
Blood-Derived Lipid and Metabolite Biomarkers in Cardiovascular Research from Clinical Studies: A Recent Update

Leibniz-Institut für Analytische Wissenschaften-ISAS-e.V., 44139 Dortmund, Germany
*
Authors to whom correspondence should be addressed.
Cells 2023, 12(24), 2796; https://doi.org/10.3390/cells12242796
Submission received: 1 September 2023
/
Revised: 24 November 2023
/
Accepted: 1 December 2023
/
Published: 8 December 2023
(This article belongs to the Special Issue The Molecular Application of Mass Spectrometry in Health and Disease)
Abstract
:The primary prevention, early detection, and treatment of cardiovascular disease (CVD) have been long-standing scientific research goals worldwide. In the past decades, traditional blood lipid profiles have been routinely used in clinical practice to estimate the risk of CVDs such as atherosclerotic cardiovascular disease (ASCVD) and as treatment targets for the primary prevention of adverse cardiac events. These blood lipid panel tests often fail to fully predict all CVD risks and thus need to be improved. A comprehensive analysis of molecular species of lipids and metabolites (defined as lipidomics and metabolomics, respectively) can provide molecular insights into the pathophysiology of the disease and could serve as diagnostic and prognostic indicators of disease. Mass spectrometry (MS) and nuclear magnetic resonance (NMR)-based lipidomics and metabolomics analysis have been increasingly used to study the metabolic changes that occur during CVD pathogenesis. In this review, we provide an overview of various MS-based platforms and approaches that are commonly used in lipidomics and metabolomics workflows. This review summarizes the lipids and metabolites in human plasma/serum that have recently (from 2018 to December 2022) been identified as promising CVD biomarkers. In addition, this review describes the potential pathophysiological mechanisms associated with candidate CVD biomarkers. Future studies focused on these potential biomarkers and pathways will provide mechanistic clues of CVD pathogenesis and thus help with the risk assessment, diagnosis, and treatment of CVD.
1. Introduction
Cardiovascular disease (CVD) is a diverse group of heart and vascular disorders that affect a significant proportion of the global population. Despite recent significant advances in various drug- and device-based treatments, CVD is still recognized as one of the leading causes of morbidity and mortality worldwide, posing a substantial socioeconomic burden on individuals and populations [1]. According to statistics before the coronavirus disease 2019 (COVID-19) pandemic, CVD accounted for approximately 18.6 million deaths, representing about one-third of all deaths worldwide [1]. Moreover, the prevalence of CVD has increased by 17.1% from the previous decade, and it is projected to rise globally [1] continuously. The increase in CVD incidence is causally related to the increase in longevity, lifestyle, and environmental-related risk factors [2,3]. Thus, changes in particular lifestyle and environmental-related risk factors, and choices should improve the current situation.
The increase in the prevalence of CVD may be linked to an increase in atherosclerotic cardiovascular disease (ASCVD) cases [4] since ASCVD, with its clinical manifestations including coronary artery disease (CAD), stroke, myocardial infarction (MI), and heart failure (HF), is the major contributing factor to all CVD fatalities [1,5,6,7]. It is a progressive and inflammatory disease characterized by arterial narrowing due to the gradual accumulation of lipids and endothelial dysfunction [8]. Dyslipidemia and hypertension [9,10] are the two main independent risk factors for ASCVD; other risk factors include insulin resistance, hyperglycemia, and obesity [11,12]. Numerous epidemiological and clinical studies [13,14,15] have established a relationship between aberrant blood lipid levels and the prevalence of CVDs such as CAD, MI, and stroke.
For decades, monitoring of the traditional lipid profile from blood tests has been used as a clinical diagnostic test for dyslipidemia, a prominent CVD risk factor which is characterized by elevated levels of low-density lipoprotein cholesterol (LDLC), triacylglycerols (TAG) [16,17], and low levels of high-density lipoprotein cholesterol (HDLC) [18,19]. Blood-derived biofluids, such as serum and plasma, are the preferred patient biospecimens for clinical cardiovascular research. Changes in traditional lipid profiles determine which patients with lipid disorders, and in the primary and secondary prevention of heart attack and stroke, would benefit from statin [20,21] or fibrate [22,23] therapy. Because CVD is a multifactorial disease, crude measurements of traditional lipids (TAG, LDLC, and HDLC) alone frequently fail to identify all individuals at risk for atherosclerotic plaque formation and subsequent thrombus formation [24,25]. This highlights the urgent need for the discovery of clinically applicable biomarkers for the prognosis, diagnosis, treatment, and monitoring of CVD.
Metabolites (including lipids) can be responsive to both genetic background and environmental exposure, thus directly providing the specific molecular phenotypes of the biological systems [26]. Their profiling reflects the pathophysiological processes underlying human diseases [27]. During the past decades, with the advances in analytical technologies, the new era of high-throughput disciplines such as metabolomics (small molecules with a molecular mass ≤ 1500 Da) for profiling of (hydrophilic) metabolites and lipidomics (usually considered a subset of metabolomics) for the profiling of (hydrophobic metabolites) lipids in a biological system have emerged.
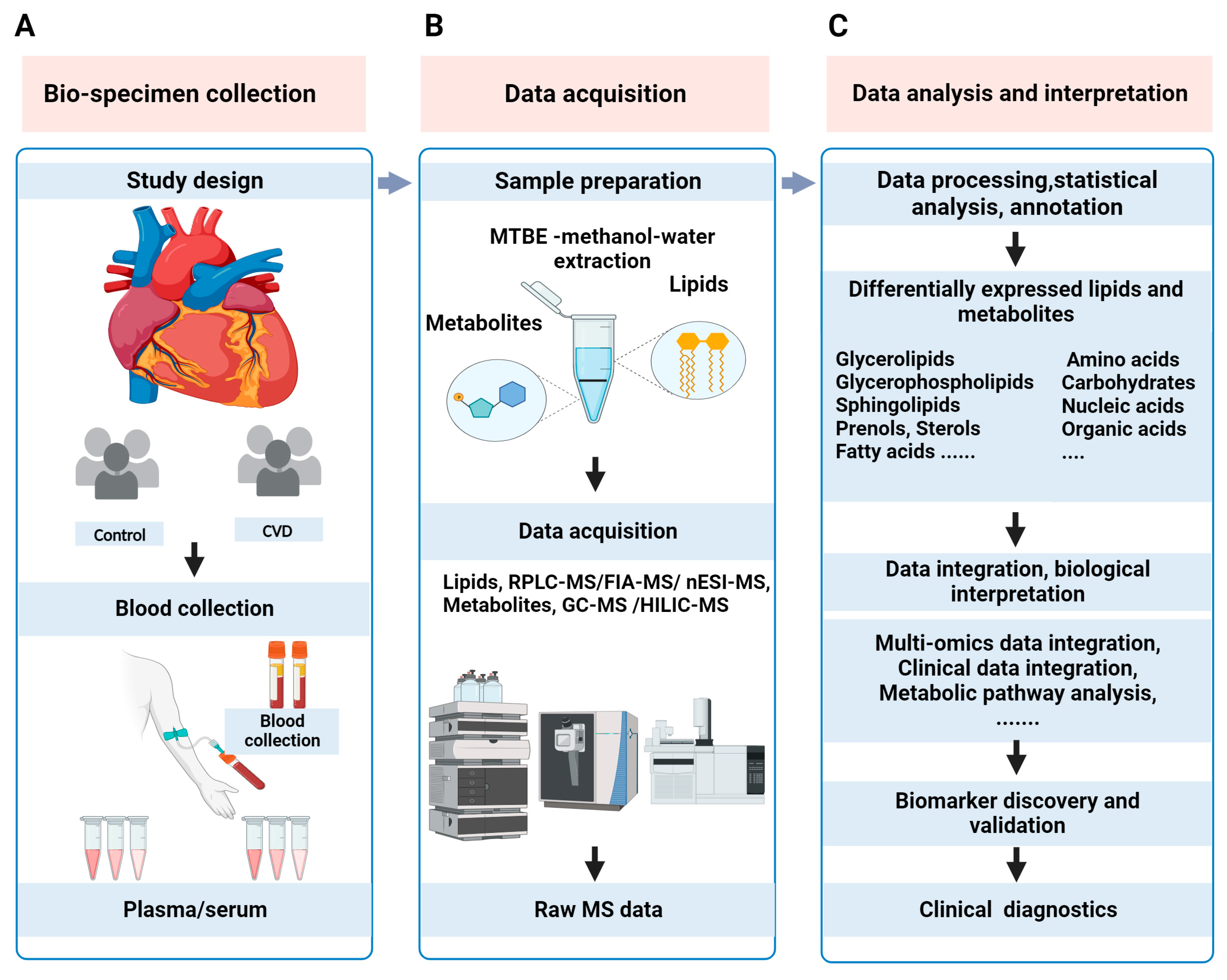
This review provides an overview of MS-based lipidomics and metabolomics (metabolipidomics) workflows, with a focus on commonly used analytical techniques and strategies. Clinical metabolipidomics study typically includes a collection of clinical samples, lipid/metabolite sample preparation, MS data acquisition, statistical analysis, biological interpretation, cross-validation of biomarker candidates, and, finally, clinical translation of biomarkers, as illustrated in Figure 1.
This review summarizes promising CVD biomarkers in human blood plasma/serum samples from recent lipidomics and metabolomics studies. Then, potential CVD pathophysiological mechanisms associated with these biomarkers are described. The association of these potential biomarkers and pathways with CVD should be evaluated in future investigations.
2. Lipidomics and Metabolomics: Analytical Workflows and Techniques
Among the many (>200) risk factors identified, an abnormal lipid panel (dyslipidemia) is the major morbidity associated with CVD. Lipids, which are essential components of cells, play three primary roles: signaling, energy storage, and structural support [29]. Plasma lipidome includes sterol lipids (cholesterol and CE), glycerophospholipids (PC, PC-O, LPC, PE, and PI), glycerolipids (MAG, DAG, and TAG), and sphingolipids (SM, DhCer, Cer, LacCer, HexCer, and FFA) as the most abundant lipids. They are frequently analyzed in blood-derived biofluids, and all cumulatively account for approximately 88% of the plasma lipidome [30] and more than 70% of the entire plasma metabolome [31]. The remaining plasma metabolome consists of hydrophilic metabolites such as carbohydrates, amino acids (involved in protein metabolism), nucleic acids (involved in DNA/RNA metabolism), organic acids, and acylcarnitines.
Lipid and metabolite analysis from complex biological samples such as plasma/serum requires dedicated sample preparation procedures and chromatographic separation modes due to the different physiochemical properties of these small biomolecules. A typical lipidomics workflow comprises sample preparation using the Bligh–Dyer [32], Matyash [33], or Folch [34] lipid extraction methods, whereas a metabolomics workflow involves sample preparation using polar organic solvents such as methanol, acetonitrile, ethanol, and their mixtures. These traditional sample preparation workflows for lipidomics and metabolomics require different aliquots of the same sample. In recent years, novel, simple, and time-saving sample preparation methods to simultaneously extract lipids and metabolites from the same sample, based either on biphasic [28,35] or monophasic [36] solvent systems, have been developed. These sample preparation workflows enable the parallel analysis of both lipids and metabolites from the same sample, thereby increasing the comparability of multiple “omics” datasets and limiting technical variability.
Most metabolipidomics studies rely on powerful analytical techniques such as NMR and MS for metabolite profiling and structural elucidation of unknown metabolites. Although NMR, being a non-destructive, unbiased, and robust technique, offers several advantages, it suffers from low sensitivity and can only detect up to 200 metabolites existing at higher concentrations (≥1 μM) in biological samples [37]. Compared to NMR, MS provides excellent sensitivity and detection limits (from the femto- to the atto-molar range) and a wide dynamic range of coverage (∼103–104). MS separates and identifies ions based on their mass-to-charge (m/z) ratio and, due to its fast scanning speed, it can simultaneously measure hundreds to thousands of analytes with high sensitivity in a single run [37].
