

Management of Rheumatoid Arthritis: Possibilities and Challenges of Mesenchymal Stromal/Stem Cell-Based Therapies
 , ,
, ,
Abstract
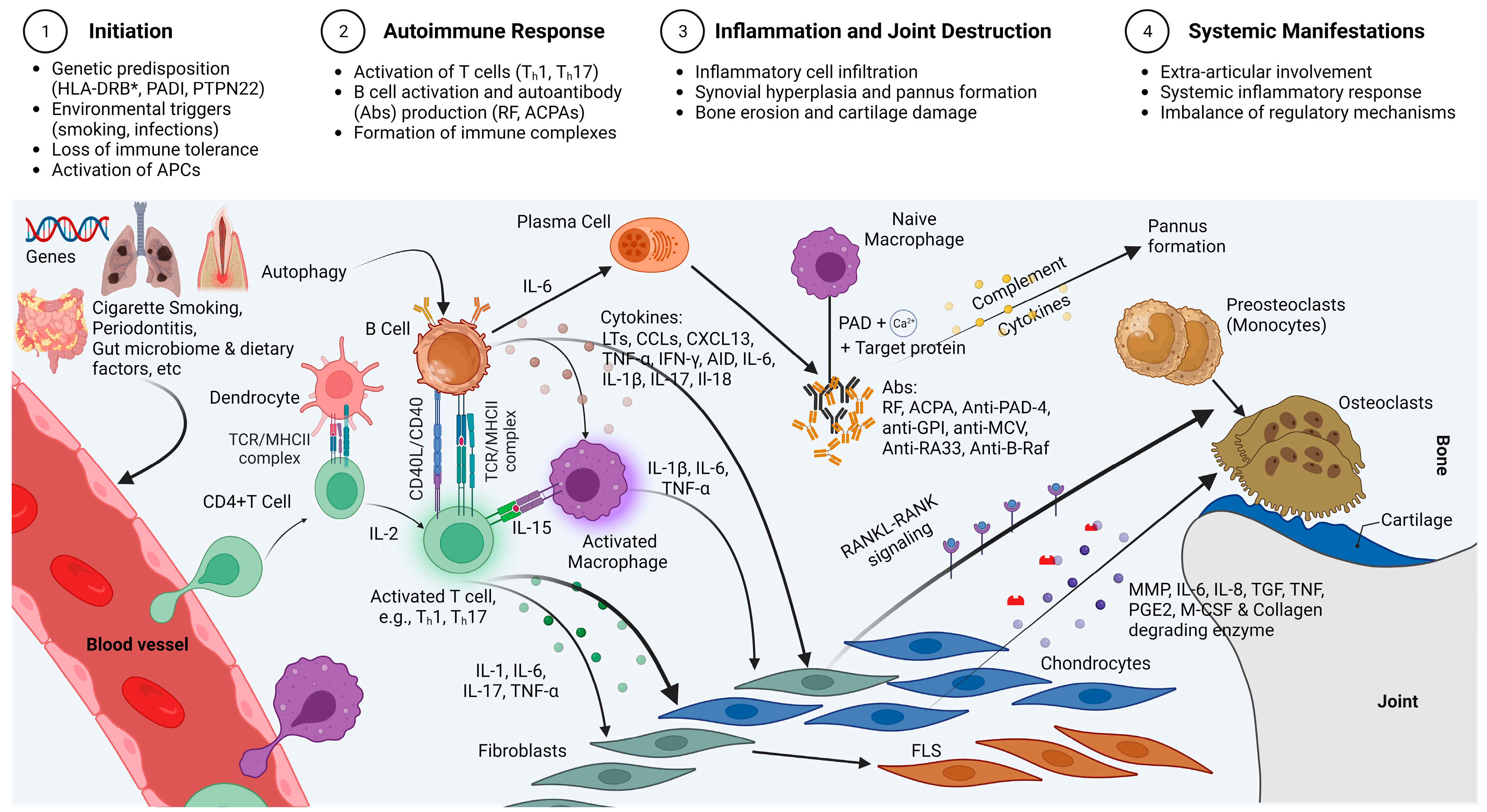
:1. Introduction
2. The Ability of MSCs to Restore Health
2.1. MSC-Based Therapies for RA
2.1.1. Mechanisms Underlying the Effectiveness of MSC-Based Treatments against RA
2.1.2. Merits of MSCs for RA Therapy
2.1.3. MSC-Based Therapy: Preclinical RA Studies
MSC Administration
MSC-Derived EVs
2.1.4. Studies on MSC-Based Therapy for RA in Japan
2.1.5. Studies on MSC-Based Therapy for RA in Other Counties

{kind=link}
| Trial Registration/ Country | Phase | Allo/Auto | Tissue Source | Regimen | Enhancement Method | Enrollment | Start Year | Completed/ Reference |
|---|---|---|---|---|---|---|---|---|
| EudraCT: 2010-021602-37; NCT01663116/Spain | Ib/IIa | Allo. | Adipose tissue | IV weekly infusions of ADMSCs: 1 million/kg (cohort A), 2 million/kg (cohort B), 4 million/kg | None | 53 active refractory RA patients | 2011 | Yes [145] |
| Unidentified/Korea | Pilot | Allo. | Adipose tissue | IV 2 doses of 3 × 108/patient and 4 doses of 2 × 108/patient and 1 dose of 2 × 108/patient; +1 IA dose of 1 × 108/patient or IV single dose of 3.5 × 108/patient + 1 IA dose 1.5 × 108/patient; | None | 4 refractory RA | 2011 | Yes [143] |
| Unidentified/China | Pilot | Allo. | Bone Marrow/Umbical Cord | IV single dose of 1 × 106 cells | None | 4 refractory RA | 2012 | Yes [144] |
| NCT01547091/China | I/II | Allo. | Umbilical Cord | IV 4 × 107 cells/patient | UC-MSC+DMARDS | 172 patients with RA were recruited/64 patients monitored for three years | 2013 | Yes [146,147] |
| NCT01851070/USA | II | Allo. | Bone Marrow | IV single dose of 1 or 2 × 106 mesenchymal precursor cells (MPCs) | None primed precursor cells | 48 refractory RA | 2013 | Yes [140,153] |
| NCT01985464/Panama | I/II | Allo. | Umbilical Cord | Unknown | Unknown | 20 refractory RA | 2013 | Unknown [extracted from ‘www.ClinicalTrials.gov’; accessed on 23 June 2023;] |
| NCT02221258/Korea | I | Allo. | Umbilical Cord Blood | IV single dose of 2.5 × 107, 5 × 107, or 1 × 108 cells of hUCB-MSCs/patient | None | 9 RA patients with moderate disease activity | 2014 | Yes [extracted from ‘www.ClinicalTrials.gov’; accessed on 23 June 2023;] |
| NCT02643823/China | I | Allo. | Umbilical Cord | IV weekly single dose of 2 × 107 hUC-MSC for 1 month | hUC-MSC + DMARDs | 40 refractory RA | 2016 | Unknown |
| ChiCTR-ONC-16008770/China | I | Allo. | Umbilical Cord | IV single dose of 1 × 106 UC-MSCs/kg of body weight | None | 53 refractory RA | 2016 | [148] |
| NCT03067870/Jordan | I | Auto. | Bone Marrow | Unknown | Unknown | 100 | 2016 | Unknown |
| NCT03333681/Iran | I | Auto. | Bone Marrow | IV single dose of 1 to 2 × 106 cells/kg of body weight | I/V hydrocortison (100 mL) and oral dimenhydrinate (10 mL) before MSCs administration + conventional therapy | 15 refractory RA | 2016 | Yes [extracted from ‘www.ClinicalTrials.gov’; accessed on 23 June 2023;] |
| ChiCTR-INR-17012462/China | I/II | Allo. | Umbilical Cord | IV single daose of 1 × 106 cells | UC-MSCs combined with recombinant IFN-γ. | 63 refractory patients | 2017 | Yes [149] |
| NCT03186417/USA | I | Allo. | Umbilical Cord | IV single dose of 2, 4 or 6 × 106 MPCs | None | 10 new onset RA actual enrolment | 2017 | Unknown [153] |
| NCT03798028/China | II/III | Allo. | Umbilical Cord | IV single dose of 1 × 106 cells/kg of body weight of UC-MSCs | +conventional treatment | 250 DMARD-resistant RA patients | 2017 | Unknown [extracted from ‘www.ClinicalTrials.gov’; accessed on 23 June 2023;] |
| NCT03618784/Korea | I/II | Allo. | Umbilical Cord Blood | IV, 5.0 or 10 × 107/patient; 3 doses | FURESTEM-RA Inj.+DMARDs | 33 refractory RA | 2018 | Unknown [extracted from ‘www.ClinicalTrials.gov’; accessed on 23 June 2023;] |
| NCT03691909/USA | I/IIa | Auto. | Adipose tissue | IV single dose of 2 × 108 adMSCs | HB-adMSCs | 15 with active RA | 2018 | Yes [extracted from ‘www.ClinicalTrials.gov’; accessed on 23 June 2023;] |
| NCT03828344/USA | I | Allo. | Umbilical Cord | IV single dose of BX-U001 at 0.75 × 106 cells/kg of body weight (Cohort 1) or 1.5 × 106 cells/kg of body weight (Cohort 2) | Unknown | 16 | 2020 | Not yet known [extracted from ‘www.ClinicalTrials.gov’; accessed on 23 June 2023;] |
| NCT02903212/France | I/II | Auto. | Peripheral blood leukocytes | Unknown | Rendered apoptotic by X-ray irradiation | 22 | 2021 | Not yet known [extracted from www.ClinicalTrials.gov; accessed on 23 June 2023;] |
| NCT04170426/USA | I/IIa | Auto. | Adipose tissue | IV three doses of 2.0–2.86 × 106 cells/kg on day 1, 4 and 7 | Celltex-AdMSCs with unknown enhancement strategy | 54 | 2021 | Not yet known [extracted from www.ClinicalTrials.gov; accessed on 23 June 2023;] |
| NCT04971980/China | I/II | Allo. | Umbilical Cord | IV single infusion of hUC-MSC at 0.5 × 106 cells/kg, or 1.0 × 106 cells/kg, or 1.5 × 106 cells/kg in three cohorts | Unknown | 9 | 2021 | Not yet known [extracted from www.ClinicalTrials.gov; accessed on 23 June 2023;] |
| NCT05003934/USA | I | Allo. | Umbilical Cord | IV single infusion of 100 million cells AlloRx | Unknown | 20 | 2022 | Not yet known [extracted from www.ClinicalTrials.gov; accessed on 23 June 2023;] |
2.1.6. Challenges and Strategies for Enhancing MSC-Based Treatments for RA
3. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hwang, J.J.; Rim, Y.A.; Nam, Y.; Ju, J.H. Recent developments in clinical applications of mesenchymal stem cells in the treatment of rheumatoid arthritis and osteoarthritis. Front. Immunol. 2021, 12, 631291. [Google Scholar] [CrossRef]
- Guo, Q.; Wang, Y.; Xu, D.; Nossent, J.; Pavlos, N.J.; Xu, J. Rheumatoid arthritis: Pathological mechanisms and modern pharmacologic therapies. Bone Res. 2018, 6, 15. [Google Scholar] [CrossRef] [Green Version]
- Ouboussad, L.; Burska, A.N.; Melville, A.; Buch, M.H. Synovial tissue heterogeneity in rheumatoid arthritis and changes with biologic and targeted synthetic therapies to inform stratified therapy. Front. Med. 2019, 6, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silman, A.J.; Pearson, J.E. Epidemiology and genetics of rheumatoid arthritis. Arthritis Res. 2002, 4 (Suppl. 3), S265–S672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firestein, G.S.; McInnes, I.B. Immunopathogenesis of rheumatoid arthritis. Immunity 2017, 46, 183–196. [Google Scholar] [CrossRef] [Green Version]
- Smolen, J.S.; Aletaha, D.; McInnes, I.N. Rheumatoid arthritis. Lancet 2016, 388, 2023–2038. [Google Scholar] [CrossRef] [PubMed]
