
Interactions between the Astrocytic Volume-Regulated Anion Channel and Aquaporin 4 in Hyposmotic Regulation of Vasopressin Neuronal Activity in the Supraoptic Nucleus


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Preparation of Acute SON Slices from Young Adult Rats
2.3. Preparation of Astrocyte Cell Culture from Rat Pups
2.4. Patch-Clamp Recordings
2.5. Immunostaining
2.6. Volume Measurement
2.7. GFAP Fillament and Colocalization Analysis
2.8. Statistical Analysis
3. Results
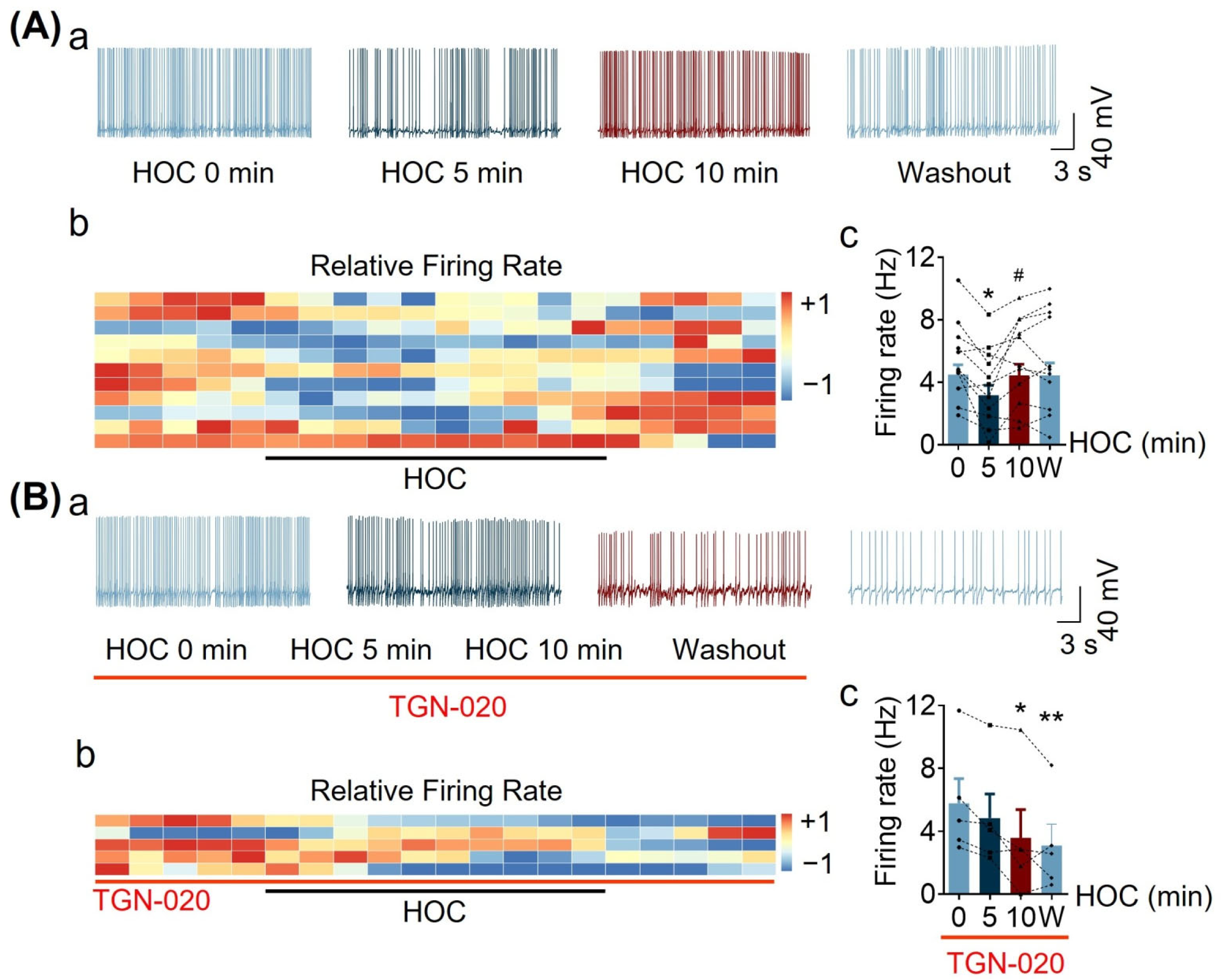
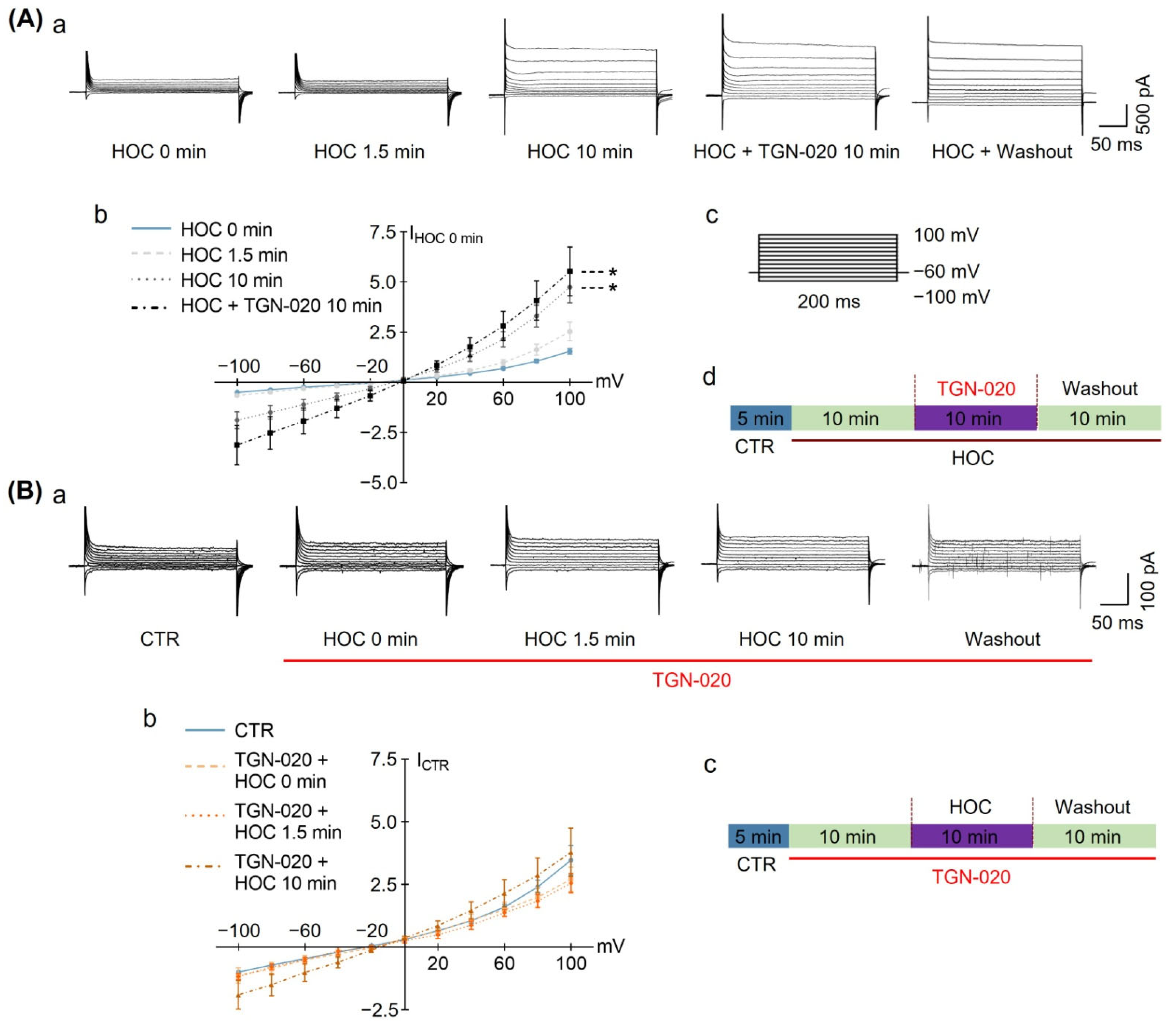
3.1. Effect of Blocking Astrocytic AQP4 Activity with TGN-020 on HOC-Modulated VP Neuronal Activity in Acute SON Slices
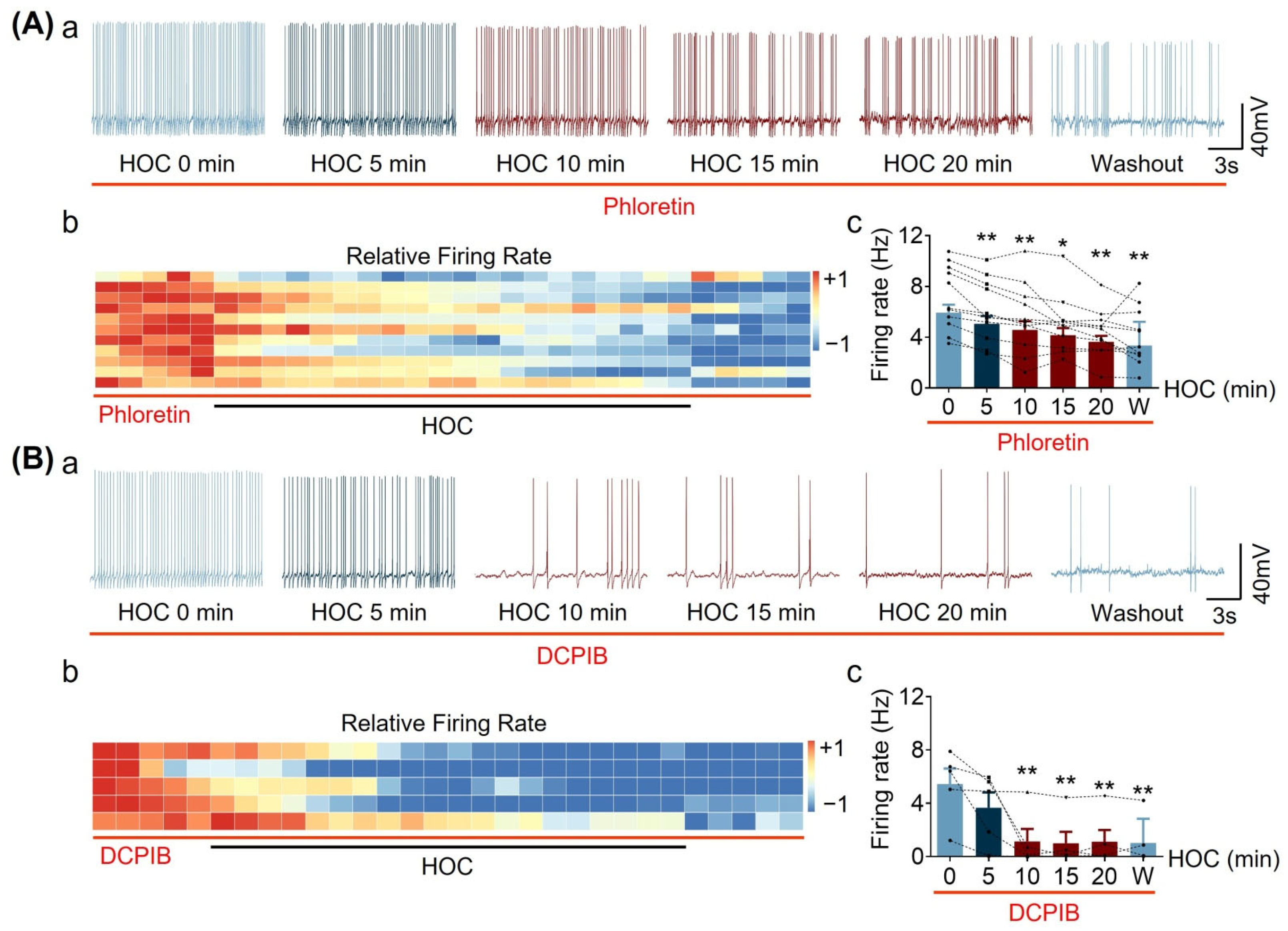
3.2. Effect of Blocking Astrocytic VRAC Activity on HOC-Modulated VP Neuronal Activity in Acute SON Slices
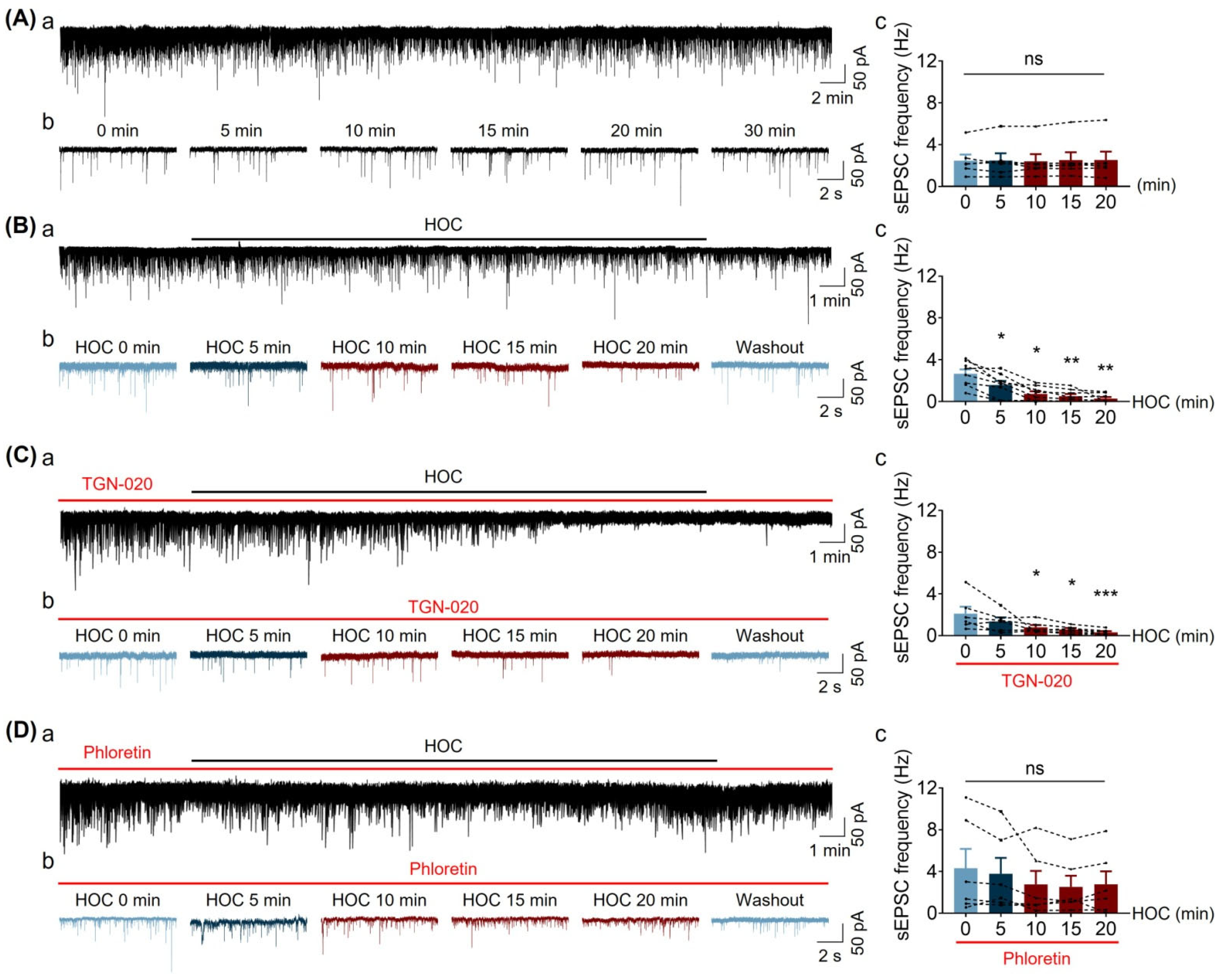
3.3. Effect of HOC on Spontaneous Excitatory Postsynaptic Current (sEPSC) Frequency of VP Neurons in Acute SON Slices
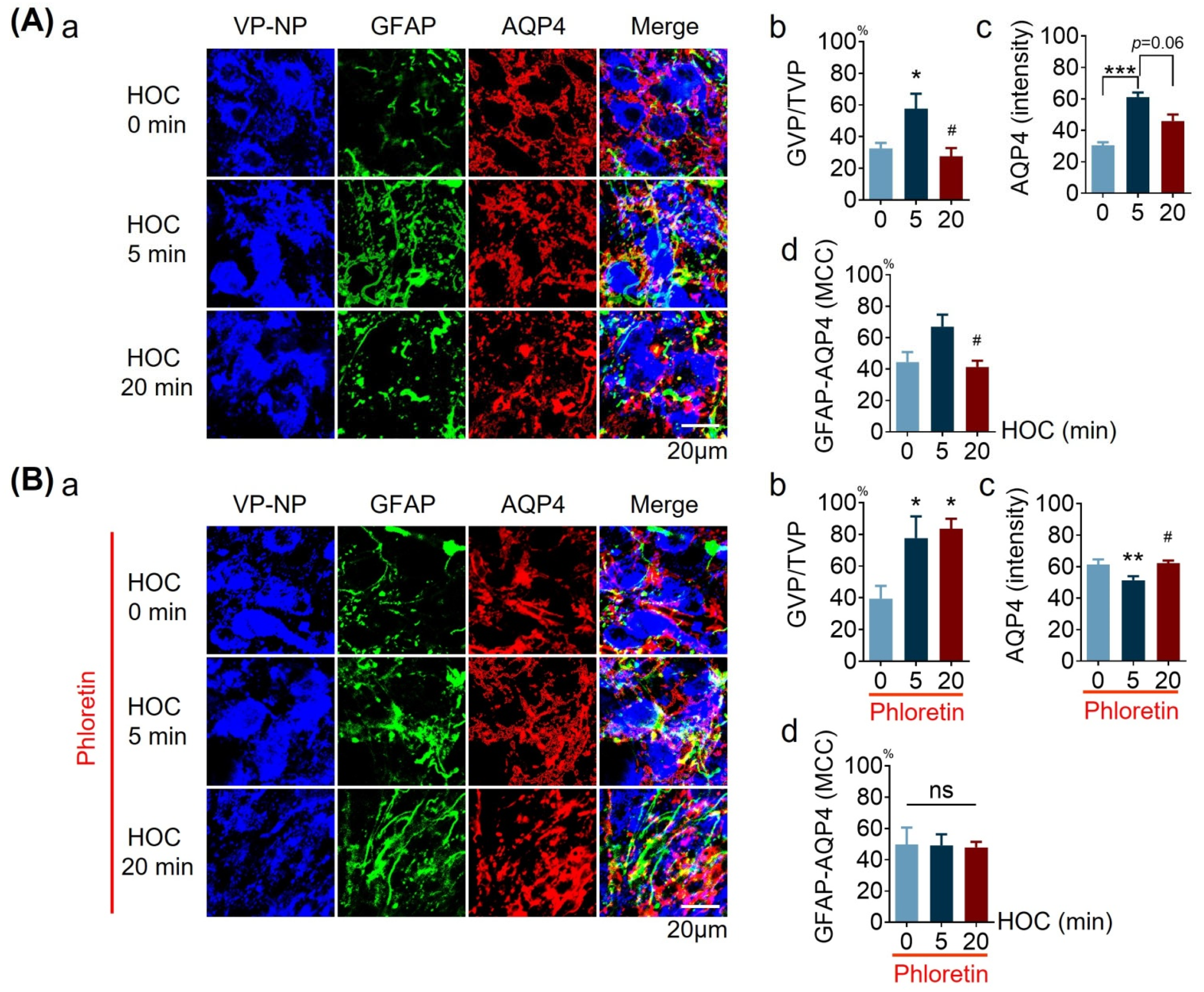
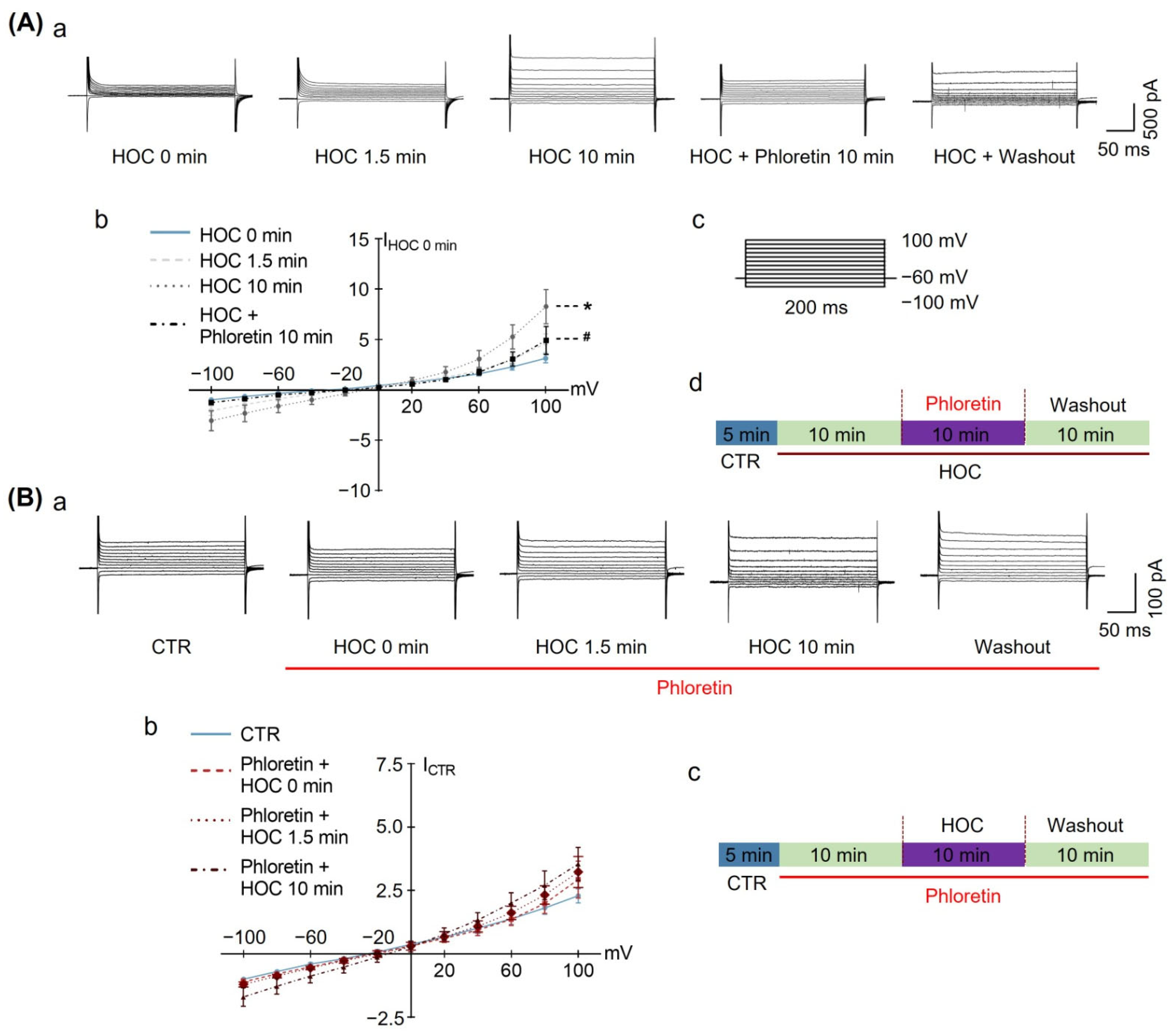
3.4. Effects of Phloretin on HOC-Evoked Changes in Distribution of GFAP Filaments, AQP4 Expresion, and Their Colocalizaion in Acute SON Slices
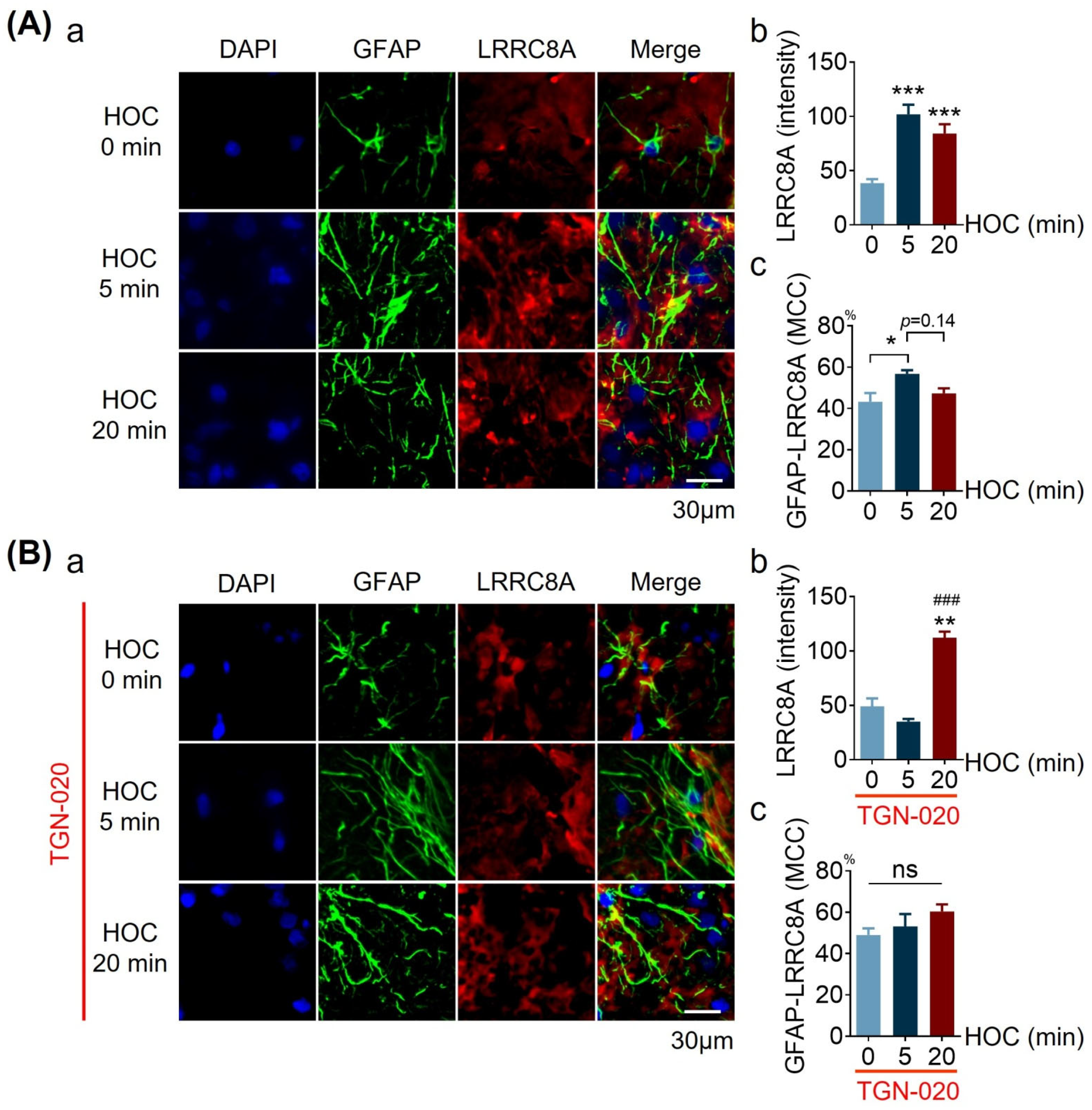
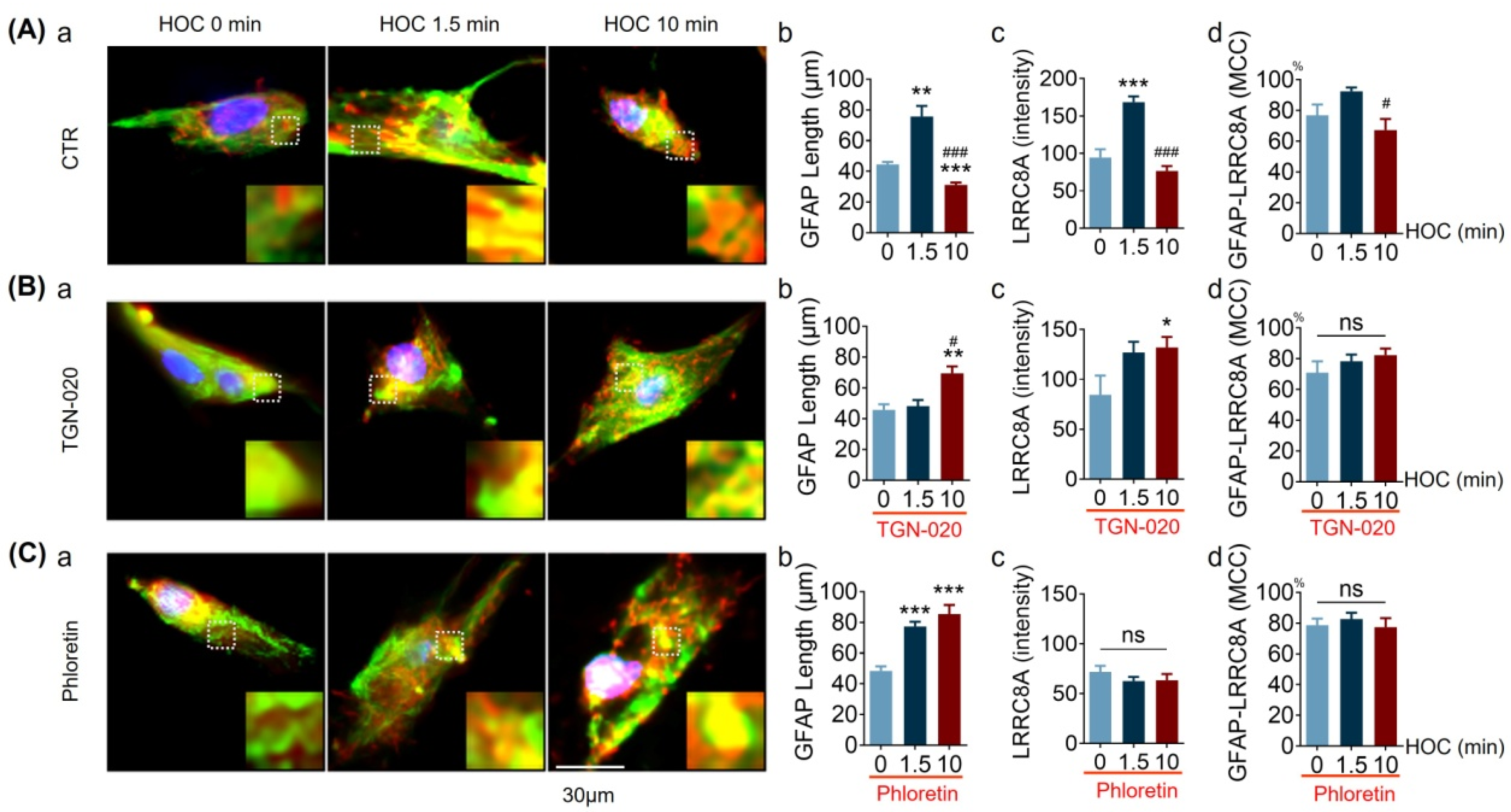
3.5. Effects of TGN-020 on LRRC8A Expression and Its Colocalization with GFAP in Acute SON Slices during HOC
3.6. Effects of HOC on VRAC Currents in Cultured Hypothalamic Astrocytes
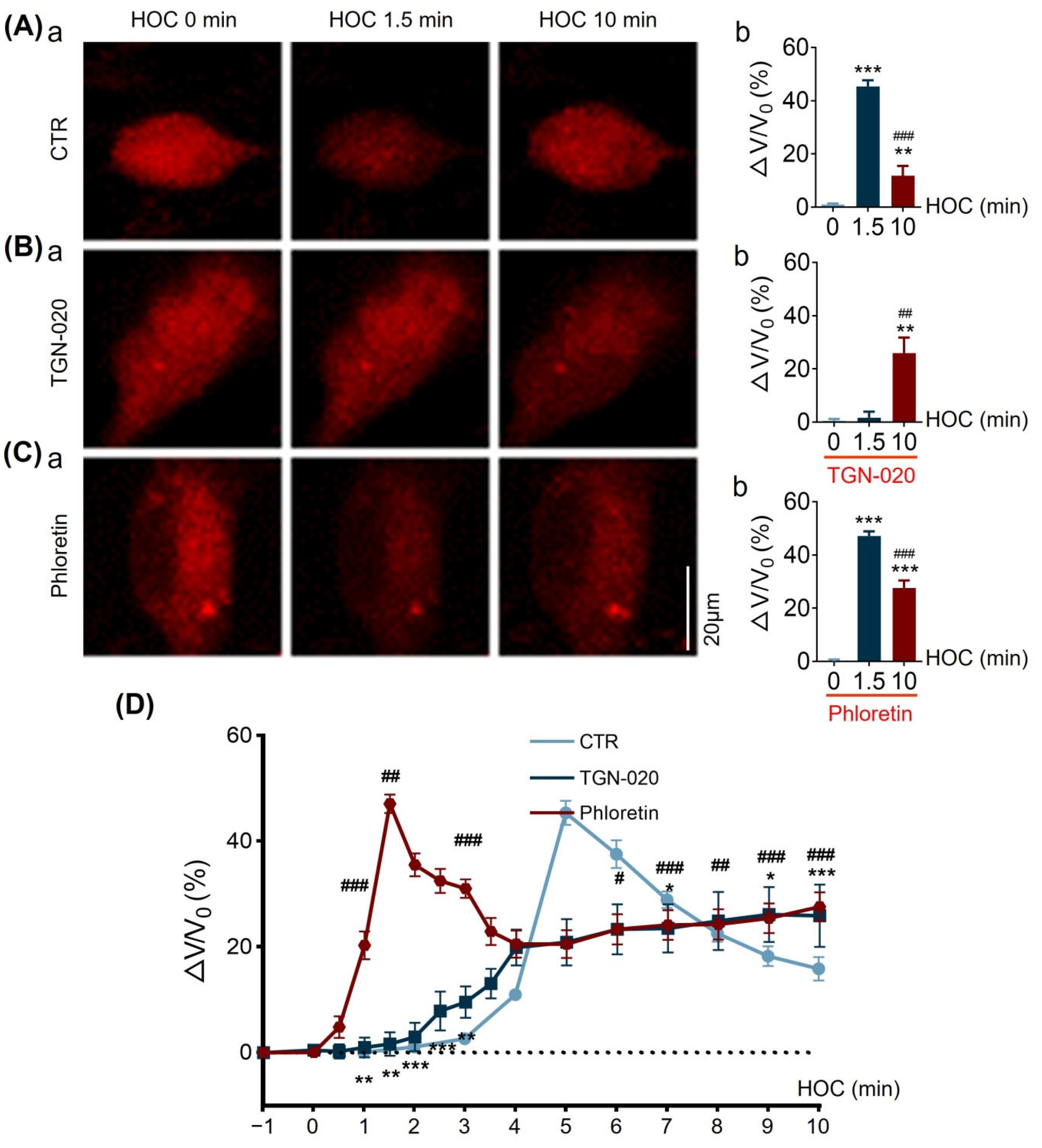
3.7. TGN-020 or Phloretin Distinctly Affects HOC-Evoked Changes in Astrocytic Volume in Cell Culture
3.8. Effects of HOC, and Associated AQP4 and VRAC Activity, on Astrocytic GFAP and LRRC8A Distribution
