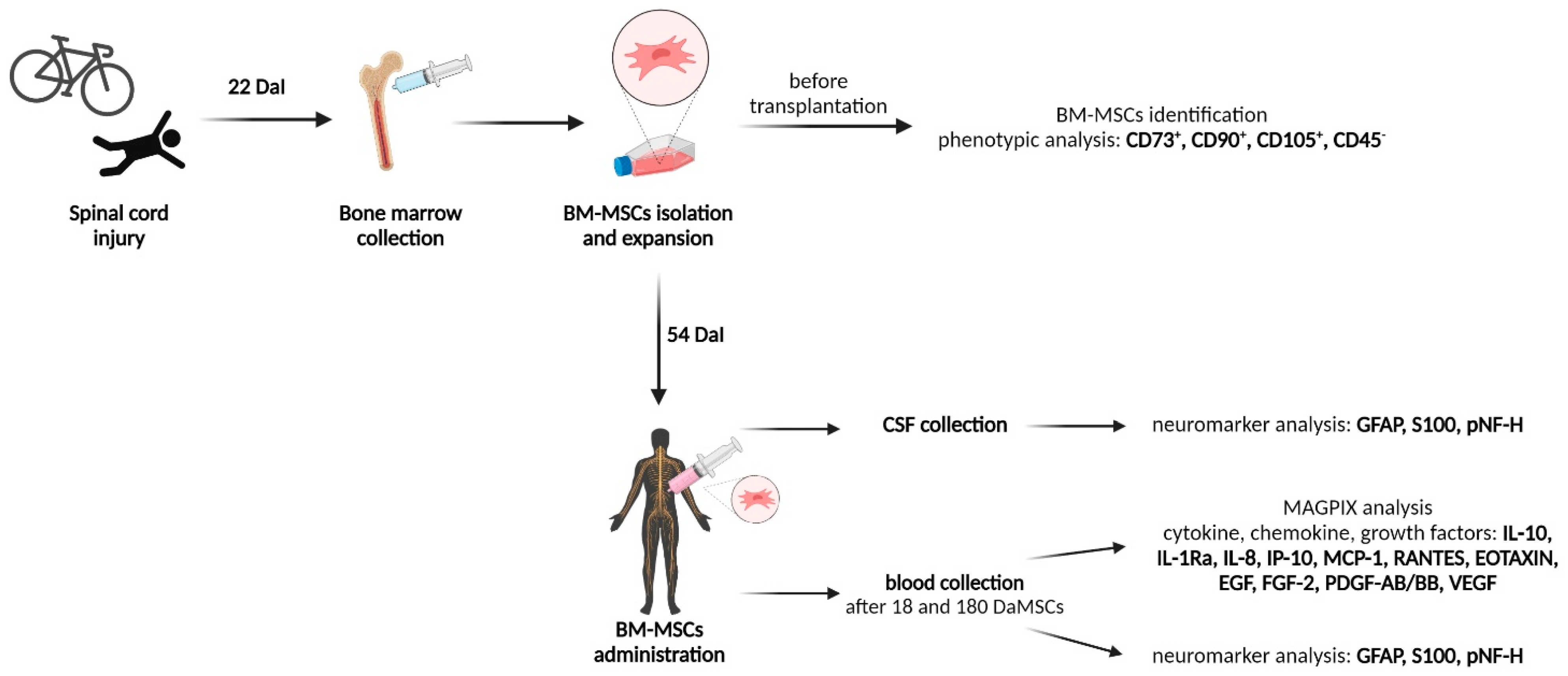
2.1. Patient
2.2. BM-MSCs Cultivation and Administration
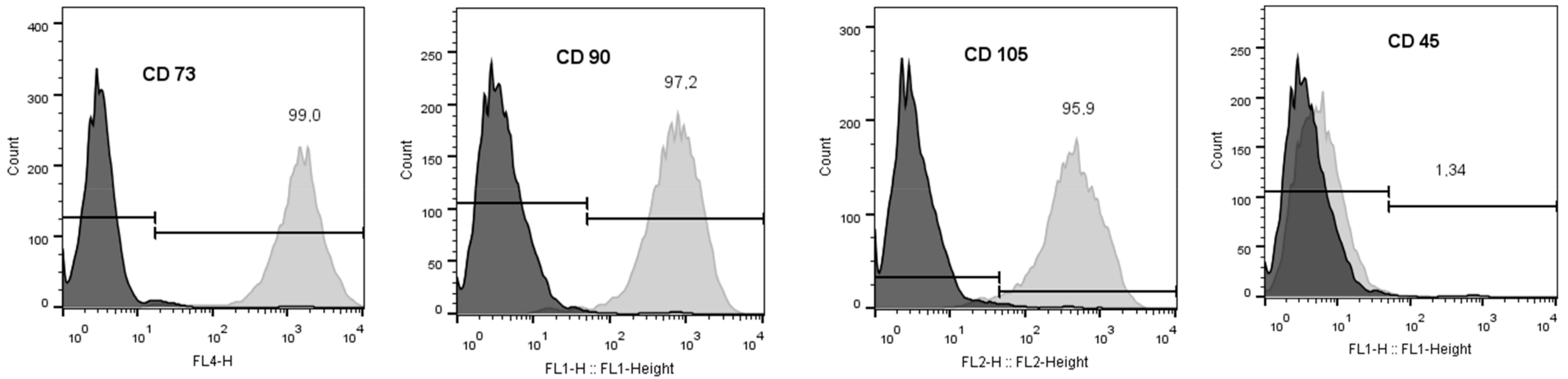
2.3. Phenotypic Analysis of BM-MSCs
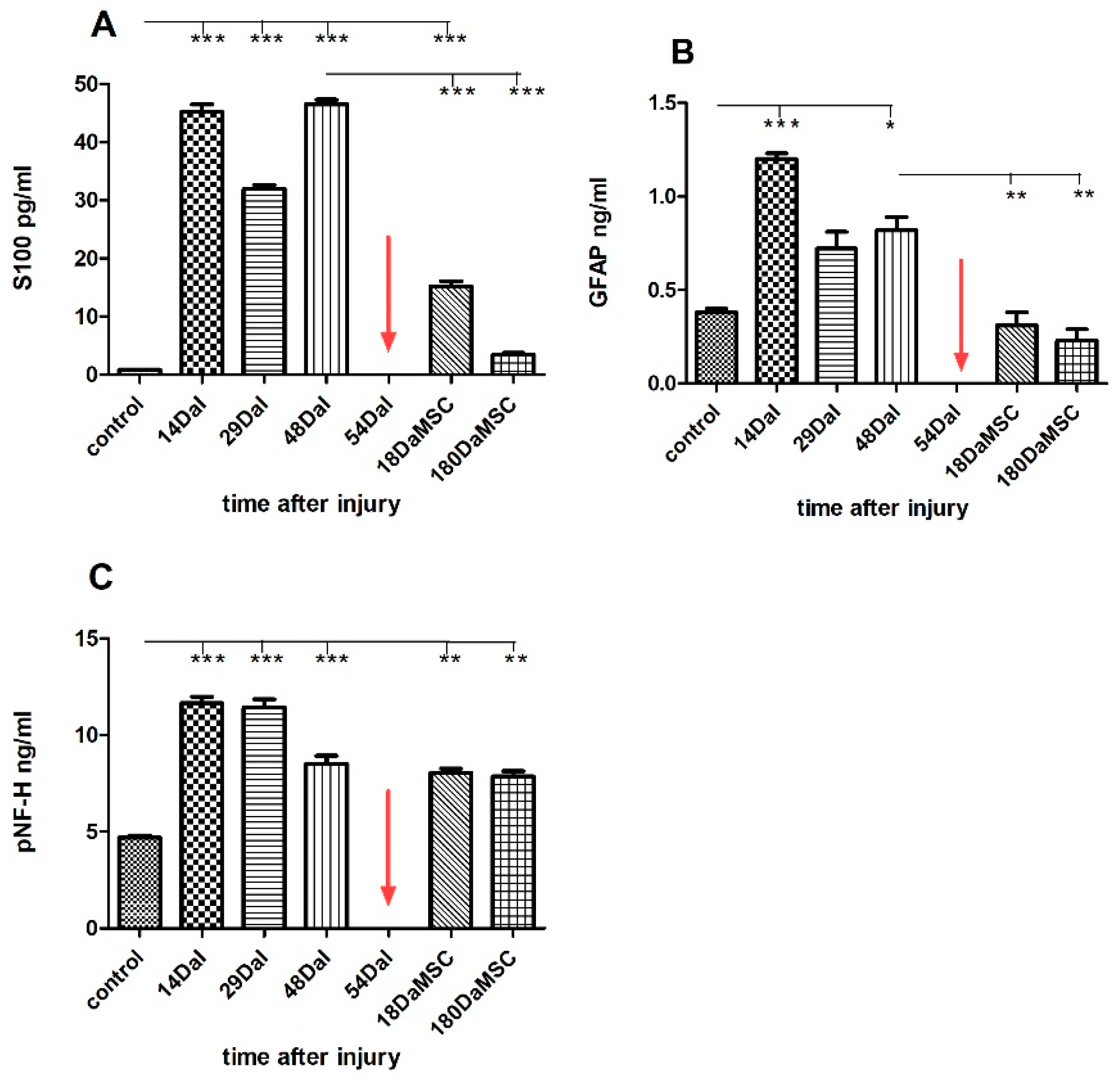
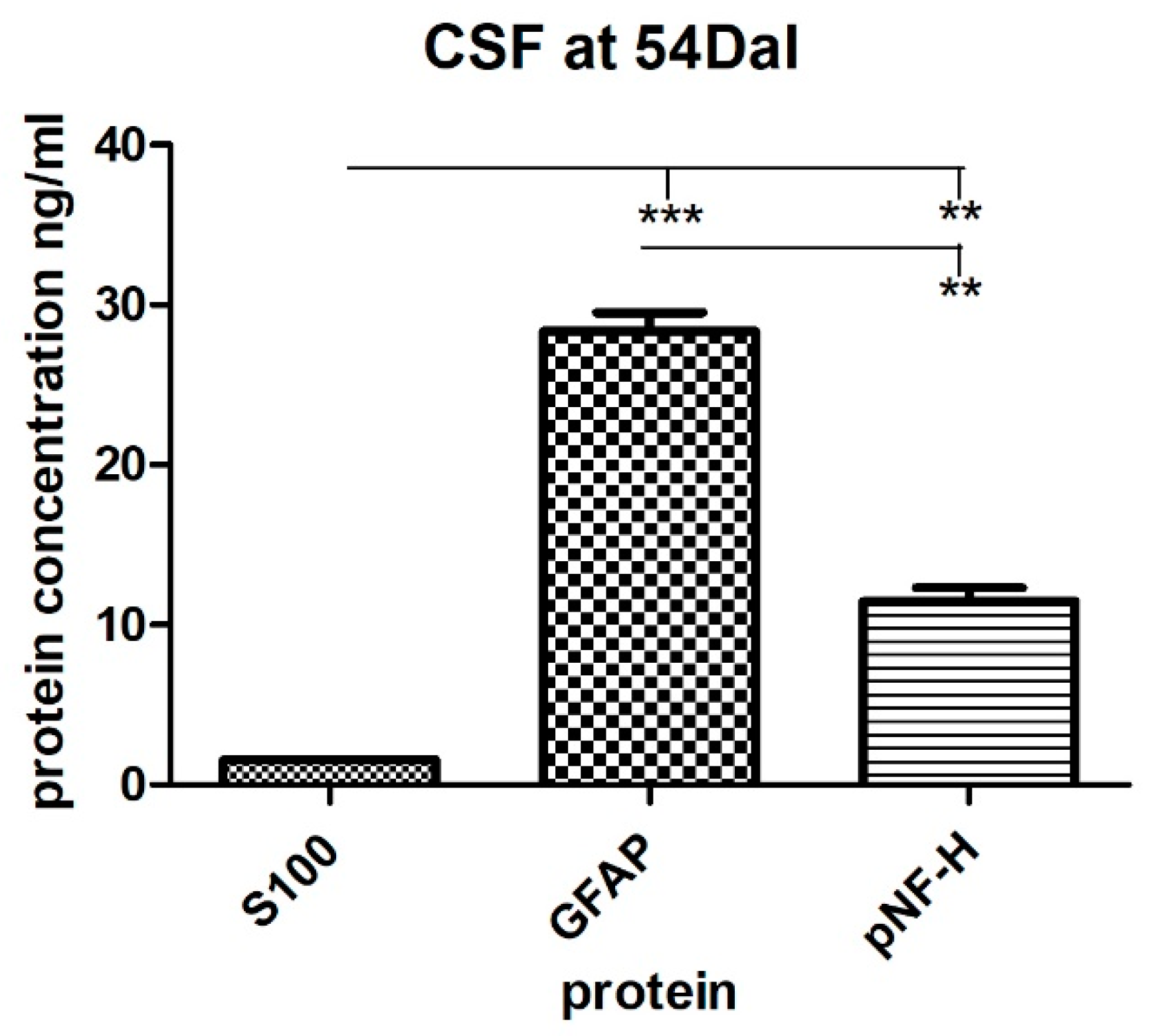
2.4. The Levels of S100, GFAP and pNF-H Protein in the Serum and the CSF of a SCI Patient Treated with Autologous BM-MSCs
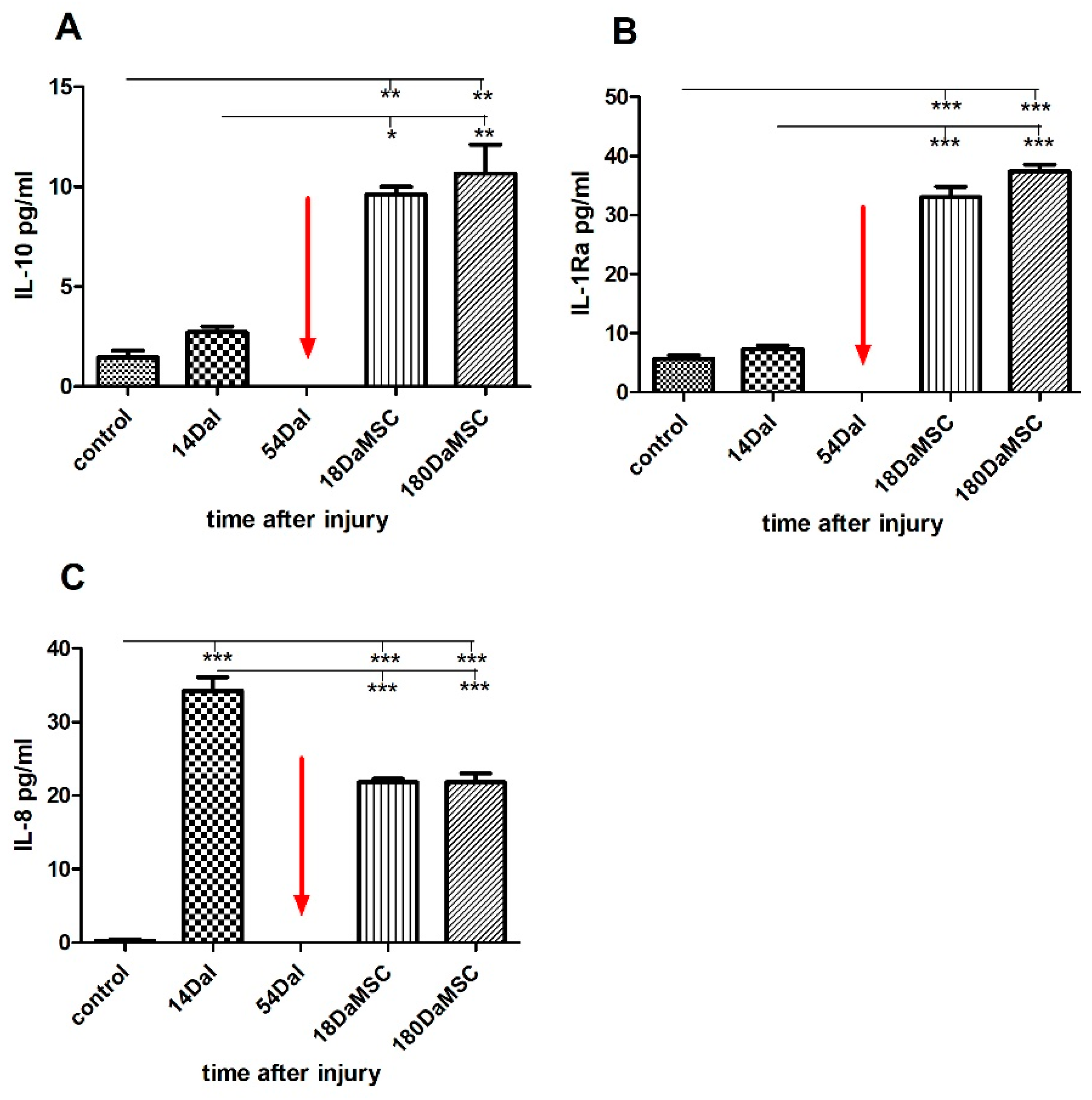
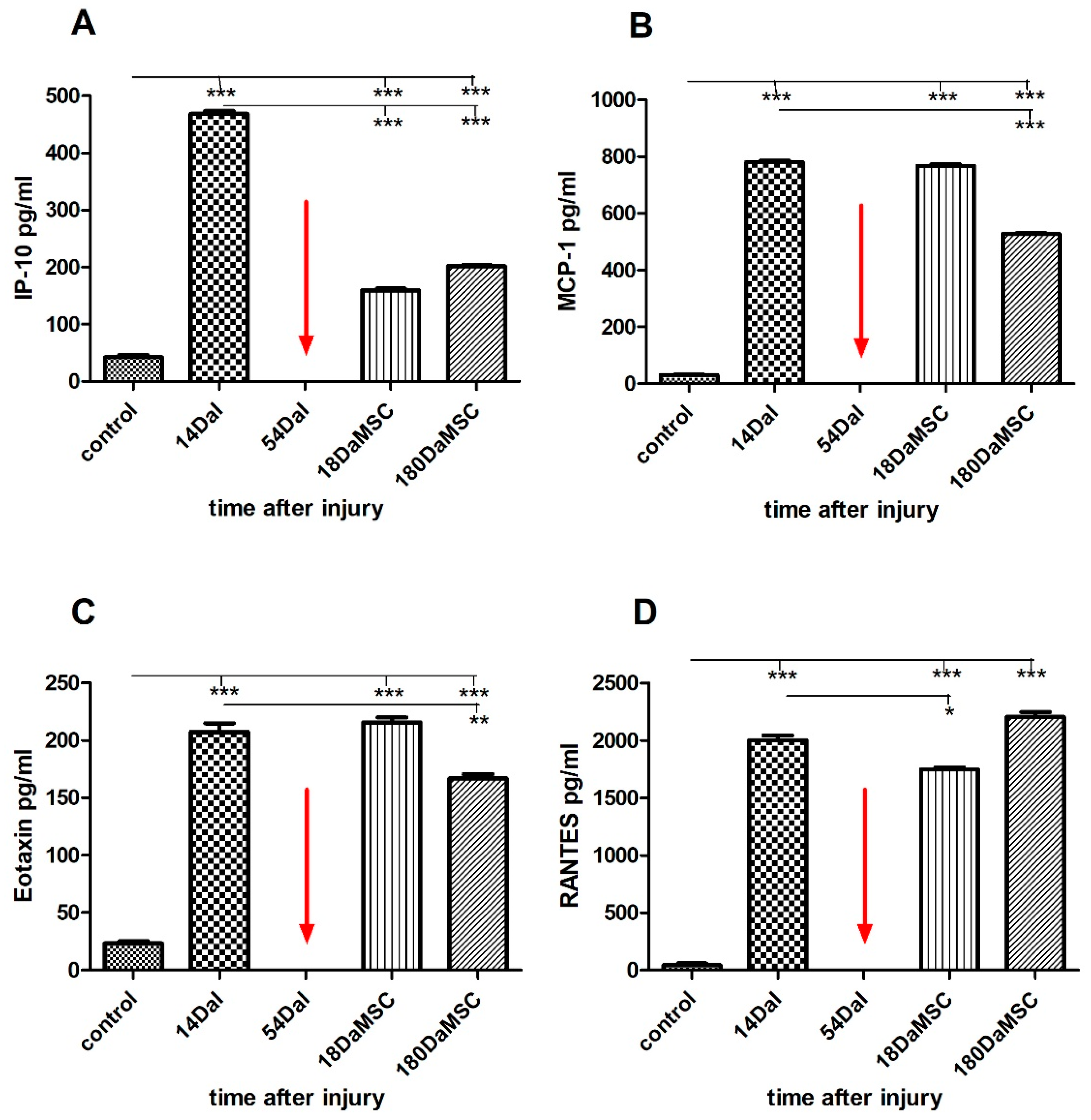
2.5. BM-MSCs Influenced Cytokine/Chemokine Levels in Injured Spinal Cord
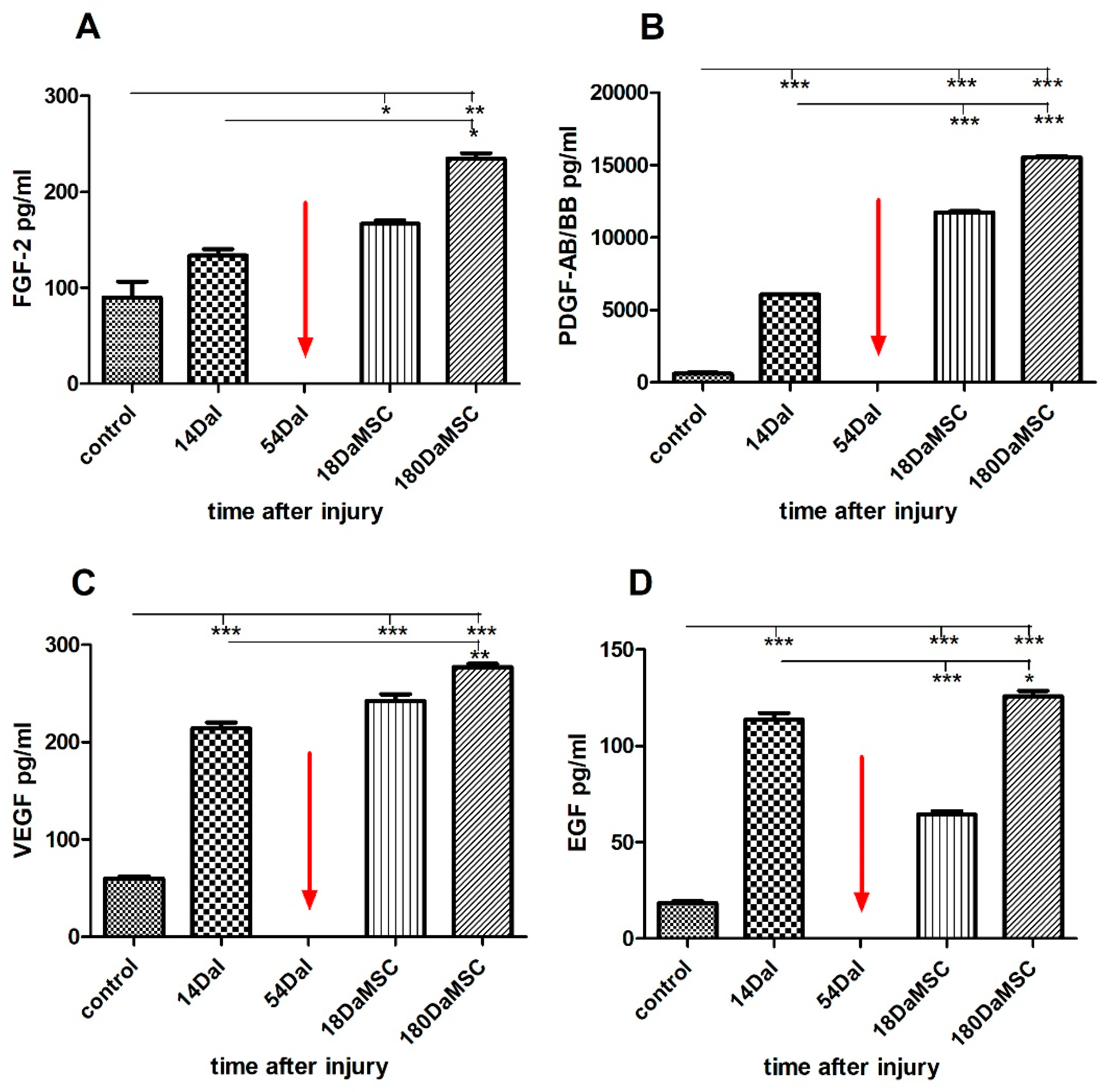
2.6. BM-MSCs Treatment Changed the Serum Levels of Growth Factors after SCI
3. Material and Methods
3.1. Patient Enrolment
3.2. Bone Marrow Collection and MSCs Isolation
3.3. Phenotypic Analysis of BM-MSCs before Application
3.4. Preparation of BM-MSCs for Clinical Application
3.5. BM-MSCs Administration
3.6. Sample Collection
3.6.1. Serum
3.6.2. Cerebrospinal Fluid
3.7. Multiplex Assay for Analysis of Cytokine/Chemokine/Growth Factors
3.8. ELISA Measurement of Human S100, GFAP, and pNF-H
3.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Grégoire, C.-A.; Goldenstein, B.L.; Floriddia, E.M.; Barnabé-Heider, F.; Fernandes, K.J.L. Endogenous neural stem cell responses to stroke and spinal cord injury. Glia 2015, 63, 1469–1482. [Google Scholar] [CrossRef] [PubMed]
- Dumont, R.J.; Okonkwo, D.O.; Verma, S.; Hurlbert, R.J.; Boulos, P.T.; Ellegala, D.B.; Dumont, A.S. Acute Spinal Cord Injury, Part I: Pathophysiologic Mechanisms. Clin. Neuropharmacol. 2001, 24, 254–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.; Dai, Y.; Chen, G.; Cui, S. Dissecting the Dual Role of the Glial Scar and Scar-Forming Astrocytes in Spinal Cord Injury. Front. Cell. Neurosci. 2020, 14, 78. [Google Scholar] [CrossRef] [Green Version]
- Hagg, T.; Oudega, M. Degenerative and Spontaneous Regenerative Processes after Spinal Cord Injury. J. Neurotrauma 2006, 23, 263–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowland, J.W.; Hawryluk, G.W.J.; Kwon, B.; Fehlings, M.G. Current status of acute spinal cord injury pathophysiology and emerging therapies: Promise on the horizon. Neurosurg. Focus 2008, 25, E2. [Google Scholar] [CrossRef] [Green Version]
- Lu, P.; Dulin, J.N. Bridging the injured spinal cord with neural stem cells. Neural Regen. Res. 2014, 9, 229–231. [Google Scholar] [CrossRef]
- Levi, A.D.; Anderson, K.D.; Okonkwo, D.O.; Park, P.; Bryce, T.N.; Kurpad, S.N.; Aarabi, B.; Hsieh, J.; Gant, K. Clinical Outcomes from a Multi-Center Study of Human Neural Stem Cell Transplantation in Chronic Cervical Spinal Cord Injury. J. Neurotrauma 2019, 36, 891–902. [Google Scholar] [CrossRef]
- Lu, P.; Wang, Y.; Graham, L.; McHale, K.; Gao, M.; Wu, D.; Brock, J.; Blesch, A.; Rosenzweig, E.; Havton, L.A.; et al. Long-Distance Growth and Connectivity of Neural Stem Cells after Severe Spinal Cord Injury. Cell 2012, 150, 1264–1273. [Google Scholar] [CrossRef] [Green Version]