In MS-based analyses, matrix-assisted laser desorption/ionization (MALDI) is a “soft” ionization technique that uses a laser energy-absorbing matrix to ionize analytes with minimal fragmentation. MALDI-MS imaging (MALDI-MSI) has been extensively used to visualize the in situ spatial distribution of metabolites from tissue samples [38,39]. Few MALDI-MS-based studies have investigated the serum metabolome profile associated with the clinical phenotype of the diseases [40,41]. The use of MALDI-MS for large-scale biofluid analysis is limited by the coverage of metabolite classes, background noise in the matrix signal, fluctuations in the analyte signal due to matrix “sweet spots”, and poor analyte quantification [40,42].
Electrospray ionization (ESI) is another ionization technique in MS that produces gas-phase ions from analytes in a solution using an electrospray. The ESI-MS interface has been extensively used in clinical research laboratories for routine analysis over several ionization methods such as electron impact (EI), chemical ionization (CI), or MALDI, etc. Biomolecules can be characterized using the (ESI-)MS technique, with prior chromatographic separation, or without prior chromatographic separation, i.e., by direct MS analysis of samples. Direct MS analysis relies on platforms such as native ESI/nano-electrospray ionization (nESI-)MS and flow injection analysis (FIA-)MS to increase samples throughput.
Hyphenated techniques such as liquid chromatography (LC-)MS (with an ESI interface) and gas chromatography (GC-)MS boast several advantages, such as the minimization of ion suppression/enhancement effects, reduction in spectral overlap, and, thus, decreased spectral complexity. Gas chromatography (GC), with an electron ionization (EI) or chemical ionization (CI) interface, is often used for the separation of volatile and thermally stable compounds. GC-MS offers, for low-abundant key metabolites such as BCAA, polyamines, glycolysis, and TCA pathway intermediates (such as pyruvate, fumarate, and citrate) [43,44,45], superior peak resolution and higher sensitivity, in comparison to LC, often employing derivatization to generate more volatile analytes. However, GC-MS is not suitable for separating species with insufficient volatility or thermal instability, such as acylcarnitines (AC) or acyl-coenzyme A (CoA) found in biofluids [46].
LC can separate a wide range of chemically diverse sets of compounds, which makes it the method of choice for routine analysis in clinical research laboratories. Correspondingly, many recent studies implement LC-MS for the comprehensive analysis of polar metabolites and non-polar lipids in CVD research [46]. In general, polar metabolites are separated on hydrophilic interaction liquid chromatography (HILIC), and non-polar lipids are separated on reverse phase (RP-)LC [47,48]. A strategy for dual separation of both polar metabolites and non-polar lipids using a two-dimensional LC(-MS) system combining HILIC separation followed by RPLC separation allows for the separation of both lipids and metabolites from the same extract in a single injection [49,50,51]. In particular, these technological advances support large-scale clinical trials with limited sample volumes and reduce the required device run time.
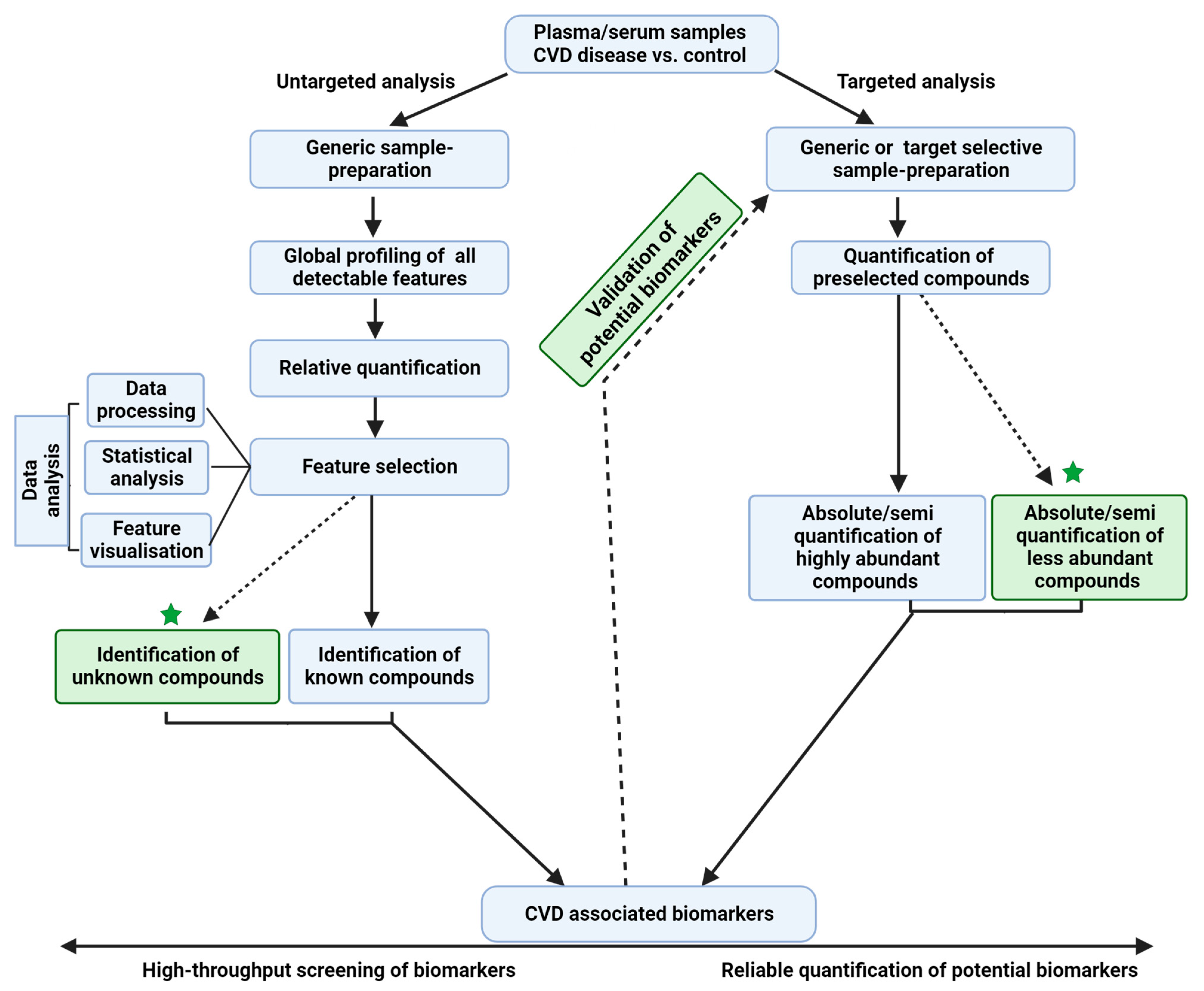
In general, MS-based metabolipidomics analysis can be performed using targeted or untargeted approaches (Figure 2), with each approach having its own advantages and limitations. While the former is a hypothesis-testing approach, the latter is a hypothesis-generating approach involving the discovery and validation of novel candidate biomarkers; however, the process is labor-intensive. The targeted analysis mainly focuses on analyzing a limited set of pre-selected target compounds (tens to hundreds) based on extensive biological hypotheses [52]. It can be performed on both high-resolution (mass resolution ≥30,000 at 200 Da) MS (HRMS) instruments and low-resolution MS (LRMS) instruments. The targeted quantification of compounds by selected/multiple reaction monitoring (SRM/MRM)- or parallel reaction monitoring (PRM)-based data acquisitions enables the accurate and reproducible quantification of analytes.
Untargeted analyses simultaneously measure a broader range of detectable (thousands) features and cover the detection of “unknown metabolites” in the biological sample. For this purpose, HRMS instruments based on mass analyzers such as the Fourier Transform Ion Cyclotron Resonance (FTICR), quadrupole orbitrap (Q-orbitrap), and quadrupole time-of-flight (QqTOF) are utilized. Often in these analyses, MS instruments first perform a full scan MS (MS1), and subsequent tandem MS (MS/MS or MS2) scans are obtained in the data-dependent acquisition (DDA) or data-independent acquisition (DIA) modes. All features obtained are processed and statistically analyzed so that differentially expressed features can be later identified. Feature identification often relies on matching MS1 and MS2 spectra and retention times to spectral libraries or by running reference standards in the same chromatographic run. This approach is often limited by the molecular diversity and complexity of the features detected, complicating subsequent data analysis and interpretation.
Non-targeted assays are commonly used for the high-throughput screening of small molecules in complex biological specimens. Despite their proven utility in the measurement of diverse molecules simultaneously, these assays are known to overlook low-abundant metabolites [53] (e.g., signaling lipids such as phosphatidyl inositol phosphates (PIPs), and eicosanoids and its metabolites). The analysis of such low-abundant metabolites needs targeted analysis, and may sometimes require additional selective sample enrichment/purification steps during the sample preparation procedures prior to MS analysis. In recent years, for the analysis of low-abundant metabolites, several new MS-based analytical methods and techniques (e.g., [54,55]) have been developed. In the future, such analytical methods could be used to comprehensively characterize the homeostasis of these low-abundance analytes under health and disease conditions.
A hybrid approach that integrates both untargeted and targeted approaches will provide comprehensive signatures of the human CVD metabolome (Figure 2). Although MS is a powerful tool for the structural elucidation of complex biological samples, it has a few limitations. For example, the distinction between multiple isomeric, or enantiomeric compounds [56] with subtle structural differences in the same coelution profile is not possible. With emerging technologies in MS such as ion mobility-mass spectrometry (IM-MS), it is feasible to separate isomers rapidly [57]. However, many advancements are to be seen in the mainstream applications of these technologies.
In recent years, several lipidomics and metabolomics studies involving large-scale epidemiological and clinical cohorts based on different analytical and methodological approaches have proposed numerous biomarkers for CVD. There is an urgent need to leverage the knowledge from current metabolipidomics studies to identify promising biomarker candidates and translate them into clinical applications. Accordingly, we aimed to summarize the CVD biomarkers (Table 1) from studies published over the last 5 years in this review. Several keywords such as “biomarker”, “metabolomics”, “lipidomics”, and “cardiovascular disease” were searched in PubMed. Reviews and animal-based research articles were excluded, and only studies based on human plasma/serum using MS analysis were included.
3. Lipid and Metabolite Biomarkers of CVD
MS-based lipidomics and metabolomics studies have shown the potential to screen biomarkers for human diseases [83,84,85]. To date, ceramides or ceramide panel combinations are the only biomarkers that are translated from metabolipidomics research into clinically approved laboratory tests. These bioactive lipids are central components of sphingolipid cell signaling pathways. They play myriad roles in cellular processes such as the regulation of apoptosis [86], cellular stress, and vascular inflammation [87,88,89,90]. Cardiometabolic diseases such as CAD [91,92] ischemic stroke [93], HF [94], lipotoxic cardiomyopathy [95], and MI [96] are associated with elevated circulating plasma ceramide levels. Higher ceramide levels are also implicated in major comorbidities of CVD, namely, type 2 diabetes mellitus (T2D) [97,98] and insulin resistance [99].
Ceramides are mostly generated in the liver, and transported into the bloodstream by lipoproteins. Two primary pathways produce ceramide (or dihydroceramide, in the event of de novo synthesis): a de novo synthesis (beginning with fatty acids) and re-acylation of sphingosine (salvage pathway), both regulated by ceramide synthases (CerS). Mammals have six isoforms of CerS (CerS1-6), each of which can preferably synthesize ceramides with a specific fatty acyl chain length (from 14 to 26 carbon atoms) [91,92], and expression patterns of CerS vary across different tissue types. Numerous studies have implicated that each CerS species has specific pathophysiological roles [100,101,102,103].
Several studies have been carried out to verify the functions of specific isoforms of CerS, and CerS6 expression in particular has been correlated with insulin resistance. Turpin et al. [101] have shown that mice lacking CerS6 were protected from glucose intolerance and high fat, diet-induced obesity. Increasing evidence suggests that CerSs are potential therapeutic targets to treat CVD, by modulating specific ceramide levels in various pathologies.
