
Effect of HPSE and HPSE2 SNPs on the Risk of Developing Primary Paraskeletal Multiple Myeloma
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
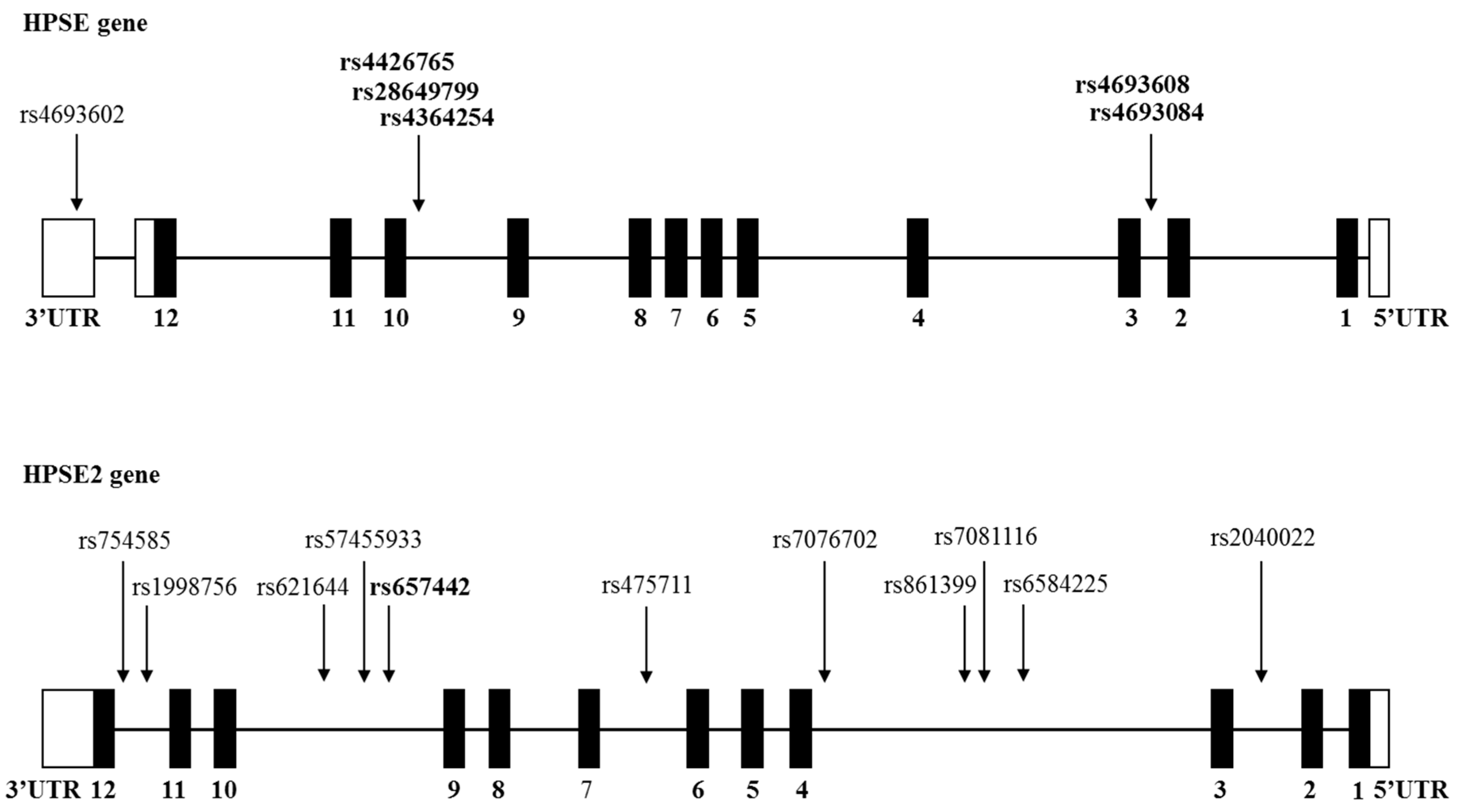
2.2. SNPs Analysis
2.3. DNA Constructs
2.4. Purification of Total RNA, Generation of cDNA, and Real-Time Quantitative RT-PCR
2.5. Statistical Analysis
3. Results
3.1. Analysis of HPSE and HPSE2 Gene SNPs among Multiple Myeloma Patients
3.2. Association between HPSE Gene SNPs and Stage of Multiple Myeloma Patients at Diagnosis
3.3. Association between HPSE Gene SNPs and Primary Bone-Related Disease in Multiple Myeloma
3.4. Association between HPSE2 Gene SNPs and Primary Extramedullary Disease (EMD) in Multiple Myeloma
3.5. Association between HPSE2 Gene SNPs and Secondary Extramedullary Disease (EMD) in Multiple Myeloma
3.6. Association between HPSE2 Gene SNPs and Bone Morbidity in Multiple Myeloma Patients
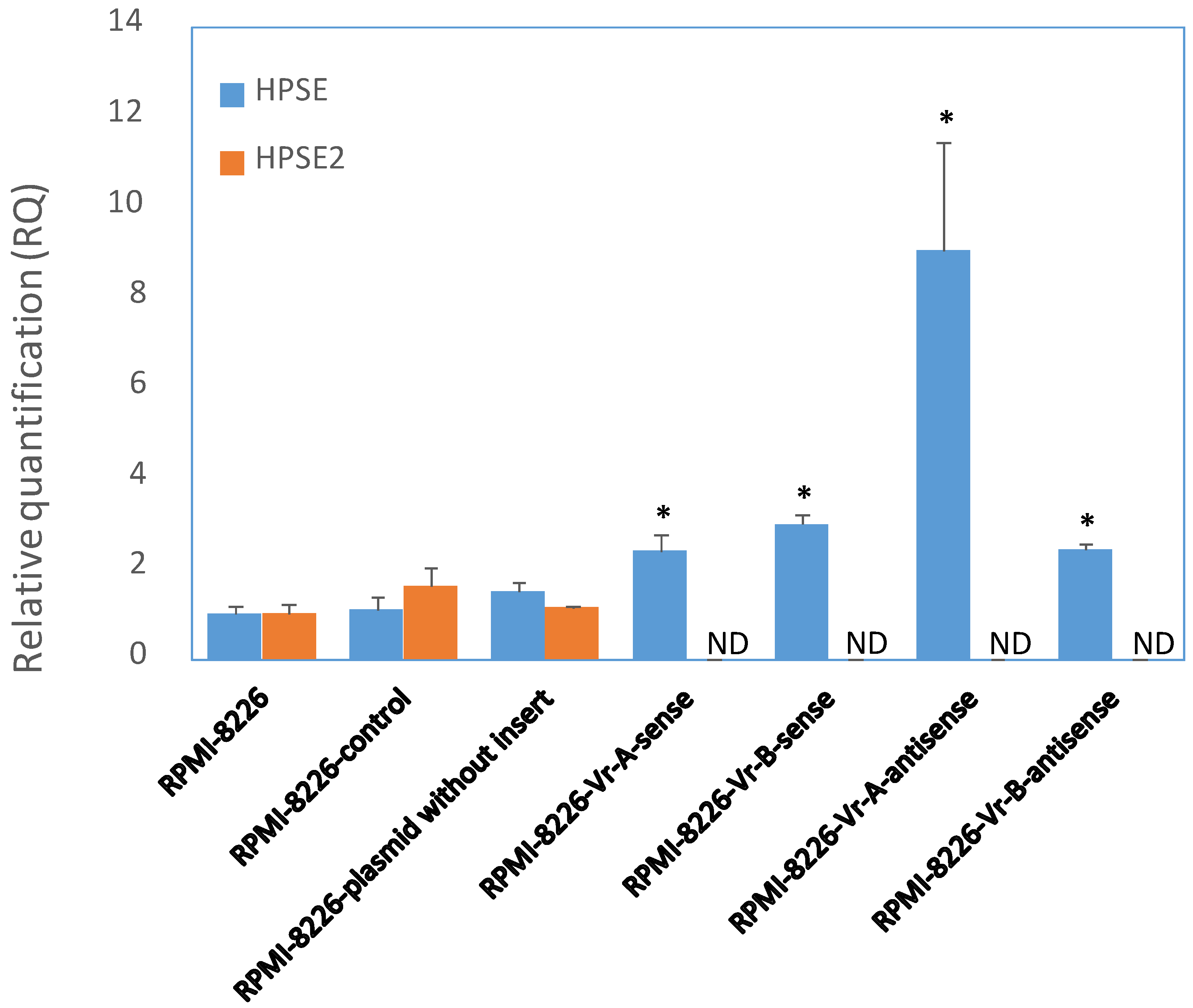
3.7. Modification of HPSE and HPSE2 Gene Expression after Transient Transfection of RPMI-8226 MM Cells with a Fragment of the Enhancer
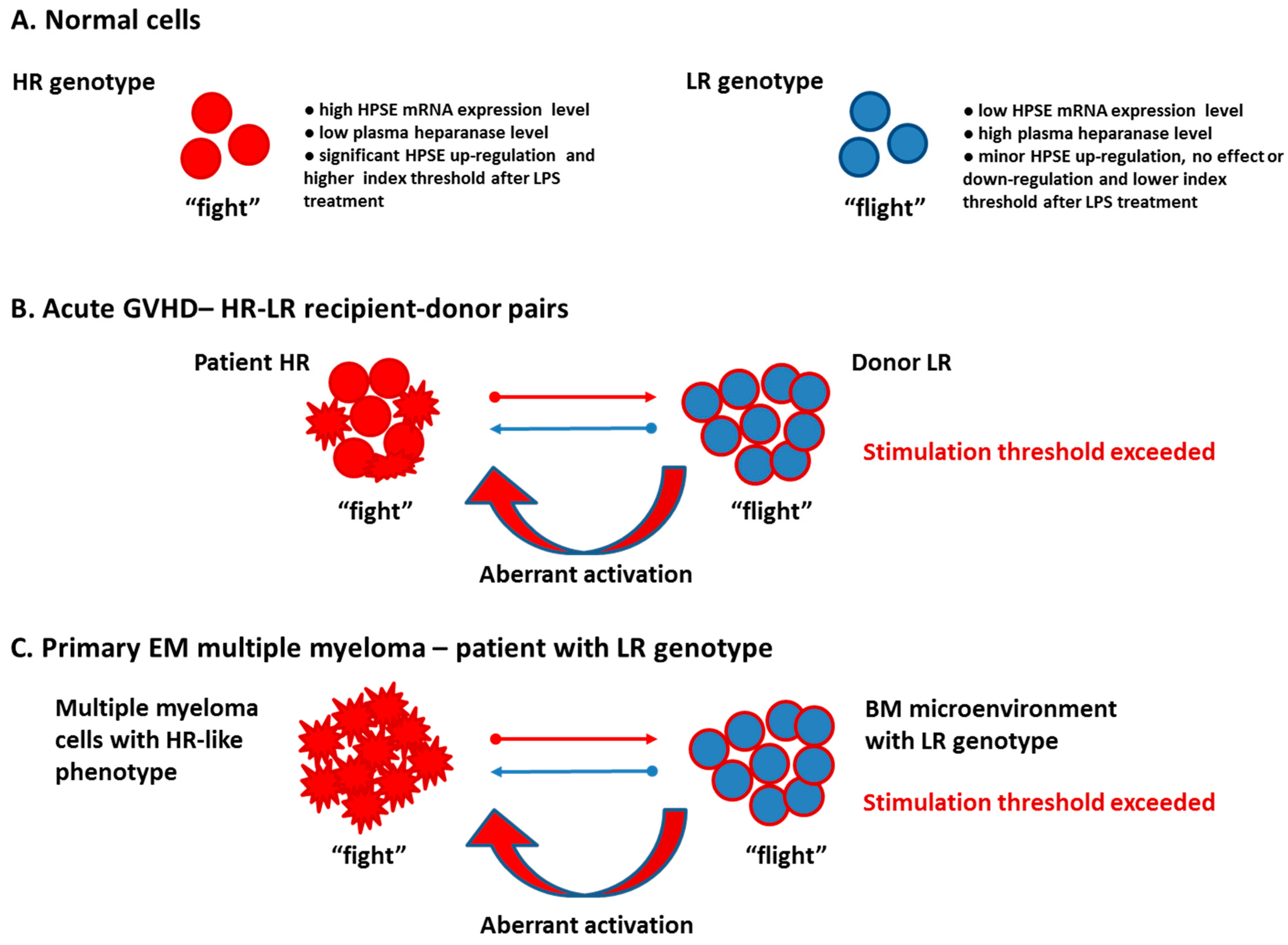
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cowan, A.J.; Green, D.J.; Kwok, M.; Lee, S.; Coffey, D.G.; Holmberg, L.A.; Tuazon, S.; Gopal, A.K.; Libby, E.N. Diagnosis and management of multiple myeloma: A review. JAMA 2022, 327, 464–477. [Google Scholar] [CrossRef]