- Coutant, F.; Miossec, P. Evolving concepts of the pathogenesis of rheumatoid arthritis with focus on the early and late stages. Curr. Opin. Rheumatol. 2020, 32, 57–63. [Google Scholar] [CrossRef]
- Lefevre, S.; Meier, F.M.; Neumann, E.; Muller-Ladner, U. Role of synovial fibroblasts in rheumatoid arthritis. Curr. Pharm. Des. 2015, 21, 130–141. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Shao, S.; Jiao, Z.; Guo, M.; Xu, H.; Wang, S. The Th17/Treg imbalance and cytokine environment in peripheral blood of patients with rheumatoid arthritis. Rheumatol. Int. 2012, 32, 887–893. [Google Scholar] [CrossRef]
- Wu, F.; Gao, J.; Kang, J.; Wang, X.; Niu, Q.; Liu, J.; Zhang, L. B cells in rheumatoid arthritis: Pathogenic mechanisms and treatment prospects. Front. Immunol. 2021, 12, 750753. [Google Scholar] [CrossRef]
- Fukui, S.; Iwamoto, N.; Takatani, A.; Igawa, T.; Shimizu, T.; Umeda, M.; Nishino, A.; Horai, Y.; Hirai, Y.; Koga, T.; et al. M1 and M2 monocytes in rheumatoid arthritis: A contribution of imbalance of M1/M2 monocytes to osteoclastogenesis. Front. Immunol. 2017, 8, 1958. [Google Scholar] [CrossRef] [Green Version]
- Jang, S.; Kwon, E.J.; Lee, J.J. Rheumatoid arthritis: Pathogenic roles of diverse immune cells. Int. J. Mol. Sci. 2022, 23, 905. [Google Scholar] [CrossRef]
- Mateen, S.; Moin, S.; Khan, A.Q.; Zafar, A.; Fatima, N. Increased reactive oxygen species formation and oxidative stress in rheumatoid arthritis. PLoS ONE 2016, 11, e0152925. [Google Scholar] [CrossRef] [Green Version]
- Miao, H.B.; Wang, F.; Lin, S.; Chen, Z. Update on the role of extracellular vesicles in rheumatoid arthritis. Expert Rev. Mol. Med. 2022, 24, e12. [Google Scholar] [CrossRef]
- Kim, H.; Takegahara, N.; Walsh, M.C.; Middleton, S.A.; Yu, J.; Shirakawa, J.; Ueda, J.; Fujihara, Y.; Ikawa, M.; Ishii, M.; et al. IgSF11 regulates osteoclast differentiation through association with the scaffold protein PSD-95. Bone Res. 2020, 8, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirakawa, J.; Takegahara, N.; Kim, H.; Lee, S.H.; Sato, K.; Yamagishi, S.; Choi, Y. Flrt2 is involved in fine-tuning of osteoclast multinucleation. BMB Rep. 2019, 52, 514–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, X.; Hu, Z.; Wang, S.; Yin, K. Netrin-1: An emerging player in inflammatory diseases. Cytokine Growth Factor Rev. 2022, 64, 4656. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Lin, C.; Stavre, Z.; Greenblatt, M.B.; Shim, J.H. Osteoblast-osteoclast communication and bone homeostasis. Cells 2020, 9, 2073. [Google Scholar] [CrossRef]
- Ding, Y.; Wang, L.; Wu, H.; Zhao, Q.; Wu, S. Exosomes derived from synovial fibroblasts under hypoxia aggravate rheumatoid arthritis by regulating Treg/Th17 balance. Exp. Biol. Med. 2020, 245, 1177–1186. [Google Scholar] [CrossRef]
- Zhang, H.G.; Liu, C.; Su, K.; Yu, S.; Zhang, L.; Zhang, S.; Wang, J.; Cao, X.; Grizzle, W.; Kimberly, R.P. A membrane form of TNF-alpha presented by exosomes delays T cell activation-induced cell death. J. Immunol. 2006, 176, 7385–7393, Erratum in J. Immunol. 2006, 177, 2025. [Google Scholar] [CrossRef]
- Burbano, C.; Rojas, M.; Muñoz-Vahos, C.; Vanegas-García, A.; Correa, L.A.; Vásquez, G.; Castaño, D. Extracellular vesicles are associated with the systemic inflammation of patients with seropositive rheumatoid arthritis. Sci. Rep. 2018, 8, 17917. [Google Scholar] [CrossRef] [Green Version]
- McInnes, I.B.; Schett, G. The pathogenesis of rheumatoid arthritis. N. Engl. J. Med. 2011, 365, 2205–2219. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.J.; Chen, Z. Cell-based therapies for rheumatoid arthritis: Opportunities and challenges. Ther. Adv. Musculoskelet Dis. 2022, 14, 1759720X221100294. [Google Scholar] [CrossRef] [PubMed]
- Buzatu, C.; Moots, R.J. Measuring disease activity and response to treatment in rheumatoid arthritis. Expert Rev. Clin. Immunol. 2019, 15, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Drosos, A.A.; Pelechas, E.; Voulgari, P.V. Treatment strategies are more important than drugs in the management of rheumatoid arthritis. Clin. Rheumatol. 2020, 39, 1363–1368. [Google Scholar] [CrossRef]
- Wilke, T.; Mueller, S.; Lee, S.C.; Majer, I.; Heisen, M. Drug survival of second biological DMARD therapy in patients with rheumatoid arthritis: A retrospective non-interventional cohort analysis. BMC Musculoskelet Disord. 2017, 18, 332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, T.H.; Wu, C.S.; Chiou, S.H.; Chang, C.H.; Liao, H.J. Adipose-derived stem cell exosomes as a novel anti-inflammatory agent and the current therapeutic targets for rheumatoid arthritis. Biomedicines 2022, 10, 1725. [Google Scholar] [CrossRef]
- Kerschbaumer, A.; Sepriano, A.; Smolen, J.S.; van der Heijde, D.; Dougados, M.; van Vollenhoven, R.; McInnes, I.B.; Bijlsma, J.W.J.; Burmester, G.R.; de Wit, M.; et al. Efficacy of pharmacological treatment in rheumatoid arthritis: A systematic literature research informing the 2019 update of the EULAR recommendations for management of rheumatoid arthritis. Ann. Rheum. Dis. 2020, 79, 744–759. [Google Scholar] [CrossRef] [Green Version]
- Hetland, M.L.; Haavardsholm, E.A.; Rudin, A.; Nordström, D.; Nurmohamed, M.; Gudbjornsson, B.; Lampa, J.; Hørslev-Petersen, K.; Uhlig, T.; Grondal, G.; et al. Active conventional treatment and three different biological treatments in early rheumatoid arthritis: Phase IV investigator initiated, randomised, observer blinded clinical trial. BMJ 2020, 371, m4328. [Google Scholar] [CrossRef]
- Köhler, B.M.; Günther, J.; Kaudewitz, D.; Lorenz, H.M. Current Therapeutic Options in the Treatment of Rheumatoid Arthritis. J. Clin. Med. 2019, 8, 938. [Google Scholar] [CrossRef] [Green Version]
- Yamaoka, K. Benefit and Risk of Tofacitinib in the Treatment of Rheumatoid Arthritis: A Focus on Herpes Zoster. Drug Saf. 2016, 39, 823–840. [Google Scholar] [CrossRef]
- Cohen, S.B.; Koenig, A.; Wang, L.; Kwok, K.; Mebus, C.A.; Riese, R.; Fleischmann, R. Efficacy and safety of tofacitinib in US and non-US rheumatoid arthritis patients: Pooled analyses of phase II and III. Clin. Exp. Rheumatol. 2016, 34, 32–36. [Google Scholar] [PubMed]
- Keystone, E.C.; Taylor, P.C.; Drescher, E.; Schlichting, D.E.; Beattie, S.D.; Berclaz, P.Y.; Lee, C.H.; Fidelus-Gort, R.K.; Luchi, M.E.; Rooney, T.P.; et al. Safety and efficacy of baricitinib at 24 weeks in patients with rheumatoid arthritis who have had an inadequate response to methotrexate. Ann. Rheum. Dis. 2015, 74, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Smolen, J.S.; Genovese, M.C.; Takeuchi, T.; Hyslop, D.L.; Macias, W.L.; Rooney, T.; Chen, L.; Dickson, C.L.; Riddle Camp, J.; Cardillo, T.E.; et al. Safety Profile of Baricitinib in Patients with Active Rheumatoid Arthritis with over 2 Years Median Time in Treatment. J. Rheumatol. 2019, 46, 7–18. [Google Scholar] [CrossRef] [Green Version]