The correlation between alterations in biomarkers in the biofluids and the cellular/tissue environment of patients with CVD may provide significant insights into the role of metabolic dysfunction in the progression of CVD pathophysiology. This correlation between biofluids ceramide metabolite markers and cellular/tissue alterations offers promising avenues for identifying potential diagnostic biomarkers, therapeutic targets, and personalized treatment strategies, thereby advancing precision medicine approaches in managing and combating cardiovascular diseases.
Given the critical role of ceramides in the development of CVD, several analytical methods have been developed and validated to quantify ceramides in human plasma and serum [104,105,106] samples.
Various research studies mentioned later have shown that clinical plasma/serum measurements of ceramides provide an independent added value to the routinely used diagnostic and prognostic tools for CVD events. The first clinical study to introduce the association of plasma ceramide levels with CVD and total mortality in CAD disease is the LURIC (Ludwigshafen Risk and cardiovascular health) [107]. Thereafter, three independent CAD cohorts, namely the Special Program University Medicine Acute Coronary Syndromes and Inflammation (SPUM-ACS) and the Bergen Coronary Angiography Cohort (BECAC) studies [108], have reconfirmed and validated the use of ceramides as predictors of CV mortality. In other studies, high levels of plasma/serum C16:0, C18:0, and C24:1 ceramides, and ratios of these ceramides to C24:0 ceramide, were shown to predict adverse cardiac events in healthy individuals and patients [108,109,110]. Further, ZoraBiosciences developed the Cardiovascular Event Risk Test (CERT)-1 risk score, comprising previously stated plasma/serum ceramide or ceramide ratios, which was used to stratify individuals into four (low, medium, increased, and high) risk categories for the primary and secondary prevention of CVD. Following this, the first diagnostic test to assess the risk of cardiovascular events in CAD patients was launched by the Mayo Clinic in the USA. In the Western Norway Coronary Angiography Cohort (WECAC) study, the CERT-1 risk score prediction was improved by including distinct phosphatidylcholines (PCs), termed the CERT-2 risk score. The CERT-2 score was verified in the LIPID and KAROLA studies [58]. Further, the sphingolipid-inclusive CAD risk score, termed the SIC score [59], for demarcating CAD patients compared to controls, and the diabetes score (dscore) [111] for predicting the onset of diabetes, have been developed.
TMAO (trimethylamine N-oxide), a circulating gut microbiota-dependent metabolite mainly derived from the oxidation of trimethylamine (TMA) and its dietary precursor [112] choline (derived from dietary PC or the hydrolysis of endogenous PC), has recently emerged as a potential biomarker for CVD. Since its discovery [112], TMAO has been implicated as a predictive biomarker for thrombosis and platelet hyperactivation [113], increased CVD risk, and atherosclerotic plaque formation [114,115,116,117] in many clinical studies. Elevated levels of TMAO in plasma are independent biomarkers of CVD for secondary prevention. The Cleveland Clinic Heart Lab commenced blood tests to measure blood TMAO levels using LC-MS/MS [118]. Although there is a broad consensus that elevated plasma TMAO levels are independent CVD biomarkers for secondary prevention [119], few recent studies [120,121] provide conflicting evidence for an association between circulatory TMAO metabolism and CVD events, suggesting that TMAO is not an independent risk factor for evaluating CV risk events. Thus, currently, traditional lipid panels, ceramide panels, and TMAO are the only established screening tests to assess future risks of CVD. Elevated levels of acylcarnitines (the transport form of activated FAs) and BCAAs are associated with an increased risk of CVD events [46,122,123,124]. This dysregulation causes a mitochondrial imbalance between mitochondrial fatty acid β-oxidation (MFAO) and glucose oxidation. Fatty acids serve as an energy source in MFAO, and acylcarnitines, which act as long-chain fatty acid carriers into the mitochondria, play important roles in mitochondrial energy homeostasis through MFAO [125,126]. MFAO is affected by BCAA catabolism in which the excess catabolic flux of BCAAs causes the accumulation of trans-endothelial fatty acid into the muscle, the accumulation of lipotoxic, incompletely esterified intermediates such as DAG, and blunted insulin signaling [127,128], thus effectively clogging the β-oxidative machinery in a manner analogous to the effect of excess fat [129]. Furthermore, both fatty acid oxidation and BCAA catabolism are closely interconnected through energy metabolism, as both can affect glycolysis through inhibiting pyruvate dehydrogenase and the TCA cycle through acetyl-CoA/succinyl-CoA/substrate (“cycle”) overload, as well as mitochondrial oxidative phosphorylation through their common target, PGC-1α [126]. These overall changes in various lipids, fatty acids, and BCAAs in circulation emphasize the disturbances in metabolic homeostasis and energy metabolism in CVD [126,130,131]. Furthermore, clogging of β-oxidation machinery may explain the decreased glucose utilization, increased glucose tolerance, and the development of insulin resistance in muscle cells, relevant for T2D [129] pathophysiology, which is a comorbidity often associated with CVD. This also highlights the role of the accumulation of BCAA in failing hearts, which was associated with myocardial insulin resistance [44,46]. Altogether, abnormal levels of circulating FAs, acylcarnitines, and BCAAs modulate insulin sensitivity, energy metabolism, and inflammation, and thus contribute to CVD risk factors such as T2D and obesity.
Tryptophan degradation via the kynurenine pathway is an important pathway in energy metabolism, leading to the biosynthesis of nicotinamide adenine dinucleotide (NAD+) [132]. Kynurenine pathway intermediates and enzymes have been proposed as inflammatory markers [133,134] in CVD [135,136]. The kynurenine pathway and CVD risk factors such as hypertension, diabetes mellitus, dyslipidemia, and obesity seem to be at play here [133,137]. Accordingly, several studies already note kynurenine pathway metabolites as diagnostic or predictive markers of ASCVD or CAD [reviewed in [137].
Along with the above pathways, inflammation, reactive oxidation species, and nitric oxide also play a role in CVD [138], however, it is difficult to capture these pathways’ alteration as the pathway intermediates are not easily detected in metabolomics analysis. In many reported studies, the mechanistic understanding of the role of newly screened biomarkers in the biological pathways underlying CVD has not yet been established or validated, and thus, they have limited clinical utility in CVD. After the discovery phase, these biomarkers should be validated and fulfill the proposed criteria for “biomarker qualification” before translating into clinical practice [139]. The addition of these biomarkers to the base model of CVD, along with other established biomarkers, could improve the mechanistic understanding of CVD as well as CVD-associated risk modeling.
4. Clinical Translation of Metabolite Biomarkers
Metabolites play diverse roles as cellular end products or byproducts, signaling molecules, enzyme modulators, and reactive species. Due to the advanced nature of these molecular indicators, mechanistic evaluation of their intended purpose with respect to clinical utility is complex, making clinical translation of metabolomic biomarkers a multifaceted process.
Pre-analytical biases in clinical “omics” can significantly affect the overall reliability of study results and the suitability of specific lipids/metabolites as biomarkers. They arise primarily from differences in sample labeling, handling, collection, processing, or storage. For example, differences in sample treatment before the analysis of blood samples affected the quantification of blood-based metabolites [28,29,30]. Reliable quantification of lipids and metabolites in clinical studies can only be achieved by standardizing pre-analytical variables. Recent reviews [31,32,33,34] have proposed several recommendations for the standardization of preanalytical sample handling procedures to obtain reliable and reproducible results. The processing and analysis of obtained MS data and its biological interpretation, including integration with other omics/clinical data and pathway analysis and the clinical translation of biomarkers, have been extensively discussed in other reviews [35,36,37] and are not the focus of this review.
For assessing metabolomic biomarkers to validate their clinical utility, performance evaluations through comparative studies (including longitudinal and population-based studies) using established standards and biobank samples is necessary. Such investigations require improved accuracy, precision, and critical data analysis. Analytical quality criteria include precision, precision, sensitivity, specificity, wide dynamic range, and robustness in biomarker assays, as well as traceability to reference standards and robustness to sample variation. In clinical biomarker discovery research, validated analytical methods must be rigorously implemented to ensure that they are suitable for the intended purpose. Thus, in our opinion, the pertaining FDA ICH M10 guidelines should also be followed for biomarker discovery research. Additionally, regulatory eligibility for qualification for biomarker research will expedite the international harmonization of the biomarker search process.
In high-throughput untargeted metabolomics, metabolites are often putatively annotated by database searches based on the spectral matching of high-resolution MS1 and MS2 data. The putative annotations of metabolites can lead to the misidentification of biomarkers and, thus, to wrong biological interpretation [56]. The Metabolomics Standards Initiative (MSI) has proposed four confidence levels for annotating and identifying metabolites [140], with level one being the highest confidence level. However, only a few untargeted metabolomics studies have reported the identification (as proposed by MSI) of metabolites at level one [141]. Thus, the correct annotation of candidate biomarkers remains a major bottleneck in large-scale clinical metabolomics studies based on untargeted approaches.
The critical evaluation, integration, and analysis of MS-based metabolomics data is crucial for the successful clinical translation of metabolite biomarkers. The prognosis and diagnosis of cardiovascular disease (CVD) may now be predicted using machine learning, which presents promising possibilities for individualized treatment plans. These computational advances should be well utilized to facilitate the broad applicability of metabolomics datasets and multi-omics data integration.
The number of metabolomics studies has increased over the past decade, necessitating the transparent reporting of the results of these studies. This will help to increase comparability across different studies and could facilitate the harmonization of biomarkers (reviewed in [142,143,144,145,146,147]). Furthermore, efforts have to be made by research communities to make minimum reporting checklists mandatory components for both lipidomics [148,149] and metabolomics studies [150].
In conclusion, the clinical translation of metabolomic biomarkers in life science research demands a comprehensive understanding of their clinical utility and pathophysiological mechanisms. Investigating their performance metrics, evaluating their complementary value in refining diagnoses and prognoses, and assessing their direct influence on decision-making processes are pivotal steps toward unlocking the full potential of metabolites. Addressing these crucial questions is paramount for propelling the field towards an integrated application of metabolomic insights within clinical practice, thereby shaping the future landscape of diagnostics and personalized medicine.
5. Conclusions
Metabolipidomics studies on blood-based biofluids have demonstrated the potential to pioneer the discovery of ceramides as clinical markers for the diagnosis and prognosis of CVD. Novel sample preparation strategies, automated data analysis workflows, machine learning algorithms, and faster MS instrumentation technologies have guided the rapid, robust, and large-scale quantification of lipids and metabolites. Comprehensive metabolomics and lipidomics analysis, in combination with proteomics and genomic analysis, can provide a complete “snapshot” of the pathophysiological mechanisms underlying CVD. This would consequently allow us to accurately identify clinically translated biomarkers and therapeutic targets of CVD. In this review (Table 1), we have noted the association of intricately interconnected biochemical processes such as fatty acid and carbohydrate, BCAA and gut microbial metabolism, glycerophospholipid, and sphingolipid pathways is reflected in many studies [58,59,60,63,65,68,70,76,78,79] where multiple metabolite classes are altered simultaneously in concerted fashion. The comprehensive monitoring of alterations in these pathway metabolites in future clinical studies as a whole (panel) will establish the causal or correlational association between CVD and metabolite biomarkers.
