1. Introduction
Cancer research has increasingly relied on 3D cell culture, specifically spheroids, as a valuable tool for testing and developing new anti-cancer drugs. Unlike traditional 2D cell culture, spheroids more accurately mimic the morphological and physiological characteristics of the in vivo microenvironment, which are critical factors influencing therapeutic efficacy [
1]. This conformation allows cells to grow and enables surface receptors to interact with neighboring cells, influencing signal translation and gene expression [
2]. Spheroids formed with a single cell type are primarily used to investigate tumor microenvironment regulators and assess responsiveness to therapy, including metabolic and proliferative gradients that can alter the sensitivity of hypoxic tumor cells or their resistance to chemotherapy [
3].
However, monocultures may not fully represent the characteristics found in the tumor microenvironment. The tumor microenvironment (TME) plays a significant role in cancer development and therapeutic resistance. It consists of two essential components: a cellular compartment that includes fibroblasts, myofibroblasts, endothelial cells, pericytes, smooth muscle cells, adipocytes, macrophages, lymphocytes, and mast cells [
4]; and a non-cellular compartment composed of a network of extracellular matrix (ECM) containing matrix proteins, glycoproteins, glycosaminoglycans, proteoglycans, transforming growth factor-β (TGF-β), vascular endothelial growth factor (VEGF), platelet-derived growth factor (PDGF), and hepatocyte growth factor (HGF) [
3,
5,
6].
Fibroblasts secrete growth factors such as hepatocyte growth factor (HGF), fibroblast growth factor (FGFs), and CXCL12 chemokine, which promote the growth and survival of malignant cells and stimulate the migration of other cells into the TME [
7]. Under normal conditions, fibroblasts remain inactive in each organ, but they can be activated in response to signals triggered by the healing process or immune system cells [
8]. Normal fibroblasts associated with cancer cells can be transformed into cancer-associated fibroblasts (CAFs) by a process known as fibroblast corruption [
9,
10].
Studies have shown that the gene expression profile of lung tumor cells changes when cultured with fibroblast-conditioned medium. Shintani et al. demonstrated that lung tumor cells benefit from increased proliferation, invasive ability, and resistance to cisplatin treatment in the presence of fibroblasts [
11].
Another crucial cell population within the TME is represented by the macrophages. They are a heterogeneous population of innate myeloid cells originating from their precursor, the monocyte. These cells may undergo differentiation or polarization in the blood or tissues and can assume multiple phenotypes in response to constant changes in the microenvironment [
12]. Cancer-associated macrophages (TAMs) are cells that drive the tumor inflammatory response, and are generally associated with a poor prognosis [
13]. TAMs are densely organized in hypoxic regions of the tumor, where the same hypoxic microenvironment stimulates cells to overexpress transcription factors such as hypoxia-inducing factor (HIF) or other molecules such as platelet-derived growth factor (PDGF), adrenomedullin (ADM), matrix metalloproteinase (MMPs) and transforming growth factor (TGF-β) [
14,
15]. Therefore, it has been reported that the polarization of TAMs depends on the TME in which they are located, and this is due to the secretion of soluble and insoluble factors [
16]. Co-culture models have shown that the interaction of monocytes with other cell types, specifically non-small cell lung cancer (NSCLC) cells, facilitates macrophage polarization [
17,
18].
Tumor cells can reside in and transform the stroma, alter the surrounding connective tissue, and modify the metabolism of resident cells, producing a permissive stroma [
19]. Several studies have demonstrated the contribution of the tumor stroma in the development and progression of various types of tumors [
6]. Immune and inflammatory cells activate the production of chemokines, cytokines, and exosomes, which lead to local tissue remodeling and can contribute to resistance to chemotherapy [
3,
19].
The study of spheroids in co-culture becomes important, as it is necessary to have a greater context for the interaction that the different cells present in the TEM. Co-culture models prove to be important in the study of tumor physiology, both for the formation of metabolite and chemical gradients, for the creation of a hypoxic environment, and for matrix–cell and cell–cell interactions. Such models can provide valuable insights into the processes that occur in vivo.
This study focuses on the generation of homotypic and heterotypic spheroids using non-small cell lung cancer (NSCLC) cell lines with either fibroblasts (FDH) or macrophages (Mcf), and the characterization of them in terms of generation time, morphological characteristics, cellular conformation, cell viability, cell cycle profile, and migratory capacity. Additionally, the response of both homotypic and heterotypic spheroids to treatment with known chemotherapeutic drugs and compounds with potential chemotherapeutic activity were evaluated. This research provides valuable insights into spheroid behavior and their response to therapeutic interventions.
2. Materials and Methods
2.1. Cell Lines
The LC-HK2 cell line, a human non-small cell lung carcinoma cell line, which was established in our laboratory [
20], the commercial line A549 (non-small cell lung cancer), and THP1 of acute monocytic leukemia were grown in Dulbecco’s modified Eagle’s minimal medium (DMEM) with F-12 nutrients from Sigma, supplemented with 10% fetal bovine serum (Cultilab). The cultures were kept at 37 °C, with the atmosphere containing ~5% CO
2. The culture medium was changed every two to three days, and the cells were subcultured regularly. Subcultures were obtained by dissociation with 0.05% trypsin and 0.02% EDTA solution.
2.2. Primary Culture of Human Fibroblasts (FDH)
A short-term culture of human dermal fibroblasts (FDH) was obtained from discarded breast reduction surgery. Fragments of at least 1 mm of skin were made and placed in bottles by the hanging drop method. After adhesion, they were maintained until the fibroblasts occupied the largest surface of the bottle. Subcultures were obtained by dissociation with a 0.05% trypsin solution and 0.02% EDTA. Discards were obtained from patients (Caucasian women, non-smokers, healthy, between 20 and 45 years old) undergoing mammoplasty surgery in collaboration with Dr. Ricardo Boggio, after approval by the Ethics Committee of the Institute of Biomedical Sciences of the University of São Paulo and after obtaining the free and informed consent term, in accordance with Good Clinical Practices (Resolution n°466/12-Brazil). All the material collected was used in the project, with no discards. CAAE: 84511418.2.0000.5467.
2.3. Differentiation of Monocytes into Macrophages
Differentiation of the THP-1 cell line into macrophages was induced, to differentiate into macrophages with the addition of PMA. A total of 25 nM of PMA was added for each 1 × 106 of cells, and after 48 h the medium was changed and differentiation into macrophages was observed.