- Rajkumar, S.V. Multiple myeloma: 2022 update on diagnosis, risk stratification, and management. AJH 2022, 97, 989–1107. [Google Scholar] [CrossRef]
- Blade, J.; Beksac, M.; Caers, J.; Jurczyszyn, A.; von Lilienfeld-Toal, M.; Moreau, P.; Rasche, L.; Rosinol, L.; Usmani, S.Z.; Zamagni, E.; et al. Extramedullary disease in multiple myeloma: A systematic literature review. Blood Cancer J. 2022, 12, 45. [Google Scholar] [CrossRef] [PubMed]
- Rosinol, L.; Beksac, M.; Zamagni, E.; Van de Donk, N.W.C.J.; Anderson, K.C.; Badros, A.; Caers, J.; Cavo, M.; Dimopoulos, M.-A.; Dispenzieri, A.; et al. Expert review on soft-tissue plasmacytomas in multiple myeloma: Definition, disease assessement and treatment considerations. Br. J. Haematol. 2021, 194, 496–507. [Google Scholar] [CrossRef]
- Rodriguez-Otero, P.; Paiva, B.; San-Miguel, J.F. Roadmap to cure multiple myeloma. Cancer Treat. Rev. 2021, 100, 102284. [Google Scholar] [CrossRef] [PubMed]
- Purushothaman, A.; Sanderson, R.D. Heparanase: A dynamic promoter of myeloma progression. Adv. Exp. Med. Biol. 2020, 1221, 331–349. [Google Scholar] [CrossRef] [PubMed]