4. Discussion
4.1. Dual HOC Effect on Astrocytic Plasticity and VP Neuronal Activity
4.2. HOC Effect on sEPSC Frequency and Role of Astrocytes
4.3. Dual HOC Effect on GFAP Plasticity and Associated APQ4 and/or VRAC Activity
4.4. Role of AQP4 in Dual HOC Effect on Astrocytic Volume
4.5. Role of VRAC in Dual HOC Effect on Astrocytic Volume and VP Neuronal Activity
4.6. Interactions of Volume-Regulating Channles in RVD: Translational Relevance
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ransom, B.R.; Ransom, C.B. Astrocytes: Multitalented stars of the central nervous system. Methods Mol. Biol. 2012, 814, 3–7. [Google Scholar] [CrossRef]
- Kohler, S.; Winkler, U.; Hirrlinger, J. Heterogeneity of Astrocytes in Grey and White Matter. Neurochem. Res. 2021, 46, 3–14. [Google Scholar] [CrossRef]
- Zorec, R.; Parpura, V.; Verkhratsky, A. Astroglial vesicular network: Evolutionary trends, physiology and pathophysiology. Acta Physiol. 2017, 222, e12915. [Google Scholar] [CrossRef] [Green Version]
- Escartin, C.; Galea, E.; Lakatos, A.; O’Callaghan, J.P.; Petzold, G.C.; Serrano-Pozo, A.; Steinhauser, C.; Volterra, A.; Carmignoto, G.; Agarwal, A.; et al. Reactive astrocyte nomenclature, definitions, and future directions. Nat. Neurosci. 2021, 24, 312–325. [Google Scholar] [CrossRef]
- Wang, S.C.; Parpura, V.; Wang, Y.F. Astroglial Regulation of Magnocellular Neuroendocrine Cell Activities in the Supraoptic Nucleus. Neurochem. Res. 2021, 46, 2586–2600. [Google Scholar] [CrossRef]
- Wang, Y.F.; Sun, M.Y.; Hou, Q.; Parpura, V. Hyposmolality differentially and spatiotemporally modulates levels of glutamine synthetase and serine racemase in rat supraoptic nucleus. Glia 2013, 61, 529–538. [Google Scholar] [CrossRef]