- Westhovens, R. Clinical efficacy of new JAK inhibitors under development. Just more of the same? Rheumatology 2019, 58 (Suppl. 1), i27–i33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serhal, L.; Edwards, C.J. Upadacitinib for the treatment of rheumatoid arthritis. Expert Rev. Clin. Immunol. 2018, 15, 13–25. [Google Scholar] [CrossRef]
- Singh, J.A.; Saag, K.G.; Bridges, S.L., Jr.; Akl, E.A.; Bannuru, R.R.; Sullivan, M.C.; Vaysbrot, E.; McNaughton, C.; Osani, M.; Shmerling, R.H.; et al. American College of Rheumatology Guideline for the treatment of rheumatoid arthritis. Arthritis Rheumatol. 2016, 68, 1–26. [Google Scholar] [CrossRef]
- Smolen, J.S.; Landewé, R.B.M.; Bijlsma, J.W.J.; Burmester, G.R.; Dougados, M.; Kerschbaumer, A.; McInnes, I.B.; Sepriano, A.; van Vollenhoven, R.F.; de Wit, M.; et al. EULAR recommendations for the management of rheumatoid arthritis with synthetic and biological disease-modifying antirheumatic drugs: 2019 update. Ann. Rheum. Dis. 2020, 79, 685–699. [Google Scholar] [CrossRef] [Green Version]
- Ntege, E.H.; Sunami, H.; Shimizu, Y. Advances in regenerative therapy: A review of the literature and future directions. Regen. Ther. 2020, 14, 136–153. [Google Scholar] [CrossRef]
- Jiang, Q.; Yang, G.; Liu, Q.; Wang, S.; Cui, D. Function and role of regulatory T cells in rheumatoid arthritis. Front. Immunol. 2021, 12, 626193. [Google Scholar] [CrossRef] [PubMed]
- Orvain, C.; Boulch, M.; Bousso, P.; Allanore, Y.; Avouac, J. Is there a place for chimeric antigen receptor-T cells in the treatment of chronic autoimmune rheumatic diseases? Arthritis Rheumatol. 2021, 73, 1954–1965. [Google Scholar] [CrossRef]
- Shimizu, Y.; Ntege, E.H.; Sunami, H. Adipose tissue-derived regenerative cell-based therapies: Current optimisation strategies for effective treatment in aesthetic surgery. In Handbook of Stem Cell Therapy; Haider, K.H., Ed.; Springer: Singapore, 2022. [Google Scholar] [CrossRef]
- Parekkadan, B.; Milwid, J.M. Mesenchymal stem cells as therapeutics. Annu. Rev. Biomed. Eng. 2010, 12, 87–117. [Google Scholar] [CrossRef] [Green Version]
- Squillaro, T.; Peluso, G.; Galderisi, U. Clinical trials with mesenchymal stem cells: An update. Cell Transpl. 2016, 25, 829–848. [Google Scholar] [CrossRef] [Green Version]
- Lukomska, B.; Stanaszek, L.; Zuba-Surma, E.; Legosz, P.; Sarzynska, S.; Drela, K. Challenges and controversies in human mesenchymal stem cell therapy. Stem Cell Int. 2019, 2019, 9628536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ankrum, J.A.; Ong, J.F.; Karp, J.M. Mesenchymal stem cells: Immune evasive, not immune privileged. Nat. Biotechnol. 2014, 32, 252–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, X.L.; Zhang, Y.; Li, X.; Fu, Q.L. Mechanisms underlying the protective effects of mesenchymal stem cell-based therapy. Cell Mol. Life Sci. 2020, 77, 2771–2794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haynesworth, S.E.; Goshima, J.; Goldberg, V.M.; Caplan, A.I. Characterization of cells with osteogenic potential from human marrow. Bone 1992, 13, 81–88. [Google Scholar] [CrossRef]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [Green Version]
- Mohamed-Ahmed, S.; Fristad, I.; Lie, S.A.; Suliman, S.; Mustafa, K.; Vindenes, H.; Idris, S.B. Adipose-derived and bone marrow mesenchymal stem cells: A donor-matched comparison. Stem Cell Res. Ther. 2018, 9, 168. [Google Scholar] [CrossRef]
- Ghoryani, M.; Shariati-Sarabi, Z.; Tavakkol-Afshari, J.; Mohammadi, M. The sufficient immunoregulatory effect of autologous bone marrow-derived mesenchymal stem cell transplantation on regulatory T cells in patients with refractory rheumatoid arthritis. J. Immunol. Res. 2020, 2020, 3562753. [Google Scholar] [CrossRef]
- Ghoryani, M.; Shariati-Sarabi, Z.; Tavakkol-Afshari, J.; Ghasemi, A.; Poursamimi, J.; Mohammadi, M. Amelioration of clinical symptoms of patients with refractory rheumatoid arthritis following treatment with autologous bone marrow-derived mesenchymal stem cells: A successful clinical trial in Iran. Biomed. Pharmacother. 2019, 109, 1834–1840. [Google Scholar] [CrossRef]
- Cosenza, S.; Toupet, K.; Maumus, M.; Luz-Crawford, P.; Blanc-Brude, O.; Jorgensen, C.; Noël, D. Mesenchymal stem cells-derived exosomes are more immunosuppressive than microparticles in inflammatory arthritis. Theranostics 2018, 8, 1399–1410. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Rong, Y.; Luo, C.; Cui, W. Bone marrow mesenchymal stem cell-derived exosomes prevent osteoarthritis by regulating synovial macrophage polarization. Aging 2020, 12, 25138–25152. [Google Scholar] [CrossRef] [PubMed]
- He, L.; He, T.; Xing, J.; Zhou, Q.; Fan, L.; Liu, C.; Chen, Y.; Wu, D.; Tian, Z.; Liu, B.; et al. Bone marrow mesenchymal stem cell-derived exosomes protect cartilage damage and relieve knee osteoarthritis pain in a rat model of osteoarthritis. Stem Cell Res. Ther. 2020, 11, 276. [Google Scholar] [CrossRef] [PubMed]
- Nagamura-Inoue, T.; He, H. Umbilical cord-derived mesenchymal stem cells: Their advantages and potential clinical utility. World J. Stem Cells 2014, 6, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.Y.; Mou, X.Z.; Du, X.C.; Xiang, C. Comparative analysis of biological characteristics of adult mesenchymal stem cells with different tissue origins. Asian Pac. J. Trop. Med. 2015, 8, 739–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Jo, C.H.; Kim, H.R.; Hwang, Y.I. Comparison of immunological characteristics of mesenchymal stem cells from the periodontal ligament, umbilical cord, and adipose tissue. Stem Cells Int. 2018, 2018, 8429042. [Google Scholar] [CrossRef]
- Kim, Y.J.; Seo, D.H.; Lee, S.H.; Lee, S.H.; An, G.H.; Ahn, H.J.; Kwon, D.; Seo, K.W.; Kang, K.S. Conditioned media from human umbilical cord blood-derived mesenchymal stem cells stimulate rejuvenation function in human skin. Biochem. Biophys. Rep. 2018, 16, 96–102. [Google Scholar] [CrossRef]
- Lu, L.L.; Liu, Y.J.; Yang, S.G.; Zhao, Q.J.; Wang, X.; Gong, W.; Han, Z.B.; Xu, Z.S.; Lu, Y.X.; Liu, D.; et al. Isolation and characterization of human umbilical cord mesenchymal stem cells with hematopoiesis-supportive function and other potentials. Haematologica 2006, 91, 1017–1026. [Google Scholar] [PubMed]