Author Contributions
D.K. and A.S.: Conceptualization, visualization; D.K.: Data curation, Figures, and tables preparation; A.F. and D.K.: methodology; literature investigation; A.F., P.P. and D.K.: writing—original draft preparation; writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
All authors acknowledge the support of the “Ministerium für Kultur und Wissenschaft des Landes Nordrhein-Westfalen” and “Berliner Senatsverwaltung für Wissenschaft, Gesundheit und Pflege”. This work was partly funded by the “Federal Ministry of Education and Research (Bundesministerium für Bildung und Forschung, BMBF) under the funding reference 161L0271”.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| AA | Amino Acid |
| AC | Acylcarnitines |
| ACS | Acute Coronary Syndromes |
| AF | Atrial Fibrillation |
| ASCVD | Atherosclerotic Cardiovascular Disease |
| AI | Atherosclerotic index |
| BCAA | Branched-Chain Amino Acids |
| BECAC | Bergen Coronary Angiography Cohort |
| CAD | Coronary Artery Disease |
| CE | Cholesteryl Ester |
| Cer | Ceramide |
| CerS | Ceramide Synthases |
| CERT | Cardiovascular Event Risk Test |
| CoA | Acyl-Coenzyme A |
| CVD | Cardiovascular Disease |
| DAG | Diacylglycerols |
| DCM | Diabetic Cardiomyopathy |
| DhCer | Dihydroceramide |
| DhSM | Dihydrosphingomylein |
| ESI | Electrospray Ionization |
| FA | Fatty Acids |
| FFA | Free Fatty Acids |
| FTICR | Fourier Transform Ion Cyclotron Resonance |
| GC-MS | Gas Chromatography-Mass Spectrometry |
| HDLC | High-Density Lipoprotein Cholesterol |
| HexCer | Hexosylceramide |
| HF | Heart Failure |
| HILIC | Hydrophilic Interaction Liquid Chromatography |
| HRMS | High-Resolution Mass Spectrometer |
| IS | Ischemic Stroke |
| KAROLA | Langzeiterfolge Der Kardiologischen Anschlussheilbehandlung |
| LacCer | Lactosylceramides |
| LC-MS | Liquid Chromatography-Mass Spectrometry |
| LDLC | Low-Density Lipoprotein Cholesterol |
| LIPID | Long-Term Intervention with Pravastatin in Ischemic Disease |
| LPC | Lysophosphatidylcholines |
| LPE | Lysophosphatidylethanolamine |
| LRMS | Low-Resolution Mass Spectrometer |
| LURIC | Ludwigshafen Risk And Cardiovascular Health |
| MAG | Monoacylglycerols |
| MFAO | Mitochondrial Fatty Acid Β-Oxidation |
| MALDI | Matrix-Assisted Laser Desorption/Ionization |
| MI | Myocardial Infarction |
| NCA | Normal Coronary Artery |
| PC | Phosphatidylcholines |
| PC-O | Alkyl Phosphatidylcholine |
| PE | Phosphatidylethanolamine |
| PI | Phosphatidylinositol |
| PIVUS | Prospective Investigation of the Vasculature in Uppsala Seniors |
| Q-orbitrap | Quadrupole Orbitrap |
| QqTOF | Quadrupole Time Of Flight |
| RP | Reverse Phase |
| SIC | Sphingolipid-Inclusive Cad Risk Score |
| SM | Sphingomyelins |
| SPL | Sphingolipids |
| SPUM-ACS | Special Program University Program-Acute Coronary Syndromes |
| ST2DM | Simple Type 2 Diabetes Mellitus |
| T2D | Type 2 Diabetes Mellitus |
| TAG | Triacylglycerol |
| TCA | Tricarboxylic Acid Cycle |
| TMA | Trimethylamine |
| TMAO | Trimethylamine N-Oxide |
| ULSAM | Uppsala Longitudinal Study Of Adult Men |
| WECAC | The Western Norway Coronary Angiography Cohort |
References
- Roth, G.A.; Mensah, G.A.; Johnson, C.O.; Addolorato, G.; Ammirati, E.; Baddour, L.M.; Barengo, N.C.; Beaton, A.Z.; Benjamin, E.J.; Benziger, C.P.; et al. Global Burden of Cardiovascular Diseases and Risk Factors, 1990-2019: Update from the GBD 2019 Study. J. Am. Coll. Cardiol. 2020, 76, 2982–3021. [Google Scholar] [CrossRef]
- Al-Kindi, S.G.; Brook, R.D.; Biswal, S.; Rajagopalan, S. Environmental determinants of cardiovascular disease: Lessons learned from air pollution. Nat. Rev. Cardiol. 2020, 17, 656–672. [Google Scholar] [CrossRef] [PubMed]
- Jousilahti, P.; Vartiainen, E.; Tuomilehto, J.; Puska, P. Sex, age, cardiovascular risk factors, and coronary heart disease: A prospective follow-up study of 14 786 middle-aged men and women in Finland. Circulation 1999, 99, 1165–1172. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, S.; Han, S.; Rane, P.P.; Fox, K.M.; Qian, Y.; Suh, H.S. Prevalence and incidence of atherosclerotic cardiovascular disease and its risk factors in Korea: A nationwide population-based study. BMC Public Health 2019, 19, 1112. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Benjamin, E.J.; Go, A.S.; Arnett, D.K.; Blaha, M.J.; Cushman, M.; Turner, M.B. Heart disease and stroke statistics—2015 update: A report from the American Heart Association. Circulation 2015, 134, e38–e360. [Google Scholar] [CrossRef] [PubMed]
- Libby, P.; Buring, J.E.; Badimon, L.; Hansson, G.K.; Deanfield, J.; Bittencourt, M.S.; Tokgözoğlu, L.; Lewis, E.F. Atherosclerosis. Nat. Rev. Dis. Primers 2019, 5, 56. [Google Scholar] [CrossRef] [PubMed]
- Mackinnon, E.S.; Goeree, R.; Goodman, S.G.; Rogoza, R.M.; Packalen, M.; Pericleous, L.; Motsepe-Ditshego, P.; Oh, P. Increasing Prevalence and Incidence of Atherosclerotic Cardiovascular Disease in Adult Patients in Ontario, Canada from 2002 to 2018. CJC Open 2022, 4, 206–213. [Google Scholar] [CrossRef]
- Lusis, A.J. Atherosclerosis. Nature 2000, 407, 233–241. [Google Scholar] [CrossRef]
- Fuchs, F.D.; Whelton, P.K. High Blood Pressure and Cardiovascular Disease. Hypertension 2020, 75, 285–292. [Google Scholar] [CrossRef]
- Hedayatnia, M.; Asadi, Z.; Zare-Feyzabadi, R.; Yaghooti-Khorasani, M.; Ghazizadeh, H.; Ghaffarian-Zirak, R.; Nosrati-Tirkani, A.; Mohammadi-Bajgiran, M.; Rohban, M.; Sadabadi, F.; et al. Dyslipidemia and cardiovascular disease risk among the MASHAD study population. Lipids Health Dis. 2020, 19, 42. [Google Scholar] [CrossRef]
- Bornfeldt, K.E.; Tabas, I. Insulin resistance, hyperglycemia, and atherosclerosis. Cell Metab. 2011, 14, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Di Pino, A.; DeFronzo, R.A. Insulin Resistance and Atherosclerosis: Implications for Insulin-Sensitizing Agents. Endocr. Rev. 2019, 40, 1447–1467. [Google Scholar] [CrossRef] [PubMed]
- Lozano, J.V.; Pallarés, V.; Cea-Calvo, L.; Llisterri, J.L.; Fernández-Pérez, C.; Martí-Canales, J.C.; Aznar, J.; Gil-Guillén, V.; Redón, J.; Investigators of the Prev-Ictus Study. Serum lipid profiles and their relationship to cardiovascular disease in the elderly: The PREV-ICTUS study. Curr. Med. Res. Opin. 2008, 24, 659–670. [Google Scholar] [CrossRef] [PubMed]
- Sherpa, L.Y.; Deji; Stigum, H.; Chongsuvivatwong, V.; Luobu, O.; Thelle, D.S.; Nafstad, P.; Bjertness, E. Lipid profile and its association with risk factors for coronary heart disease in the highlanders of Lhasa, Tibet. High Alt. Med. Biol. 2011, 12, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Castelli, W.P. Epidemiology of coronary heart disease: The Framingham study. Am. J. Med. 1984, 76, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, J.L.; Brown, M.S. A century of cholesterol and coronaries: From plaques to genes to statins. Cell 2015, 161, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Ference, B.A.; Yoo, W.; Alesh, I.; Mahajan, N.; Mirowska, K.K.; Mewada, A.; Kahn, J.; Afonso, L.; Williams, K.A., Sr.; Flack, J.M. Effect of long-term exposure to lower low-density lipoprotein cholesterol beginning early in life on the risk of coronary heart disease: A Mendelian randomization analysis. J. Am. Coll. Cardiol. 2012, 60, 2631–2639. [Google Scholar] [CrossRef] [PubMed]
- Geyer, P.E.; Holdt, L.M.; Teupser, D.; Mann, M. Revisiting biomarker discovery by plasma proteomics. Mol. Syst. Biol. 2017, 13, 942. [Google Scholar] [CrossRef]
- Muschitz, C.; Kautzky-Willer, A.; Rauner, M.; Winhöfer-Stöckl, Y.; Haschka, J. Diagnosis and management of patients with diabetes and co-existing osteoporosis (Update 2019): Common guideline of the Autrian Society for Bone and Mineral Research and the Austrian Diabetes Society. Wien. Klin. Wochenschr. 2019, 131, 174–185. [Google Scholar] [CrossRef]
- Vrecer, M.; Turk, S.; Drinovec, J.; Mrhar, A. Use of statins in primary and secondary prevention of coronary heart disease and ischemic stroke. Meta-analysis of randomized trials. Int. J. Clin. Pharmacol. Ther. 2003, 41, 567–577. [Google Scholar] [CrossRef]
- Virani, S.S.; Smith, S.C., Jr.; Stone, N.J.; Grundy, S.M. Secondary Prevention for Atherosclerotic Cardiovascular Disease: Comparing Recent US and European Guidelines on Dyslipidemia. Circulation 2020, 141, 1121–1123. [Google Scholar] [CrossRef] [PubMed]
- Jakob, T.; Nordmann, A.J.; Schandelmaier, S.; Ferreira-González, I.; Briel, M. Fibrates for primary prevention of cardiovascular disease events. Cochrane Database Syst. Rev. 2016, 11, CD009753. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.H.; Kim, S.G. Fibrates Revisited: Potential Role in Cardiovascular Risk Reduction. Diabetes Metab. J. 2020, 44, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Sachdeva, A.; Cannon, C.P.; Deedwania, P.C.; Labresh, K.A.; Smith, S.C., Jr.; Dai, D.; Hernandez, A.; Fonarow, G.C. Lipid levels in patients hospitalized with coronary artery disease: An analysis of 136,905 hospitalizations in Get with The Guidelines. Am. Heart J. 2009, 157, 111–117.e112. [Google Scholar] [CrossRef] [PubMed]
- Wong, Y.-K.; Tse, H.-F. Circulating Biomarkers for Cardiovascular Disease Risk Prediction in Patients with Cardiovascular Disease. Front. Cardiovasc. Med. 2021, 8, 713191. [Google Scholar] [CrossRef] [PubMed]
- Rattray, N.J.W.; Deziel, N.C.; Wallach, J.D.; Khan, S.A.; Vasiliou, V.; Ioannidis, J.P.A.; Johnson, C.H. Beyond genomics: Understanding exposotypes through metabolomics. Hum. Genom. 2018, 12, 4. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S. Metabolomics for Investigating Physiological and Pathophysiological Processes. Physiol. Rev. 2019, 99, 1819–1875. [Google Scholar] [CrossRef] [PubMed]
- Coman, C.; Solari, F.A.; Hentschel, A.; Sickmann, A.; Zahedi, R.P.; Ahrends, R. Simultaneous Metabolite, Protein, Lipid Extraction (SIMPLEX): A Combinatorial Multimolecular Omics Approach for Systems Biology. Mol. Cell. Proteom. 2016, 15, 1453–1466. [Google Scholar] [CrossRef]
- Jaishy, B.; Abel, E.D. Lipids, lysosomes, and autophagy. J. Lipid Res. 2016, 57, 1619–1635. [Google Scholar] [CrossRef]
- Quehenberger, O.; Dennis, E.A. The human plasma lipidome. N. Engl. J. Med. 2011, 365, 1812–1823. [Google Scholar] [CrossRef]
- Gallart-Ayala, H.; Teav, T.; Ivanisevic, J. Metabolomics meets lipidomics: Assessing the small molecule component of metabolism. Bioessays 2020, 42, e2000052. [Google Scholar] [CrossRef] [PubMed]