- Wang, X.; Li, T.; Liu, Y.; Jia, S.; Liu, X.; Jiang, Y.; Wang, P.; Parpura, V.; Wang, Y.F. Aquaporin 4 differentially modulates osmotic effects on vasopressin neurons in rat supraoptic nucleus. Acta Physiol. 2021, 232, e13672. [Google Scholar] [CrossRef]
- Benfenati, V.; Ferroni, S. Water transport between CNS compartments: Functional and molecular interactions between aquaporins and ion channels. Neuroscience 2010, 168, 926–940. [Google Scholar] [CrossRef]
- Kitchen, P.; Salman, M.M.; Halsey, A.M.; Clarke-Bland, C.; MacDonald, J.A.; Ishida, H.; Vogel, H.J.; Almutiri, S.; Logan, A.; Kreida, S.; et al. Targeting Aquaporin-4 Subcellular Localization to Treat Central Nervous System Edema. Cell 2020, 181, 784–799. [Google Scholar] [CrossRef]
- Han, Y.E.; Kwon, J.; Won, J.; An, H.; Jang, M.W.; Woo, J.; Lee, J.S.; Park, M.G.; Yoon, B.E.; Lee, S.E.; et al. Tweety-homolog (Ttyh) Family Encodes the Pore-forming Subunits of the Swelling-dependent Volume-regulated Anion Channel (VRACswell) in the Brain. Exp. Neurobiol. 2019, 28, 183–215. [Google Scholar] [CrossRef]
- Jiang, Y.H.; Li, T.; Liu, Y.; Liu, X.; Jia, S.; Hou, C.; Chen, G.; Wang, H.; Ling, S.; Gao, Q.; et al. Contribution of inwardly rectifying K(+) channel 4.1 of supraoptic astrocytes to the regulation of vasopressin neuronal activity by hypotonicity. Glia 2023, 71, 704–719. [Google Scholar] [CrossRef]
- Benfenati, V.; Caprini, M.; Dovizio, M.; Mylonakou, M.N.; Ferroni, S.; Ottersen, O.P.; Amiry-Moghaddam, M. An aquaporin-4/transient receptor potential vanilloid 4 (AQP4/TRPV4) complex is essential for cell-volume control in astrocytes. Proc. Natl. Acad. Sci. USA 2011, 108, 2563–2568. [Google Scholar] [CrossRef] [Green Version]