- Ramani, V.C.; Vlodavsky, I.; Ng, M.; Zhang, Y.; Barbieri, P.; Noseda, A.; Sanderson, R.D. Chemotherapy induces expression and release of heparanase leading to changes associated with an aggressive tumor phenotype. Matrix Biol. 2016, 55, 22–34. [Google Scholar] [CrossRef]
- Ramani, V.C.; Zhan, F.; He, M.; Barbieri, P.; Noseda, A.; Tricot, G.; Sanderson, R.D. Targeting heparanase overcomes chemoresistance and diminishes relapse in myeloma. Oncotarget 2016, 7, 1598–1607. [Google Scholar] [CrossRef] [Green Version]
- Kelly, T.; Miao, H.Q.; Yang, Y.; Navarro, E.; Kussie, P.; Huang, Y.; MacLeod, V.; Casciano, J.; Joseph, L.; Zhan, F.; et al. High heparanase activity in multiple myeloma is associated with elevated microvessel density. Cancer Res. 2003, 63, 8749–8756. [Google Scholar]
- Barash, U.; Zohar, Y.; Wildbaum, G.; Beider, K.; Nagler, A.; Karin, N.; Ilan, N.; Vlodavsky, I. Heparanase enhancers myeloma progression via CXCL10 downregulation. Leukemia 2014, 28, 2178–2187. [Google Scholar] [CrossRef]
- Yang, Y.; Ren, Y.; Ramani, V.C.; Nan, L.; Suva, L.J.; Sanderson, R.D. Heparanase enhances local and systemic osteolysis in multiple myeloma by upregulating the expression and secretion of RANKL. Cancer Res. 2010, 70, 8329–8338. [Google Scholar] [CrossRef] [Green Version]