- Sündermann, A.; Eggers, L.F.; Schwudke, D. Liquid Extraction: Bligh and Dyer. In Encyclopedia of Lipidomics; Wenk, M.R., Ed.; Springer: Dordrecht, The Netherlands, 2016; pp. 1–4. [Google Scholar] [CrossRef]
- Matyash, V.; Liebisch, G.; Kurzchalia, T.V.; Shevchenko, A.; Schwudke, D. Lipid extraction by methyl-tert-butyl ether for high-throughput lipidomics. J. Lipid Res. 2008, 49, 1137–1146. [Google Scholar] [CrossRef] [PubMed]
- Ulmer, C.Z.; Jones, C.M.; Yost, R.A.; Garrett, T.J.; Bowden, J.A. Optimization of Folch, Bligh-Dyer, and Matyash sample-to-extraction solvent ratios for human plasma-based lipidomics studies. Anal. Chim. Acta 2018, 1037, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Hoene, M.; Li, J.; Li, Y.; Zhao, X.; Häring, H.-U.; Schleicher, E.D.; Weigert, C.; Xu, G.; Lehmann, R. Simultaneous extraction of metabolome and lipidome with methyl tert-butyl ether from a single small tissue sample for ultra-high performance liquid chromatography/mass spectrometry. J. Chromatogr. A 2013, 1298, 9–16. [Google Scholar] [CrossRef]
- Muehlbauer, L.K.; Jen, A.; Zhu, Y.; He, Y.; Shishkova, E.; Overmyer, K.A.; Coon, J.J. Rapid Multi-Omics Sample Preparation for Mass Spectrometry. Anal. Chem. 2023, 95, 659–667. [Google Scholar] [CrossRef]
- Marshall, D.D.; Powers, R. Beyond the paradigm: Combining mass spectrometry and nuclear magnetic resonance for metabolomics. Prog. Nucl. Magn. Reson. Spectrosc. 2017, 100, 1–16. [Google Scholar] [CrossRef]
- Andersen, M.K.; Høiem, T.S.; Claes, B.S.R.; Balluff, B.; Martin-Lorenzo, M.; Richardsen, E.; Krossa, S.; Bertilsson, H.; Heeren, R.M.A.; Rye, M.B.; et al. Spatial differentiation of metabolism in prostate cancer tissue by MALDI-TOF MSI. Cancer Metab. 2021, 9, 9. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, C.; Tian, L.; Wu, L.; Jie, Y.; Wang, N.; Liu, R.; Wang, L. In situ metabolic profile and spatial distribution of ocular tissues: New insights into dry eye disease. Ocul. Surf. 2022, 24, 51–63. [Google Scholar] [CrossRef]
- Lai, X.; Guo, K.; Huang, W.; Su, Y.; Chen, S.; Li, Q.; Liang, K.; Gao, W.; Wang, X.; Chen, Y.; et al. Combining MALDI-MS with machine learning for metabolomic characterization of lung cancer patient sera. Anal. Methods 2022, 14, 499–507. [Google Scholar] [CrossRef]
- Wolrab, D.; Jirásko, R.; Cífková, E.; Höring, M.; Mei, D.; Chocholoušková, M.; Peterka, O.; Idkowiak, J.; Hrnčiarová, T.; Kuchař, L.; et al. Lipidomic profiling of human serum enables detection of pancreatic cancer. Nat. Commun. 2022, 13, 124. [Google Scholar] [CrossRef]
- Wang, P.; Giese, R.W. Recommendations for quantitative analysis of small molecules by matrix-assisted laser desorption ionization mass spectrometry. J. Chromatogr. A 2017, 1486, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Spagou, K.; Theodoridis, G.; Wilson, I.; Raikos, N.; Greaves, P.; Edwards, R.; Nolan, B.; Klapa, M.I. A GC-MS metabolic profiling study of plasma samples from mice on low- and high-fat diets. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2011, 879, 1467–1475. [Google Scholar] [CrossRef] [PubMed]
- Sansbury, B.E.; DeMartino, A.M.; Xie, Z.; Brooks, A.C.; Brainard, R.E.; Watson, L.J.; DeFilippis, A.P.; Cummins, T.D.; Harbeson, M.A.; Brittian, K.R.; et al. Metabolomic analysis of pressure-overloaded and infarcted mouse hearts. Circ. Heart Fail. 2014, 7, 634–642. [Google Scholar] [CrossRef] [PubMed]
- Fiehn, O. Metabolomics by Gas Chromatography-Mass Spectrometry: Combined Targeted and Untargeted Profiling. Curr. Protoc. Mol. Biol. 2016, 114, 30.34.31–30.34.32. [Google Scholar] [CrossRef] [PubMed]
- McGarrah, R.W.; Crown, S.B.; Zhang, G.-F.; Shah, S.H.; Newgard, C.B. Cardiovascular Metabolomics. Circ. Res. 2018, 122, 1238–1258. [Google Scholar] [CrossRef] [PubMed]
- Dettmer, K.; Aronov, P.A.; Hammock, B.D. Mass spectrometry-based metabolomics. Mass Spectrom. Rev. 2007, 26, 51–78. [Google Scholar] [CrossRef] [PubMed]
- Lam, S.M.; Wang, Y.; Li, B.; Du, J.; Shui, G. Metabolomics through the lens of precision cardiovascular medicine. J. Genet. Genom. 2017, 44, 127–138. [Google Scholar] [CrossRef]
- Wang, S.; Zhou, L.; Wang, Z.; Shi, X.; Xu, G. Simultaneous metabolomics and lipidomics analysis based on novel heart-cutting two-dimensional liquid chromatography-mass spectrometry. Anal. Chim. Acta 2017, 966, 34–40. [Google Scholar] [CrossRef]
- Feng, J.; Zhong, Q.; Kuang, J.; Liu, J.; Huang, T.; Zhou, T. Simultaneous Analysis of the Metabolome and Lipidome Using Polarity Partition Two-Dimensional Liquid Chromatography–Mass Spectrometry. Anal. Chem. 2021, 93, 15192–15199. [Google Scholar] [CrossRef]
- Schwaiger, M.; Schoeny, H.; El Abiead, Y.; Hermann, G.; Rampler, E.; Koellensperger, G. Merging metabolomics and lipidomics into one analytical run. Analyst 2018, 144, 220–229. [Google Scholar] [CrossRef]
- Lu, W.; Bennett, B.D.; Rabinowitz, J.D. Analytical strategies for LC-MS-based targeted metabolomics. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2008, 871, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Basit, A.; Pontis, S.; Piomelli, D.; Armirotti, A. Ion mobility mass spectrometry enhances low-abundance species detection in untargeted lipidomics. Metabolomics 2016, 12, 50. [Google Scholar] [CrossRef] [PubMed]
- Kale, D.; Kikul, F.; Phapale, P.; Beedgen, L.; Thiel, C.; Brügger, B. Quantification of Dolichyl Phosphates Using Phosphate Methylation and Reverse-Phase Liquid Chromatography–High Resolution Mass Spectrometry. Anal. Chem. 2023, 95, 3210–3217. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Lämmerhofer, M. Isomer Selective Comprehensive Lipidomics Analysis of Phosphoinositides in Biological Samples by Liquid Chromatography with Data Independent Acquisition Tandem Mass Spectrometry. Anal. Chem. 2021, 93, 9583–9592. [Google Scholar] [CrossRef] [PubMed]
- Theodoridis, G.; Gika, H.; Raftery, D.; Goodacre, R.; Plumb, R.S.; Wilson, I.D. Ensuring Fact-Based Metabolite Identification in Liquid Chromatography–Mass Spectrometry-Based Metabolomics. Anal. Chem. 2023, 95, 3909–3916. [Google Scholar] [CrossRef] [PubMed]
- Olajide, O.E.; Donkor, B.; Hamid, A.M. Systematic Optimization of Ambient Ionization Ion Mobility Mass Spectrometry for Rapid Separation of Isomers. J. Am. Soc. Mass Spectrom. 2022, 33, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Hilvo, M.; Meikle, P.J.; Pedersen, E.R.; Tell, G.S.; Dhar, I.; Brenner, H.; Schöttker, B.; Lääperi, M.; Kauhanen, D.; Koistinen, K.M.; et al. Development and validation of a ceramide- and phospholipid-based cardiovascular risk estimation score for coronary artery disease patients. Eur. Heart J. 2020, 41, 371–380. [Google Scholar] [CrossRef]
- Poss, A.M.; Maschek, J.A.; Cox, J.E.; Hauner, B.J.; Hopkins, P.N.; Hunt, S.C.; Holland, W.L.; Summers, S.A.; Playdon, M.C. Machine learning reveals serum sphingolipids as cholesterol-independent biomarkers of coronary artery disease. J. Clin. Investig. 2020, 130, 1363–1376. [Google Scholar] [CrossRef]
- Surendran, A.; Atefi, N.; Ismail, U.; Shah, A.; Ravandi, A. Impact of myocardial reperfusion on human plasma lipidome. iScience 2022, 25, 103828. [Google Scholar] [CrossRef]
- Xuan, C.; Li, H.; Tian, Q.W.; Guo, J.J.; He, G.W.; Lun, L.M.; Wang, Q. Quantitative Assessment of Serum Amino Acids and Association with Early-Onset Coronary Artery Disease. Clin. Interv. Aging 2021, 16, 465–474. [Google Scholar] [CrossRef]
- Xue, Y.; Jiang, W.; Ma, Q.; Wang, X.; Jia, P.; Li, Q.; Chen, S.; Song, B.; Wang, Y.; Zhang, J.; et al. U-shaped association between plasma sphingosine-1-phosphate levels and mortality in patients with chronic systolic heart failure: A prospective cohort study. Lipids Health Dis. 2020, 19, 125. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Liu, J.; Zhang, Q.; Zhong, W.; Li, B.; Li, C.; Liu, Z.; Yang, M.; Zhao, P. Targeted metabolomic analysis of plasma metabolites in patients with coronary heart disease in southern China. Medicine 2019, 98, e14309. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Canela, M.; Guasch-Ferré, M.; Razquin, C.; Toledo, E.; Hernández-Alonso, P.; Clish, C.B.; Li, J.; Wittenbecher, C.; Dennis, C.; Alonso-Gómez, Á.; et al. Plasma acylcarnitines and risk of incident heart failure and atrial fibrillation: The Prevención con dieta mediterránea study. Rev. Esp. Cardiol. 2022, 75, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Gander, J.; Carrard, J.; Gallart-Ayala, H.; Borreggine, R.; Teav, T.; Infanger, D.; Colledge, F.; Streese, L.; Wagner, J.; Klenk, C.; et al. Metabolic Impairment in Coronary Artery Disease: Elevated Serum Acylcarnitines Under the Spotlights. Front. Cardiovasc. Med. 2021, 8, 792350. [Google Scholar] [CrossRef] [PubMed]
- Wittenbecher, C.; Cuadrat, R.; Johnston, L.; Eichelmann, F.; Jäger, S.; Kuxhaus, O.; Prada, M.; Del Greco, M.F.; Hicks, A.A.; Hoffman, P.; et al. Dihydroceramide- and ceramide-profiling provides insights into human cardiometabolic disease etiology. Nat. Commun. 2022, 13, 936. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Liu, M.L.; Huang, B.; Zhao, F.R.; Li, Y.; Cui, X.T.; Lin, R. Acetylcarnitine Is Associated with Cardiovascular Disease Risk in Type 2 Diabetes Mellitus. Front. Endocrinol. 2021, 12, 806819. [Google Scholar] [CrossRef] [PubMed]
- Eichelmann, F.; Sellem, L.; Wittenbecher, C.; Jäger, S.; Kuxhaus, O.; Prada, M.; Cuadrat, R.; Jackson, K.G.; Lovegrove, J.A.; Schulze, M.B. Deep Lipidomics in Human Plasma: Cardiometabolic Disease Risk and Effect of Dietary Fat Modulation. Circulation 2022, 146, 21–35. [Google Scholar] [CrossRef]
- Zheng, D.-m.; An, Z.-n.; Ge, M.-h.; Wei, D.-z.; Jiang, D.-w.; Xing, X.-j.; Shen, X.-l.; Liu, C. Medium & long-chain acylcarnitine’s relation to lipid metabolism as potential predictors for diabetic cardiomyopathy: A metabolomic study. Lipids Health Dis. 2021, 20, 151. [Google Scholar] [CrossRef]
- Ottosson, F.; Emami Khoonsari, P.; Gerl, M.J.; Simons, K.; Melander, O.; Fernandez, C. A plasma lipid signature predicts incident coronary artery disease. Int. J. Cardiol. 2021, 331, 249–254. [Google Scholar] [CrossRef]
- Cavus, E.; Karakas, M.; Ojeda, F.M.; Kontto, J.; Veronesi, G.; Ferrario, M.M.; Linneberg, A.; Jørgensen, T.; Meisinger, C.; Thorand, B.; et al. Association of Circulating Metabolites with Risk of Coronary Heart Disease in a European Population: Results From the Biomarkers for Cardiovascular Risk Assessment in Europe (BiomarCaRE) Consortium. JAMA Cardiol. 2019, 4, 1270–1279. [Google Scholar] [CrossRef]