- Stanko, P.; Kaiserova, K.; Altanerova, V.; Altaner, C. Comparison of human mesenchymal stem cells derived from dental pulp, bone marrow, adipose tissue, and umbilical cord tissue by gene expression. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc Czech Repub. 2014, 158, 373–377. [Google Scholar] [CrossRef] [Green Version]
- Schmelzer, E.; McKeel, D.T.; Gerlach, J.C. Characterization of human mesenchymal stem cells from different tissues and their membrane encasement for prospective transplantation therapies. Biomed. Res. Int. 2019, 2019, 6376271. [Google Scholar] [CrossRef]
- Um, S.; Ha, J.; Choi, S.J.; Oh, W.; Jin, H.J. Prospects for the therapeutic development of umbilical cord blood-derived mesenchymal stem cells. World J. Stem Cells 2020, 12, 1511–1528. [Google Scholar] [CrossRef]
- Jin, H.J.; Bae, Y.K.; Kim, M.; Kwon, S.J.; Jeon, H.B.; Choi, S.J.; Kim, S.W.; Yang, Y.S.; Oh, W.; Chang, J.W. Comparative analysis of human mesenchymal stem cells from bone marrow, adipose tissue, and umbilical cord blood as sources of cell therapy. Int. J. Mol. Sci. 2013, 14, 17986–18001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, F.; Lu, M.; Cheung, K.M.; Leung, V.Y.; Zhou, G. Intrinsic properties of mesemchymal stem cells from human bone marrow, umbilical cord and umbilical cord blood comparing the different sources of MSC. Curr. Stem Cell Res. Ther. 2012, 7, 389–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Salazar, M.; Gonzalez-Galofre, Z.N.; Casamitjana, J.; Crisan, M.; James, A.W.; Péault, B. Five Decades Later, Are Mesenchymal Stem Cells Still Relevant? Front. Bioeng. Biotechnol. 2020, 8, 148. [Google Scholar] [CrossRef] [Green Version]
- Nepali, S.; Park, M.; Lew, H.; Kim, O. Comparative analysis of human adipose-derived mesenchymal stem cells from orbital and abdominal fat. Stem Cells Int. 2018, 2018, 3932615. [Google Scholar] [CrossRef] [PubMed]
- Heidari, N.; Abbasi-Kenarsari, H.; Namaki, S.; Baghaei, K.; Zali, M.R.; Ghaffari Khaligh, S.; Hashemi, S.M. Adipose-derived mesenchymal stem cell-secreted exosome alleviates dextran sulfate sodium-induced acute colitis by Treg cell induction and inflammatory cytokine reduction. J. Cell Physiol. 2021, 236, 5906–5920. [Google Scholar] [CrossRef]
- Blazquez, R.; Sanchez-Margallo, F.M.; de la Rosa, O.; Dalemans, W.; Alvarez, V.; Tarazona, R.; Casado, J.G. Immunomodulatory potential of human adipose mesenchymal stem cells derived exosomes on in vitro stimulated T cells. Front. Immunol. 2014, 5, 556. [Google Scholar] [CrossRef] [Green Version]
- Serejo, T.R.T.; Silva-Carvalho, A.É.; Braga, L.D.C.F.; Neves, F.A.R.; Pereira, R.W.; Carvalho, J.L.; Saldanha-Araujo, F. Assessment of the immunosuppressive potential of INF-γ licensed adipose mesenchymal stem cells, their secretome and extracellular vesicles. Cells 2019, 8, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.G.; He, Z.Y.; Liang, S.; Yang, Q.; Cheng, P.; Chen, A.M. Comprehensive proteomic analysis of exosomes derived from human bone marrow, adipose tissue, and umbilical cord mesenchymal stem cells. Stem Cell Res. Ther. 2020, 11, 511. [Google Scholar] [CrossRef]
- Li, Q.; Yu, H.; Sun, M.; Yang, P.; Hu, X.; Ao, Y.; Cheng, J. The tissue origin effect of extracellular vesicles on cartilage and bone regeneration. Acta Biomater. 2021, 125, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Woo, C.H.; Kim, H.K.; Jung, G.Y.; Jung, Y.J.; Lee, K.S.; Yun, Y.E.; Han, J.; Lee, J.; Kim, W.S.; Choi, J.S.; et al. Small extracellular vesicles from human adipose-derived stem cells attenuate cartilage degeneration. J. Extracell. Vesicles 2020, 9, 1735249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Bari, C.; Dell’Accio, F.; Tylzanowski, P.; Luyten, F.P. Multipotent mesenchymal stem cells from adult human synovial membrane. Arthritis Rheum. 2001, 44, 1928–1942. [Google Scholar] [CrossRef] [PubMed]
- Murata, Y.; Uchida, S.; Utsunomiya, H.; Hatakeyama, A.; Nakashima, H.; Chang, A.; Sekiya, I.; Sakai, A. Synovial mesenchymal stem cells derived from the cotyloid fossa synovium have higher self-renewal and differentiation potential than those from the paralabral synovium in the hip joint. Am. J. Sport. Med. 2018, 46, 2942–2953. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Gao, J.; Mi, L.; Zhang, G.; Zhang, L.; Zhang, N.; Huo, R.; Hu, J.; Xu, K. Synovial membrane mesenchymal stem cells: Past life, current situation, and application in bone and joint diseases. Stem Cell Res. Ther. 2020, 11, 381. [Google Scholar] [CrossRef] [PubMed]
- Kubosch, E.J.; Lang, G.; Furst, D.; Kubosch, D.; Izadpanah, K.; Rolauffs, B.; Sudkamp, N.P.; Schmal, H. The potential for synovium-derived stem cells in cartilage repair. Curr. Stem Cell Res. Ther. 2018, 13, 174–184. [Google Scholar] [CrossRef]
- Mochizuki, T.; Muneta, T.; Sakaguchi, Y.; Nimura, A.; Yokoyama, A.; Koga, H.; Sekiya, I. Higher chondrogenic potential of fibrous synovium- and adipose synovium-derived cells compared with subcutaneous fat-derived cells: Distinguishing properties of mesenchymal stem cells in humans. Arthritis Rheum. 2006, 54, 843–853. [Google Scholar] [CrossRef]
- Ogata, Y.; Mabuchi, Y.; Yoshida, M.; Suto, E.G.; Suzuki, N.; Muneta, T.; Sekiya, I.; Akazawa, C. Purified human synovium mesenchymal stem cells as a good resource for cartilage regeneration. PLoS ONE 2015, 10, e0129096. [Google Scholar] [CrossRef]
- Sakaguchi, Y.; Sekiya, I.; Yagishita, K.; Muneta, T. Comparison of human stem cells derived from various mesenchymal tissues: Superiority of synovium as a cell source. Arthritis Rheum. 2005, 52, 2521–2529. [Google Scholar] [CrossRef]
- Nikfarjam, S.; Rezaie, J.; Zolbanin, N.M.; Jafari, R. Mesenchymal stem cell derived-exosomes: A modern approach in translational medicine. J. Transl. Med. 2020, 18, 449. [Google Scholar] [CrossRef]
- Sakai, D.; Schol, J.; Foldager, C.B.; Sato, M.; Watanabe, M. Regenerative technologies to bed side: Evolving the regulatory framework. J. Orthop. Translat. 2017, 9, 1–7, Erratum in J. Orthop. Transl. 2018, 12, 93. [Google Scholar] [CrossRef]
- Spaggiari, G.M.; Capobianco, A.; Abdelrazik, H.; Becchetti, F.; Mingari, M.C.; Moretta, L. Mesenchymal stem cells inhibit natural killer-cell proliferation, cytotoxicity, and cytokine production: Role of indoleamine 2,3-dioxygenase and prostaglandin E2. Blood 2008, 111, 1327–1333. [Google Scholar] [CrossRef] [PubMed]
- Nasef, A.; Chapel, A.; Mazurier, C.; Bouchet, S.; Lopez, M.; Mathieu, N.; Sensebé, L.; Zhang, Y.; Gorin, N.C.; Thierry, D.; et al. Identification of IL-10 and TGF-β transcripts involved in the inhibition of T-lymphocyte proliferation during cell contact with human mesenchymal stem cells. Gene Expr. 2007, 13, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Wu, X.R.; Liu, H.S.; Liu, X.-H.; Liu, G.H.; Zheng, X.B.; Hu, T.; Liang, Z.X.; He, X.W.; Wu, X.J.; et al. Immunomodulatory effect of urine-derived stem Cells on inflammatory bowel diseases via downregulating Th1/Th17 immune responses in a PGE2-dependent manner. J. Crohn’s Colitis 2019, 14, 654–668. [Google Scholar] [CrossRef] [PubMed]