- Reed, M.M.; Blazer-Yost, B. Channels and Transporters in Astrocyte Volume Regulation in Health and Disease. Cell. Physiol. Biochem. 2022, 56, 12–30. [Google Scholar] [CrossRef]
- Qiu, Z.; Dubin, A.E.; Mathur, J.; Tu, B.; Reddy, K.; Miraglia, L.J.; Reinhardt, J.; Orth, A.P.; Patapoutian, A. SWELL1, a plasma membrane protein, is an essential component of volume-regulated anion channel. Cell 2014, 157, 447–458. [Google Scholar] [CrossRef] [Green Version]
- Voss, F.K.; Ullrich, F.; Münch, J.; Lazarow, K.; Lutter, D.; Mah, N.; Andrade-Navarro, M.A.; von Kries, J.P.; Stauber, T.; Jentsch, T.J. Identification of LRRC8 heteromers as an essential component of the volume-regulated anion channel VRAC. Science 2014, 344, 634–638. [Google Scholar] [CrossRef] [Green Version]
- Formaggio, F.; Saracino, E.; Mola, M.G.; Rao, S.B.; Amiry-Moghaddam, M.; Muccini, M.; Zamboni, R.; Nicchia, G.P.; Caprini, M.; Benfenati, V. LRRC8A is essential for swelling-activated chloride current and for regulatory volume decrease in astrocytes. FASEB J. 2019, 33, 101–113. [Google Scholar] [CrossRef] [Green Version]
- Benfenati, V.; Nicchia, G.P.; Svelto, M.; Rapisarda, C.; Frigeri, A.; Ferroni, S. Functional down-regulation of volume-regulated anion channels in AQP4 knockdown cultured rat cortical astrocytes. J. Neurochem. 2007, 100, 87–104. [Google Scholar] [CrossRef]
- Mola, M.G.; Saracino, E.; Formaggio, F.; Amerotti, A.G.; Barile, B.; Posati, T.; Cibelli, A.; Frigeri, A.; Palazzo, C.; Zamboni, R.; et al. Cell Volume Regulation Mechanisms in Differentiated Astrocytes. Cell Physiol. Biochem. 2021, 55, 196–212. [Google Scholar] [CrossRef]