2.4. Generation of Homotypic and Heterotypic Spheroids
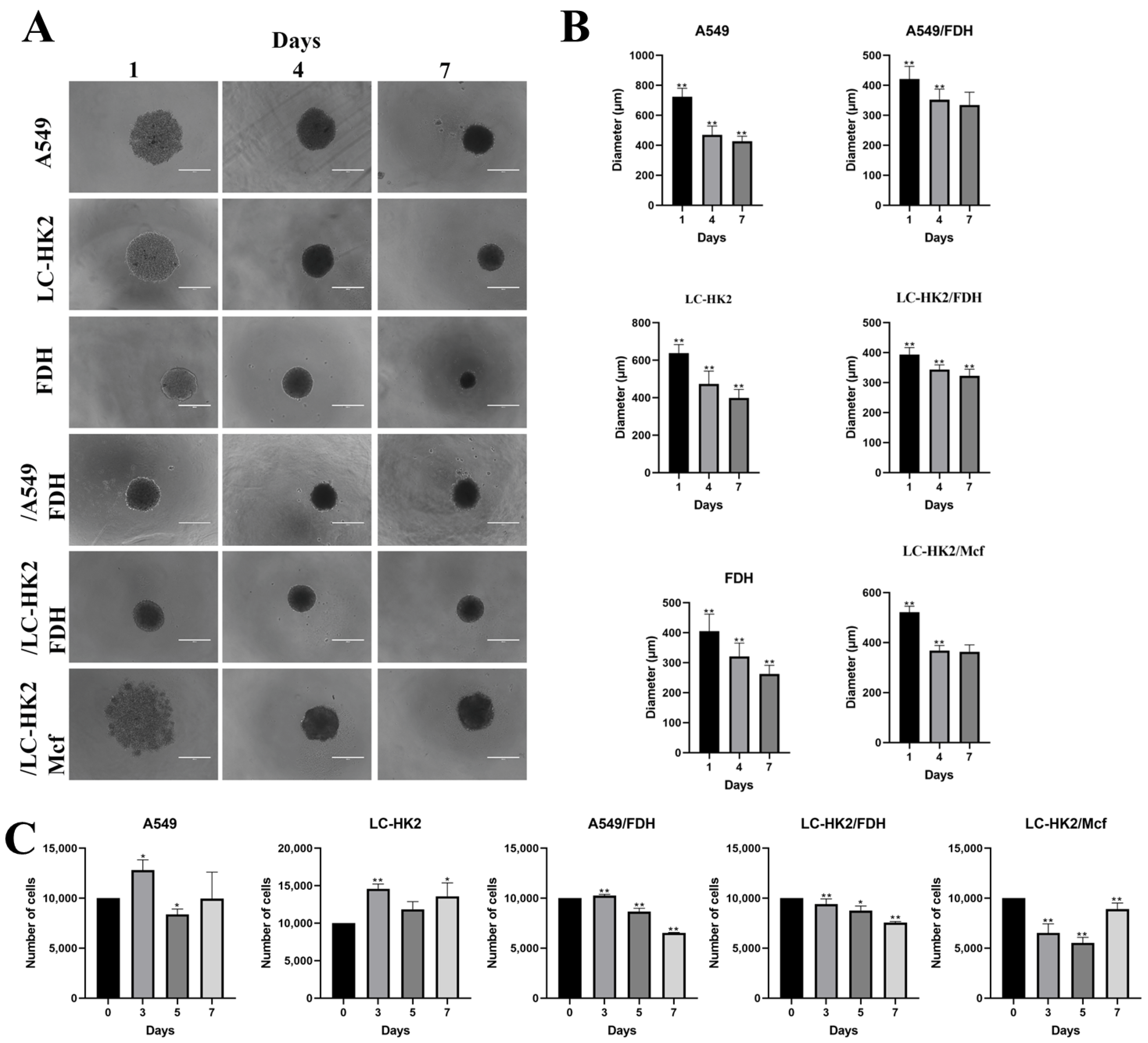
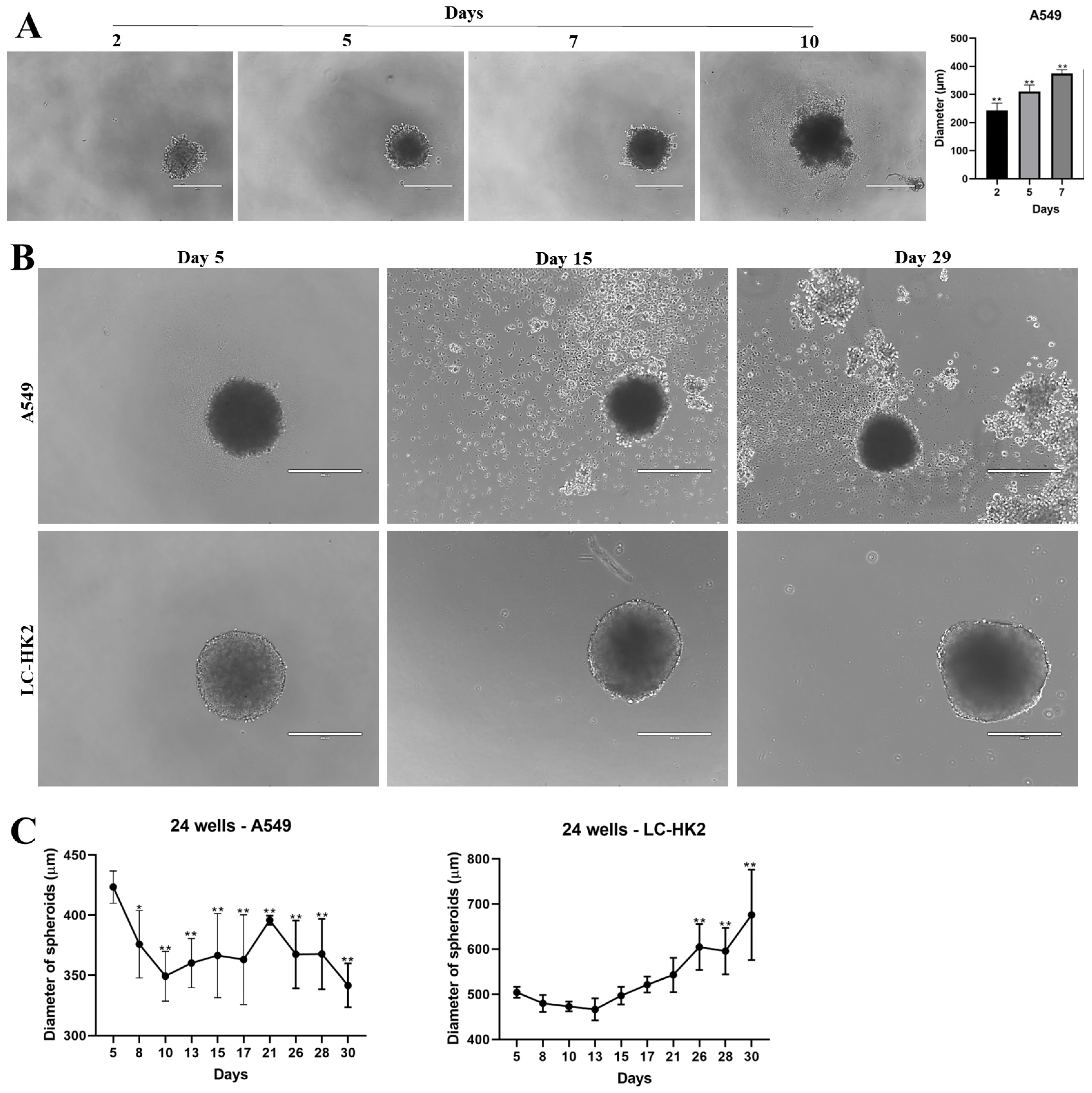
Cells grown in a monolayer were subjected to enzymatic dissociation with a solution of 0.2% trypsin +0.02% EDTA. After neutralizing the action of trypsin with culture medium, the cells were counted in a Guava EasyCycle mini flow cytometer (Millipore Biosciences, Temecula, CA, USA) and 1 × 104 cells were added in 96-well plates with agarose bottom (1%). For the generation of heterotypic spheroids, 5 × 103 cells were plated for each cellular type (1:1 proportion). Spheroids were sustained in DMEM-F12 medium supplemented with 10% FBS; the medium was changed every three days, and the plates with the spheroids were kept in the oven under the conditions mentioned above. The images of the spheroids were obtained by the digital inverted light microscope EVOS AME-3302 (AD, Leusden, The Netherlands).
2.5. Measurement of the Diameter of the Spheroids
The spheroids were kept in the oven for 7 days, after which the spheroids were photographed with an inverted microscope Evos AME-3302 (AD, Leusden, The Netherlands). The measurement of spheroids was performed with the ImageJ program; 10 spheroids were measured on different days in 3 independent experiments.
2.6. Cell Migration Assay
After 7 days in culture, the spheroids were transferred to an adherent surface, and 72 h later, the plates with the migrating cells from the spheroid were photographed, and we proceeded to immunofluorescence reaction, and 5 spheroids were measured in 3 independent experiments using the ImageJ program.