- Zhou, Q.; Ren, D.; Xiao, Y.; Yi, L.; Zhou, Z. Plasma fatty acid metabolic profiling coupled with clinical research reveals the risk factors for atherosclerosis development in type 2 diabetes mellitus. RSC Adv. 2019, 9, 36162–36170. [Google Scholar] [CrossRef] [PubMed]
- Stenemo, M.; Ganna, A.; Salihovic, S.; Nowak, C.; Sundström, J.; Giedraitis, V.; Broeckling, C.D.; Prenni, J.E.; Svensson, P.; Magnusson, P.K.E.; et al. The metabolites urobilin and sphingomyelin (30:1) are associated with incident heart failure in the general population. ESC Heart Fail. 2019, 6, 764–773. [Google Scholar] [CrossRef] [PubMed]
- Lind, L.; Salihovic, S.; Sundström, J.; Broeckling, C.D.; Magnusson, P.K.; Prenni, J.; Fall, T.; Ärnlöv, J. Multicohort Metabolomics Analysis Discloses 9-Decenoylcarnitine to Be Associated with Incident Atrial Fibrillation. J. Am. Heart Assoc. 2021, 10, e017579. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, L.; Yin, D.; Zhou, Q.; Lv, L.; Dong, Z.; Shi, Y. Integration of proteomic and metabolomic characterization in atrial fibrillation-induced heart failure. BMC Genom. 2022, 23, 789. [Google Scholar] [CrossRef]
- Zhong, W.; Deng, Q.; Deng, X.; Zhong, Z.; Hou, J. Plasma Metabolomics of Acute Coronary Syndrome Patients Based on Untargeted Liquid Chromatography–Mass Spectrometry. Front. Cardiovasc. Med. 2021, 8, 616081. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Li, S.; Sun, G.; Song, L.; Feng, W.; Li, R.; Liu, H.; Dong, Y.; Chen, S.; Yang, S.; et al. Early Warning of Ischemic Stroke Based on Atherosclerosis Index Combined with Serum Markers. J. Clin. Endocrinol. Metab. 2022, 107, 1956–1964. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Chen, X.; Chen, W.; Zhong, L.; Cui, M. Comprehensive plasma metabolomic and lipidomic analyses reveal potential biomarkers for heart failure. Mol. Cell. Biochem. 2021, 476, 3449–3460. [Google Scholar] [CrossRef]
- Ferreira-Divino, L.F.; Suvitaival, T.; Rotbain Curovic, V.; Tofte, N.; Trošt, K.; Mattila, I.M.; Theilade, S.; Winther, S.A.; Hansen, T.W.; Frimodt-Møller, M.; et al. Circulating metabolites and molecular lipid species are associated with future cardiovascular morbidity and mortality in type 1 diabetes. Cardiovasc. Diabetol. 2022, 21, 135. [Google Scholar] [CrossRef]
- Xue, H.; Chen, X.; Yu, C.; Deng, Y.; Zhang, Y.; Chen, S.; Chen, X.; Chen, K.; Yang, Y.; Ling, W. Gut Microbially Produced Indole-3-Propionic Acid Inhibits Atherosclerosis by Promoting Reverse Cholesterol Transport and Its Deficiency Is Causally Related to Atherosclerotic Cardiovascular Disease. Circ. Res. 2022, 131, 404–420. [Google Scholar] [CrossRef]
- Zhao, M.; Wei, H.; Li, C.; Zhan, R.; Liu, C.; Gao, J.; Yi, Y.; Cui, X.; Shan, W.; Ji, L.; et al. Gut microbiota production of trimethyl-5-aminovaleric acid reduces fatty acid oxidation and accelerates cardiac hypertrophy. Nat. Commun. 2022, 13, 1757. [Google Scholar] [CrossRef]
- Shah, H.S.; Moreno, L.O.; Morieri, M.L.; Tang, Y.; Mendonca, C.; Jobe, J.M.; Thacker, J.B.; Mitri, J.; Monti, S.; Niewczas, M.A.; et al. Serum Orotidine: A Novel Biomarker of Increased CVD Risk in Type 2 Diabetes Discovered Through Metabolomics Studies. Diabetes Care 2022, 45, 1882–1892. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, Y.; Li, X.; Deng, X.; Kong, Y.; Wang, W.; Zhou, Y. Machine learning of plasma metabolome identifies biomarker panels for metabolic syndrome: Findings from the China Suboptimal Health Cohort. Cardiovasc. Diabetol. 2022, 21, 288. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.P.; Mishra, B.H.; Lyytikäinen, L.-P.; Hilvo, M.; Juonala, M.; Kähönen, M.; Hutri-Kähönen, N.; Fotiadis, D.I.; Raitakari, O.T.; Laaksonen, R.; et al. Assessment of plasma ceramides as predictor for subclinical atherosclerosis. Atheroscler. Plus 2021, 45, 25–31. [Google Scholar] [CrossRef] [PubMed]
- McGurk, K.A.; Keavney, B.D.; Nicolaou, A. Circulating ceramides as biomarkers of cardiovascular disease: Evidence from phenotypic and genomic studies. Atherosclerosis 2021, 327, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Beckonert, O.; Keun, H.C.; Ebbels, T.M.D.; Bundy, J.; Holmes, E.; Lindon, J.C.; Nicholson, J.K. Metabolic profiling, metabolomic and metabonomic procedures for NMR spectroscopy of urine, plasma, serum and tissue extracts. Nat. Protoc. 2007, 2, 2692–2703. [Google Scholar] [CrossRef] [PubMed]
- de Mello, V.D.F.; Lankinen, M.; Schwab, U.; Kolehmainen, M.; Lehto, S.; Seppänen-Laakso, T.; Oresic, M.; Pulkkinen, L.; Uusitupa, M.; Erkkilä, A.T. Link between plasma ceramides, inflammation and insulin resistance: Association with serum IL-6 concentration in patients with coronary heart disease. Diabetologia 2009, 52, 2612–2615. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, I.; Mastrandrea, L.D. Serum sphingolipids and inflammatory mediators in adolescents at risk for metabolic syndrome. Endocrine 2012, 41, 442–449. [Google Scholar] [CrossRef] [PubMed]
- Holland, W.L.; Adams, A.C.; Brozinick, J.T.; Bui, H.H.; Miyauchi, Y.; Kusminski, C.M.; Bauer, S.M.; Wade, M.; Singhal, E.; Cheng, C.C.; et al. An FGF21-adiponectin-ceramide axis controls energy expenditure and insulin action in mice. Cell Metab. 2013, 17, 790–797. [Google Scholar] [CrossRef]
- Vandanmagsar, B.; Youm, Y.-H.; Ravussin, A.; Galgani, J.E.; Stadler, K.; Mynatt, R.L.; Ravussin, E.; Stephens, J.M.; Dixit, V.D. The NLRP3 inflammasome instigates obesity-induced inflammation and insulin resistance. Nat. Med. 2011, 17, 179–188. [Google Scholar] [CrossRef]
- Meikle, P.J.; Wong, G.; Tsorotes, D.; Barlow, C.K.; Weir, J.M.; Christopher, M.J.; MacIntosh, G.L.; Goudey, B.; Stern, L.; Kowalczyk, A.; et al. Plasma lipidomic analysis of stable and unstable coronary artery disease. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 2723–2732. [Google Scholar] [CrossRef]
- Peterson, L.R.; Xanthakis, V.; Duncan, M.S.; Gross, S.; Friedrich, N.; Völzke, H.; Felix, S.B.; Jiang, H.; Sidhu, R.; Nauck, M.; et al. Ceramide Remodeling and Risk of Cardiovascular Events and Mortality. J. Am. Heart Assoc. 2018, 7, 7931. [Google Scholar] [CrossRef] [PubMed]
- Gui, Y.-K.; Li, Q.; Liu, L.; Zeng, P.; Ren, R.-F.; Guo, Z.-F.; Wang, G.-H.; Song, J.-G.; Zhang, P. Plasma levels of ceramides relate to ischemic stroke risk and clinical severity. Brain Res. Bull. 2020, 158, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Lemaitre, R.N.; Jensen, P.N.; Hoofnagle, A.; McKnight, B.; Fretts, A.M.; King, I.B.; Siscovick, D.S.; Psaty, B.M.; Heckbert, S.R.; Mozaffarian, D.; et al. Plasma ceramides and sphingomyelins in relation to heart failure risk. Circ. Heart Fail. 2019, 12, e005708. [Google Scholar] [CrossRef] [PubMed]
- Park, T.-S.; Hu, Y.; Noh, H.-L.; Drosatos, K.; Okajima, K.; Buchanan, J.; Tuinei, J.; Homma, S.; Jiang, X.-C.; Abel, E.D.; et al. Ceramide is a cardiotoxin in lipotoxic cardiomyopathy. J. Lipid Res. 2008, 49, 2101–2112. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho, L.P.; Tan, S.H.; Ow, G.-S.; Tang, Z.; Ching, J.; Kovalik, J.-P.; Poh, S.C.; Chin, C.-T.; Richards, A.M.; Martinez, E.C.; et al. Plasma Ceramides as Prognostic Biomarkers and Their Arterial and Myocardial Tissue Correlates in Acute Myocardial Infarction. JACC Basic Transl. Sci. 2018, 3, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Chew, W.S.; Torta, F.; Ji, S.; Choi, H.; Begum, H.; Sim, X.; Khoo, C.M.; Khoo, E.Y.H.; Ong, W.-Y.; Van Dam, R.M.; et al. Large-scale lipidomics identifies associations between plasma sphingolipids and T2DM incidence. JCI Insight 2019, 5, 126925. [Google Scholar] [CrossRef] [PubMed]
- Yaribeygi, H.; Bo, S.; Ruscica, M.; Sahebkar, A. Ceramides and diabetes mellitus: An update on the potential molecular relationships. Diabet. Med. 2020, 37, 11–19. [Google Scholar] [CrossRef]
- Sokolowska, E.; Blachnio-Zabielska, A. The Role of Ceramides in Insulin Resistance. Front. Endocrinol. 2019, 10, 577. [Google Scholar] [CrossRef]
- Raichur, S.; Wang, S.T.; Chan, P.W.; Li, Y.; Ching, J.; Chaurasia, B.; Dogra, S.; Öhman, M.K.; Takeda, K.; Sugii, S.; et al. CerS2 Haploinsufficiency Inhibits β-Oxidation and Confers Susceptibility to Diet-Induced Steatohepatitis and Insulin Resistance. Cell Metab. 2014, 20, 919. [Google Scholar] [CrossRef]
- Turpin, S.M.; Nicholls, H.T.; Willmes, D.M.; Mourier, A.; Brodesser, S.; Wunderlich, C.M.; Mauer, J.; Xu, E.; Hammerschmidt, P.; Brönneke, H.S.; et al. Obesity-induced CerS6-dependent C16:0 ceramide production promotes weight gain and glucose intolerance. Cell Metab. 2014, 20, 678–686. [Google Scholar] [CrossRef]
- Menuz, V.; Howell, K.S.; Gentina, S.; Epstein, S.; Riezman, I.; Fornallaz-Mulhauser, M.; Hengartner, M.O.; Gomez, M.; Riezman, H.; Martinou, J.-C. Protection of C. elegans from anoxia by HYL-2 ceramide synthase. Science 2009, 324, 381–384. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-W.; Park, W.-J.; Futerman, A.H. Ceramide synthases as potential targets for therapeutic intervention in human diseases. Biochim. Biophys. Acta 2014, 1841, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Kauhanen, D.; Sysi-Aho, M.; Koistinen, K.M.; Laaksonen, R.; Sinisalo, J.; Ekroos, K. Development and validation of a high-throughput LC-MS/MS assay for routine measurement of molecular ceramides. Anal. Bioanal. Chem. 2016, 408, 3475–3483. [Google Scholar] [CrossRef] [PubMed]
- Basit, A.; Piomelli, D.; Armirotti, A. Rapid evaluation of 25 key sphingolipids and phosphosphingolipids in human plasma by LC-MS/MS. Anal. Bioanal. Chem. 2015, 407, 5189–5198. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Hao, S.; Yao, X.; You, J.; Li, X.; Lai, D.; Han, C.; Schilling, J.; Hwa, K.Y.; Thyparambil, S.; et al. Quantitative LCMS for ceramides/dihydroceramides: Pregnancy baseline biomarkers and potential metabolic messengers. bioRxiv 2020. [Google Scholar] [CrossRef]
- Sigruener, A.; Kleber, M.E.; Heimerl, S.; Liebisch, G.; Schmitz, G.; Maerz, W. Glycerophospholipid and sphingolipid species and mortality: The Ludwigshafen Risk and Cardiovascular Health (LURIC) study. PLoS ONE 2014, 9, e85724. [Google Scholar] [CrossRef] [PubMed]
- Laaksonen, R.; Ekroos, K.; Sysi-Aho, M.; Hilvo, M.; Vihervaara, T.; Kauhanen, D.; Suoniemi, M.; Hurme, R.; März, W.; Scharnagl, H.; et al. Plasma ceramides predict cardiovascular death in patients with stable coronary artery disease and acute coronary syndromes beyond LDL-cholesterol. Eur. Heart J. 2016, 37, 1967–1976. [Google Scholar] [CrossRef] [PubMed]
- Meeusen, J.W.; Donato, L.J.; Bryant, S.C.; Baudhuin, L.M.; Berger, P.B.; Jaffe, A.S. Plasma ceramides. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 1933–1939. [Google Scholar] [CrossRef]