- McCarthy, K.D.; de Vellis, J. Preparation of separate astroglial and oligodendroglial cell cultures from rat cerebral tissue. J. Cell Biol. 1980, 85, 890–902. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, W.E.; Foehring, R.C.; Kirchner, M.K.; Sladek, C.D. Electrophysiological properties of identified oxytocin and vasopressin neurones. J. Neuroendocrinol. 2019, 31, e12666. [Google Scholar] [CrossRef]
- Nimmerjahn, A.; Helmchen, F. In vivo labeling of cortical astrocytes with sulforhodamine 101 (SR101). Cold. Spring Harb. Protoc. 2012, 2012, 326–334. [Google Scholar] [CrossRef]
- Murphy, T.R.; Davila, D.; Cuvelier, N.; Young, L.R.; Lauderdale, K.; Binder, D.K.; Fiacco, T.A. Hippocampal and Cortical Pyramidal Neurons Swell in Parallel with Astrocytes during Acute Hypoosmolar Stress. Front. Cell. Neurosci. 2017, 11, 275. [Google Scholar] [CrossRef] [Green Version]
- Dunn, K.W.; Kamocka, M.M.; McDonald, J.H. A practical guide to evaluating colocalization in biological microscopy. Am. J. Physiol. Cell Physiol. 2011, 300, C723–C742. [Google Scholar] [CrossRef] [Green Version]
- Abdullaev, I.F.; Rudkouskaya, A.; Schools, G.P.; Kimelberg, H.K.; Mongin, A.A. Pharmacological comparison of swelling-activated excitatory amino acid release and Cl- currents in cultured rat astrocytes. J. Physiol. 2006, 572, 677–689. [Google Scholar] [CrossRef]
- Dierckx, T.; Haidar, M.; Grajchen, E.; Wouters, E.; Vanherle, S.; Loix, M.; Boeykens, A.; Bylemans, D.; Hardonniere, K.; Kerdine-Romer, S.; et al. Phloretin suppresses neuroinflammation by autophagy-mediated Nrf2 activation in macrophages. J. Neuroinflamm. 2021, 18, 148. [Google Scholar] [CrossRef]
- Sabirov, R.Z.; Kurbannazarova, R.S.; Melanova, N.R.; Okada, Y. Volume-sensitive anion channels mediate osmosensitive glutathione release from rat thymocytes. PLoS ONE 2013, 8, e55646. [Google Scholar] [CrossRef] [Green Version]