2.7. Viability Assay with Hoechst and PI
After treatments, the spheroids were stained with a solution of HO (Hoechst 33342) and PI (propidium iodide) and left in the study for at least 2 h, after which the 3 spheroids each condition were photographed with a Lionheart microscope from Biotek.
2.8. Cell Cycle Analysis
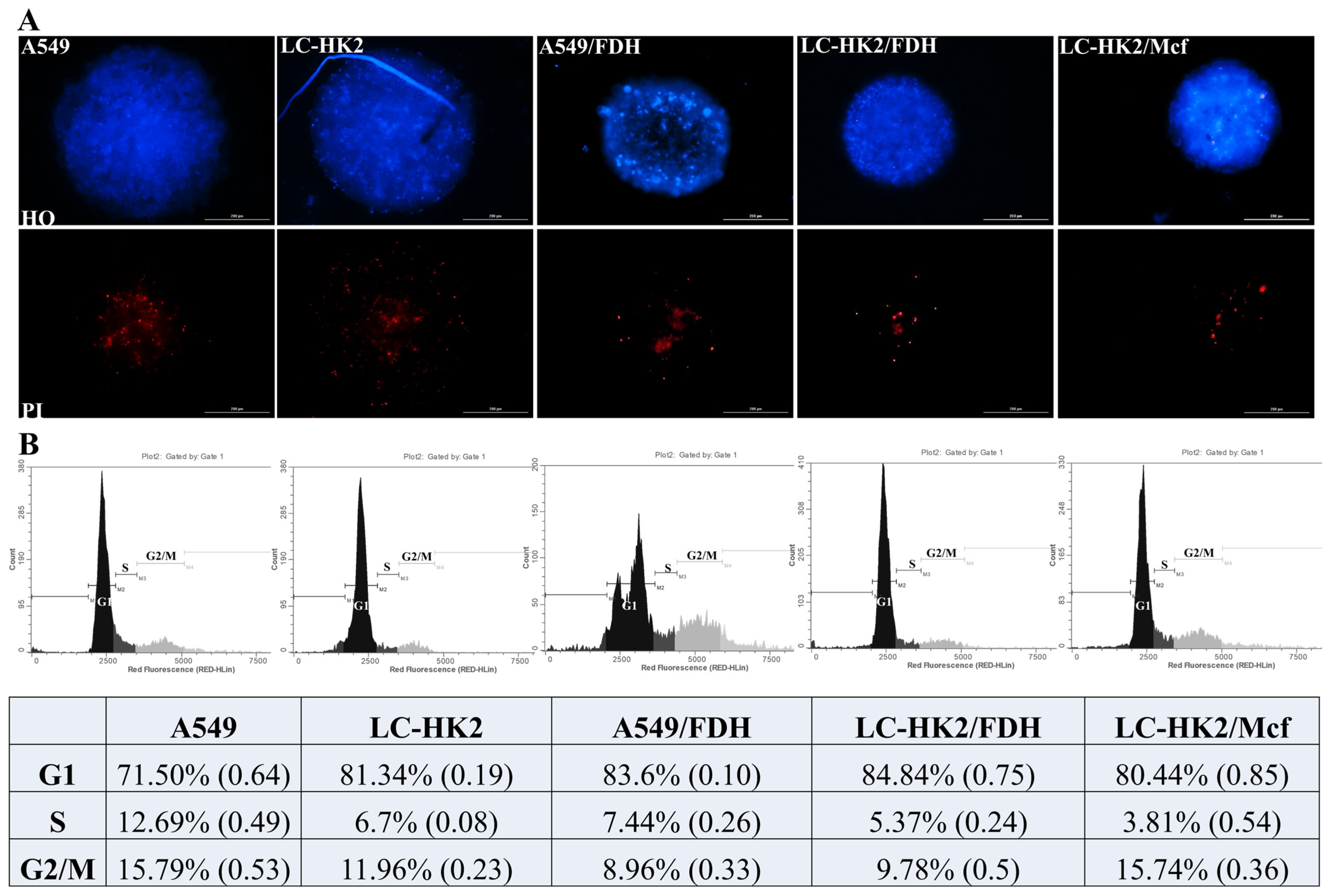
After treatments, spheroid cells were resuspended using trypsin-EDTA, centrifuged at 1200 rpm for 10 min, washed with phosphate buffered saline A (PBSA, without Ca+ and Mg+) and centrifuged. Samples were fixed with 75% methanol at 4 °C for 1h and washed with PBSA. The DNA was stained with PI (10 μg/mL) and treated with RNase (10 µg/mL) at 4 °C for 1 h and quantified by flow cytometer (GUAVA EasyCyte Plus, Hayward, CA, USA). The assay was conducted three times in replicates, and the results were expressed as mean ± SD of the percentage of cell distribution in each cell cycle phase (G1, S, and G2/M).
2.9. Immunofluorescence and Fluorescence Assay
For immunostaining, samples were fixed with 3.7% formaldehyde (Sigma-Aldrich, HAVERHILL, MA, USA) for 30 min, permeabilized with Triton X-100 (0.5%) for 30 min and incubated overnight with primary monoclonal antibodies and for 2 h with secondary antibodies. Nuclei were labeled with DAPI (1:100) or propidium iodide (10 mg/mL) from Sigma-Aldrich (St. Louis, MO, USA). The slides were mounted with Vecta-Shield from Vector Laboratories (Newark, CA, USA) and analyzed by laser scanning confocal microscopy from Leica TCS SP8 (HE, DE) or with fluorescent microscopy Lionheart from Biotek.
2.10. Transmission Electron Microscopy (TEM)
Spheroids were fixed for 2 h with 2.5% glutaraldehyde and 2% formaldehyde in 0.1 M sodium cacodylate buffer, pH 7.2. The fixed samples were washed in 0.1 M sodium cacodylate buffer, pH 7.2, and post-fixed in 1% osmium tetroxide. The tissues were dehydrated in a graded ethanol and propylene oxide series. Resin infiltration was done with a 1:1 mixture of propylene oxide and EPON (Electron Microscopy Science, Hatfield, PA, USA) for 5 h, followed by pure Epon for 5 h. Next, the material was embedded in Epon and polymerized for 48 h. Semi-thin sections were cut using an ultra-microtome and stained with toluidine blue. Appropriate regions of the spheroids were then thin sectioned at 70 and 90 nm and stained with 4% uranyl acetate and a 10% lead citrate solution. The material was analyzed with a Jeol 1010 transmission electron microscope at 80 kV.