- Kanno, H.; Pearse, D.D.; Ozawa, H.; Itoi, E.; Bunge, M.B. Schwann cell transplantation for spinal cord injury repair: Its significant therapeutic potential and prospectus. Rev. Neurosci. 2015, 26, 121–128. [Google Scholar] [CrossRef]
- Tan, C.; Yang, C.; Liu, H.; Tang, C.; Huang, S. Effect of Schwann cell transplantation combined with electroacupuncture on axonal regeneration and remyelination in rats with spinal cord injury. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2021, 304, 2506–2520. [Google Scholar] [CrossRef]
- Raisman, G.; Carlstedt, T.; Choi, D.; Li, Y. Clinical prospects for transplantation of OECs in the repair of brachial and lumbosacral plexus injuries: Opening a door. Exp. Neurol. 2011, 229, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Xu, C.-C.; Li, J.; Guan, X.-Y.; Gao, L.; Ma, L.-X.; Li, R.-X.; Peng, Y.-W.; Zhu, G.-P. Transplantation of Oligodendrocyte Precursor Cells Improves Locomotion Deficits in Rats with Spinal Cord Irradiation Injury. PLoS ONE 2013, 8, e57534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, B.; Sun, L.; Li, P.; Tian, M.; Luo, Y.; Ren, X. Transplantation of oligodendrocyte precursor cells improves myelination and promotes functional recovery after spinal cord injury. Injury 2012, 43, 794–801. [Google Scholar] [CrossRef] [PubMed]
- Tetzlaff, W.; Okon, E.; Karimi-Abdolrezaee, S.; Hill, C.E.; Sparling, J.; Plemel, J.; Plunet, W.T.; Tsai, E.; Baptiste, D.; Smithson, L.J.; et al. A Systematic Review of Cellular Transplantation Therapies for Spinal Cord Injury. J. Neurotrauma 2011, 28, 1611–1682. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- DiGirolamo, C.M.; Stokes, D.; Colter, D.; Phinney, D.; Class, R.; Prockop, D.J. Propagation and senescence of human marrow stromal cells in culture: A simple colony-forming assay identifies samples with the greatest potential to propagate and differentiate. Br. J. Haematol. 1999, 107, 275–281. [Google Scholar] [CrossRef]
- Zuk, P.A.; Zhu, M.; Ashjian, P.; De Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human adipose tissue is a source of multipotent stem cells. Mol. Biol. Cell 2002, 13, 4279–4295. [Google Scholar] [CrossRef]
- Kim, D.-W.; Staples, M.; Shinozuka, K.; Pantcheva, P.; Kang, S.-D.; Borlongan, C.V. Wharton’s Jelly-Derived Mesenchymal Stem Cells: Phenotypic Characterization and Optimizing Their Therapeutic Potential for Clinical Applications. Int. J. Mol. Sci. 2013, 14, 11692–11712. [Google Scholar] [CrossRef] [Green Version]
- Rocca, G.L. Editorial—Connecting the Dots: The Promises of Wharton’s Jelly Mesenchymal Stem Cells forTissue Repair and Regen-eration. Open Tissue Eng. Regen. Med. J. 2011, 4, 3–5. [Google Scholar] [CrossRef] [Green Version]
- Pelekanos, R.A.; Sardesai, V.S.; Futrega, K.; Lott, W.B.; Kuhn, M.; Doran, M.R. Isolation and Expansion of Mesenchymal Stem/Stromal Cells Derived from Human Placenta Tissue. J. Vis. Exp. 2016, 10, e54204. [Google Scholar] [CrossRef] [Green Version]