- Bowens, N.H.; Dohare, P.; Kuo, Y.H.; Mongin, A.A. DCPIB, the proposed selective blocker of volume-regulated anion channels, inhibits several glutamate transport pathways in glial cells. Mol. Pharmacol. 2013, 83, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Lv, J.; Liang, Y.; Zhang, S.; Lan, Q.; Xu, Z.; Wu, X.; Kang, L.; Ren, J.; Cao, Y.; Wu, T.; et al. DCPIB, an Inhibitor of Volume-Regulated Anion Channels, Distinctly Modulates K2P Channels. ACS Chem. Neurosci. 2019, 10, 2786–2793. [Google Scholar] [CrossRef]
- Afzal, A.; Figueroa, E.E.; Kharade, S.V.; Bittman, K.; Matlock, B.K.; Flaherty, D.K.; Denton, J.S. The LRRC8 volume-regulated anion channel inhibitor, DCPIB, inhibits mitochondrial respiration independently of the channel. Physiol. Rep. 2019, 7, e14303. [Google Scholar] [CrossRef] [Green Version]
- Yagil, C.; Sladek, C.D. Osmotic regulation of vasopressin and oxytocin release is rate sensitive in hypothalamoneurohypophysial explants. Am. J. Physiol. 1990, 258, R492–R500. [Google Scholar] [CrossRef]
- Syeda, R.; Qiu, Z.; Dubin, A.E.; Murthy, S.E.; Florendo, M.N.; Mason, D.E.; Mathur, J.; Cahalan, S.M.; Peters, E.C.; Montal, M.; et al. LRRC8 Proteins Form Volume-Regulated Anion Channels that Sense Ionic Strength. Cell 2016, 164, 499–511. [Google Scholar] [CrossRef] [Green Version]
- Nilius, B. Is the volume-regulated anion channel VRAC a “water-permeable” channel? Neurochem. Res. 2004, 29, 3–8. [Google Scholar] [CrossRef]
- Pajarillo, E.; Rizor, A.; Lee, J.; Aschner, M.; Lee, E. The role of astrocytic glutamate transporters GLT-1 and GLAST in neurological disorders: Potential targets for neurotherapeutics. Neuropharmacology 2019, 161, 107559. [Google Scholar] [CrossRef]