2.11. CellTiter-Glo 3D Cell Viability Assay
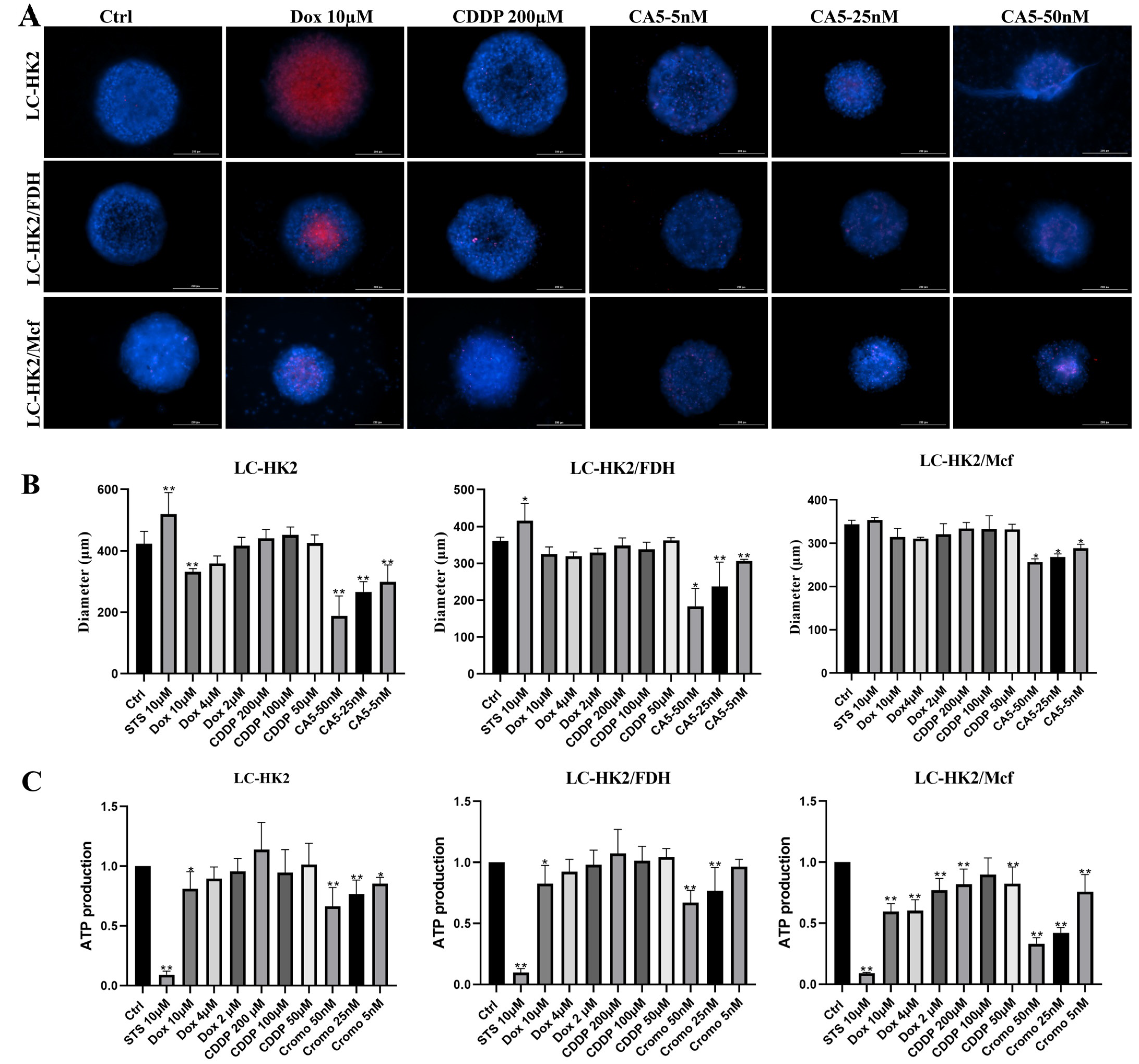
After of treatment for 48h with Staurosporine 10 µM, Doxorubicin (10, 4 and 2 µM), CDDP (cisplatin 200, 100 and 50 µM) and CA5 (50, 25 and 5 nM). The viability of the spheroids was assessed by ATP quantification utilizing the CellTiter-Glo 3D Cell Viability kit (Promega, Madison, WI, USA) according to the manufacturer’s instructions. The measurement represents the average of 4 spheroids per condition in 3 independent experiments.
2.12. Statistical Analysis
Results were expressed as means (SD). Data were submitted to the ANOVA test, followed by Dunnet’s test for multiple comparisons with the control. p-value < 0.05 was considered statistically signific.
4. Discussion
Currently, one of our challenges is developing cell culture models that reflect the conditions found in vivo. This way, we can develop tools that improve the study of drugs and increase their effectiveness against cancer.
Uniform spheroids can be formed by using microwells that prevent cells from adhering to the wells and aggregating between them [
21]. Then, in order to achieve less variability in the generation and size of the spheroids, we can use the liquid overlay method in microwells with an agarose bottom, thus preventing the adhesion of the spheroids to the bottom of the plate. We were able to generate both homotypic and heterotypic spheroids, with diameters showing relatively minor variation. It is noteworthy that the dermal fibroblast spheroids exhibited an even smaller degree of diameter variation. This finding could be attributed to the properties of dermal fibroblasts, which are known to play a crucial role in wound healing and tissue regeneration. The ability to form spheroids with a consistent diameter is crucial for the accurate evaluation of various cellular parameters, such as viability, proliferation, and migration.