- Havulinna, A.S.; Sysi-Aho, M.; Hilvo, M.; Kauhanen, D.; Hurme, R.; Ekroos, K.; Salomaa, V.; Laaksonen, R. Circulating Ceramides Predict Cardiovascular Outcomes in the Population-Based FINRISK 2002 Cohort. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 2424–2430. [Google Scholar] [CrossRef]
- Hilvo, M.; Salonurmi, T.; Havulinna, A.S.; Kauhanen, D.; Pedersen, E.R.; Tell, G.S.; Meyer, K.; Teeriniemi, A.-M.; Laatikainen, T.; Jousilahti, P.; et al. Ceramide stearic to palmitic acid ratio predicts incident diabetes. Diabetologia 2018, 61, 1424–1434. [Google Scholar] [CrossRef]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; Dugar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.-M.; et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Gregory, J.C.; Org, E.; Buffa, J.A.; Gupta, N.; Wang, Z.; Li, L.; Fu, X.; Wu, Y.; Mehrabian, M.; et al. Gut Microbial Metabolite TMAO Enhances Platelet Hyperreactivity and Thrombosis Risk. Cell 2016, 165, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.H.W.; Wang, Z.; Kennedy, D.J.; Wu, Y.; Buffa, J.A.; Agatisa-Boyle, B.; Li, X.S.; Levison, B.S.; Hazen, S.L. Gut microbiota-dependent trimethylamine N-oxide (TMAO) pathway contributes to both development of renal insufficiency and mortality risk in chronic kidney disease. Circ. Res. 2015, 116, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Lever, M.; George, P.M.; Slow, S.; Bellamy, D.; Young, J.M.; Ho, M.; McEntyre, C.J.; Elmslie, J.L.; Atkinson, W.; Molyneux, S.L.; et al. Betaine and Trimethylamine-N-Oxide as Predictors of Cardiovascular Outcomes Show Different Patterns in Diabetes Mellitus: An Observational Study. PLoS ONE 2014, 9, e114969. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Nemet, I.; Wang, Z.; Lai, H.T.M.; de Oliveira Otto, M.C.; Lemaitre, R.N.; Fretts, A.M.; Sotoodehnia, N.; Budoff, M.; DiDonato, J.A.; et al. Longitudinal Plasma Measures of Trimethylamine N-Oxide and Risk of Atherosclerotic Cardiovascular Disease Events in Community-Based Older Adults. J. Am. Heart Assoc. 2021, 10, e020646. [Google Scholar] [CrossRef] [PubMed]
- Roncal, C.; Martínez-Aguilar, E.; Orbe, J.; Ravassa, S.; Fernandez-Montero, A.; Saenz-Pipaon, G.; Ugarte, A.; Estella-Hermoso de Mendoza, A.; Rodriguez, J.A.; Fernández-Alonso, S.; et al. Trimethylamine-N-Oxide (TMAO) Predicts Cardiovascular Mortality in Peripheral Artery Disease. Sci. Rep. 2019, 9, 15580. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Levison, B.S.; Hazen, J.E.; Donahue, L.; Li, X.-M.; Hazen, S.L. Measurement of trimethylamine-N-oxide by stable isotope dilution liquid chromatography tandem mass spectrometry. Anal. Biochem. 2014, 455, 35–40. [Google Scholar] [CrossRef]
- Matsuzawa, Y.; Nakahashi, H.; Konishi, M.; Sato, R.; Kawashima, C.; Kikuchi, S.; Akiyama, E.; Iwahashi, N.; Maejima, N.; Okada, K.; et al. Microbiota-derived Trimethylamine N-oxide Predicts Cardiovascular Risk After STEMI. Sci. Rep. 2019, 9, 11647. [Google Scholar] [CrossRef]
- Ringel, C.; Dittrich, J.; Gaudl, A.; Schellong, P.; Beuchel, C.F.; Baber, R.; Beutner, F.; Teren, A.; Engel, C.; Wirkner, K.; et al. Association of plasma trimethylamine N-oxide levels with atherosclerotic cardiovascular disease and factors of the metabolic syndrome. Atherosclerosis 2021, 335, 62–67. [Google Scholar] [CrossRef]
- Bjørnestad, E.Ø.; Dhar, I.; Svingen, G.F.T.; Pedersen, E.R.; Ørn, S.; Svenningsson, M.M.; Tell, G.S.; Ueland, P.M.; Sulo, G.; Laaksonen, R.; et al. Circulating trimethylamine N-oxide levels do not predict 10-year survival in patients with or without coronary heart disease. J. Intern. Med. 2022, 292, 915–924. [Google Scholar] [CrossRef]
- Tobias, D.K.; Lawler, P.R.; Harada, P.H.; Demler, O.V.; Ridker, P.M.; Manson, J.E.; Cheng, S.; Mora, S. Circulating Branched-Chain Amino Acids and Incident Cardiovascular Disease in a Prospective Cohort of US Women. Circ. Genom. Precis. Med. 2018, 11, e002157. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-W.; Zhao, X.; Guo, Y.-L.; Zhu, C.-G.; Wu, N.-Q.; Sun, J.; Liu, G.; Dong, Q.; Li, J.-J. Free fatty acids and cardiovascular outcome: A Chinese cohort study on stable coronary artery disease. Nutr. Metab. 2017, 14, 41. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.; Granger, C.B.; Craig, D.; Haynes, C.; Bain, J.; Stevens, R.D.; Hauser, E.R.; Newgard, C.B.; Kraus, W.E.; Newby, L.K.; et al. Validation of the association between a branched chain amino acid metabolite profile and extremes of coronary artery disease in patients referred for cardiac catheterization. Atherosclerosis 2014, 232, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Gao, D.; Jiang, Y. Function, Detection and Alteration of Acylcarnitine Metabolism in Hepatocellular Carcinoma. Metabolites 2019, 9, 36. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Wang, S.; Zhang, C.; Zhao, Y. Coordinated Modulation of Energy Metabolism and Inflammation by Branched-Chain Amino Acids and Fatty Acids. Front. Endocrinol. 2020, 11, 617. [Google Scholar] [CrossRef] [PubMed]
- Jang, C.; Oh, S.F.; Wada, S.; Rowe, G.C.; Liu, L.; Chan, M.C.; Rhee, J.; Hoshino, A.; Kim, B.; Ibrahim, A.; et al. A branched-chain amino acid metabolite drives vascular fatty acid transport and causes insulin resistance. Nat. Med. 2016, 22, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Newgard, C.B.; An, J.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Lien, L.F.; Haqq, A.M.; Shah, S.H.; Arlotto, M.; Slentz, C.A.; et al. A branched-chain amino acid-related metabolic signature that differentiates obese and lean humans and contributes to insulin resistance. Cell Metab. 2009, 9, 311–326. [Google Scholar] [CrossRef]
- Newgard, C.B. Interplay between lipids and branched-chain amino acids in development of insulin resistance. Cell Metab. 2012, 15, 606–614. [Google Scholar] [CrossRef]
- Tobias, D.K.; Mora, S.; Verma, S.; Lawler, P.R. Altered branched chain amino acid metabolism: Toward a unifying cardiometabolic hypothesis. Curr. Opin. Cardiol. 2018, 33, 558–564. [Google Scholar] [CrossRef]
- Wang, L.-Y.; Chen, C. Energy metabolism homeostasis in cardiovascular diseases. J. Geriatr. Cardiol. 2021, 18, 1044–1057. [Google Scholar] [CrossRef]
- Savitz, J. The kynurenine pathway: A finger in every pie. Mol. Psychiatry 2020, 25, 131–147. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, D.; Song, P.; Zou, M.-H. Tryptophan-kynurenine pathway is dysregulated in inflammation, and immune activation. Front. Biosci. 2015, 20, 1116–1143. [Google Scholar] [CrossRef]
- Badawy, A.A.B. Kynurenine Pathway of Tryptophan Metabolism: Regulatory and Functional Aspects. Int. J. Tryptophan Res. 2017, 10, 1178646917691938. [Google Scholar] [CrossRef] [PubMed]
- Zakynthinos, E.; Pappa, N. Inflammatory biomarkers in coronary artery disease. J. Cardiol. 2009, 53, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Xian, X.; Wang, Z.; Bi, Y.; Chen, Q.; Han, X.; Tang, D.; Chen, R. Research Progress on the Relationship between Atherosclerosis and Inflammation. Biomolecules 2018, 8, 80. [Google Scholar] [CrossRef] [PubMed]
- Gáspár, R.; Halmi, D.; Demján, V.; Berkecz, R.; Pipicz, M.; Csont, T. Kynurenine Pathway Metabolites as Potential Clinical Biomarkers in Coronary Artery Disease. Front. Immunol. 2021, 12, 768560. [Google Scholar] [CrossRef] [PubMed]
- Siti, H.N.; Kamisah, Y.; Kamsiah, J. The role of oxidative stress, antioxidants and vascular inflammation in cardiovascular disease (a review). Vasc. Pharmacol. 2015, 71, 40–56. [Google Scholar] [CrossRef]
- Goodsaid, F.M.; Frueh, F.W.; Mattes, W. Strategic paths for biomarker qualification. Toxicology 2008, 245, 219–223. [Google Scholar] [CrossRef]
- Fiehn, O.; Robertson, D.; Griffin, J.; van der Werf, M.; Nikolau, B.; Morrison, N.; Sumner, L.W.; Goodacre, R.; Hardy, N.W.; Taylor, C.; et al. The metabolomics standards initiative (MSI). Metabolomics 2007, 3, 175–178. [Google Scholar] [CrossRef]
- Kodra, D.; Pousinis, P.; Vorkas, P.A.; Kademoglou, K.; Liapikos, T.; Pechlivanis, A.; Virgiliou, C.; Wilson, I.D.; Gika, H.; Theodoridis, G. Is Current Practice Adhering to Guidelines Proposed for Metabolite Identification in LC-MS Untargeted Metabolomics? A Meta-Analysis of the Literature. J. Proteome Res. 2022, 21, 590–598. [Google Scholar] [CrossRef]
- Creek, D.J.; Dunn, W.B.; Fiehn, O.; Griffin, J.L.; Hall, R.D.; Lei, Z.; Mistrik, R.; Neumann, S.; Schymanski, E.L.; Sumner, L.W.; et al. Metabolite identification: Are you sure? And how do your peers gauge your confidence? Metabolomics 2014, 10, 350–353. [Google Scholar] [CrossRef]
- Schymanski, E.L.; Jeon, J.; Gulde, R.; Fenner, K.; Ruff, M.; Singer, H.P.; Hollender, J. Identifying small molecules via high resolution mass spectrometry: Communicating confidence. Environ. Sci. Technol. 2014, 48, 2097–2098. [Google Scholar] [CrossRef] [PubMed]
- Chaleckis, R.; Meister, I.; Zhang, P.; Wheelock, C.E. Challenges, progress and promises of metabolite annotation for LC-MS-based metabolomics. Curr. Opin. Biotechnol. 2019, 55, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Considine, E.C.; Salek, R.M. A Tool to Encourage Minimum Reporting Guideline Uptake for Data Analysis in Metabolomics. Metabolites 2019, 9, 43. [Google Scholar] [CrossRef] [PubMed]
- Koelmel, J.P.; Ulmer, C.Z.; Jones, C.M.; Yost, R.A.; Bowden, J.A. Common cases of improper lipid annotation using high-resolution tandem mass spectrometry data and corresponding limitations in biological interpretation. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2017, 1862, 766–770. [Google Scholar] [CrossRef] [PubMed]
- Köfeler, H.C.; Eichmann, T.O.; Ahrends, R.; Bowden, J.A.; Danne-Rasche, N.; Dennis, E.A.; Fedorova, M.; Griffiths, W.J.; Han, X.; Hartler, J.; et al. Quality control requirements for the correct annotation of lipidomics data. Nat. Commun. 2021, 12, 4771. [Google Scholar] [CrossRef] [PubMed]
- McDonald, J.G.; Ejsing, C.S.; Kopczynski, D.; Holčapek, M.; Aoki, J.; Arita, M.; Arita, M.; Baker, E.S.; Bertrand-Michel, J.; Bowden, J.A.; et al. Introducing the Lipidomics Minimal Reporting Checklist. Nat. Metab. 2022, 4, 1086–1088. [Google Scholar] [CrossRef]
- O’Donnell, V.B.; FitzGerald, G.A.; Murphy, R.C.; Liebisch, G.; Dennis, E.A.; Quehenberger, O.; Subramaniam, S.; Wakelam, M.J.O. Steps Toward Minimal Reporting Standards for Lipidomics Mass Spectrometry in Biomedical Research Publications. Circ. Genom. Precis. Med. 2020, 13, e003019. [Google Scholar] [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef]
Figure 1.