- Araújo, A.B.; Furlan, J.M.; Salton, G.D.; Schmalfuss, T.; Röhsig, L.M.; Silla, L.M.R.; Passos, E.P.; Paz, A.H. Isolation of human mesenchymal stem cells from amnion, chorion, placental decidua and umbilical cord: Comparison of four enzymatic protocols. Biotechnol. Lett. 2018, 40, 989–998. [Google Scholar] [CrossRef] [PubMed]
- Secunda, R.; Vennila, R.; Mohanashankar, A.M.; Rajasundari, M.; Jeswanth, S.; Surendran, R. Isolation, expansion and characterisation of mesenchymal stem cells from human bone marrow, adipose tissue, umbilical cord blood and matrix: A comparative study. Cytotechnology 2015, 67, 793–807. [Google Scholar] [CrossRef] [PubMed]
- Abrams, M.B.; Dominguez, C.; Pernold, K.; Reger, R.; Wiesenfeld-Hallin, Z.; Olson, L.; Prockop, D. Multipotent mesenchymal stromal cells attenuate chronic inflammation and injury-induced sensitivity to me-chanical stimuli in experimental spinal cord injury. Restor. Neurol. Neurosci. 2009, 27, 307–321. [Google Scholar] [PubMed]
- Jiang, W.; Xu, J. Immune modulation by mesenchymal stem cells. Cell Prolif. 2019, 53, e12712. [Google Scholar] [CrossRef] [PubMed]
- Yan, K.; Zhang, R.; Sun, C.; Chen, L.; Li, P.; Liu, Y.; Peng, L.; Sun, H.; Qin, K.; Chen, F.; et al. Bone marrow-derived mesenchymal stem cells maintain the resting phenotype of microglia and inhibit microglial acti-vation. PLoS ONE 2013, 8, e84116. [Google Scholar] [CrossRef] [PubMed]
- Kruminis-Kaszkiel, E.; Osowski, A.; Bejer-Oleńska, E.; Dziekoński, M.; Wojtkiewicz, J. Differentiation of Human Mesenchymal Stem Cells from Wharton’s Jelly Towards Neural Stem Cells Using a Feasible and Repeatable Protocol. Cells 2020, 9, 739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liau, L.L.; Looi, Q.H.; Chia, W.C.; Subramaniam, T.; Ng, M.H.; Law, J.X. Treatment of spinal cord injury with mesenchymal stem cells. Cell Biosci. 2020, 10, 1–17. [Google Scholar] [CrossRef]
- Voulgari-Kokota, A.; Fairless, R.; Karamita, M.; Kyrargyri, V.; Tseveleki, V.; Evangelidou, M.; Delorme, B.; Charbord, P.; Diem, R.; Probert, L. Mesenchymal stem cells protect CNS neurons against glutamate excitotoxicity by inhibiting glutamate receptor expression and function. Exp. Neurol. 2012, 236, 161–170. [Google Scholar] [CrossRef]
- Cantinieaux, D.; Quertainmont, R.; Blacher, S.; Rossi, L.; Wanet, T.; Noel, A.; Brook, G.; Schoenen, J.; Franzen, R. Conditioned Medium from Bone Marrow-Derived Mesenchymal Stem Cells Improves Recovery after Spinal Cord Injury in Rats: An Original Strategy to Avoid Cell Transplantation. PLoS ONE 2013, 8, e69515. [Google Scholar] [CrossRef]
- Crigler, L.; Robey, R.C.; Asawachaicharn, A.; Gaupp, D.; Phinney, D. Human mesenchymal stem cell subpopulations express a variety of neuro-regulatory molecules and promote neuronal cell survival and neuritogenesis. Exp. Neurol. 2006, 198, 54–64. [Google Scholar] [CrossRef]
- Aggarwal, S.; Pittenger, M.F. Human mesenchymal stem cells modulate allogeneic immune cell responses. Blood 2005, 105, 1815–1822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chopp, M.; Li, Y. Treatment of neural injury with marrow stromal cells. Lancet Neurol. 2002, 1, 92–100. [Google Scholar] [CrossRef]