Tumor cells can survive in unfavorable conditions such as deprivation of nutrients, oxygen, or growth factors, inhibition by contact with neighboring cells, or loss of anchorage in the cell layer [
6]. Cell adhesion, aggregation, and growth in 3D cultures can be artificially produced by adding extracellular homologs such as collagen or matrigel. Other methods for generating 3D cultures include liquid overlay, fiber mesh made of biocompatible polymers, solid or porous granules, and extracellular matrices and their substitutes. These methods also require the addition of artificially produced supplements. The search for mimicking the ECM in the tumor microenvironment has led to the emergence of different biomaterials, as well as the addition of ECM components [
22,
23].
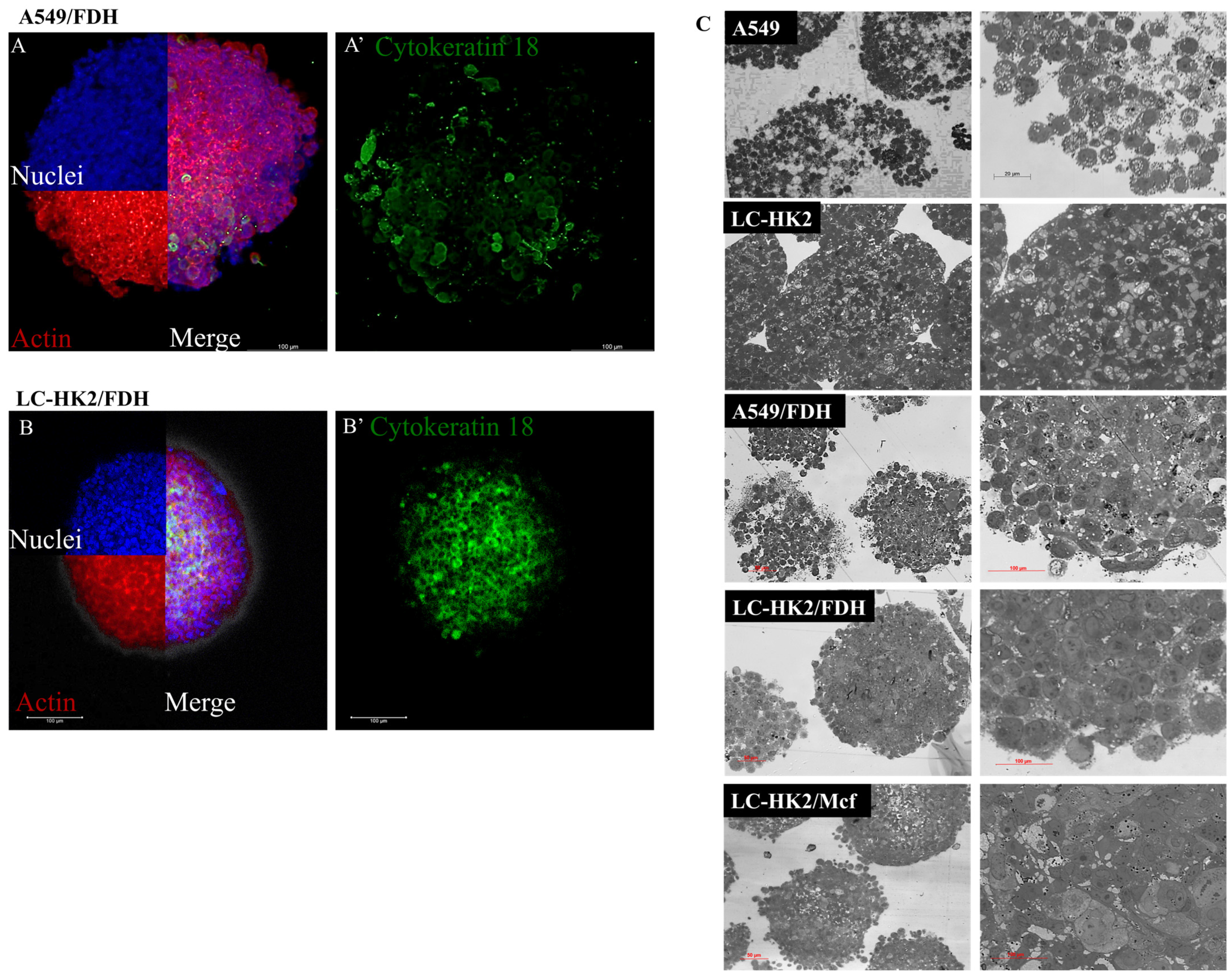
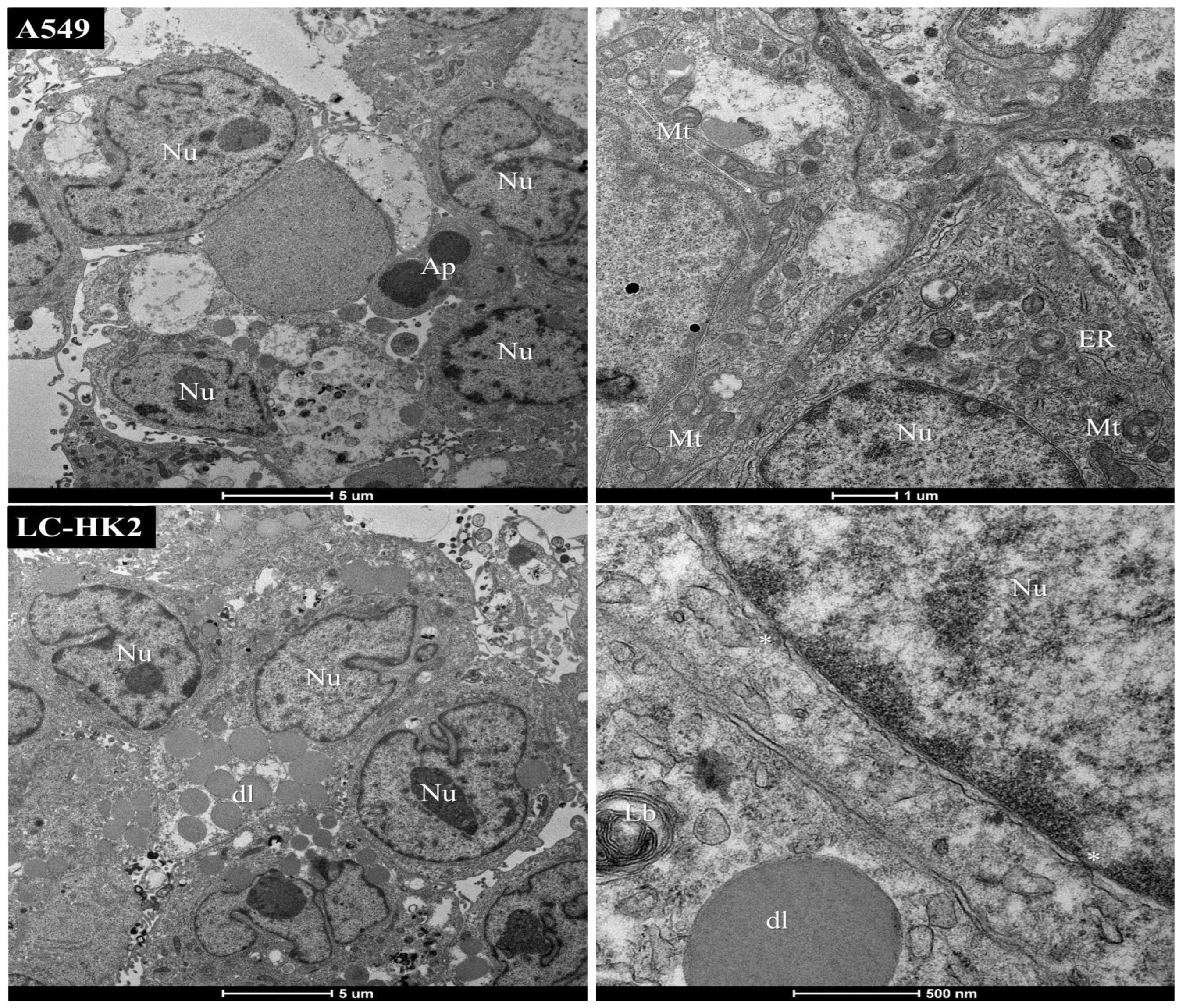
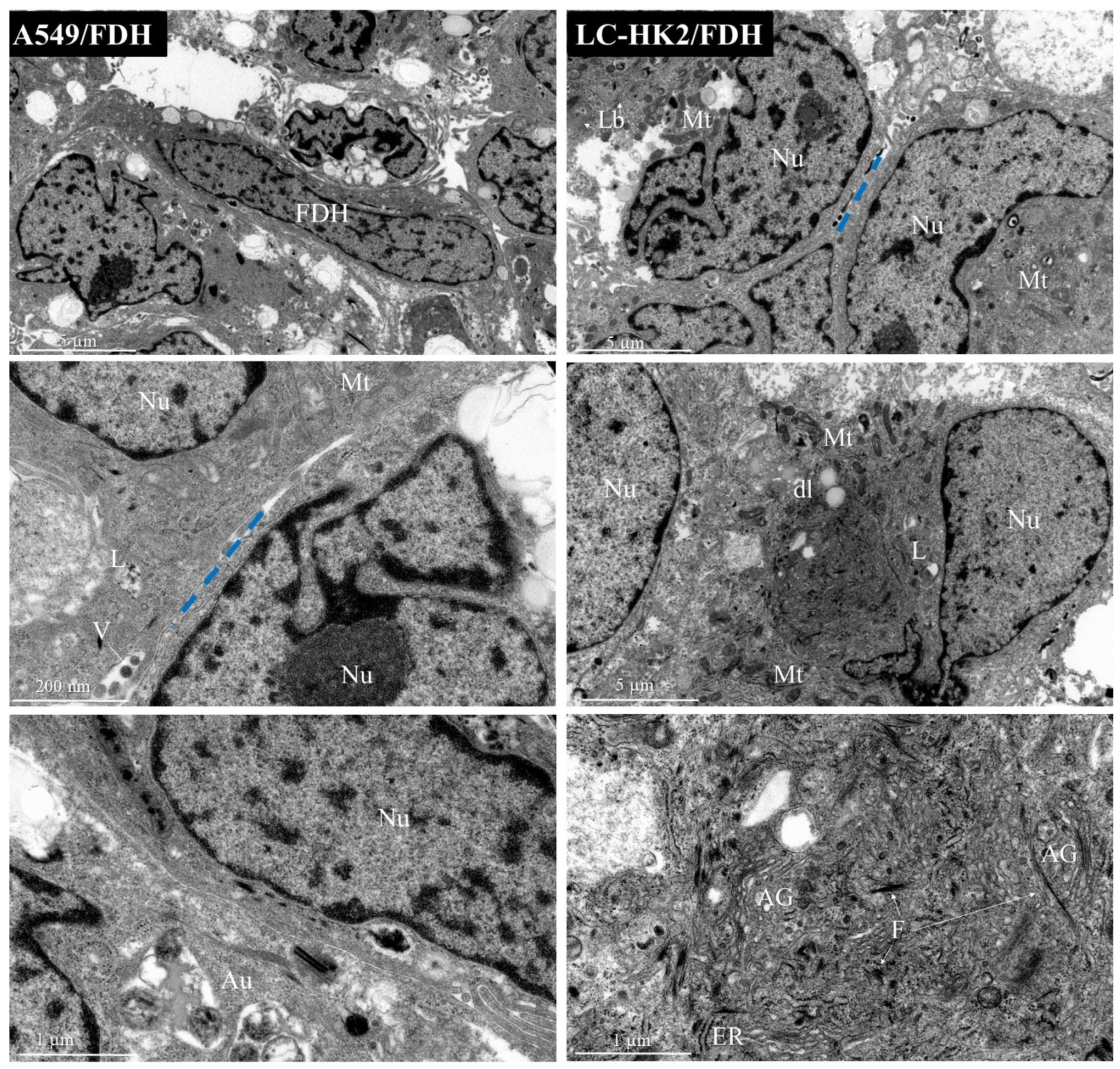
This study sheds light on the contribution of fibroblasts to the complexity of spheroids, as evidenced by cytoplasmic changes observed through transmission electron microscopy of LC-HK2/FDH spheroids. Interestingly, the incorporation of FDH into the spheroid composition was found to expedite the aggregation of the constituent cells, which differs from the aggregation time observed in homotypic spheroids. These findings suggest that FDH association could be leveraged to enhance the efficiency of spheroid generation assays for investigating cellular behavior in 3D microenvironments.
