
Unraveling Adipose Tissue Dysfunction: Molecular Mechanisms, Novel Biomarkers, and Therapeutic Targets for Liver Fat Deposition

 and
and 
Abstract
:1. Introduction
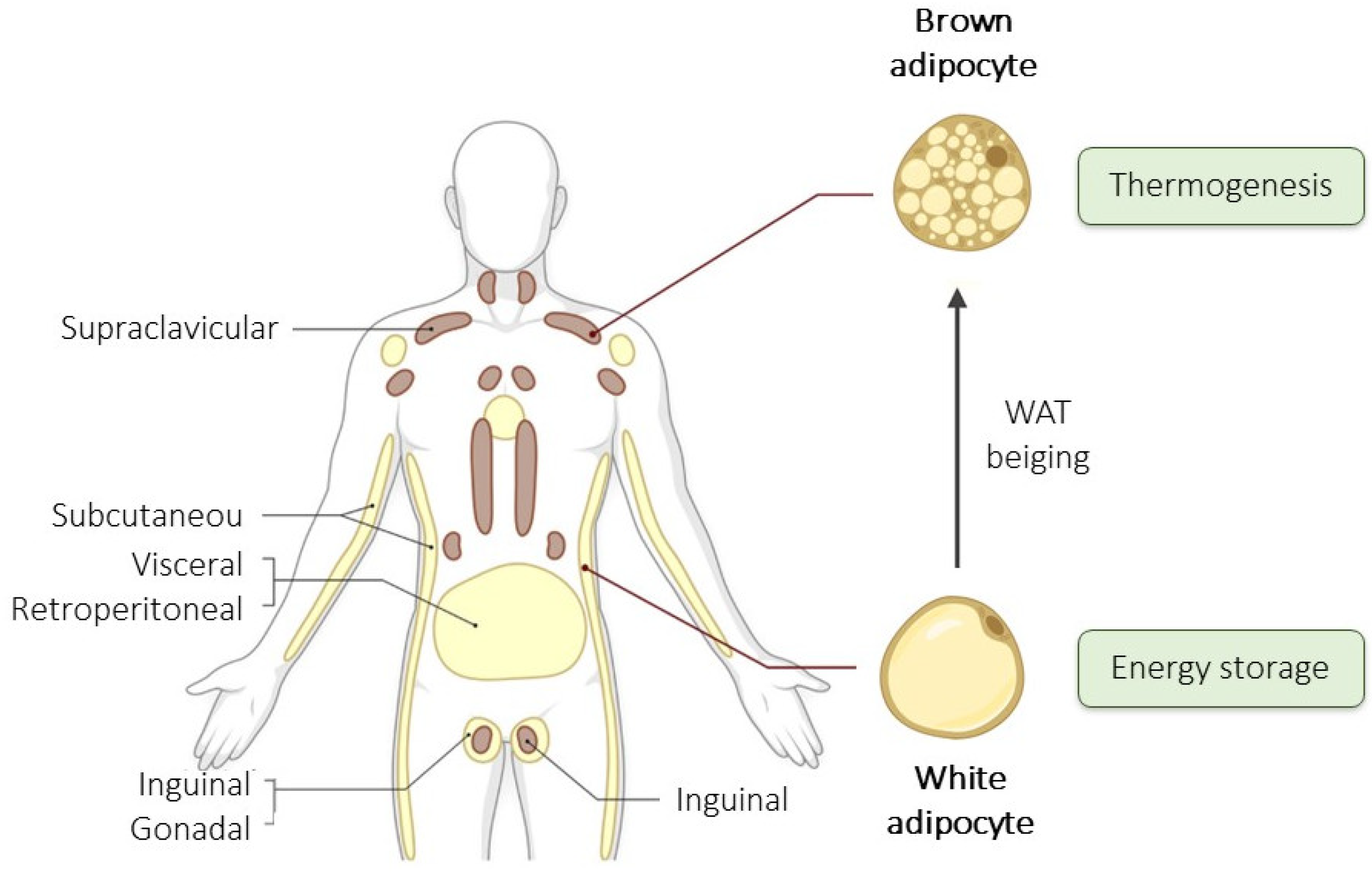
2. Types and Composition of Adipose Tissue
2.1. Types and Location of Adipose Tissue

2.2. Composition and Cellular Types of White Adipose Tissue
- Endothelial cells: Form the lining of blood vessels within AT, playing a crucial role in nutrient transport, angiogenesis, and tissue oxygenation [16].
- Blood cells: Include various types of cells involved in the immune response and the transport of oxygen and nutrients [17].
- Fibroblasts: Contribute to the structural integrity of AT, producing extracellular matrix components that support adipocytes [18].
- Pericytes: Surround endothelial cells, regulate blood flow and capillary stability, and are involved in angiogenesis [19].
- Macrophages: Particularly important in visWAT, these immune cells can contribute to inflammation in AT, especially in the context of obesity and metabolic dysfunction. They play a dual role, maintaining tissue homeostasis and mediating inflammatory responses [20].
- Immune Cells: Including T cells, B cells, and others, they are involved in the immune surveillance of AT and can contribute to inflammation and insulin resistance in obesity [21].
3. Metabolic Functions and Dysfunctions of Adipose Tisue
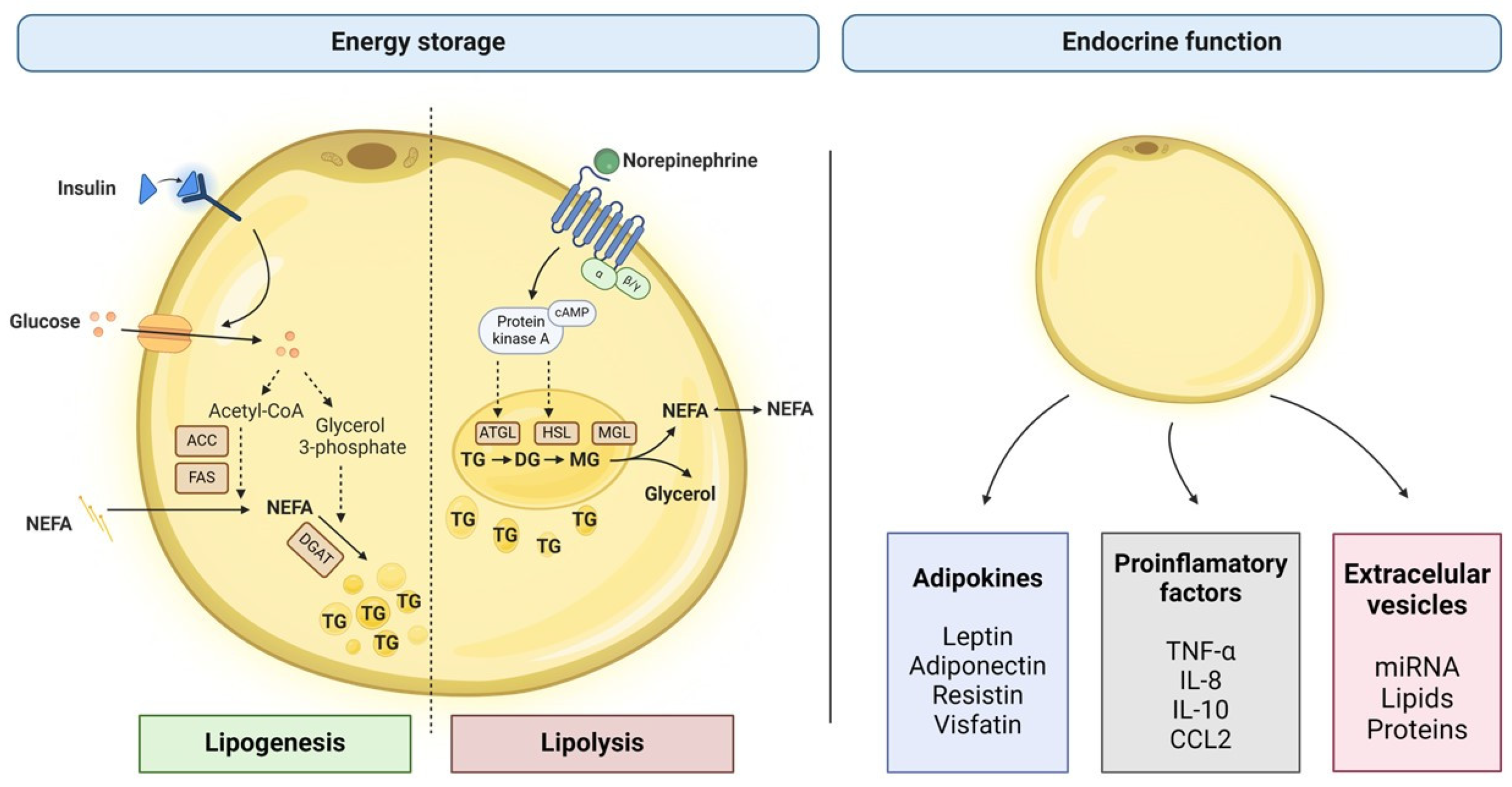
3.1. Metabolic Functions of White Adipose Tissue
3.1.1. Lipids Storage and Mobilization
3.1.2. Endocrine Function
3.2. Adipose Tissue Dysfunction in Obesity
3.2.1. Adipocyte Size and Number
3.2.2. Adipocyte Turnover
3.2.3. Insulin Sensitivity
3.2.4. Lipolytic Activity
3.2.5. Secretory Profile
3.2.6. Oxidative Stress
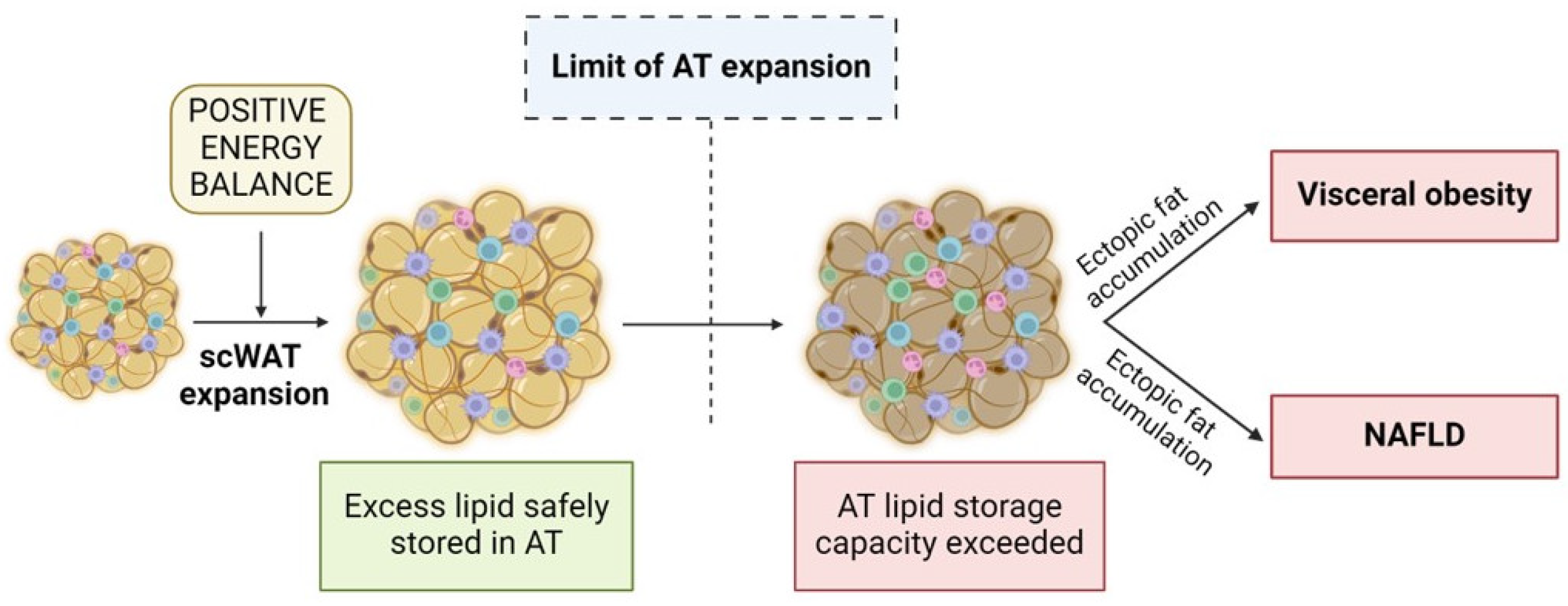
4. Adipose Tissue Expandability and Liver Fat Deposition
4.1. Connection between AT Expandability and NAFLD
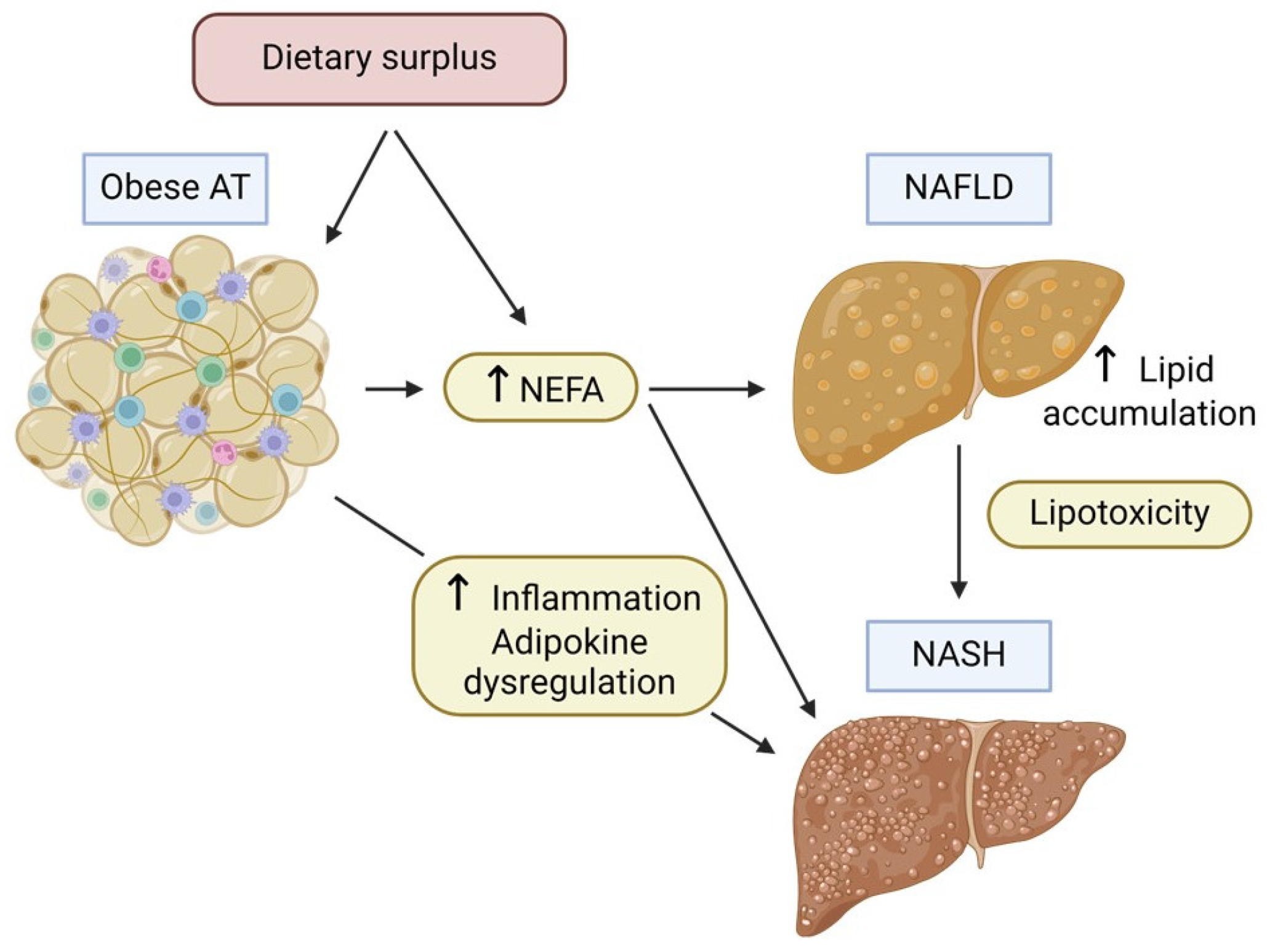
4.2. Hepatic Response to Ectopic Fat Accumulation