- Jakob, F.; Limbert, C.; Schilling, T.; Benisch, P.; Seefried, L.; Ebert, R. Biology of Mesenchymal Stem Cells. Curr. Rheumatol. Rev. 2008, 4, 148–154. [Google Scholar] [CrossRef]
- Hamano, K.; Li, T.-S.; Kobayashi, T.; Kobayashi, S.; Matsuzaki, M.; Esato, K. Angiogenesis Induced by the Implantation of Self-Bone Marrow Cells: A New Material for Therapeutic Angiogenesis. Cell Transplant. 2000, 9, 439–443. [Google Scholar] [CrossRef]
- Park, C.W.; Kim, K.-S.; Bae, S.; Son, H.K.; Myung, P.-K.; Hong, H.J.; Kim, H. Cytokine Secretion Profiling of Human Mesenchymal Stem Cells by Antibody Array. Int. J. Stem Cells 2009, 2, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Schiff, L.; Hadker, N.; Weiser, S.; Rausch, C. A literature review of the feasibility of glial fibrillary acidic protein as a biomarker for stroke and traumatic brain injury. Mol. Diagn. Ther. 2012, 16, 79–92. [Google Scholar] [CrossRef]
- Mayer, C.A.; Brunkhorst, R.; Niessner, M.; Pfeilschifter, W.; Steinmetz, H.; Foerch, C. Blood Levels of Glial Fibrillary Acidic Protein (GFAP) in Patients with Neurological Diseases. PLoS ONE 2013, 8, e62101. [Google Scholar] [CrossRef]
- Sedaghat, F.; Notopoulos, A. S100 protein family and its application in clinical practice. Hippokratia 2008, 12, 198–204. [Google Scholar]
- Cao, F.; Yang, X.F.; Liu, W.G.; Hu, W.W.; Li, G.; Zheng, X.J.; Shen, F.; Zhao, X.Q.; Lv, S.T. Elevation of neuron-specific enolase and S-100beta protein level in experimental acute spinal cord injury. J. Clin. Neurosci. 2008, 15, 541–544. [Google Scholar] [CrossRef]
- Faridaalee, G.; Khajeh, F.K. Serum and Cerebrospinal Fluid Levels of S-100β Is A Biomarker for Spinal Cord Injury; a Systematic Review and Meta-Analysis. Arch. Acad. Emerg. Med. 2019, 7, e19. [Google Scholar]
- Shaw, G.; Yang, C.; Ellis, R.; Anderson, K.; Mickle, J.P.; Scheff, S.; Pike, B.; Anderson, D.K.; Howland, D.R. Hyperphosphorylated neurofilament NF-H is a serum biomarker of axonal injury. Biochem. Biophys. Res. Commun. 2005, 336, 1268–1277. [Google Scholar] [CrossRef] [PubMed]
- Boylan, K.B.; Glass, J.D.; Crook, J.E.; Yang, C.; Thomas, C.S.; Desaro, P.; Johnston, A.; Overstreet, K.; Kelly, C.; Polak, M.; et al. Phosphorylated neurofilament heavy subunit (pNF-H) in peripheral blood and CSF as a potential prognostic bi-omarker in amyotrophic lateral sclerosis. J. Neurol. Neurosurg. Psychiatry 2013, 84, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.B.; Wolper, R.A.; Miralia, L.; Yang, C.; Shaw, G. Detection of phosphorylated NF-H in the cerebrospinal fluid and blood of aneurysmal subarachnoid hemorrhage patients. J. Cereb. Blood Flow Metab. 2008, 28, 1261–1271. [Google Scholar] [CrossRef] [PubMed]
- St, M.I.; Ungureanu, D.; Tascu, A.; Costachescu, B.; Iencean, A.S.; Poeata, I. CSF phosphorylated neurofilament subunit NF-H (pNF-H) levels are biomarkers of Spinal Cord Injury. Rom. Neurosurg. 2013, 148–450. [Google Scholar]