Fibroblasts are responsible for synthesizing and organizing ECM components, generating biological signals, growth factors, angiogenic factors, and remodeling enzymes such as matrix metalloproteinase (MMP) [
24]. The composition and physical properties of the ECM influence the functional behavior, cell growth, morphology, and survival of the cell population. Changes in these properties play a key role in tumor development, progression, and metastasis [
25,
26].
In cancer, fibroblasts remain activated, and there is a failure to remove them by apoptosis, with them thus becoming important contributors to tumorigenesis [
19]. It is currently believed that most active fibroblasts and CAFs are derived from resident fibroblasts that have transdifferentiated in response to factors secreted by tumors such as TGF-β [
27]. The study by Fromigué and colleagues demonstrated that co-culture between A549 cells (NSCLC) and normal fibroblasts induce the expression of ST3, a protein expressed by invading fibroblasts. They also observed an increase in the expression of MMPs [
24].
In addition to playing an important role in ECM deposition, active fibroblasts are responsible for the formation of a capsule that surrounds the tumor. This capsule is associated with a lower capacity for tumor cell invasion, but, at the same time, it makes it more difficult for the chemotherapy drugs to penetrate [
4,
28]. When generating heterotypic spheroids with FDH, it was noticed that the cell resuspension process of the spheroids with trypsin took longer than that of the homotypic spheroids and heterotypic spheroids with macrophages. Thus, we can infer that in this case, the fibroblasts were responsible for a greater deposition of ECM, an issue that can also be observed in the penetration of drugs, since the LC-HK2/FDH spheroids had the lowest decrease in cell viability.
Unlike findings from other studies involving NSCLC and fibroblasts, we were able to form spheroids from the A549 cell line. Additionally, there were differences observed in the morphology of A549 cells and fibroblasts, with the latter having an elongated morphology, as seen in TEM. Curiously, when LC-HK2 cells and short-lived fibroblasts (FDH) were combined to form heterotypic spheroids, no discernible morphological difference between the two cell types was observed. However, this contrast in morphology had already been reported in a previous study involving Colo 699 cell lines (NSCLC) and SV80 fibroblasts (immortalized cell lines). In this same study, the production of ECM was demonstrated and associated with a more rounded morphology, as well as a more homogeneous surface of the spheroid when fibroblasts were added [
23].
The compaction of the spheroids can be attributed to both cellular death and an increase in the production of ECM components, which contribute to enhanced cell cohesion. The phenomenon of cellular death in this case is a result of the anoikis process. It is noteworthy that not all tumor cells are able to withstand anoikis [
6]. It is interesting to observe that FDH cells, despite experiencing cellular death, were able to form tightly packed spheroids.
Spheroids exhibit crucial characteristics, such as a proliferative and metabolic gradient, which impact pharmacological efficacy. The necrotic center in spheroids is formed due to restricted access to nutrients and oxygen, leading to low pH and the presence of metabolites. Cells in the center adapt and become quiescent. Distinct zones were formed within spheroids, with proliferative cells on the outer edges and quiescent and necrotic cells in the center, similar to solid tumors. Spheroids, without an external matrix, secrete extracellular matrix to create a tissue-like microenvironment, resulting in hypoxia [
29,
30].
In the tumor, only a small subset of cells performs an action, while most become dormant or die. These solitary cells are known as dormant tumor cells (DTCs), and they are characterized by their lack of proliferation and death while they remain silent [
3]. In the spheroids generated, a center of dead cells was observed, which can be seen in the viability assay by staining with HO and IP. In addition, a population of quiescent cells was observed, characterized by a high percentage of cells in the G1 phase and a decrease in the S phase in all spheroids. The spheroids studied had all the expected cell populations, including proliferative cells on the outer edge.
In both tumors and organs, the average interstitial oxygen decreases as one moves away from the blood vessels, and different degrees of hypoxia are observed, ranging from mild to severe. This is due to the limited oxygen diffusion in tissue, which depends on the rate of tissue oxygen consumption and is restricted to a tissue thickness of approximately 130 µm [
4]. Spheroids with an average diameter size between 150 and 300 µm show an intermediate normoxic quiescent zone, and a hypoxic zone in the center [
29,
31]. As observed, the spheroids generated had a final diameter between 300 and 400 µm after 7 days in culture, which could indicate zonation within the spheroids.
Hypoxia induces cellular processes reprogramming through hypoxia-inducible factors (HIFs), a family of transcription factors. These HIFs mediate primary adaptive responses to changes in oxygen levels in the environment [
4]. Hypoxia is a strong stimulus in the tumor microenvironment and leads to the activation of autophagy. Cancer cells employ autophagy as a mechanism to support their survival and preserve cell integrity [
32]. The homotypic and heterotypic spheroids also showed cellular processes such as apoptosis and autophagy.
Autophagy is the lysosomal degradation process that packages damaged proteins and organelles into double-membrane vesicles and transports them to the lysosome for degradation [
33]. It is involved in several pathologies and has a dual role in tumorigenesis, both promoting cell progression and resistance to chemotherapy [
34].
Studies of NSCLC have shown that an acidic pH can induce autophagy flux, promoting lung cancer cell survival in vivo and in vitro. This pH can also cause ROS production, resulting in endoplasmic reticulum stress [
33]. The acidic tumor microenvironment contributes to drug resistance by creating a physiological barrier to weakly basic chemotherapy drugs, thereby preventing their access to cancer cells [
35,
36].