- López-Santalla, M.; Mancheño-Corvo, P.; Menta, R.; Lopez-Belmonte, J.; Delarosa, O.; Bueren, J.A.; Dalemans, W.; Lombardo, E.; Garín, M. Human adipose-derived mesenchymal stem cells modulate experimental autoimmune arthritis by modifying early adaptive T cell responses. Stem Cells 2015, 33, 3493–3503. [Google Scholar] [CrossRef]
- López-García, L.; Castro-Manrreza, M.E. TNF-α and IFN-γ participate in improving the immunoregulatory capacity of mesenchymal stem/stromal cells: Importance of cell-cell contact and extracellular vesicles. Int. J. Mol. Sci. 2021, 22, 9531. [Google Scholar] [CrossRef]
- Szydlak, R. Biological, chemical and mechanical factors regulating migration and homing of mesenchymal stem cells. World J. Stem Cells 2021, 13, 619–631. [Google Scholar] [CrossRef]
- Zachar, L.; Bačenková, D.; Rosocha, J. Activation, homing, and role of the mesenchymal stem cells in the inflammatory environment. J. Inflam. Res. 2016, 9, 231–240. [Google Scholar] [CrossRef] [Green Version]
- Le Blanc, K.; Mougiakakos, D. Multipotent mesenchymal stromal cells and the innate immune system. Nat. Rev. Immunol. 2012, 12, 383–396. [Google Scholar] [CrossRef]
- Jiang, W.; Xu, J. Immune modulation by mesenchymal stem cells. Cell Prolif. 2020, 53, e12712. [Google Scholar] [CrossRef] [Green Version]
- Bačenková, D.; Trebuňová, M.; Morochovič, R.; Dosedla, E.; Findrik Balogová, A.; Gašparová, P.; Živčák, J. Interaction between mesenchymal stem cells and the immune system in rheumatoid arthritis. Pharmaceuticals 2022, 15, 941. [Google Scholar] [CrossRef] [PubMed]
- Tatara, R.; Ozaki, K.; Kikuchi, Y.; Hatanaka, K.; Oh, I.; Meguro, A.; Matsu, H.; Sato, K.; Ozawa, K. Mesenchymal stromal cells inhibit Th17 but not regulatory T-cell differentiation. Cytotherapy 2011, 13, 686–694. [Google Scholar] [CrossRef]
- Galland, S.; Stamenkovic, I. Mesenchymal stromal cells in cancer: A review of their immunomodulatory functions and dual effects on tumor progression. J. Pathol. 2020, 250, 555–572. [Google Scholar] [CrossRef] [Green Version]
- Ren, G.; Zhao, X.; Zhang, L.; Zhang, J.; L’Huillier, A.; Ling, W.; Roberts, A.I.; Le, A.D.; Shi, S.; Shao, C.; et al. Inflammatory cytokine-induced intercellular adhesion molecule-1 and vascular cell adhesion molecule-1 in mesenchymal stem cells are critical for immunosuppression. J. Immunol. 2010, 184, 2321–2328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najar, M.; Raicevic, G.; Fayyad-Kazan, H.; De Bruyn, C.; Bron, D.; Toungouz, M.; Lagneaux, L. Impact of different mesenchymal stromal cell types on T-cell activation, proliferation and migration. Int. Immunopharmacol. 2013, 15, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Luque-Campos, N.; Contreras-López, R.A.; Jose Paredes-Martínez, M.; Torres, M.J.; Bahraoui, S.; Wei, M.; Espinoza, F.; Djouad, F.; Elizondo-Vega, R.J.; Luz-Crawford, P. Mesenchymal stem cells improve rheumatoid arthritis progression by controlling memory T cell response. Front. Immunol. 2019, 10, 798. [Google Scholar] [CrossRef] [PubMed]
- Berthelot, J.M.; Le Goff, B.; Maugars, Y. Bone marrow mesenchymal stem cells in rheumatoid arthritis, spondyloarthritis, and ankylosing spondylitis: Problems rather than solutions? Arthritis Res. Ther. 2019, 21, 239. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Zhou, X.; Wang, X.; Cheng, W.; Hu, X.; Wang, Y.; Luo, B.; Huang, W.; Gu, J. miR-34a in extracellular vesicles from bone marrow mesenchymal stem cells reduces rheumatoid arthritis inflammation via the cyclin I/ATM/ATR/p53 axis. J. Cell Mol. Med. 2021, 25, 1896–1910. [Google Scholar] [CrossRef]
- Farinazzo, A.; Angiari, S.; Turano, E.; Bistaffa, E.; Dusi, S.; Ruggieri, S.; Bonafede, R.; Mariotti, R.; Constantin, G.; Bonetti, B. Nanovesicles from adipose-derived mesenchymal stem cells inhibit T lymphocyte trafficking and ameliorate chronic experimental autoimmune encephalomyelitis. Sci. Rep. 2018, 8, 7473. [Google Scholar] [CrossRef]
- Tian, J.; Zhu, Q.; Zhang, Y.; Bian, Q.; Hong, Y.; Shen, Z.; Xu, H.; Rui, K.; Yin, K.; Wang, S. Olfactory ecto-mesenchymal stem cell-derived exosomes ameliorate experimental colitis via modulating Th1/Th17 and Treg cell responses. Front. Immunol. 2020, 11, 598322. [Google Scholar] [CrossRef]
- Cocucci, E.; Meldolesi, J. Ectosomes and exosomes: Shedding the confusion between extracellular vesicles. Trends Cell Biol. 2015, 25, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, N.; Yerneni, S.S.; Azambuja, J.H.; Gillespie, D.G.; Menshikova, E.V.; Jackson, E.K.; Whiteside, T.L. Tumour-derived exosomes promote angiogenesis via adenosine A2B receptor signalling. Angiogenesis 2020, 23, 599–610. [Google Scholar] [CrossRef]
- Lindenbergh, M.F.S.; Wubbolts, R.; Borg, E.G.F.; van ′t Veld, E.M.; Boes, M.; Stoorvogel, W. Dendritic cells release exosomes together with a phagocytosed pathogen; potential implications for the role of exosomes in antigen presentation. J. Extracell. Vesicles 2020, 9, 1798606. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Ma, D.; Zhang, G.; Gao, J.; Su, Y.; Liu, S.; Liu, Y.; Han, J.; Tian, M.; Wei, C.; et al. Human umbilical cord mesenchymal stem cell-derived small extracellular vesicles ameliorate collagen-induced arthritis via immunomodulatory T lymphocytes. Mol. Immunol. 2021, 135, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Xu, K.; Zhang, G.; Liu, Y.; Gao, J.; Tian, M.; Wei, C.; Li, J.; Zhang, L. Immunomodulatory effect of human umbilical cord mesenchymal stem cells on T lymphocytes in rheumatoid arthritis. Int. Immunopharmacol. 2019, 74, 105687. [Google Scholar] [CrossRef]
- Di Trapani, M.; Bassi, G.; Midolo, M.; Gatti, A.; Kamga, P.T.; Cassaro, A.; Carusone, R.; Adamo, A.; Krampera, M. Differential and transferable modulatory effects of mesenchymal stromal cell-derived extracellular vesicles on T, B and NK cell functions. Sci. Rep. 2016, 6, 24120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, K.S.; Kang, S.A.; Kim, S.D.; Mun, S.J.; Yu, H.S.; Roh, H.J. Dendritic cells and M2 macrophage play an important role in suppression of Th2-mediated inflammation by adipose stem cells-derived extracellular vesicles. Stem Cell Res. 2019, 39, 101500. [Google Scholar] [CrossRef]
- Khare, D.; Or, R.; Resnick, I.; Barkatz, C.; Almogi-Hazan, O.; Avni, B. Mesenchymal stromal cell-derived exosomes affect mRNA expression and function of B-lymphocytes. Front. Immunol. 2018, 9, 3053. [Google Scholar] [CrossRef] [Green Version]