- Lipotoxicity: Hepatocytes exposed to high levels of lipids, particularly saturated fatty acids and other toxic lipid species such as diacylglycerol (DAG) and ceramide, undergo lipotoxic stress. This can lead to cell dysfunction and apoptosis [50].
- Inflammatory Response: Ectopic fat in the liver can trigger an inflammatory response, attracting immune cells and producing pro-inflammatory cytokines. Moreover, chronic inflammation leads to significant histological changes in the liver, such as hepatocyte necrosis and apoptosis, neutrophil chemotaxis, activation of hepatic stellate cells, and production of Mallory bodies (aggregates of cytokeratin intermediate filaments). This plays a significant role in the disease’s development from basic steatosis to NASH and fibrosis. Furthermore, persistent inflammation may promote carcinogenesis and hence contribute to the progression of the disease to hepatocarcinoma (HCC) [51].
- Oxidative Stress: When lipid flow surpasses the capacities of both mitochondria and peroxisomes, respiratory oxidation becomes compromised, resulting in disturbances in lipid homeostasis, the generation of harmful metabolites, and an excess production of reactive oxygen species (ROS) [52]. These molecules precipitate oxidative stress and exacerbate hepatic necro-inflammatory processes, further aggravating mitochondrial damage. Moreover, ROS and oxidized low-density lipoproteins (LDL) can activate Kupffer and hepatic stellate cells, thus resulting in collagen deposition and secondary liver fibrosis [53,54].
- Endoplasmic Reticulum (ER) Stress: The accumulation of lipids can disturb ER function in hepatocytes, leading to unfolded protein response and further contributing to cellular stress and apoptosis [55].
- Altered Metabolism: Hepatocytes in a fatty liver have altered carbohydrate and lipid metabolism, often associated with insulin resistance. This metabolic dysfunction can exacerbate the accumulation of lipids in the liver and impair liver function [56].