The typical workflow of MS-based clinical metabolipidomics analysis in CVD research. (A) Pre-analytical phase for clinical metabolipidomics study includes planning of study design, collection of blood samples, and separation of plasma/serum from whole blood. (B) Lipids and metabolites are extracted from plasma/serum samples, and then subjected to MS-based data acquisition. A schematic representation of the biphasic separation of lipids (top phase) and metabolites (lower phase) using the SIMPLEX [28] method is shown as a reference. (C) Data analysis is performed to obtain differentially expressed features. Further, outcomes of metabolipidomics data can be integrated with clinical or other “omics” data and interpreted biologically to provide insight into the pathophysiological mechanisms of CVD. After cross-validation, these biomarkers can be clinically translated for routine applications.
Figure 1.
The typical workflow of MS-based clinical metabolipidomics analysis in CVD research. (A) Pre-analytical phase for clinical metabolipidomics study includes planning of study design, collection of blood samples, and separation of plasma/serum from whole blood. (B) Lipids and metabolites are extracted from plasma/serum samples, and then subjected to MS-based data acquisition. A schematic representation of the biphasic separation of lipids (top phase) and metabolites (lower phase) using the SIMPLEX [28] method is shown as a reference. (C) Data analysis is performed to obtain differentially expressed features. Further, outcomes of metabolipidomics data can be integrated with clinical or other “omics” data and interpreted biologically to provide insight into the pathophysiological mechanisms of CVD. After cross-validation, these biomarkers can be clinically translated for routine applications.

Figure 2.
Overview of targeted and untargeted approaches used in metabolipidomics. The untargeted approach offers an unbiased approach for identifying novel biomarkers and generating new hypotheses related to CVD biology, while the targeted approach is useful for validation of these hypotheses and for quantification of low-abundant metabolites. A comprehensive analysis of the metabolome using a combination of these approaches will help to further our understanding of metabolite–CVD associations.
Figure 2.
Overview of targeted and untargeted approaches used in metabolipidomics. The untargeted approach offers an unbiased approach for identifying novel biomarkers and generating new hypotheses related to CVD biology, while the targeted approach is useful for validation of these hypotheses and for quantification of low-abundant metabolites. A comprehensive analysis of the metabolome using a combination of these approaches will help to further our understanding of metabolite–CVD associations.


{kind=link}
{kind=link}
Table 1.
Summary of promising lipid and metabolite biomarkers recently discovered from human CVD studies using different analytical approaches.
Table 1.
Summary of promising lipid and metabolite biomarkers recently discovered from human CVD studies using different analytical approaches.
| Analytical Technique | Analytes of Interest | Potential Biomarker | Disease | Case, n | Controls, n | |
|---|---|---|---|---|---|---|
| ||||||
| LC-MS | Ceramides, phospholipids | PC-16:0/22:5, 14:0/22:6, 16:0/16:0; Cer-C16:0, C18:0, C24:1, C24:0 | CVD in ASCVD | 340 #1 | 3449 #1 | [58] |
| 503 #2 | 5448 #2 | |||||
| 105 #3 | 918 #3 | |||||
| Sphingolipids (SPL)s | dhCer C18:0, Cer C22:0, sphingosine, dhSM 24:1, SM- C24:0, C18:0 | CAD | 462 | 212 | [59] | |
| Abundant lipids | LPC 17:1, AC 18:2, TAG 51:0 | STEMI | 80 | 50 | [60] | |
| AAs | 4-Hydroxyproline | CAD | 128 | 64 | [61] | |
| Sphingosine 1-phosphate | Sphingosine 1-phosphate | HF | 210 | 54 | [62] | |
| TMAO, choline, creatinine, carnitine | CAD | 302 | 59 | [63] | ||
| AAs, ACs, polar metabolites | AF: AC- C16:0, C18:0, C18:2 | HF | 326 | 426 | [64] | |
| HF: AC- C14:0, C16:0, C18:0, C18:2 | AF | 509 | 617 | |||
| ACs, BCAAs | AC-C3:0, C4:0, C4:0-OH, C5:0, C5:1-O2, C5:1, C5:0-OH, C6:0, C8:0, C8:0 DC, C8:1, C10:0, C10:1, C12:0, C12:1, C14:1, C16:1, C16:0, C18:1, C20:4; carnitine, valine, isoleucine | CAD | 54 | 116 | [65] | |
| Direct MS | DhCer, Cer a | DhCer C22:2, C26:1; Cer C16:0, C20:0 | CVD | 551 | 1137 | [66] |
| ACs | Acetylcarnitine (AC-C2:0) | CVD in T2D | 356 | 676 | [67] | |
| Abundant lipids a,b | CE-20:(0 to 3), 22:(0 to 4), 24:0, 24:1; FFA- 15:0, 16:0, 18:4, 20:0, 20:3, 20:4, 22:1, 24:0, 24:1; MAG- 14:0, 15:0, 20:0, 18:1, 18:2, 18:3, 20:3, 20:4; 20:5; FAs: DAG 16:0, 18:0, 22:4; TAG 18:0; PE 16:1, 18:0, 22:4, P-22:4; P-22:5; Cer 14:0; dhCer 22:2; PC 20:2; PI 22:4; LacCer 16:0, 18:0, 18:1, 20:1, 26:0; HexCer 18:1, 24:1 | CVD | 551 | 1148 | [68] | |
| ACs | AC- 14:0, 12:0, 14:0 DC, 14:0-OH, 20:0, 18:0, 16:1-OH, 8:0, 6:0, 10:0 | DCM | 50 | 50 | [69] | |
| Abundant lipids c | PC 15:0_18:2, 17:0_20:3, 16:0_20:1, O-16:2_18:0; SM 34:1; CE 18:0, DAG 18:1_18:3; PI 16:0_20:4 | CAD | 536 | 3329 | [70] | |
| LC-MS, Direct MS | AAs, biogenic amines, glycerophospholipids sugars, ACs, SPLs d | PC O-40:6, O-38:6, 38:6, 38:5, 40:6 | CAD | 2166 | 8575 | [71] |
| GC-MS | FAs | FA 20:0, 22:6n-3 | ASCVD in T2D | 44 | 74 | [72] |
| ||||||
| LC-MS | Urobilin; SM 30:1 | Incident HF | 74 #4 183 #5 84 #6 | 920 #4 1121 #5 1797 #6 | [73] | |
| 9-Decenoylcarnitine | Prevalent AF | 38 #4 20 #5 35 #6 | 897 #4 1118 #5 1935 #6 | [74] | ||
| Diosmetin, α-Ergocryptine, Fludrocortisone acetate, Quinic acid, 2,6-ditert-butylbenzoquinone | AF with HF | 20 | 20 | [75] | ||
| 5-oxo-D-proline, LPC 20:4, creatinine, LPE 16:0 | ACS | 284 | 130 | [76] | ||
| SM 18:0/14:0, LPC 18:0/0:0, 1-Methylpyrrolidinium, PC18:2/18:2, 18:0/18:0 | IS | Discovery: | [77] | |||
| 120 | 40 | |||||
| Validation: | ||||||
| 160 | 80 | |||||
| LPC 18:2, CE 18:1, alanine, choline, fructose | HF | 87 | 49 | [78] | ||
| LC-MS, GC.GC-MS | 4-hydroxyphenyl acetic acid, Threonine, PC 36:2, PC-O 34:2, 34:3, SM 34:1 | CVD in T1D | 95 | 669 | [79] | |
| ||||||
| LC-MS | Indole-3-Propionic Acid | CAD ASCVD | 30 | 30 | [80] | |
| 256 | 256 | |||||
| Trimethyl-5 amino valeric acid | Incident HF | 1647 | [81] | |||
| Orotidine e | CVD in T2D | Discovery: | [82] | |||
| 113 | 112 | |||||
| Validation: | ||||||
| 300 | 299 | |||||
Abbreviations: n—number of participants. Profiling based on commercial metabolomics platforms: a Complex lipid panel™ Metabolon (Metabolon Inc., Morrisville, NC, USA); b Lipid classes containing specific FA chain length; c Lipotype shotgun lipidomics (Lipotype: Dresden, Germany); d Biocrates (MxP® Quant 500 kit) (Biocrates, Innsbruck, Austria); and e Global discovery panel™ Metabolon (Metabolon Inc., USA). Clinical study names: #1: WECAC; #2: LIPID; #3: KAROLA; #4: PIVUS; #5: ULSAM; #6: TwinGene.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kale, D.; Fatangare, A.; Phapale, P.; Sickmann, A. Blood-Derived Lipid and Metabolite Biomarkers in Cardiovascular Research from Clinical Studies: A Recent Update. Cells 2023, 12, 2796. https://doi.org/10.3390/cells12242796
AMA Style
Kale D, Fatangare A, Phapale P, Sickmann A. Blood-Derived Lipid and Metabolite Biomarkers in Cardiovascular Research from Clinical Studies: A Recent Update. Cells. 2023; 12(24):2796. https://doi.org/10.3390/cells12242796
Chicago/Turabian StyleKale, Dipali, Amol Fatangare, Prasad Phapale, and Albert Sickmann. 2023. "Blood-Derived Lipid and Metabolite Biomarkers in Cardiovascular Research from Clinical Studies: A Recent Update" Cells 12, no. 24: 2796. https://doi.org/10.3390/cells12242796
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.