- An, L.; Chu, T.; Wang, L.; An, S.; Li, Y.; Hao, H.; Zhang, Z.; Yue, H. Frequent injections of high-dose human umbilical cord mesenchymal stem cells slightly aggravate arthritis and skeletal muscle cachexia in collagen-induced arthritic mice. Exp. Ther. Med. 2021, 22, 1272. [Google Scholar] [CrossRef]
- Zhai, Q.; Dong, J.; Zhang, X.; He, X.; Fei, D.; Jin, Y.; Li, B.; Jin, F. Mesenchymal stem cells enhance therapeutic effect and prevent adverse gastrointestinal reaction of methotrexate treatment in collagen-induced arthritis. Stem Cells Int. 2021, 2021, 8850820. [Google Scholar] [CrossRef]
- Nam, Y.; Jung, S.M.; Rim, Y.A.; Jung, H.; Lee, K.; Park, N.; Kim, J.; Jang, Y.; Park, Y.B.; Park, S.H.; et al. Intraperitoneal infusion of mesenchymal stem cell attenuates severity of collagen antibody induced arthritis. PLoS ONE 2018, 13, e0198740. [Google Scholar] [CrossRef] [PubMed]
- Shu, J.; Pan, L.; Huang, X.; Wang, P.; Li, H.; He, X.; Cai, Z. Transplantation of human amnion mesenchymal cells attenuates the disease development in rats with collagen-induced arthritis. Clin. Exp. Rheumatol. 2015, 33, 484–490. [Google Scholar] [PubMed]
- Li, X.; Lu, C.; Fan, D.; Lu, X.; Xia, Y.; Zhao, H.; Xu, H.; Zhu, Y.; Li, J.; Liu, H.; et al. Human umbilical mesenchymal stem cells display therapeutic potential in rheumatoid arthritis by regulating interactions between immunity and gut microbiota via the aryl hydrocarbon receptor. Front. Cell Dev. Biol. 2020, 8, 131. [Google Scholar] [CrossRef] [PubMed]
- Abdelmawgoud, H.; Saleh, A. Anti-inflammatory and antioxidant effects of mesenchymal and hematopoietic stem cells in a rheumatoid arthritis rat model. Adv. Clin. Exp. Med. 2018, 27, 873–880. [Google Scholar] [CrossRef]
- Haikal, S.M.; Abdeltawab, N.F.; Rashed, L.A.; Abd El-Galil, T.I.; Elmalt, H.A.; Amin, M.A. Combination therapy of mesenchymal stromal cells and interleukin-4 attenuates rheumatoid arthritis in a collagen-induced murine model. Cells 2019, 8, 823. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Li, X.; Zhang, Z.; Zhou, M.; Sun, Y.; Su, D.; Feng, X.; Gao, X.; Shi, S.; Chen, W.; et al. Allogeneic mesenchymal stem cells inhibited T follicular helper cell generation in rheumatoid arthritis. Sci. Rep. 2015, 5, 12777. [Google Scholar] [CrossRef] [Green Version]
- Roudsari, P.P.; Alavi-Moghadam, S.; Rezaei-Tavirani, M.; Goodarzi, P.; Tayanloo-Beik, A.; Sayahpour, F.A.; Larijani, B.; Arjmand, B. The outcome of stem cell-based therapies on the immune responses in rheumatoid arthritis. Adv. Exp. Med. Biol. 2021, 1326, 159–186. [Google Scholar] [CrossRef]
- Gao, J.; Zhang, G.; Xu, K.; Ma, D.; Ren, L.; Fan, J.; Hou, J.; Han, J.; Zhang, L. Bone marrow mesenchymal stem cells improve bone erosion in collagen-induced arthritis by inhibiting osteoclasia-related factors and differentiating into chondrocytes. Stem Cell Res. Ther. 2020, 11, 171. [Google Scholar] [CrossRef]
- Zheng, J.; Zhu, L.; Iok In, I.; Chen, Y.; Jia, N.; Zhu, W. Bone marrow-derived mesenchymal stem cells-secreted exosomal microRNA-192-5p delays inflammatory response in rheumatoid arthritis. Int. Immunopharmacol. 2020, 78, 105985. [Google Scholar] [CrossRef]
- Meng, Q.; Qiu, B. Exosomal microRNA-320a derived from mesenchymal stem cells regulates rheumatoid arthritis fibroblast-like synoviocyte activation by suppressing CXCL9 expression. Front. Physiol. 2020, 11, 441. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, H.; Xia, Y.; Yan, F.; Lu, Y. Therapeutic potential of mesenchymal cell-derived miRNA-150-5p-expressing exosomes in rheumatoid arthritis mediated by the modulation of MMP14 and VEGF. J. Immunol. 2018, 201, 2472–2482. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.; Liu, Y.; Ma, C.; Guan, C.; Ma, X.; Meng, S. Mesenchymal stem cell-originated exosomal lncRNA HAND2-AS1 impairs rheumatoid arthritis fibroblast-like synoviocyte activation through miR-143-3p/TNFAIP3/NF-κB pathway. J. Orthop. Surg. Res. 2021, 16, 116. [Google Scholar] [CrossRef]
- Tsujimaru, K.; Takanashi, M.; Sudo, K.; Ishikawa, A.; Mineo, S.; Ueda, S.; Kumagai, K.; Kuroda, M. Extracellular microvesicles that originated adipose tissue derived mesenchymal stem cells have the potential ability to improve rheumatoid arthritis on mice. Regen. Ther. 2020, 15, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Fathollahi, A.; Hashemi, S.M.; Haji Molla Hoseini, M.; Tavakoli, S.; Farahani, E.; Yeganeh, F. Intranasal administration of small extracellular vesicles derived from mesenchymal stem cells ameliorated the experimental autoimmune encephalomyelitis. Int. Immunopharmacol. 2021, 90, 107207. [Google Scholar] [CrossRef] [PubMed]
- Jafarinia, M.; Alsahebfosoul, F.; Salehi, H.; Eskandari, N.; Azimzadeh, M.; Mahmoodi, M.; Asgary, S.; Ganjalikhani Hakemi, M. Therapeutic effects of extracellular vesicles from human adipose-derived mesenchymal stem cells on chronic experimental autoimmune encephalomyelitis. J. Cell Physiol. 2020, 235, 8779–8790. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.S.; Choi, Y.; Kim, H.O. Adipose-derived mesenchymal stem cells promote M2 macrophage phenotype through exosomes. Stem Cells Int. 2019, 2019, 7921760. [Google Scholar] [CrossRef] [PubMed]
- Domenis, R.; Cifù, A.; Quaglia, S.; Pistis, C.; Moretti, M.; Vicario, A.; Parodi, P.C.; Fabris, M.; Niazi, K.R.; Soon-Shiong, P.; et al. Pro inflammatory stimuli enhance the immunosuppressive functions of adipose mesenchymal stem cells-derived exosomes. Sci. Rep. 2018, 8, 13325. [Google Scholar] [CrossRef]
- Bolandi, Z.; Mokhberian, N.; Eftekhary, M.; Sharifi, K.; Soudi, S.; Ghanbarian, H.; Hashemi, S.M. Adipose derived mesenchymal stem cell exosomes loaded with miR-10a promote the differentiation of Th17 and Treg from naive CD4+ T cell. Life Sci. 2020, 259, 118218. [Google Scholar] [CrossRef]
- Cao, D.; van Vollenhoven, R.; Klareskog, L.; Trollmo, C.; Malmström, V. CD25brightCD4+ regulatory T cells are enriched in inflamed joints of patients with chronic rheumatic disease. Arthritis Res. Ther. 2004, 6, 335–346. [Google Scholar] [CrossRef] [Green Version]
- Flores-Borja, F.; Jury, E.C.; Mauri, C.; Ehrenstein, M.R. Defects in CTLA-4 are associated with abnormal regulatory T cell function in rheumatoid arthritis. Proc. Natl. Acad. Sci. USA 2008, 105, 19396–19401. [Google Scholar] [CrossRef]
- Nie, H.; Zheng, Y.; Li, R.; Guo, T.B.; He, D.; Fang, L.; Liu, X.; Xiao, L.; Chen, X.; Wan, B.; et al. Phosphorylation of FOXP3 controls regulatory T cell function and is inhibited by TNF-α in rheumatoid arthritis. Nat. Med. 2013, 19, 322–328. [Google Scholar] [CrossRef]