5. Diagnostic Approaches and Biomarkers for Adipose Tissue Dysfunction
5.1. Anthropometric Parameters
5.2. Imaging Techniques for Adipose Tissue Analysis
5.2.1. Bioimpedance Analysis (BIA)
5.2.2. Dual-Energy X-ray Absorptiometry (DXA)
5.2.3. Computed Tomography (CT) and Magnetic Resonance Imaging (MRI)
5.3. Circulating Biomarkers
5.3.1. Adipokines
5.3.2. Markers of Glucose-Insulin Homeostasis
5.3.3. Inflammatory Biomarkers
5.4. Omics-Based Biomarkers
5.4.1. Genomics
5.4.2. Transcriptomics
5.4.3. Metabolomics
5.5. Challenges and Future Directions
6. Therapeutic Strategies and Future Directions
6.1. Lifestyle, Surgical Options, and Pharmacological Interventions
6.2. Innovative Therapies
6.2.1. ADMSC-Based Therapies
6.2.2. Genetic Modification of ADMSC: CRISPR/Cas9 Gene Editing
6.3. Future Directions
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cinti, S. The adipose organ. Prostaglandins Leukot Essent Fat. Acids 2005, 73, 9–15. [Google Scholar] [CrossRef]
- Zwick, R.K.; Guerrero-Juarez, C.F.; Horsley, V.; Plikus, M.V. Anatomical, Physiological, and Functional Diversity of Adipose Tissue. Cell Metab. 2018, 27, 68–83. [Google Scholar] [CrossRef]
- Arner, P.; Rydén, M. Human White Adipose Tissue: A Highly Dynamic Metabolic Organ. J. Intern. Med. 2022, 291, 611–621. [Google Scholar] [CrossRef]
- Carpentier, A.C.; Blondin, D.P.; Haman, F.; Richard, D. Brown Adipose Tissue-A Translational Perspective. Endocr. Rev. 2023, 44, 143–192. [Google Scholar] [CrossRef]
- Sakers, A.; De Siqueira, M.K.; Seale, P.; Villanueva, C.J. Adipose-Tissue Plasticity in Health and Disease. Cell 2022, 185, 419–446. [Google Scholar] [CrossRef]
- Kwok, K.H.M.; Lam, K.S.L.; Xu, A. Heterogeneity of White Adipose Tissue: Molecular Basis and Clinical Implications. Exp. Mol. Med. 2016, 48, e215. [Google Scholar] [CrossRef] [PubMed]
- Saponaro, C.; Gaggini, M.; Carli, F.; Gastaldelli, A. The Subtle Balance between Lipolysis and Lipogenesis: A Critical Point in Metabolic Homeostasis. Nutrients 2015, 7, 9453–9474. [Google Scholar] [CrossRef] [PubMed]
- Scherer, P.E. Adipose Tissue: From Lipid Storage Compartment to Endocrine Organ. Diabetes 2006, 55, 1537–1545. [Google Scholar] [CrossRef] [PubMed]
- Virtue, S.; Vidal-Puig, A. Adipose Tissue Expandability, Lipotoxicity and the Metabolic Syndrome—An Allostatic Perspective. Biochim. Biophys Acta 2010, 1801, 338–349. [Google Scholar] [CrossRef] [PubMed]
- Quek, J.; Chan, K.E.; Wong, Z.Y.; Tan, C.; Tan, B.; Lim, W.H.; Tan, D.J.H.; Tang, A.S.P.; Tay, P.; Xiao, J.; et al. Global prevalence of non-alcoholic fatty liver disease and non-alcoholic steatohepatitis in the overweight and obese population: A systematic review and meta-analysis. Lancet Gastroenterol. Hepatol. 2023, 8, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Pilkington, A.C.; Paz, H.A.; Wankhade, U.D. Beige Adipose Tissue Identification and Marker Specificity—Overview. Front. Endocrinol. 2021, 12, 599134. [Google Scholar] [CrossRef]
- Ibrahim, M.M. Subcutaneous and Visceral Adipose Tissue: Structural and Functional Differences. Obes. Rev. 2010, 11, 11–18. [Google Scholar] [CrossRef]
- Patel, P.; Abate, N. Body Fat Distribution and Insulin Resistance. Nutrients 2013, 5, 2019–2027. [Google Scholar] [CrossRef]
- Torres Irizarry, V.C.; Jiang, Y.; He, Y.; Xu, P. Hypothalamic Estrogen Signaling and Adipose Tissue Metabolism in Energy Homeostasis. Front. Endocrinol. 2022, 13, 898139. [Google Scholar] [CrossRef]
- Ambele, M.A.; Dhanraj, P.; Giles, R.; Pepper, M.S. Adipogenesis: A Complex Interplay of Multiple Molecular Determinants and Pathways. Int. J. Mol. Sci. 2020, 21, 4283. [Google Scholar] [CrossRef]
- Festa, J.; AlZaim, I.; Kalucka, J. Adipose Tissue Endothelial Cells: Insights into Their Heterogeneity and Functional Diversity. Curr. Opin. Genet. Dev. 2023, 81, 102055. [Google Scholar] [CrossRef] [PubMed]
- Herold, J.; Kalucka, J. Angiogenesis in Adipose Tissue: The Interplay Between Adipose and Endothelial Cells. Front. Physiol. 2021, 11, 624903. [Google Scholar] [CrossRef]
- Ejaz, A.; Hatzmann, F.M.; Hammerle, S.; Ritthammer, H.; Mattesich, M.; Zwierzina, M.; Waldegger, P.; Zwerschke, W. Fibroblast Feeder Layer Supports Adipogenic Differentiation of Human Adipose Stromal/Progenitor Cells. Adipocyte 2019, 8, 178–189. [Google Scholar] [CrossRef]
- Ahmed, T.A.; Shousha, W.G.; Abdo, S.M.; Mohamed, I.K.; El-Badri, N. Human Adipose-Derived Pericytes: Biological Characterization and Reprogramming into Induced Pluripotent Stem Cells. Cell. Physiol. Biochem. 2020, 54, 271–286. [Google Scholar] [PubMed]
- Russo, L.; Lumeng, C.N. Properties and Functions of Adipose Tissue Macrophages in Obesity. Immunology 2018, 155, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Autieri, M.V.; Scalia, R. Adipose Tissue Inflammation and Metabolic Dysfunction in Obesity. Am. J. Physiol. Cell Physiol. 2021, 320, C375–C391. [Google Scholar] [CrossRef]
- Esteve Ràfols, M. Adipose Tissue: Cell Heterogeneity and Functional Diversity. Endocrinol. Nutr. 2014, 61, 100–112. [Google Scholar] [CrossRef]
- Massier, L.; Jalkanen, J.; Elmastas, M.; Zhong, J.; Wang, T.; Nono Nankam, P.A.; Frendo-Cumbo, S.; Bäckdahl, J.; Subramanian, N.; Sekine, T.; et al. An Integrated Single Cell and Spatial Transcriptomic Map of Human White Adipose Tissue. Nat. Commun. 2023, 14, 1438. [Google Scholar] [CrossRef]
- Luo, L.; Liu, M. Adipose Tissue in Control of Metabolism. J. Endocrinol. 2016, 231, R77–R99. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Xiaoli, A.M.; Yang, F. Regulation and Metabolic Significance of De Novo Lipogenesis in Adipose Tissues. Nutrients 2018, 10, 1383. [Google Scholar] [CrossRef] [PubMed]
- Carpentier, A.C. 100th anniversary of the Discovery of Insulin Perspective: Insulin and Adipose Tissue Fatty Acid Metabolism. Am. J. Physiol. Endocrinol. Metab. 2021, 320, E653–E670. [Google Scholar] [CrossRef]
- Grabner, G.F.; Xie, H.; Schweiger, M.; Zechner, R. Lipolysis: Cellular Mechanisms for Lipid Mobilization from Fat Stores. Nat. Metab. 2021, 3, 1445–1465. [Google Scholar] [CrossRef]
- Duncan, R.E.; Ahmadian, M.; Jaworski, K.; Sarkadi-Nagy, E.; Sul, H.S. Regulation of Lipolysis in Adipocytes. Annu. Rev. Nutr. 2007, 27, 79–101. [Google Scholar] [CrossRef]
- Lass, A.; Zimmermann, R.; Oberer, M.; Zechner, R. Lipolysis—A Highly Regulated Multi-Enzyme Complex Mediates the Catabolism of Cellular Fat Stores. Prog. Lipid Res. 2011, 50, 14–27. [Google Scholar] [CrossRef]
- Scheja, L.; Heeren, J. The Endocrine Function of Adipose Tissues in Health and Cardiometabolic Disease. Nat. Rev. Endocrinol. 2019, 15, 507–524. [Google Scholar] [CrossRef] [PubMed]
- Trayhurn, P. Endocrine and signalling role of adipose tissue: New perspectives on fat. Acta. Physiol. Scand. 2005, 184, 285–293. [Google Scholar] [CrossRef]