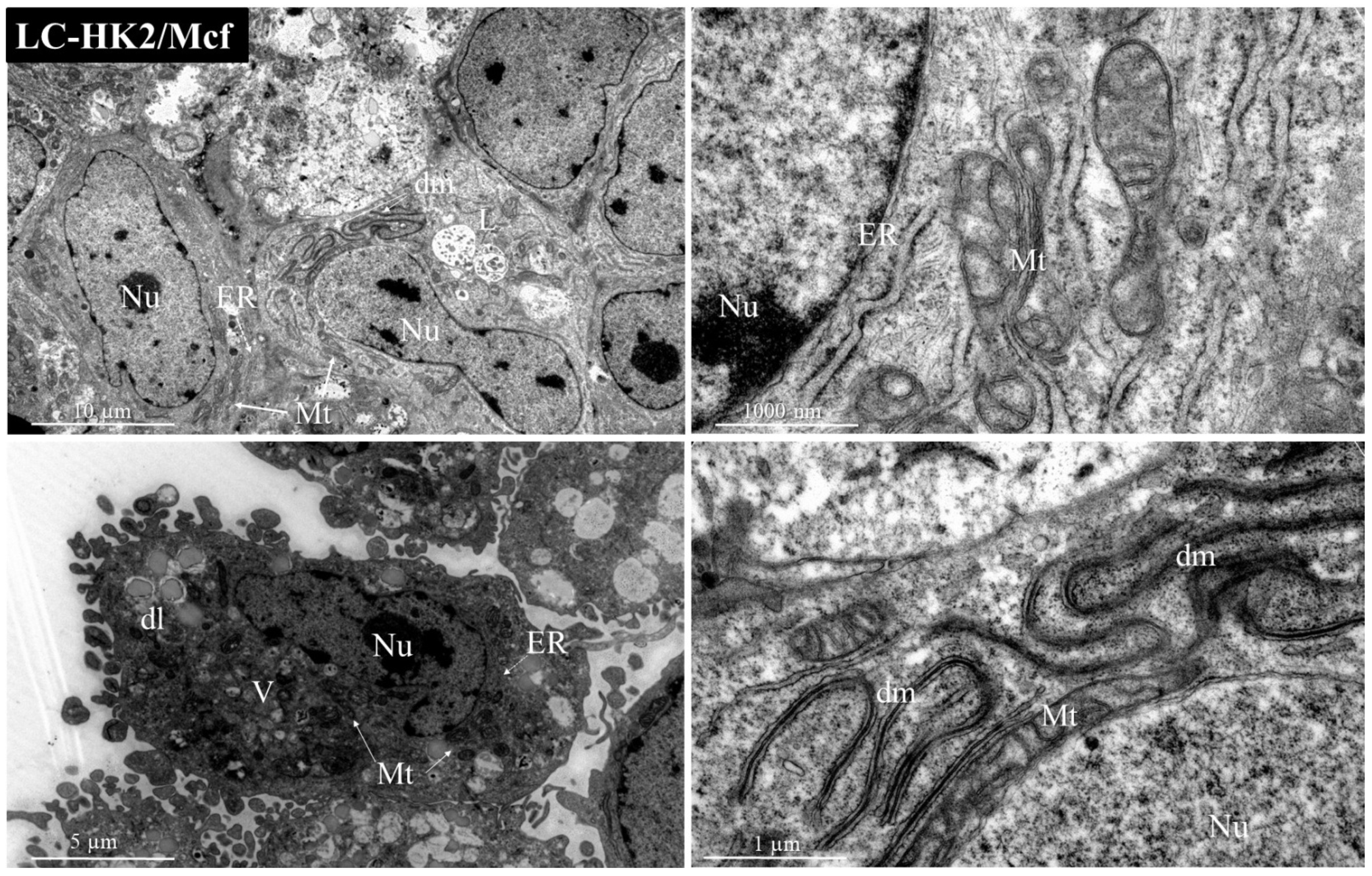
In the homotypic and heterotypic spheroids generated, lamellar bodies were observed. Lamellar bodies are organelles associated with lysosomes to produce surfactant substances, and were also associated with autophagosomes in mouse lung pneumocytes [
37,
38]. These lamellar structures can be present in normal cells, but also in cells that have suffered some type of stress or in pathological conditions [
39].
Many mitochondria were observed, as well as a variety of morphology and size in both homotypic and heterotypic spheroids. The shape, size, and number of mitochondria are controlled by the dynamics of two opposing processes, fission and fusion, and mitochondrial fission is predominant in tumor cells. Mitochondrial fragmentation is a result of excessive fission [
40]. Mitochondria form a network that contacts the nucleus, endoplasmic reticulum, Golgi complex, and cytoskeleton. Mitochondrial morphology changes based on cellular demands, and can include branching, bending, retractions, and changes in the shape and number of cristae [
41].
Cancer mitochondria have heterogeneous ultrastructure, with changes including outer membrane buckling, crystal disorganization, matrix myelin figures, vacuoles, and distorted shapes. Despite these changes, cancer mitochondria do not differ much from normal mitochondria in function, substrate oxidation rate, phosphate ion transport, passive swelling, and other factors [
42]. In tumors such as pancreatic or salivary duct carcinoma and others, many mitochondria have been observed. Mitochondria exhibit variability and abnormalities in number, size, and shape, even within the same specimen, as well as a degree of severity of internal ultrastructural changes [
40]. Moreover, in heterotypic spheroids with macrophages, the presence of double membranes can be observed, which could be an indicator of mitochondrial rearrangement.
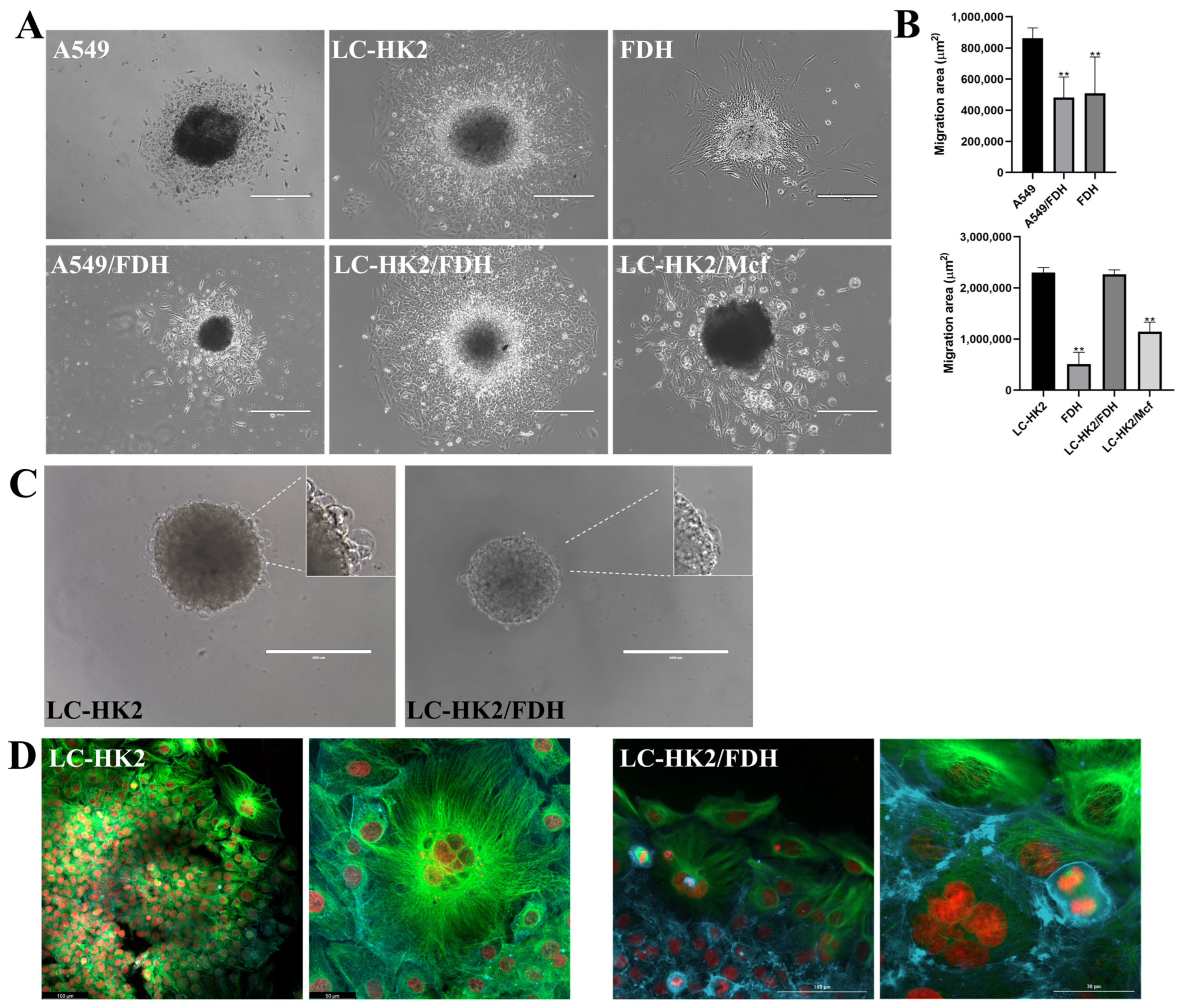
Migrating cells survive in the circulation, with their extravasation into a nearby or distant organ/tissue subsequently colonizing a secondary site [
3]. For invasion and metastasis, the interaction between fibroblasts, immune cells, angiogenic cells, and their factors is essential [
19]. Unlike other studies with spheroids, where the size and invasiveness of cells reflect their migratory capacity [
29], the migration of cells on an adherent surface in LC-HK2 spheroids did not depend on their size. Heterotypic spheroids (LC-HK2/FDH), which had a slightly smaller final diameter than homotypic spheroids (LC-HK2), showed a migration area that was very similar to each other. In contrast, the LC-HK2/Mcf heterotypic spheroids had a smaller migration area than the LC-HK2 spheroids. Previous work on cancer pancreatic cells demonstrated that cells selected for resistance to anoikis had higher rates of migration and invasion than cells that were not selected [
43].