- Bartlett, R.D.; Burley, S.; Ip, M.; Phillips, J.B.; Choi, D. Cell Therapies for Spinal Cord Injury: Trends and Challenges of Current Clinical Trials. Neurosurgery 2020, 87, E456–E472. [Google Scholar] [CrossRef] [PubMed]
- Karamouzian, S.; Nematollahi-Mahani, S.N.; Nakhaee, N.; Eskandary, H. Clinical safety and primary efficacy of bone marrow mesenchymal cell transplantation in subacute spinal cord injured patients. Clin. Neurol. Neurosurg. 2012, 114, 935–939. [Google Scholar] [CrossRef]
- Upadhyayula, P.S.; Martin, J.R.; Rennert, R.C.; Ciacci, J.D. Review of operative considerations in spinal cord stem cell therapy. World J. Stem Cells 2021, 13, 168–176. [Google Scholar] [CrossRef]
- Yamaguchi, S.; Horie, N.; Satoh, K.; Ishikawa, T.; Mori, T.; Maeda, H.; Fukuda, Y.; Hiu, T.; Morofuji, Y.; Izumo, T. Age of donor of human mesenchymal stem cells affects structural and functional recovery after cell therapy following ischaemic stroke. Br. J. Pharmacol. 2017, 38, 1199–1212. [Google Scholar] [CrossRef]
- Rezniczek, G.A.; Kumbruch, S.; Scheich, J.; Jensen, A.; Tempfer, C.B. Factors influencing yield and neuronal differentiation of mesenchymal stem cells from umbilical cord blood and matrix. Regen. Med. 2016, 11, 465–474. [Google Scholar] [CrossRef]
- Takahashi, H.; Aoki, Y.; Nakajima, A.; Sonobe, M.; Terajima, F.; Saito, M.; Taniguchi, S.; Yamada, M.; Watanabe, F.; Furuya, T.; et al. Phosphorylated neurofilament subunit NF-H becomes elevated in the cerebrospinal fluid of patients with acutely worsening symptoms of compression myelopathy. J. Clin. Neurosci. 2014, 21, 2175–2178. [Google Scholar] [CrossRef]
- Ahadi, R.; Khodagholi, F.; Daneshi, A.; Vafaei, A.; Mafi, A.A.; Jorjani, M. Diagnostic Value of Serum Levels of GFAP, pNF-H, and NSE Compared with Clinical Findings in Severity Assessment of Human Traumatic Spinal Cord Injury. Spine 2015, 40, E823–E830. [Google Scholar] [CrossRef] [Green Version]
- Hayakawa, K.; Okazaki, R.; Ishii, K.; Ueno, T.; Izawa, N.; Tanaka, Y.; Toyooka, S.; Matsuoka, N.; Morioka, K.; Ohori, Y.; et al. Phosphorylated neurofilament subunit NF-H as a biomarker for evaluating the severity of spinal cord injury patients, a pilot study. Spinal Cord 2012, 50, 493–496. [Google Scholar] [CrossRef]
- Singh, A.; Kumar, V.; Ali, S.; Mahdi, A.A.; Srivastava, R.N. Phosphorylated neurofilament heavy: A potential blood biomarker to evaluate the severity of acute spinal cord injuries in adults. Int. J. Crit. Illn. Inj. Sci. 2017, 7, 212–217. [Google Scholar] [CrossRef]
- Abdelhak, A.; Huss, A.; Kassubek, J.; Tumani, H.; Otto, M. Serum GFAP as a biomarker for disease severity in multiple sclerosis. Sci. Rep. 2018, 8, 1–7. [Google Scholar] [CrossRef]
- Matute-Blanch, C.; Montalban, X.; Comabella, M. Multiple sclerosis, and other demyelinating and autoimmune inflammatory diseases of the central nervous system. Cerebellum Embryol. Diagn. Investig. 2018, 146, 67–84. [Google Scholar] [CrossRef]