- Jiang, S.; Tian, G.; Yang, Z.; Gao, X.; Wang, F.; Li, J.; Tian, Z.; Huang, B.; Wei, F.; Sang, X.; et al. Enhancement of acellular cartilage matrix scaffold by Wharton’s jelly mesenchymal stem cell-derived exosomes to promote osteochondral regeneration. Bioact. Mater. 2021, 6, 2711–2728. [Google Scholar] [CrossRef] [PubMed]
- Rui, K.; Zhang, Z.; Tian, J.; Lin, X.; Wang, X.; Ma, J.; Tang, X.; Xu, H.; Lu, L.; Wang, S. Olfactory ecto-mesenchymal stem cells possess immunoregulatory function and suppress autoimmune arthritis. Cell Mol. Immunol. 2016, 13, 401–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Ji, Q.; Meng, C.; Liu, H.; Fan, C.; Lipkind, S.; Wang, Z.; Xu, Q. Role of gingival mesenchymal stem cell exosomes in macrophage polarization under inflammatory conditions. Int. Immunopharmacol. 2020, 81, 106030. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Z.; Shi, B.; Li, Y.; Wang, R.; Sun, J.; Hu, Y.; Yuan, C.; Xu, Q. Effect of gingival mesenchymal stem cell-derived exosomes on inflammatory macrophages in a high-lipid microenvironment. Int. Immunopharmacol. 2021, 94, 107455. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Wang, Y.; Zhao, B.; Niu, X.; Hu, B.; Li, Q.; Zhang, J.; Ding, J.; Chen, Y.; Wang, Y. Comparison of exosomes secreted by induced pluripotent stem cell-derived mesenchymal stem cells and synovial membrane-derived mesenchymal stem cells for the treatment of osteoarthritis. Stem Cell Res. Ther. 2017, 8, 64. [Google Scholar] [CrossRef] [Green Version]
- Zavatti, M.; Beretti, F.; Casciaro, F.; Bertucci, E.; Maraldi, T. Comparison of the therapeutic effect of amniotic fluid stem cells and their exosomes on monoiodoacetate-induced animal model of osteoarthritis. Biofactors 2020, 46, 106–117. [Google Scholar] [CrossRef]
- Gatti, M.; Beretti, F.; Zavatti, M.; Bertucci, E.; Ribeiro Luz, S.; Palumbo, C.; Maraldi, T. Amniotic fluid stem cell-derived extracellular vesicles counteract steroid-induced osteoporosis in vitro. Int. J. Mol. Sci. 2020, 22, 38. [Google Scholar] [CrossRef]
- Lopez-Santalla, M.; Fernandez-Perez, R.; Garin, M.I. Mesenchymal Stem/Stromal Cells for Rheumatoid Arthritis Treatment: An Update on Clinical Applications. Cells 2020, 9, 1852. [Google Scholar] [CrossRef]
- Lopez-Santalla, M.; Hervas-Salcedo, R.; Fernandez-Garcia, M.; Bueren, J.A.; Garín, M. Cell Therapy with Mesenchymal Stem Cells Induces an Innate Immune Memory Response That Attenuates Experimental Colitis in the Long Term. J. Crohn’s Coliti 2020, 14, 1424–1435. [Google Scholar] [CrossRef]
- Panés, J.; Garcia-Olmo, D.; Van Assche, G.; Colombel, J.F.; Reinisch, W.; Baumgart, D.C.; Dignass, A.; Nachury, M.; Ferrante, M.; Kazemi-Shirazi, L.; et al. Long-term Efficacy and Safety of Stem Cell Therapy (Cx601) for Complex Perianal Fistulas in Patients with Crohn’s Disease. Gastroenterology 2018, 154, 1334–1342. [Google Scholar] [CrossRef] [Green Version]
- Ra, J.C.; Kang, S.K.; Shin, I.S.; Park, H.G.; Joo, S.A.; Kim, J.G.; Kang, B.-C.; Lee, Y.S.; Nakama, K.; Piao, M.; et al. Stem cell treatment for patients with autoimmune disease by systemic infusion of culture-expanded autologous adipose tissue derived mesenchymal stem cells. J. Transl. Med. 2011, 9, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, J.; Li, X.; Zhang, H.; Wang, D.; Feng, X.; Wang, H.; Hua, B.; Liu, B.; Sun, L. Allogeneic mesenchymal stem cells transplantation in patients with refractory RA. Clin. Rheumatol. 2011, 31, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Álvaro-Gracia, J.M.; Jover, J.A.; García-Vicuña, R.; Carreño, L.; Alonso, A.; Marsal, S.; Blanco, F.; Martínez-Taboada, V.M.; Taylor, P.; Martín-Martín, C.; et al. Intravenous administration of expanded allogeneic adipose-derived mesenchymal stem cells in refractory rheumatoid arthritis (Cx611): Results of a multicentre, dose escalation, randomised, single-blind, placebo-controlled phase Ib/IIa clinical trial. Ann. Rheum. Dis. 2017, 76, 196–202. [Google Scholar] [CrossRef]
- Wang, L.; Wang, L.; Cong, X.; Liu, G.; Zhou, J.; Bai, B.; Li, Y.; Bai, W.; Li, M.; Ji, H.; et al. Human Umbilical Cord Mesenchymal Stem Cell Therapy for Patients with Active Rheumatoid Arthritis: Safety and Efficacy. Stem Cells Dev. 2013, 22, 3192–3202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Huang, S.; Li, S.; Li, M.; Shi, J.; Bai, W.; Wang, Q.; Zheng, L.; Liu, Y. Efficacy and Safety of Umbilical Cord Mesenchymal Stem Cell Therapy for Rheumatoid Arthritis Patients: A Prospective Phase I/II Study. Drug Des. Dev. Ther. 2019, 13, 4331–4340. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; He, X.; Zhao, R.-S.; Guo, W.; Zhu, M.; Xing, W.; Jiang, D.; Liu, C.-Y.; Xu, X. Serum IFN-γ levels predict the therapeutic effect of mesenchymal stem cell transplantation in active rheumatoid arthritis. J. Transl. Med. 2018, 16, 165. [Google Scholar] [CrossRef] [Green Version]
- Alvaro-Gracia, J.M.; Jover, J.A.; Garcia-Vicuña, R.; Carreño, L.; Alonso, A.; Marsal, S.; Blanco, F.J.; Martínez-Taboada, V.M.; Taylor, P.C.; Díaz-González, F.; et al. Phase IB/IIA study on intravenous administration of expanded allogeneic adipose-derived mesenchymal stem cells in refractory rheumatoid arthritis patients. In Proceeding of 2013 ACR/ARHP Annual Meeting, San Diego, CA, USA, 25–30 October 2013; p. 2644. [Google Scholar]
- Shabgah, A.G.; Shariati-Sarabi, Z.; Tavakkol-Afshari, J.; Ghasemi, A.; Ghoryani, M.; Mohammadi, M. A significant decrease of BAFF, APRIL, and BAFF receptors following mesenchymal stem cell transplantation in patients with refractory rheumatoid arthritis. Gene 2020, 732, 144336. [Google Scholar] [CrossRef]
- Shabgah, A.G.; Shariati-Sarabi, Z.; Tavakkol-Afshari, J.; Ghoryani, M.; Mohammadi, M. Possible anti-inflammatory effects of mesenchymal stem cells transplantation via changes in CXCL8 levels in patients with refractory rheumatoid arthritis. Int. J. Mol. Cell. Med. 2019, 8, 191–199. [Google Scholar]
- Park, E.H.; Lim, H.S.; Lee, S.; Roh, K.; Seo, K.W.; Kang, K.S.; Shin, K. Intravenous infusion of umbilical cord blood-derived mesenchymal stem cells in rheumatoid arthritis: A phase Ia clinical trial. Stem Cells Transl. Med. 2018, 7, 636–642. [Google Scholar] [CrossRef] [Green Version]
- Kafaja, S.; Segal, K.; Skerrett, D.; Itescu, S.; Furst, D. FRI0220 Allogeneic mesenchymal precursor cells (MPCS): A novel approach to treating biologic refractory rheumatoid arthritis. Ann. Rheum. Dis. 2017, 76, 566–567. [Google Scholar]