- Clemente-Suárez, V.J.; Redondo-Flórez, L.; Beltrán-Velasco, A.I.; Martín-Rodríguez, A.; Martínez-Guardado, I.; Navarro-Jiménez, E.; Laborde-Cárdenas, C.C.; Tornero-Aguilera, J.F. The Role of Adipokines in Health and Disease. Biomedicines 2023, 11, 1290. [Google Scholar] [CrossRef]
- Friedman, J.M. Leptin and the Endocrine Control of Energy Balance. Nat. Metab. 2019, 1, 754–764. [Google Scholar] [CrossRef] [PubMed]
- Karbowska, J.; Kochan, Z. Role of Adiponectin in the Regulation of Carbohydrate and Lipid Metabolism. J. Physiol. Pharmacol. 2006, 57 (Suppl. S6), 103–113. [Google Scholar] [PubMed]
- Al-Mansoori, L.; Al-Jaber, H.; Prince, M.S.; Elrayess, M.A. Role of Inflammatory Cytokines, Growth Factors and Adipokines in Adipogenesis and Insulin Resistance. Inflammation 2022, 45, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Jo, J.; Gavrilova, O.; Pack, S.; Jou, W.; Mullen, S.; Sumner, A.E.; Cushman, S.W.; Periwal, V. Hypertrophy and/or Hyperplasia: Dynamics of Adipose Tissue Growth. PLoS Comput. Biol. 2009, 5, e1000324. [Google Scholar] [CrossRef] [PubMed]
- Trayhurn, P. Hypoxia and Adipose Tissue Function and Dysfunction in Obesity. Physiol. Rev. 2013, 93, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Zorena, K.; Jachimowicz-Duda, O.; Ślęzak, D.; Robakowska, M.; Mrugacz, M. Adipokines and Obesity. Potential Link to Metabolic Disorders and Chronic Complications. Int. J. Mol. Sci. 2020, 21, 3570. [Google Scholar] [CrossRef] [PubMed]
- Wondmkun, Y.T. Obesity, Insulin Resistance, and Type 2 Diabetes: Associations and Therapeutic Implications. Diabetes Metab. Syndr. Obes. 2020, 13, 3611–3616. [Google Scholar] [CrossRef]
- McQuaid, S.E.; Hodson, L.; Neville, M.J.; Dennis, A.L.; Cheeseman, J.; Humphreys, S.M.; Ruge, T.; Gilbert, M.; Fielding, B.A.; Frayn, K.N.; et al. Downregulation of Adipose Tissue Fatty Acid Trafficking in Obesity: A Driver for Ectopic Fat Deposition? Diabetes 2011, 60, 47–55. [Google Scholar] [CrossRef]
- Ipsen, D.H.; Lykkesfeldt, J.; Tveden-Nyborg, P. Molecular Mechanisms of Hepatic Lipid Accumulation in Non-Alcoholic Fatty Liver Disease. Cell. Mol. Life Sci. 2018, 75, 3313–3327. [Google Scholar] [CrossRef]
- Horwitz, A.; Birk, R. Adipose Tissue Hyperplasia and Hypertrophy in Common and Syndromic Obesity—The Case of BBS Obesity. Nutrients 2023, 15, 3445. [Google Scholar] [CrossRef]
- Hocking, S.; Samocha-Bonet, D.; Milner, K.L.; Greenfield, J.R.; Chisholm, D.J. Adiposity and Insulin Resistance in Humans: The Role of the Different Tissue and Cellular Lipid Depots. Endocr. Rev. 2013, 34, 463–500. [Google Scholar] [CrossRef]
- Verboven, K.; Wouters, K.; Gaens, K.; Hansen, D.; Bijnen, M.; Wetzels, S.; Stehouwer, C.D.; Goossens, G.H.; Schalkwijk, C.G.; Blaak, E.E.; et al. Abdominal Subcutaneous and Visceral Adipocyte Size, Lipolysis and Inflammation Relate to Insulin Resistance in Male Obese Humans. Sci. Rep. 2018, 8, 1–8. [Google Scholar] [CrossRef]
- Korac, A.; Srdic-Galic, B.; Kalezic, A.; Stancic, A.; Otasevic, V.; Korac, B.; Jankovic, A. Adipokine Signatures of Subcutaneous and Visceral Abdominal Fat in Normal-Weight and Obese Women with Different Metabolic Profiles. Arch. Med. Sci. 2021, 17, 323–336. [Google Scholar] [CrossRef]
- Masschelin, P.M.; Cox, A.R.; Chernis, N.; Hartig, S.M. The Impact of Oxidative Stress on Adipose Tissue Energy Balance. Front. Physiol. 2020, 10, 1638. [Google Scholar] [CrossRef] [PubMed]
- Gray, S.L.; Vidal-Puig, A.J. Adipose Tissue Expandability in the Maintenance of Metabolic Homeostasis. Nutr. Rev. 2007, 65, 7–12. [Google Scholar] [CrossRef]
- Lee, E.; Korf, H.; Vidal-Puig, A. An Adipocentric Perspective on the Development and Progression of Non-Alcoholic Fatty Liver Disease. J. Hepatol. 2023, 78, 1048–1062. [Google Scholar] [CrossRef] [PubMed]
- Azzu, V.; Vacca, M.; Virtue, S.; Allison, M.; Vidal-Puig, A. Adipose Tissue-Liver Cross Talk in the Control of Whole-Body Metabolism: Implications in Non-alcoholic Fatty Liver Disease. Gastroenterology 2020, 158, 1899–1912. [Google Scholar] [CrossRef] [PubMed]
- Alkhouri, N.; Dixon, L.J.; Feldstein, A.E. Lipotoxicity in Non-alcoholic Fatty Liver Disease: Not All Lipids Are Created Equal. Expert. Rev. Gastroenterol. Hepatol. 2009, 3, 445–451. [Google Scholar] [CrossRef] [PubMed]
- Du Plessis, J.; Van Pelt, J.; Korf, H.; Mathieu, C.; Van Der Schueren, B.; Lannoo, M.; Oyen, T.; Topal, B.; Fetter, G.; Nayler, S.; et al. Association of Adipose Tissue Inflammation with Histologic Severity of Non-alcoholic Fatty Liver Disease. Gastroenterology 2015, 149, e14–e648. [Google Scholar] [CrossRef]
- Wang, J.; Wang, J.; He, W.; He, W.; Tsai, P.J.; Chen, P.H.; Ye, M.; Guo, J.; Su, Z. Mutual Interaction between Endoplasmic Reticulum and Mitochondria in Non-alcoholic Fatty Liver Disease. Lipids Health Dis. 2020, 19, 72. [Google Scholar] [CrossRef] [PubMed]
- Delli Bovi, A.P.; Marciano, F.; Mandato, C.; Siano, M.A.; Savoia, M.; Vajro, P. Oxidative Stress in Non-Alcoholic Fatty Liver Disease. An Updated Mini Review. Front. Med. 2021, 8, 165. [Google Scholar] [CrossRef]
- Paradies, G.; Paradies, V.; Ruggiero, F.M.; Petrosillo, G. Oxidative Stress, Cardiolipin and Mitochondrial Dysfunction in Non-alcoholic Fatty Liver Disease. World J. Gastroenterol. 2014, 20, 14205–14218. [Google Scholar] [CrossRef]
- Zhang, X.Q.; Xu, C.F.; Yu, C.H.; Chen, W.X.; Li, Y.M. Role of Endoplasmic Reticulum Stress in the Pathogenesis of Non-alcoholic Fatty Liver Disease. World J. Gastroenterol. 2014, 20, 1768–1776. [Google Scholar] [CrossRef] [PubMed]
- Jacome-Sosa, M.M.; Parks, E.J. Fatty Acid Sources and Their Fluxes as They Contribute to Plasma Triglyceride Concentrations and Fatty Liver in Humans. Curr. Opin. Lipidol. 2014, 25, 213–220. [Google Scholar] [CrossRef]
- Pujia, R.; Tarsitano, M.G.; Arturi, F.; De Lorenzo, A.; Lenzi, A.; Pujia, A.; Montalcini, T. Advances in Phenotyping Obesity and in Its Dietary and Pharmacological Treatment: A Narrative Review. Front. Nutr. 2022, 9, 804719. [Google Scholar] [CrossRef]
- Górnicka, M.; Szewczyk, K.; Białkowska, A.; Jancichova, K.; Habanova, M.; Górnicki, K.; Hamulka, J. Anthropometric Indices as Predictive Screening Tools for Obesity in Adults; The Need to Define Sex-Specific Cut-Off Points for Anthropometric Indices. Appl. Sci. 2022, 12, 6165. [Google Scholar] [CrossRef]
- Khanna, D.; Peltzer, C.; Kahar, P.; Parmar, M.S. Body Mass Index (BMI): A Screening Tool Analysis. Cureus 2022, 14, e22119. [Google Scholar] [CrossRef]
- Ross, R.; Neeland, I.J.; Yamashita, S.; Shai, I.; Seidell, J.; Magni, P.; Santos, R.D.; Arsenault, B.; Cuevas, A.; Hu, F.B.; et al. Waist Circumference as a Vital Sign in Clinical Practice: A Consensus Statement from the IAS and ICCR Working Group on Visceral Obesity. Nat. Rev. Endocrinol. 2020, 16, 177–189. [Google Scholar] [CrossRef]
- Wall-Medrano, A.; Faramarzi, E.; Romero-Saldaña, M.; Es, Z. Diagnostic Accuracy of the Waist-to-Height Ratio and Other Anthropometric Indices for Metabolically Healthy Obesity in the Working Population. Front. Nutr. 2022, 9, 962054. [Google Scholar]