- Holmström, U.; Tsitsopoulos, P.P.; Holtz, A.; Salci, K.; Shaw, G.; Mondello, S.; Marklund, N. Cerebrospinal fluid levels of GFAP and pNF-H are elevated in patients with chronic spinal cord injury and neurological deterioration. Acta Neurochir. 2020, 162, 2075–2086. [Google Scholar] [CrossRef]
- Petzold, A.; Shaw, G. Comparison of two ELISA methods for measuring levels of the phosphorylated neurofilament heavy chain. J. Immunol. Methods 2007, 319, 34–40. [Google Scholar] [CrossRef]
- Harrop, J.S.; Hashimoto, R.; Norvell, D.; Raich, A.; Aarabi, B.; Grossman, R.G.; Guest, J.D.; Tator, C.H.; Chapman, J.; Fehlings, M. Evaluation of clinical experience using cell-based therapies in patients with spinal cord injury: A systematic review. J. Neurosurg. Spine 2012, 17, 230–246. [Google Scholar] [CrossRef]
- Sofroniew, M.V. Molecular dissection of reactive astrogliosis and glial scar formation. Trends Neurosci. 2009, 32, 638–647. [Google Scholar] [CrossRef] [Green Version]
- Borlongan, C.V.; Zhang, Y.; Yu, S.; Tuazon, J.P.; Lee, J.-Y.; Corey, S.; Kvederis, L.; Kingsbury, C.; Kaneko, Y. Neuroprotective effects of human bone marrow mesenchymal stem cells against cerebral ischemia are mediated in part by an anti-apoptotic mechanism. Neural Regen. Res. 2019, 14, 597–604. [Google Scholar] [CrossRef]
- Huang, W.; Lv, B.; Zeng, H.; Shi, D.; Liu, Y.; Chen, F.; Li, F.; Liu, X.; Zhu, R.; Yu, L.; et al. Paracrine Factors Secreted by MSCs Promote Astrocyte Survival Associated with GFAP Downregulation After Ischemic Stroke via p38 MAPK and JNK. J. Cell. Physiol. 2015, 230, 2461–2475. [Google Scholar] [CrossRef]
- Moore, K.W.; de Waal Malefyt, R.; Coffman, R.L.; O’Garra, A. Interleukin-10 and the interleukin-10 receptor. Annu. Rev. Immunol. 2001, 19, 683–765. [Google Scholar] [CrossRef]
- Zhou, Z.; Peng, X.; Insolera, R.; Fink, D.J.; Mata, M. Interleukin-10 provides direct trophic support to neurons. J. Neurochem. 2009, 110, 1617–1627. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Peng, X.; Insolera, R.; Fink, D.J.; Mata, M. IL-10 promotes neuronal survival following spinal cord injury. Exp. Neurol. 2009, 220, 183–190. [Google Scholar] [CrossRef] [Green Version]
- Plunkett, J.A.; Yu, C.-G.; Easton, J.M.; Bethea, J.R.; Yezierski, R.P. Effects of Interleukin-10 (IL-10) on Pain Behavior and Gene Expression Following Excitotoxic Spinal Cord Injury in the Rat. Exp. Neurol. 2001, 168, 144–154. [Google Scholar] [CrossRef]
- Yu, Z.; Sun, X.; Xia, R.; Chen, Q.; Wu, Q.; Zheng, W. Modulation of inflammatory factors predicts the outcome following spinal cord injury. J. Orthop. Surg. Res. 2020, 15, 199. [Google Scholar] [CrossRef]
- Palmer, G.; Guerne, P.A.; Mezin, F.; Maret, M.; Guicheux, J.; Goldring, M.B.; Gabay, C. Production of interleukin-1 receptor antagonist by human articular chondrocytes. Arthritis Res. 2002, 4, 226–231. [Google Scholar] [CrossRef]
- Bickel, M. The role of interleukin-8 in inflammation and mechanisms of regulation. J. Periodontol. 1993, 64, 456–460. [Google Scholar]