- Yang, J.; Vitery, M.D.C.; Chen, J.; Osei-Owusu, J.; Chu, J.; Qiu, Z. Glutamate-Releasing SWELL1 Channel in Astrocytes Modulates Synaptic Transmission and Promotes Brain Damage in Stroke. Neuron 2019, 102, 813–827. [Google Scholar] [CrossRef]
- Hyzinski-García, M.C.; Rudkouskaya, A.; Mongin, A.A. LRRC8A protein is indispensable for swelling-activated and ATP-induced release of excitatory amino acids in rat astrocytes. J. Physiol. 2014, 592, 4855–4862. [Google Scholar] [CrossRef]
- Deleuze, C.; Duvoid, A.; Hussy, N. Properties and glial origin of osmotic-dependent release of taurine from the rat supraoptic nucleus. J. Physiol. 1998, 507 Pt 2, 463–471. [Google Scholar] [CrossRef]
- Wang, Y.F.; Parpura, V. Astroglial Modulation of Hydromineral Balance and Cerebral Edema. Front. Mol. Neurosci. 2018, 11, 204. [Google Scholar] [CrossRef] [Green Version]
- Verkhratsky, A.; Semyanov, A.; Zorec, R. Physiology of Astroglial Excitability. Function 2020, 1, zqaa016. [Google Scholar] [CrossRef]
- Jorgacevski, J.; Zorec, R.; Potokar, M. Insights into Cell Surface Expression, Supramolecular Organization, and Functions of Aquaporin 4 Isoforms in Astrocytes. Cells 2020, 9, 2622. [Google Scholar] [CrossRef]
- Creighton, B.A.; Ruffins, T.W.; Lorenzo, D.N. Visualizing and Analyzing Intracellular Transport of Organelles and Other Cargos in Astrocytes. J. Vis. Exp. 2019, 150, e60230. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effect | Supporting Role | Effect | Supporting Role | |||||
|---|---|---|---|---|---|---|---|---|
| Measurements | Figure | Early HOC | AQP4 | VRAC | Late HOC | AQP4 | VRAC | |
| Acute Slice | Firing rate | Figure 1 and Figure 2 | ↓ | Y | Y | R | Y | Y |
| sEPSC frequency | Figure 3 | ↓ | Y | Y | ↓ | N | Y | |