- Sommer, I.; Teufer, B.; Szelag, M.; Nussbaumer-Streit, B.; Titscher, V.; Klerings, I.; Gartlehner, G. The Performance of Anthropometric Tools to Determine Obesity: A Systematic Review and Meta-Analysis. Sci. Rep. 2020, 10, 12699. [Google Scholar] [CrossRef] [PubMed]
- Piqueras, P.; Ballester, A.; Durá-Gil, J.V.; Martinez-Hervas, S.; Redón, J.; Real, J.T. Anthropometric Indicators as a Tool for Diagnosis of Obesity and Other Health Risk Factors: A Literature Review. Front. Psychol. 2021, 12, 631179. [Google Scholar] [CrossRef]
- Kim, S.R.; Lerman, L.O. Diagnostic Imaging in the Management of Patients with Metabolic Syndrome. Transl. Res. 2018, 194, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Shuster, A.; Patlas, M.; Pinthus, J.H.; Mourtzakis, M. The Clinical Importance of Visceral Adiposity: A Critical Review of Methods for Visceral Adipose Tissue Analysis. Br. J. Radiol. 2012, 85, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Brunani, A.; Perna, S.; Soranna, D.; Rondanelli, M.; Zambon, A.; Bertoli, S.; Vinci, C.; Capodaglio, P.; Lukaski, H.; Cancello, R. Body Composition Assessment Using Bioelectrical Impedance Analysis (BIA) in a Wide Cohort of Patients Affected with Mild to Severe Obesity. Clin. Nutr. 2021, 40, 3973–3981. [Google Scholar] [CrossRef]
- Onuma, T.; Kamishima, T.; Sasaki, T.; Sakata, M. Absolute Reliability of Adipose Tissue Volume Measurement by Computed Tomography: Application of Low-Dose Scan and Minimal Detectable Change—A Phantom Study. Radiol. Phys. Technol. 2015, 8, 312–319. [Google Scholar] [CrossRef]
- Kjønigsen, L.J.; Harneshaug, M.; Fløtten, A.M.; Karterud, L.K.; Petterson, K.; Skjolde, G.; Eggesbø, H.B.; Weedon-Fekjær, H.; Henriksen, H.B.; Lauritzen, P.M. Reproducibility of Semiautomated Body Composition Segmentation of Abdominal Computed Tomography: A Multiobserver Study. Eur. Radiol. Exp. 2019, 3, 42. [Google Scholar] [CrossRef]
- Fukuda, T.; Bouchi, R.; Takeuchi, T.; Nakano, Y.; Murakami, M.; Minami, I.; Izumiyama, H.; Hashimoto, K.; Yoshimoto, T.; Ogawa, Y. Ratio of Visceral-to-Subcutaneous Fat Area Predicts Cardiovascular Events in Patients with Type 2 Diabetes. J. Diabetes Investig. 2018, 9, 396–402. [Google Scholar] [CrossRef]
- Kwon, S.H.; Han, A.L. The Correlation between the Ratio of Visceral Fat Area to Subcutaneous Fat Area on Computed Tomography and Lipid Accumulation Product as Indexes of Cardiovascular Risk. J. Obes. Metab. Syndr. 2019, 28, 186–193. [Google Scholar] [CrossRef]
- Ladeiras-Lopes, R.; Sampaio, F.; Bettencourt, N.; Fontes-Carvalho, R.; Ferreira, N.; Leite-Moreira, A.; Gama, V. The Ratio Between Visceral and Subcutaneous Abdominal Fat Assessed by Computed Tomography Is an Independent Predictor of Mortality and Cardiac Events. Rev. Española De Cardiol. (Engl. Ed.) 2017, 70, 331–337. [Google Scholar] [CrossRef]
- Lopez-Yus, M.; Casamayor, C.; Soriano-Godes, J.J.; Borlan, S.; Gonzalez-Irazabal, Y.; Garcia-Sobreviela, M.P.; Garcia-Rodriguez, B.; del Moral-Bergos, R.; Calmarza, P.; Artigas, J.M.; et al. Isthmin-1 (ISM1), a Novel Adipokine That Reflects Abdominal Adipose Tissue Distribution in Individuals with Obesity. Cardiovasc. Diabetol. 2023, 22, 335. [Google Scholar] [CrossRef]
- Cinteza, E.E.; Cinteza, M. Biomarkers in Obesity. Rev. Rom. Med. Lab. 2018, 26, 353–358. [Google Scholar] [CrossRef]
- Nimptsch, K.; Konigorski, S.; Pischon, T. Diagnosis of Obesity and Use of Obesity Biomarkers in Science and Clinical Medicine. Metabolism 2019, 92, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Aleksandrova, K.; Mozaffarian, D.; Pischon, T. Addressing the Perfect Storm: Biomarkers in Obesity and Pathophysiology of Cardiometabolic Risk. Clin. Chem. 2018, 64, 142–153. [Google Scholar] [CrossRef] [PubMed]
- Unamuno, X.; Gómez-Ambrosi, J.; Rodríguez, A.; Becerril, S.; Frühbeck, G.; Catalán, V. Adipokine Dysregulation and Adipose Tissue Inflammation in Human Obesity. Eur. J. Clin. Investig. 2018, 48, e12997. [Google Scholar] [CrossRef] [PubMed]
- Würfel, M.; Blüher, M.; Stumvoll, M.; Ebert, T.; Kovacs, P.; Tönjes, A.; Breitfeld, J. Adipokines as Clinically Relevant Therapeutic Targets in Obesity. Biomedicines 2023, 11, 1427. [Google Scholar] [CrossRef] [PubMed]
- Liuzzi, A.; Savia, G.; Tagliaferri, M.; Lucantoni, R.; Petroni, M.L.; De Medici, C.; Viberti, G.C. Serum Leptin Concentration in Moderate and Severe Obesity: Relationship with Clinical, Anthropometric and Metabolic Factors. Int. J. Obes. Relat. Metab. Disord. 1999, 23, 1066–1073. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, H.; Shimomura, Y.; Hayashi, R.; Ohtani, K.; Sato, N.; Futawatari, T.; Mori, M. Serum Leptin Concentration Is Associated with Total Body Fat Mass, but Not Abdominal Fat Distribution. Int. J. Obes. Relat. Metab. Disord. 1997, 21, 536–541. [Google Scholar] [CrossRef] [PubMed]
- Perakakis, N.; Farr, O.M.; Mantzoros, C.S. Leptin in Leanness and Obesity: JACC State-of-the-Art Review. J. Am. Coll. Cardiol. 2021, 77, 745–760. [Google Scholar] [CrossRef]
- Gruber, T.; Pan, C.; Contreras, R.E.; Wiedemann, T.; Morgan, D.A.; Skowronski, A.A.; Lefort, S.; De Bernardis Murat, C.; Le Thuc, O.; Legutko, B.; et al. Obesity-Associated Hyperleptinemia Alters the Gliovascular Interface of the Hypothalamus to Promote Hypertension. Cell Metab. 2021, 33, 1155–1170.e10. [Google Scholar] [CrossRef] [PubMed]
- Poetsch, M.S.; Strano, A.; Guan, K. Role of Leptin in Cardiovascular Diseases. Front. Endocrinol. 2020, 11, 354. [Google Scholar] [CrossRef] [PubMed]
- Chai, S.B.; Sun, F.; Nie, X.L.; Wang, J. Leptin and Coronary Heart Disease: A Systematic Review and Meta-Analysis. Atherosclerosis 2014, 233, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Kishida, K.; Funahashi, T.; Shimomura, I. Adiponectin as a Routine Clinical Biomarker. Best Pract. Res. Clin. Endocrinol. Metab. 2014, 28, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.Y.; Huang, K.C.; Chang, L.C.; Huang, Y.S.; Chi, Y.C.; Su, T.C.; Chen, C.L.; Yang, W.S. Adiponectin: A Biomarker of Obesity-Induced Insulin Resistance in Adipose Tissue and Beyond. J. Biomed. Sci. 2008, 15, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Hong, X.; Zhang, X.; You, L.; Li, F.; Lian, H.; Wang, J.; Mao, N.; Ren, M.; Li, Y.; Wang, C.; et al. Association between Adiponectin and Newly Diagnosed Type 2 Diabetes in Population with the Clustering of Obesity, Dyslipidaemia and Hypertension: A Cross-Sectional Study. BMJ Open 2023, 13, e060377. [Google Scholar] [CrossRef]
- Srikanthan, K.; Feyh, A.; Visweshwar, H.; Shapiro, J.I.; Sodhi, K. Systematic Review of Metabolic Syndrome Biomarkers: A Panel for Early Detection, Management, and Risk Stratification in the West Virginian Population. Int. J. Med. Sci. 2016, 13, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Liang, S.; Que, S.; Zhou, L.; Zheng, S.; Mardinoglu, A. Meta-Analysis of Adiponectin as a Biomarker for the Detection of Metabolic Syndrome. Front. Physiol. 2018, 9, 1238. [Google Scholar] [CrossRef]