- Wang, L.; Li, Y.; Zhang, X.; Liu, N.; Shen, S.; Sun, S.; Jiang, Y.; Li, P.; Jin, H.; Shen, L. Paracrine interleukin-8 affects mesenchymal stem cells through the Akt pathway and enhances human umbilical vein endothelial cell proliferation and migration. Biosci. Rep. 2021, 41. [Google Scholar] [CrossRef]
- Liu, M.; Guo, S.; Hibbert, J.M.; Jain, V.; Singh, N.; Wilson, N.O.; Stiles, J.K. CXCL10/IP-10 in infectious diseases pathogenesis and potential therapeutic implications. Cytokine Growth Factor Rev. 2011, 22, 121–130. [Google Scholar] [CrossRef]
- Yu, Q.; Tian, D.L.; Tian, Y.; Zhao, X.T.; Yang, X.Y. Elevation of the Chemokine Pair CXCL10/CXCR3 Initiates Sequential Glial Activation and Crosstalk During the De-velopment of Bimodal Inflammatory Pain after Spinal Cord Ischemia Reperfusion. Cell. Physiol. Biochem. 2018, 9, 2214–2228. [Google Scholar] [CrossRef] [PubMed]
- Babcock, A.A.; Kuziel, W.A.; Rivest, S.; Owens, T. Chemokine Expression by Glial Cells Directs Leukocytes to Sites of Axonal Injury in the CNS. J. Neurosci. 2003, 23, 7922–7930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaaqoq, A.M.; Namas, R.; Almahmoud, K.; Azhar, N.; Mi, Q.; Zamora, R.; Brienza, D.M.; Billiar, T.R.; Vodovotz, Y. Inducible Protein-10, a Potential Driver of Neurally Controlled Interleukin-10 and Morbidity in Human Blunt Trauma*. Crit. Care Med. 2014, 42, 1487–1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yagura, K.; Ohtaki, H.; Tsumuraya, T.; Sato, A.; Miyamoto, K.; Kawada, N.; Suzuki, K.; Nakamura, M.; Kanzaki, K.; Dohi, K.; et al. The enhancement of CCL2 and CCL5 by human bone marrow-derived mesenchymal stem/stromal cells might contribute to inflammatory suppression and axonal extension after spinal cord injury. PLoS ONE 2020, 15, e0230080. [Google Scholar] [CrossRef] [PubMed]
- Matthews, A.N.; Friend, D.S.; Zimmermann, N.; Wert, S.E.; Rothenberg, M.E. Eotaxin is required for the baseline level of tissue eosinophils. Proc. Natl. Acad. Sci. USA 1998, 95, 6273–6278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niemi, J.; DeFrancesco-Lisowitz, A.; Cregg, J.; Howarth, M.; Zigmond, R.E. Overexpression of the monocyte chemokine CCL2 in dorsal root ganglion neurons causes a conditioning-like increase in neurite outgrowth and does so via a STAT3 dependent mechanism. Exp. Neurol. 2016, 275, 25–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Santana Nunes, A.K.; Rapôso, C.; de Oliveira, W.H.; Thomé, R.; Verinaud, L.; Tovar-Moll, F.; Peixoto, C.A. Phosphodiesterase-5 inhibition promotes remyelination by MCP-1/CCR-2 and MMP-9 regulation in a cu-prizone-induced demyelination model. Exp. Neurol. 2016, 275, 143–153. [Google Scholar] [CrossRef]
- Tokami, H.; Ago, T.; Sugimori, H.; Kuroda, J.; Awano, H.; Suzuki, K.; Kiyohara, Y.; Kamouchi, M.; Kitazono, T. RANTES has a potential to play a neuroprotective role in an autocrine/paracrine manner after ischemic stroke. Brain Res. 2013, 1517, 122–132. [Google Scholar] [CrossRef]
- Olbrich, L.; Foehring, D.; Happel, P.; Brand-Saberi, B.; Theiss, C. Fast rearrangement of the neuronal growth cone’s actin cytoskeleton following VEGF stimulation. Histochem. Cell Biol. 2013, 139, 431–445. [Google Scholar] [CrossRef]