Compared to A549 spheroids, those formed with A549/FDH cells exhibited a smaller migration area, indicating a potential influence of fibroblasts on the migratory response of A549 cells. These findings highlight the impact of spheroid composition on cellular processes, and demonstrate how they can differ depending on the cell type. Specifically, our results emphasize the role of fibroblasts in regulating cell migration and highlight the need for further investigation into complex interactions between different cell types in 3D microenvironments.
The spheroids generated in this study exhibited both common and individual characteristics that are interconnected and subject to further study. The importance of a 3D system that allows for the observation of interactions between different cell types and the creation of necessary characteristics is highlighted. Co-culture models, where different cell types are grown in the same culture dish, provide a greater degree of in vivo similarity compared to monocultures and can be used to study cell–cell interactions ex vivo. However, analyzing cellular interactions becomes challenging, since these interactions change depending on the environment, and others occur within a defined pH range [
22,
30]. Three-dimensional models, such as spheroids, generally reflect cellular behavior in tissues and recapitulate cellular heterogeneity in tumors. The highly compact spheroids formed in this study indicate cell–cell and cell–matrix contact [
21,
44].
LC-HK2/FDH spheroids exhibited enhanced resistance to drug penetration compared to other generated spheroids, even when higher concentrations were used, surpassing those typically employed in monolayer studies. The configuration of spheroids can significantly influence drug delivery and efficacy outcomes. Therefore, it is essential to thoroughly characterize spheroids to minimize variability across experimental setups, enabling a better result comparison and reproducibility.
As observed, the concentration of the compounds used in the spheroids was higher than the typically employed IC50 concentration in monolayer cultures. It is also known that the gene expression profiles of the 3D culture have been shown to more accurately reflect the expression profiles observed in the 2D culture [
31]. The lack of similarity in architecture between monolayer culture and in vivo conditions can lead to erroneous conclusions. Some assays and drugs that exhibit good activity against cancer cells in monolayer culture may not have the same response when translated to clinical trials [
30,
44]. Therefore, it is crucial to include 3D studies in the screening of compounds to support the findings of monolayer studies [
30]. It is important to consider that conducting multiple feasibility tests is necessary to demonstrate, with greater accuracy, the effects of drugs on spheroids. Staurosporine, which was used as cell death control in this study at a high concentration, is a drug isolated from
Streptomyces staurosporeus (now Lentzea albida). Initially, it was characterized as an exceptionally powerful inhibitor of protein kinase C, displaying remarkable potency. However, further studies have revealed that the compound exhibits a high degree of promiscuity, interacting with numerous other kinases in addition to its primary target [
45].
Cisplatin induces cell death in various cell lines through multiple mechanisms. It triggers oxidative stress by generating reactive oxygen species and causing lipid peroxidation. Additionally, cisplatin activates p53 signaling, leading to cell cycle arrest. It down-regulates proto-oncogenes and anti-apoptotic proteins while activating both intrinsic and extrinsic pathways of apoptosis. These combined effects contribute to the cytotoxicity of cisplatin and its ability to eliminate cancer cells [
46,
47]. Cisplatin is a well-known chemotherapeutic widely used in NSCLC, with an IC50 of 22.5 µM in the A549 cell monolayer [
48].
Doxorubicin, like cisplatin, is used in the treatment of NSCLC; it is characterized as a DNA damaging agent, and in A549 cells it has an IC50 of 1.48 µM [
49]. Doxorubicin was extracted from
Streptomyces peucetius. The antitumor activity of doxorubicin (Dox) has been attributed to its ability to intercalate DNA and its association with proteins involved in DNA replication and transcription. Dox is classified as a topoisomerase II poison [
50]. Another mechanism of action of Dox may be its ability to generate free radicals that induce damage to cell membranes and DNA [
51]. The high therapeutic effects and clinical applications of doxorubicin are compromised due to its high hydrophilicity, short half-life, and the requirement of high doses for treatment effectiveness resulting in cardiotoxicity, extravasation, nephrotoxicity, and myelosuppression [
52].
Chromomycin A5, in turn, is a glycosylated tricyclic polyketide, a member of the aureole acid group of antitumor antibiotics. Studies suggest that chromomycin A5 is associated with the transcription factor T-box 2 (TBX2), and could be associated with its anti-proliferative and potential anti-metastatic effects [
53]. CA5 would also act as an inducer of ICD (immunogenic cell death) [
54]. Chromomycins are highly cytotoxic, and can induce autophagy in melanoma cells [
55]. In the monolayer of melanoma, breast cancer, and rhabdomyosarcoma cell lines, CA5 presents an IC50 between 0.8 and 6.5 nM [
53]. In this study, CA5 was the compound that had the most pronounced effect on the spheroid cell viability. This effect was observed through a decrease spheroid size and a reduction in ATP production.
It was observed that cisplatin increases ATP production, indicating changes in metabolic rates, as observed in other studies, but this is not associated with cell viability. Something similar can be observed with doxorubicin, but in an inverse way. In the stained result obtained with HO and IP, a center appears much more stained in red, but it does not reflect what happens with ATP production. When analyzing the response to compounds, it was observed that fibroblasts can not only provide a physical barrier, but also influence the metabolic response. In the case of macrophages, they may interact not as physical barriers, but likely through the production of signals that enable the action of compounds on cell viability. However, further studies on the role of these macrophages are necessary. Homotypic spheroids had a larger diameter than heterotypic spheroids. This difference in size suggested that the penetration of compounds into the spheroids could be different. However, despite the variation in diameter, heterotypic spheroids with fibroblasts responded to the tested compounds similarly to homotypic spheroids in terms of ATP production. It is important to consider the impact of spheroid size when analyzing compound responses.
The observation indicates that cisplatin increased the ATP production, suggesting changes in metabolic rates, which align with findings from other studies. However, this increase in ATP production does not align with cell viability. Similarly, doxorubicin showed a similar pattern, but in an opposite manner. In the viability reactions with HO and IP, the central region of the spheroids was more red-stained. Nevertheless, the staining pattern does not correspond to the observed ATP production or cell viability. It could be just doxorubicin accumulation.
In the analysis of compound responses, it was observed that fibroblasts not only act as physical barriers, but also play a role in the production of signals that facilitate the action of compounds on cell viability. These observations highlight the complexity of the interaction between different cell types and compounds in the experimental setup.
The response to treatments may also be modulated by cell–cell interaction, including the interaction between two different cell types, which can influence the process. Much of our knowledge about how ECM can modulate drug responses and contribute to resistance comes from studies of cancer cell interactions with tumor stroma [
5,
6]. At this stage, the significance lies in generating not only homotypic spheroids, but also heterotypic spheroids composed of stromal cells, along with tumor cells. This combination reflects the interplay between different cell types within the tumor microenvironment and enhances the relevance of studying tumor relapses.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}