- Kabat, M.; Bobkov, I.; Kumar, S.; Grumet, M. Trends in mesenchymal stem cell clinical trials 2004–2018: Is efficacy optimal in a narrow dose range? Stem Cells Transl. Med. 2019, 9, 17–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galipeau, J.; Sensébé, L. Mesenchymal Stromal Cells: Clinical Challenges and Therapeutic Opportunities. Cell Stem Cell 2018, 22, 824–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarsenova, M.; Issabekova, A.; Abisheva, S.; Rutskaya-Moroshan, K.; Ogay, V.; Saparov, A. Mesenchymal Stem Cell-Based Therapy for Rheumatoid Arthritis. Int. J. Mol. Sci. 2021, 22, 11592. [Google Scholar] [CrossRef] [PubMed]
- Choudhery, M.S. Strategies to improve regenerative potential of mesenchymal stem cells. World J. Stem Cells 2021, 13, 1845–1862. [Google Scholar] [CrossRef]
- Pevsner-Fischer, M.; Levin, S.; Zipori, D. The origins of mesenchymal stromal cell heterogeneity. Stem Cell Rev. Rep. 2011, 7, 560–568. [Google Scholar] [CrossRef]
- Olmedo-Moreno, L.; Aguilera, Y.; Baliña-Sánchez, C.; Martín-Montalvo, A.; Capilla-González, V. Heterogeneity of In Vitro Expanded Mesenchymal Stromal Cells and Strategies to Improve Their Therapeutic Actions. Pharmaceutics 2022, 14, 1112. [Google Scholar] [CrossRef]
- Jimenez-Puerta, G.J.; Marchal, J.A.; López-Ruiz, E.; Gálvez-Martín, P. Role of Mesenchymal Stromal Cells as Therapeutic Agents: Potential Mechanisms of Action and Implications in Their Clinical Use. J. Clin. Med. 2020, 9, 445. [Google Scholar] [CrossRef] [Green Version]
- Zhou, T.; Yuan, Z.; Weng, J.; Pei, D.; Du, X.; He, C.; Lai, P. Challenges and advances in clinical applications of mesenchymal stromal cells. J. Hematol. Oncol. 2021, 14, 24. [Google Scholar] [CrossRef]
- Damasceno, P.K.F.; de Santana, T.A.; Santos, G.C.; Orge, I.D.; Silva, D.N.; Albuquerque, J.F.; Golinelli, G.; Grisendi, G.; Pinelli, M.; Ribeiro Dos Santos, R.; et al. Genetic Engineering as a Strategy to Improve the Therapeutic Efficacy of Mesenchymal Stem/Stromal Cells in Regenerative Medicine. Front. Cell Dev. Biol. 2020, 8, 737. [Google Scholar] [CrossRef]
- Mastrolia, I.; Foppiani, E.M.; Murgia, A.; Candini, O.; Samarelli, A.V.; Grisendi, G.; Veronesi, E.; Horwitz, E.M.; Dominici, M. Challenges in Clinical Development of Mesenchymal Stromal/Stem Cells: Concise Review. Stem Cells Transl. Med. 2019, 8, 1135–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeske, R.; Yuan, X.; Fu, Q.; Bunnell, B.A.; Logan, T.M.; Li, Y. In Vitro Culture Expansion Shifts the Immune Phenotype of Human Adipose-Derived Mesenchymal Stem Cells. Front. Immunol. 2021, 12, 621744. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.Q.; Zhu, J.; Ankrum, J.A. Manufacturing of primed mesenchymal stromal cells for therapy. Nat. Biomed. Eng. 2019, 3, 90–104. [Google Scholar] [CrossRef]
- Tannenbaum, S.E.; Reubinoff, B.E. Advances in hPSC expansion towards therapeutic entities: A review. Cell Prolif. 2022, 55, e13247. [Google Scholar] [CrossRef]
- Jankovic, M.G.; Stojkovic, M.; Bojic, S.; Jovicic, N.; Kovacevic, M.M.; Ivosevic, Z.; Juskovic, A.; Kovacevic, V.; Ljujic, B. Scaling up human mesenchymal stem cell manufacturing using bioreactors for clinical uses. Curr. Res. Transl. Med. 2023, 71, 103393. [Google Scholar] [CrossRef]
- Freitas-Ribeiro, S.; Reis, R.L.; Pirraco, R.P. Long-term and short-term preservation strategies for tissue engineering and regenerative medicine products: State of the art and emerging trends. PNAS Nexus 2022, 1, pgac212. [Google Scholar] [CrossRef]
- Lv, B.; Zhang, X.; Yuan, J.; Chen, Y.; Ding, H.; Cao, X.; Huang, A. Biomaterial-supported MSC transplantation enhances cell-cell communication for spinal cord injury. Stem Cell Res. Ther. 2021, 12, 36. [Google Scholar] [CrossRef] [PubMed]
- Noronha, N.C.; Mizukami, A.; Caliári-Oliveira, C.; Cominal, J.G.; Rocha, J.L.M.; Covas, D.T.; Swiech, K.; Malmegrim, K.C.R. Priming approaches to improve the efficacy of mesenchymal stromal cell-based therapies. Stem Cell Res. Ther. 2019, 10, 131, Erratum in Stem Cell Res Ther. 2019, 10, 132. [Google Scholar] [CrossRef] [Green Version]
- Conforti, A.; Scarsella, M.; Starc, N.; Giorda, E.; Biagini, S.; Proia, A.; Carsetti, R.; Locatelli, F.; Bernardo, M.E. Microvescicles derived from mesenchymal stromal cells are not as effective as their cellular counterpart in the ability to modulate immune responses in vitro. Stem Cells Dev. 2014, 23, 2591–2599. [Google Scholar] [CrossRef] [Green Version]
- Carreras-Planella, L.; Monguió-Tortajada, M.; Borràs, F.E.; Franquesa, M. Immunomodulatory effect of MSC on B cells is independent of secreted extracellular vesicles. Front. Immunol. 2019, 10, 1288, Erratum in Front. Immunol. 2019, 10, 2413. [Google Scholar] [CrossRef] [Green Version]
- Zidan, A.A.; Al-Hawwas, M.; Perkins, G.B.; Mourad, G.M.; Stapledon, C.J.M.; Bobrovskaya, L.; Zhou, X.F.; Hurtado, P.R. Characterization of urine stem cell-derived extracellular vesicles reveals B cell stimulating cargo. Int. J. Mol. Sci. 2021, 22, 459. [Google Scholar] [CrossRef] [PubMed]

| Tissue of Origin | Potential Advantages | Limitations | References |
|---|---|---|---|
| Bone marrow (BM-MSCs) |
|
| [47,48,49,50,51,52,53,54] |
| Umbilical cord (UC-MSCs) |
|
| [55,56,57,58,59,60,61,62] |
| Umbilical cord blood (UCB-MSCs) |
|
| [63,64,65,66] |
| Adipose tissue (AD-MSCs) |
|
| [50,58,67,68,69,70,71,72,73] |
| Synovial membrane (SM-MSCs) |
|
| [74,75,76,77,78,79,80] |
| Tissue Microenvironment | MSC Phenotype | Characteristics | Expressed Molecules |
|---|---|---|---|
| Low inflammatory condition (Low levels of TNF-α and IFN-γ) | Proinflammatory mesenchymal stem cells (MSC1) |
|
|
| High inflammatory microenvironment (High levels of TNF-α and IFN-γ) | Anti-inflammatory mesenchymal stem cells (MSC2) |
|
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shimizu, Y.; Ntege, E.H.; Azuma, C.; Uehara, F.; Toma, T.; Higa, K.; Yabiku, H.; Matsuura, N.; Inoue, Y.; Sunami, H. Management of Rheumatoid Arthritis: Possibilities and Challenges of Mesenchymal Stromal/Stem Cell-Based Therapies. Cells 2023, 12, 1905. https://doi.org/10.3390/cells12141905
Shimizu Y, Ntege EH, Azuma C, Uehara F, Toma T, Higa K, Yabiku H, Matsuura N, Inoue Y, Sunami H. Management of Rheumatoid Arthritis: Possibilities and Challenges of Mesenchymal Stromal/Stem Cell-Based Therapies. Cells. 2023; 12(14):1905. https://doi.org/10.3390/cells12141905
Chicago/Turabian StyleShimizu, Yusuke, Edward Hosea Ntege, Chinatsu Azuma, Fuminari Uehara, Takashi Toma, Kotaro Higa, Hiroki Yabiku, Naoki Matsuura, Yoshikazu Inoue, and Hiroshi Sunami. 2023. "Management of Rheumatoid Arthritis: Possibilities and Challenges of Mesenchymal Stromal/Stem Cell-Based Therapies" Cells 12, no. 14: 1905. https://doi.org/10.3390/cells12141905