- Marques, V.; Afonso, M.B.; Bierig, N.; Duarte-Ramos, F.; Santos-Laso, Á.; Jimenez-Agüero, R.; Eizaguirre, E.; Bujanda, L.; Pareja, M.J.; Luís, R.; et al. Adiponectin, Leptin, and IGF-1 Are Useful Diagnostic and Stratification Biomarkers of NAFLD. Front. Med. 2021, 8, 683250. [Google Scholar] [CrossRef]
- Barrios, V.; Gómez-Huelgas, R.; Rodríguez, R.; de Pablos-Velasco, P.; Barrios Alonso, V. Adiponectin: An Emerging Cardiovascular Risk Factor. Rev. Esp. Cardiol. 2008, 61, 1159–1167. [Google Scholar] [CrossRef]
- Djoussé, L.; Bartz, T.M.; Ix, J.H.; Kochar, J.; Kizer, J.R.; Gottdiener, J.S.; Tracy, R.P.; Mozaffarian, D.; Siscovick, D.S.; Mukamal, K.J.; et al. Fatty Acid-Binding Protein 4 and Incident Heart Failure: The Cardiovascular Health Study. Eur. J. Heart Fail. 2013, 15, 394–399. [Google Scholar] [CrossRef] [PubMed]
- Romacho, T.; Sánchez-Ferrer, C.F.; Peiró, C. Visfatin/Nampt: An Adipokine with Cardiovascular Impact. Mediat. Inflamm. 2013, 2013, 946427. [Google Scholar] [CrossRef]
- Kahn, S.E.; Hull, R.L.; Utzschneider, K.M. Mechanisms Linking Obesity to Insulin Resistance and Type 2 Diabetes. Nature 2006, 444, 840–846. [Google Scholar] [CrossRef] [PubMed]
- Wiebe, N.; Muntner, P.; Tonelli, M. Associations of Body Mass Index, Fasting Insulin, and Inflammation with Mortality: A Prospective Cohort Study. Int. J. Obes. 2022, 46, 2107–2113. [Google Scholar] [CrossRef] [PubMed]
- Xun, P.; Wu, Y.; He, Q.; He, K. Fasting Insulin Concentrations and Incidence of Hypertension, Stroke, and Coronary Heart Disease: As Meta-Analysis of Prospective Cohort Studies 1–3. Am. J. Clin. Nutr. 2013, 98, 1543–1554. [Google Scholar] [CrossRef]
- Patel, N.; Taveira, T.H.; Choudhary, G.; Whitlatch, H.; Wu, W.C. Fasting Serum C-Peptide Levels Predict Cardiovascular and Overall Death in Non-diabetic Adults. J. Am. Heart Assoc. 2012, 1, e003152. [Google Scholar] [CrossRef]
- Pikkemaat, M.; Melander, O.; Mölstad, S.; Garberg, G.; Boström, K.B. C-Peptide Concentration, Mortality and Vascular Complications in People with Type 2 Diabetes. The Skaraborg Diabetes Register. Diabet. Med. 2015, 32, 85–89. [Google Scholar] [CrossRef]
- Talia, C.; Connolly, L.; Fowler, P.A. The Insulin-like Growth Factor System: A Target for Endocrine Disruptors? Environ. Int. 2021, 147, 106311. [Google Scholar] [CrossRef]
- Sandhu, M.S.; Heald, A.H.; Gibson, J.M.; Cruickshank, J.K.; Dunger, D.B.; Wareham, N.J. Circulating Concentrations of Insulin-like Growth Factor-I and Development of Glucose Intolerance: A Prospective Observational Study. The Lancet 2002, 359, 1740–1745. [Google Scholar] [CrossRef]
- Drogan, D.; Schulze, M.B.; Boeing, H.; Pischon, T. Insulin-Like Growth Factor 1 and Insulin-Like Growth Factor-Binding Protein 3 in Relation to the Risk of Type 2 Diabetes Mellitus: Results from the EPIC-Potsdam Study. Am. J. Epidemiol. 2016, 183, 553–560. [Google Scholar] [CrossRef]
- Khanna, D.; Khanna, S.; Khanna, P.; Kahar, P.; Patel, B.M. Obesity: A Chronic Low-Grade Inflammation and Its Markers. Cureus 2022, 14, e22711. [Google Scholar] [CrossRef]
- Graßmann, S.; Wirsching, J.; Eichelmann, F.; Aleksandrova, K. Association Between Peripheral Adipokines and Inflammation Markers: A Systematic Review and Meta-Analysis. Obesity 2017, 25, 1776–1785. [Google Scholar] [CrossRef] [PubMed]
- Emerging Risk Factors Collaboration; Kaptoge, S.; Di Angelantonio, E.; Lowe, G.; Pepys, M.B.; Thompson, S.G.; Collins, R.; Danesh, J. C-reactive protein concentration and risk of coronary heart disease, stroke, and mortality: An individual participant meta-analysis. Lancet 2010, 375, 132–140. [Google Scholar] [PubMed]
- Zhang, Q.; Meng, X.H.; Qiu, C.; Shen, H.; Zhao, Q.; Zhao, L.J.; Tian, Q.; Sun, C.Q.; Deng, H.W. Integrative Analysis of Multi-Omics Data to Detect the Underlying Molecular Mechanisms for Obesity in Vivo in Humans. Hum. Genom. 2022, 16, 022–00388. [Google Scholar] [CrossRef] [PubMed]
- Aleksandrova, K.; Egea Rodrigues, C.; Floegel, A.; Ahrens, W. Omics Biomarkers in Obesity: Novel Etiological Insights and Targets for Precision Prevention. Curr. Obes. Rep. 2020, 9, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Loos, R.J.F.; Yeo, G.S.H. The Genetics of Obesity: From Discovery to Biology. Nat. Rev. Genet. 2022, 23, 120–133. [Google Scholar] [CrossRef] [PubMed]
- Loos, R.J.F.; Janssens, A.C.J.W. Predicting Polygenic Obesity Using Genetic Information. Cell Metab. 2017, 25, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.J.; Geisler, C.; Blundell, J.; Dulloo, A.; Schutz, Y.; Krawczak, M.; Bosy-Westphal, A.; Enderle, J.; Heymsfield, S.B. The Case of GWAS of Obesity: Does Body Weight Control Play by the Rules? Int. J. Obes. 2018, 42, 1395–1405. [Google Scholar] [CrossRef] [PubMed]
- Yengo, L.; Sidorenko, J.; Kemper, K.E.; Zheng, Z.; Wood, A.R.; Weedon, M.N.; Frayling, T.M.; Hirschhorn, J.; Yang, J.; Visscher, P.M. Meta-Analysis of Genome-Wide Association Studies for Height and Body Mass Index in ~700 000 Individuals of European Ancestry. Hum. Mol. Genet. 2018, 27, 3641–3649. [Google Scholar] [CrossRef]
- Locke, A.E.; Kahali, B.; Berndt, S.I.; Justice, A.E.; Pers, T.H.; Day, F.R.; Powell, C.; Vedantam, S.; Buchkovich, M.L.; Yang, J.; et al. Genetic Studies of Body Mass Index Yield New Insights for Obesity Biology. Nature 2015, 518, 197–206. [Google Scholar] [CrossRef]
- Fawcett, K.A.; Barroso, I. The Genetics of Obesity: FTO Leads the Way. Trends Genet. 2010, 26, 266–274. [Google Scholar] [CrossRef]
- Ehrlich, A.C.; Friedenberg, F.K. Genetic Associations of Obesity: The Fat-Mass and Obesity-Associated (FTO) Gene. Clin. Transl. Gastroenterol. 2016, 7, e140. [Google Scholar] [CrossRef]
- Melhorn, S.J.; Askren, M.K.; Chung, W.K.; Kratz, M.; Bosch, T.A.; Tyagi, V.; Webb, M.F.; De Leon, M.R.B.; Grabowski, T.J.; Leibel, R.L.; et al. FTO Genotype Impacts Food Intake and Corticolimbic Activation. Am. J. Clin. Nutr. 2018, 107, 145–154. [Google Scholar] [CrossRef]
- Magno, F.C.C.M.; Guaraná, H.C.; Fonseca, A.C.P.; Cabello, G.M.K.; Carneiro, J.R.I.; Pedrosa, A.P.; Ximenes, A.C.; Rosado, E.L. Influence of FTO Rs9939609 Polymorphism on Appetite, Ghrelin, Leptin, IL6, TNFα Levels, and Food Intake of Women with Morbid Obesity. Diabetes Metab. Syndr. Obes. 2018, 11, 199–207. [Google Scholar] [CrossRef]
- Cecil, J.E.; Tavendale, R.; Watt, P.; Hetherington, M.M.; Palmer, C.N.A. An Obesity-Associated FTO Gene Variant and Increased Energy Intake in Children. N. Engl. J. Med. 2008, 359, 2558–2566. [Google Scholar] [CrossRef]
- Speakman, J.R. FTO Effect on Energy Demand versus Food Intake. Nature 2010, 464, E1. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Y.; Shie, S.S.; Hsieh, I.C.; Tsai, M.L.; Wen, M.S. FTO Modulates Circadian Rhythms and Inhibits the CLOCK-BMAL1-Induced Transcription. Biochem. Biophys Res. Commun. 2015, 464, 826–832. [Google Scholar] [CrossRef] [PubMed]