| GVP/TVP | Figure 4 | ↑ | NT | N | R | NT | Y | |
| AQP4 intensity/level | Figure 4 | ↑ | NT | Y | R, t | NT | ND-t | |
| GFAP-AQP4 MCC | Figure 4 | ↑, t | NT | ND-t | R | NT | Y | |
| LRRC8A intensity/level | Figure 5 | ↑ | Y | NT | ↑ | N | NT | |
| GFAP-LRRC8A MCC | Figure 5 | ↑ | Y | NT | R, t | ND-t | NT | |
| Astro Culture | VRAC current | Figure 6 and Figure 7 | NC | NA | NA | ↑ | N; Y-pt | Y |
| ∆V/V0 | Figure 8 | ↑ | Y | N | RVD | Y | Y | |
| GFAP filament length | Figure 9 | ↑ | Y | N | R | Y | Y | |
| LRRC8A intensity/level | Figure 9 | ↑ | Y | Y | R | Y | N | |
| GFAP-LRRC8A MCC | Figure 9 | ↑, t | ND-t | ND-t | ↓ | Y | Y | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Wang, X.-R.; Jiang, Y.-H.; Li, T.; Ling, S.; Wang, H.-Y.; Yu, J.-W.; Jia, S.-W.; Liu, X.-Y.; Hou, C.-M.; et al. Interactions between the Astrocytic Volume-Regulated Anion Channel and Aquaporin 4 in Hyposmotic Regulation of Vasopressin Neuronal Activity in the Supraoptic Nucleus. Cells 2023, 12, 1723. https://doi.org/10.3390/cells12131723
Liu Y, Wang X-R, Jiang Y-H, Li T, Ling S, Wang H-Y, Yu J-W, Jia S-W, Liu X-Y, Hou C-M, et al. Interactions between the Astrocytic Volume-Regulated Anion Channel and Aquaporin 4 in Hyposmotic Regulation of Vasopressin Neuronal Activity in the Supraoptic Nucleus. Cells. 2023; 12(13):1723. https://doi.org/10.3390/cells12131723
Chicago/Turabian StyleLiu, Yang, Xiao-Ran Wang, Yun-Hao Jiang, Tong Li, Shuo Ling, Hong-Yang Wang, Jia-Wei Yu, Shu-Wei Jia, Xiao-Yu Liu, Chun-Mei Hou, and et al. 2023. "Interactions between the Astrocytic Volume-Regulated Anion Channel and Aquaporin 4 in Hyposmotic Regulation of Vasopressin Neuronal Activity in the Supraoptic Nucleus" Cells 12, no. 13: 1723. https://doi.org/10.3390/cells12131723