- Schlau, M.; Terheyden-Keighley, D.; Theis, V.; Mannherz, H.G.; Theiss, C. VEGF Triggers the Activation of Cofilin and the Arp2/3 Complex within the Growth Cone. Int. J. Mol. Sci. 2018, 19, 384. [Google Scholar] [CrossRef] [Green Version]
- Silva-Hucha, S.; Pastor, A.; Morcuende, S. Neuroprotective Effect of Vascular Endothelial Growth Factor on Motoneurons of the Oculomotor System. Int. J. Mol. Sci. 2021, 22, 814. [Google Scholar] [CrossRef] [PubMed]
- Figley, S.A.; Liu, Y.; Karadimas, S.K.; Satkunendrarajah, K.; Fettes, P.; Spratt, S.K.; Lee, G.; Ando, D.; Surosky, R.; Giedlin, M.; et al. Delayed administration of a bio-engineered zinc-finger VEGF-A gene therapy is neuroprotective and attenuates allodynia following traumatic spinal cord injury. PLoS ONE 2014, 9, e96137. [Google Scholar] [CrossRef] [PubMed]
- Idrisova, K.F.; Rizvanov, A.A.; Zeinalova, A.K.; Masgutova, G.A.; Bogov, A.A.; Allegrucci, C.; Syromiatnikova, V.Y.; SalafutdinovI, I.; Garanina, E.E.; Andreeva, D.I.; et al. Application of neurotrophic and proangiogenic factors as therapy after peripheral nervous system injury. Neural Regen. Res. 2022, 17, 1240. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Neufeld, A.H. Activation of epidermal growth factor receptor causes astrocytes to form cribriform structures. Glia 2004, 46, 153–168. [Google Scholar] [CrossRef]
- Li, Z.W.; Li, J.J.; Wang, L.; Zhang, J.P.; Wu, J.J.; Mao, X.Q.; Shi, G.F.; Wang, Q.; Wang, F.; Zou, J. Epidermal growth factor receptor inhibitor ameliorates excessive astrogliosis and improves the regeneration microen-vironment and functional recovery in adult rats following spinal cord injury. J. Neuroinflamm. 2014, 11, 71. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Slovinska, L.; Harvanova, D.; Janockova, J.; Matejova, J.; Cibur, P.; Moravek, M.; Spakova, T.; Rosocha, J. Mesenchymal Stem Cells in the Treatment of Human Spinal Cord Injury: The Effect on Individual Values of pNF-H, GFAP, S100 Proteins and Selected Growth Factors, Cytokines and Chemokines. Curr. Issues Mol. Biol. 2022, 44, 578-596. https://doi.org/10.3390/cimb44020040
Slovinska L, Harvanova D, Janockova J, Matejova J, Cibur P, Moravek M, Spakova T, Rosocha J. Mesenchymal Stem Cells in the Treatment of Human Spinal Cord Injury: The Effect on Individual Values of pNF-H, GFAP, S100 Proteins and Selected Growth Factors, Cytokines and Chemokines. Current Issues in Molecular Biology. 2022; 44(2):578-596. https://doi.org/10.3390/cimb44020040
Chicago/Turabian StyleSlovinska, Lucia, Denisa Harvanova, Jana Janockova, Jana Matejova, Peter Cibur, Marko Moravek, Timea Spakova, and Jan Rosocha. 2022. "Mesenchymal Stem Cells in the Treatment of Human Spinal Cord Injury: The Effect on Individual Values of pNF-H, GFAP, S100 Proteins and Selected Growth Factors, Cytokines and Chemokines" Current Issues in Molecular Biology 44, no. 2: 578-596. https://doi.org/10.3390/cimb44020040