- Lan, N.; Lu, Y.; Zhang, Y.; Pu, S.; Xi, H.; Nie, X.; Liu, J.; Yuan, W. FTO—A Common Genetic Basis for Obesity and Cancer. Front. Genet. 2020, 11, 559138. [Google Scholar] [CrossRef] [PubMed]
- Smemo, S.; Tena, J.J.; Kim, K.H.; Gamazon, E.R.; Sakabe, N.J.; Gómez-Marín, C.; Aneas, I.; Credidio, F.L.; Sobreira, D.R.; Wasserman, N.F.; et al. Obesity-Associated Variants within FTO Form Long-Range Functional Connections with IRX3. Nature 2014, 507, 371–375. [Google Scholar] [CrossRef]
- Fan, S.H.; Say, Y.H. Leptin and Leptin Receptor Gene Polymorphisms and Their Association with Plasma Leptin Levels and Obesity in a Multi-Ethnic Malaysian Suburban Population. J. Physiol. Anthropol. 2014, 33, 15. [Google Scholar] [CrossRef] [PubMed]
- Howlader, M.; Sultana, M.I.; Akter, F.; Hossain, M.M. Adiponectin Gene Polymorphisms Associated with Diabetes Mellitus: A Descriptive Review. Heliyon 2021, 7, e07851. [Google Scholar] [CrossRef] [PubMed]
- Nasias, D.; Dalakoura-Karagkouni, K.; Vassou, D.; Papagiannakis, G.; Papadaki, A.; Kardassis, D. Transcriptome Analysis of the Adipose Tissue in a Mouse Model of Metabolic Syndrome Identifies Gene Signatures Related to Disease Pathogenesis. Genomics 2020, 112, 4053–4062. [Google Scholar] [CrossRef] [PubMed]
- Hildreth, A.D.; Ma, F.; Wong, Y.Y.; Sun, R.; Pellegrini, M.; O’Sullivan, T.E. Single-cell sequencing of human white adipose tissue identifies new cell states in health and obesity. Nat. Immunol. 2021, 22, 639–653. [Google Scholar] [CrossRef] [PubMed]
- Bäckdahl, J.; Franzén, L.; Massier, L.; Li, Q.; Jalkanen, J.; Gao, H.; Andersson, A.; Bhalla, N.; Thorell, A.; Rydén, M.; et al. Spatial mapping reveals human adipocyte subpopulations with distinct sensitivities to insulin. Cell Metab. 2021, 33, 2301. [Google Scholar] [CrossRef]
- Homuth, G.; Wahl, S.; Müller, C.; Schurmann, C.; Mäder, U.; Blankenberg, S.; Carstensen, M.; Dörr, M.; Endlich, K.; Englbrecht, C.; et al. Extensive Alterations of the Whole-Blood Transcriptome Are Associated with Body Mass Index: Results of an MRNA Profiling Study Involving Two Large Population-Based Cohorts. BMC Med. Genom. 2015, 8, 65. [Google Scholar] [CrossRef]
- Ghosh, S.; Dent, R.; Harper, M.E.; Gorman, S.A.; Stuart, J.S.; McPherson, R. Gene Expression Profiling in Whole Blood Identifies Distinct Biological Pathways Associated with Obesity. BMC Med. Genom. 2010, 3, 56. [Google Scholar] [CrossRef] [PubMed]
- Mattick, J.S. Non-Coding RNAs: The Architects of Eukaryotic Complexity. EMBO Rep. 2001, 2, 986–991. [Google Scholar] [CrossRef]
- Iacomino, G.; Siani, A. Role of MicroRNAs in Obesity and Obesity-Related Diseases. Genes Nutr. 2017, 12, 23. [Google Scholar] [CrossRef]
- Jean-François, L.; Derghal, A.; Mounien, L. Micrornas in Obesity and Related Metabolic Disorders. Cells 2019, 8, 859. [Google Scholar]
- Ortiz-Dosal, A.; Rodil-García, P.; Salazar-Olivo, L.A. Circulating MicroRNAs in Human Obesity: A Systematic Review. Biomarkers 2019, 24, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Ji, C.; Guo, X. The Clinical Potential of Circulating MicroRNAs in Obesity. Nat. Rev. Endocrinol. 2019, 15, 731–743. [Google Scholar] [CrossRef]
- Rangel-Huerta, O.D.; Pastor-Villaescusa, B.; Gil, A. Are We Close to Defining a Metabolomic Signature of Human Obesity? A Systematic Review of Metabolomics Studies. Metabolomics 2019, 15, 93. [Google Scholar] [CrossRef]
- Floegel, A.; Wientzek, A.; Bachlechner, U.; Jacobs, S.; Drogan, D.; Prehn, C.; Adamski, J.; Krumsiek, J.; Schulze, M.B.; Pischon, T.; et al. Linking Diet, Physical Activity, Cardiorespiratory Fitness and Obesity to Serum Metabolite Networks: Findings from a Population-Based Study. Int. J. Obes. 2014, 38, 1388–1396. [Google Scholar] [CrossRef]
- Alves, A.; Bassot, A.; Bulteau, A.L.; Pirola, L.; Morio, B. Glycine Metabolism and Its Alterations in Obesity and Metabolic Diseases. Nutrients 2019, 11, 56. [Google Scholar] [CrossRef]
- Tulipani, S.; Palau-Rodriguez, M.; Miñarro Alonso, A.; Cardona, F.; Marco-Ramell, A.; Zonja, B.; Lopez de Alda, M.; Muñoz-Garach, A.; Sanchez-Pla, A.; Tinahones, F.J.; et al. Biomarkers of Morbid Obesity and Prediabetes by Metabolomic Profiling of Human Discordant Phenotypes. Clinica Chim. Acta 2016, 463, 53–61. [Google Scholar] [CrossRef]
- Siddik, M.A.B.; Shin, A.C. Recent Progress on Branched-Chain Amino Acids in Obesity, Diabetes, and Beyond. Endocrinol. Metab. 2019, 34, 234–246. [Google Scholar] [CrossRef]
- Wenk, M.R. Lipidomics: New Tools and Applications. Cell 2010, 143, 888–895. [Google Scholar] [CrossRef] [PubMed]
- Mousa, A.; Naderpoor, N.; Mellett, N.; Wilson, K.; Plebanski, M.; Meikle, P.J.; de Courten, B. Lipidomic Profiling Reveals Early-Stage Metabolic Dysfunction in Overweight or Obese Humans. Biochim. Biophys Acta Mol. Cell Biol. Lipids 2019, 1864, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Klop, B.; Elte, J.W.F.; Cabezas, M.C. Dyslipidemia in Obesity: Mechanisms and Potential Targets. Nutrients 2013, 5, 1218–1240. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Vinces, S.; Garcia, P.H.D.; Silva, A.A.R.; de Fernandes, A.M.A.P.; Barreto, J.A.; Duarte, G.H.B.; Antonio, M.A.; Birbrair, A.; Porcari, A.M.; de Carvalho, P.O. Mass-Spectrometry-Based Lipidomics Discriminates Specific Changes in Lipid Classes in Healthy and Dyslipidemic Adults. Metabolites 2023, 13, 222. [Google Scholar] [CrossRef] [PubMed]
- Mihalik, S.J.; Goodpaster, B.H.; Kelley, D.E.; Chace, D.H.; Vockley, J.; Toledo, F.G.; DeLany, J.P. Increased levels of plasma acylcarnitines in obesity and type 2 diabetes and identification of a marker of glucolipotoxicity. Obesity 2010, 18, 1695–1700. [Google Scholar] [CrossRef] [PubMed]
- Guasch-Ferré, M.; Ruiz-Canela, M.; Li, J.; Zheng, Y.; Bulló, M.; Wang, D.D.; Toledo, E.; Clish, C.; Corella, D.; Estruch, R.; et al. Plasma Acylcarnitines and Risk of Type 2 Diabetes in a Mediterranean Population at High Cardiovascular Risk. J. Clin. Endocrinol. Metab. 2019, 104, 1508–1519. [Google Scholar] [CrossRef]
- Spiller, S.; Blüher, M.; Hoffmann, R. Plasma Levels of Free Fatty Acids Correlate with Type 2 Diabetes Mellitus. Diabetes Obes. Metab. 2018, 20, 2661–2669. [Google Scholar] [CrossRef] [PubMed]
- Xia, Q.; Campbell, J.A.; Ahmad, H.; Si, L.; de Graaff, B.; Palmer, A.J. Bariatric Surgery Is a Cost-Saving Treatment for Obesity—A Comprehensive Meta-Analysis and Updated Systematic Review of Health Economic Evaluations of Bariatric Surgery. Obes. Rev. 2020, 21, e12932. [Google Scholar] [CrossRef] [PubMed]
- Wilding, J.P.H.; Batterham, R.L.; Calanna, S.; Davies, M.; Van Gaal, L.F.; Lingvay, I.; McGowan, B.M.; Rosenstock, J.; Tran, M.T.D.; Wadden, T.A.; et al. Once-Weekly Semaglutide in Adults with Overweight or Obesity. N. Engl. J. Med. 2021, 384, 989–1002. [Google Scholar] [CrossRef]
- Jastreboff, A.M.; Aronne, L.J.; Ahmad, N.N.; Wharton, S.; Connery, L.; Alves, B.; Kiyosue, A.; Zhang, S.; Liu, B.; Bunck, M.C.; et al. Tirzepatide Once Weekly for the Treatment of Obesity. N. Engl. J. Med. 2022, 387, 205–216. [Google Scholar] [CrossRef]
- Lin, X.; Li, H. Obesity: Epidemiology, Pathophysiology, and Therapeutics. Front. Endocrinol. 2021, 12, 706978. [Google Scholar] [CrossRef]
- Lopez-Yus, M.; García-Sobreviela, M.P.; del Moral-Bergos, R.; Arbones-Mainar, J.M. Gene Therapy Based on Mesenchymal Stem Cells Derived from Adipose Tissue for the Treatment of Obesity and Its Metabolic Complications. Int. J. Mol. Sci. 2023, 24, 7468. [Google Scholar] [CrossRef]
- Pittenger, M.F.; Discher, D.E.; Péault, B.M.; Phinney, D.G.; Hare, J.M.; Caplan, A.I. Mesenchymal Stem Cell Perspective: Cell Biology to Clinical Progress. NPJ Regen. Med. 2019, 4, 22. [Google Scholar] [CrossRef]
- Andrzejewska, A.; Lukomska, B.; Janowski, M. Concise Review: Mesenchymal Stem Cells: From Roots to Boost. Stem. Cells 2019, 37, 855–864. [Google Scholar] [CrossRef]
- Jovic, D.; Yu, Y.; Wang, D.; Wang, K.; Li, H.; Xu, F.; Liu, C.; Liu, J.; Luo, Y. A Brief Overview of Global Trends in MSC-Based Cell Therapy. Stem. Cell Rev. Rep. 2022, 18, 1525–1545. [Google Scholar] [CrossRef]
- Munir, H.; Ward, L.S.C.; Sheriff, L.; Kemble, S.; Nayar, S.; Barone, F.; Nash, G.B.; McGettrick, H.M. Adipogenic Differentiation of Mesenchymal Stem Cells Alters Their Immunomodulatory Properties in a Tissue-Specific Manner. Stem. Cells 2017, 35, 1636–1646. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, X.; Cao, W.; Shi, Y. Plasticity of Mesenchymal Stem Cells in Immunomodulation: Pathological and Therapeutic Implications. Nat. Immunol. 2014, 15, 1009–1016. [Google Scholar] [CrossRef]
- Yang, X.; Meng, Y.; Han, Z.; Ye, F.; Wei, L.; Zong, C. Mesenchymal Stem Cell Therapy for Liver Disease: Full of Chances and Challenges. Cell Biosci. 2020, 10, 123. [Google Scholar] [CrossRef]
- Melief, S.M.; Zwaginga, J.J.; Fibbe, W.E.; Roelofs, H. Adipose Tissue-Derived Multipotent Stromal Cells Have a Higher Immunomodulatory Capacity Than Their Bone Marrow-Derived Counterparts. Stem. Cells Transl. Med. 2013, 2, 455–463. [Google Scholar] [CrossRef]
- Cao, M.; Pan, Q.; Dong, H.; Yuan, X.; Li, Y.; Sun, Z.; Dong, X.; Wang, H. Adipose-Derived Mesenchymal Stem Cells Improve Glucose Homeostasis in High-Fat Diet-Induced Obese Mice. Stem. Cell Res. Ther. 2015, 12, 277. [Google Scholar] [CrossRef]
- Lee, C.-W.; Hsiao, W.-T.; Lee, O.K.-S. Mesenchymal Stromal Cell-Based Therapies Reduce Obesity and Metabolic Syndromes Induced by a High-Fat Diet. Transl. Res. 2017, 182, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Karaoz, E.; Okcu, A.; Ünal, Z.S.; Subasi, C.; Saglam, O.; Duruksu, G. Adipose Tissue-Derived Mesenchymal Stromal Cells Efficiently Differentiate into Insulin-Producing Cells in Pancreatic Islet Microenvironment Both in Vitro and in Vivo. Cytotherapy 2013, 15, 557–570. [Google Scholar] [CrossRef] [PubMed]
- Nam, J.S.; Kang, H.M.; Kim, J.; Park, S.; Kim, H.; Ahn, C.W.; Park, J.O.; Kim, K.R. Transplantation of Insulin-Secreting Cells Differentiated from Human Adipose Tissue-Derived Stem Cells into Type 2 Diabetes Mice. Biochem. Biophys Res. Commun. 2014, 443, 775–781. [Google Scholar] [CrossRef] [PubMed]
- Pan, F.; Liao, N.; Zheng, Y.; Wang, Y.; Gao, Y.; Wang, S.; Jiang, Y.; Liu, X. Intrahepatic Transplantation of Adipose-Derived Stem Cells Attenuates the Progression of Non-Alcoholic Fatty Liver Disease in Rats. Mol. Med. Rep. 2015, 12, 3725–3733. [Google Scholar] [CrossRef]
- Shree, N.; Venkategowda, S.; Venkatranganna, M.V.; Datta, I.; Bhonde, R.R. Human Adipose Tissue Mesenchymal Stem Cells as a Novel Treatment Modality for Correcting Obesity Induced Metabolic Dysregulation. Int. J. Obes. 2019, 43, 2107–2118. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; Oh, J.W. Development of CRISPR/Cas9 System for Targeted DNA Modifications and Recent Improvements in Modification Efficiency and Specificity. BMB Rep. 2020, 53, 341–348. [Google Scholar] [CrossRef]
- Kantor, A.; McClements, M.E.; Maclaren, R.E. Crispr-Cas9 Dna Base-Editing and Prime-Editing. Int. J. Mol. Sci. 2020, 21, 6240. [Google Scholar] [CrossRef]
- Lundh, M.; Pluciñska, K.; Isidor, M.S.; Petersen, P.S.S.; Emanuelli, B. Bidirectional Manipulation of Gene Expression in Adipocytes Using CRISPRa and SiRNA. Mol. Metab. 2017, 6, 1313–1320. [Google Scholar] [CrossRef]
- Wang, C.H.; Lundh, M.; Fu, A.; Kriszt, R.; Huang, T.L.; Lynes, M.D.; Leiria, L.O.; Shamsi, F.; Darcy, J.; Greenwood, B.P.; et al. CRISPR-Engineered Human Brown-like Adipocytes Prevent Diet-Induced Obesity and Ameliorate Metabolic Syndrome in Mice. Sci. Transl. Med. 2020, 12, eaaz8664. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Cohen, J.L.; Nicoloro, S.M.; Kelly, M.; Yenilmez, B.; Henriques, F.; Tsagkaraki, E.; Edwards, Y.J.K.; Hu, X.; Friedline, R.H.; et al. CRISPR-Delivery Particles Targeting Nuclear Receptor–Interacting Protein 1 (Nrip1) in Adipose Cells to Enhance Energy Expenditure. J. Biol. Chem. 2018, 293, 17291–17305. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Pros | Cons | |
|---|---|---|---|
| Anthropometric parameters | BMI WC WHR | Non-invasive Cost-effective tests | Do not consider adiposity and AT distribution |
| Imaging techniques | BIA DXA CT MRI | Allow quantification of the fat volume and distribution High accuracy and reproducibility | Complex, require computerized processing Lack of guidelines and thresholds |
| Circulating biomarkers | Adipokines Insulin pathway Inflammatory markers | Non-invasive Cost-effective tests | Levels influenced by multiple factors Lack of guidelines and thresholds |
| Omics-based biomarkers | Genomics Transcriptomics Metabolomics Lipidomics | Integrate several data Allow more personalized diagnosis | Complex, require computerized processing Validation of its reproducibility is needed |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopez-Yus, M.; Hörndler, C.; Borlan, S.; Bernal-Monterde, V.; Arbones-Mainar, J.M. Unraveling Adipose Tissue Dysfunction: Molecular Mechanisms, Novel Biomarkers, and Therapeutic Targets for Liver Fat Deposition. Cells 2024, 13, 380. https://doi.org/10.3390/cells13050380
Lopez-Yus M, Hörndler C, Borlan S, Bernal-Monterde V, Arbones-Mainar JM. Unraveling Adipose Tissue Dysfunction: Molecular Mechanisms, Novel Biomarkers, and Therapeutic Targets for Liver Fat Deposition. Cells. 2024; 13(5):380. https://doi.org/10.3390/cells13050380
Chicago/Turabian StyleLopez-Yus, Marta, Carlos Hörndler, Sofia Borlan, Vanesa Bernal-Monterde, and Jose M. Arbones-Mainar. 2024. "Unraveling Adipose Tissue Dysfunction: Molecular Mechanisms, Novel Biomarkers, and Therapeutic Targets for Liver Fat Deposition" Cells 13, no. 5: 380. https://doi.org/10.3390/cells13050380