- Purushothaman, A.; Uyama, T.; Kobayashi, F.; Yamada, S.; Sugahara, K.; Rapraeger, A.C.; Sanderson, R.D. Heparanase-enhanced shedding of syndecan-1 by myeloma cells promotes endothelial invasion and angiogenesis. Blood 2010, 115, 2449–2457. [Google Scholar] [CrossRef] [Green Version]
- Ramani, V.C.; Purushothaman, A.; Stewart, M.D.; Thompson, C.A.; Vlodavsky, I.; Sanderson, R.D. The heparanase/syndecan-1 axis in cancer: Mechanisms and therapies. FEBS J. 2013, 280, 2294–2306. [Google Scholar] [CrossRef] [Green Version]
- Jung, O.; Trapp-Stamborski, V.; Purushothaman, A.; Jin, H.; Wang, H.; Sanderson, R.D.; Rapraeger, A.C. Heparanase-induced shedding of syndecan-1/CD138 in myeloma and endothelial cells activates VEGFR2 and an invasive phenotype: Prevention by novel synstatins. Oncogene 2016, 5, e202. [Google Scholar] [CrossRef] [Green Version]
- Vlodavsky, I.; Singh, P.; Boyango, I.; Gutter-Kapon, L.; Elkin, M.; Sanderson, R.D.; Ilan, N. Heparanase: From basic research to therapeutic applications in cancer and inflammation. Drug Resist. Updates 2016, 29, 54–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, C.A.; Purushothaman, A.; Ramani, V.C.; Vlodavsky, I.; Sanderson, R.D. Heparanase regulates secretion, composition, and function of tumor cell-derived exosomes. J. Biol. Chem. 2013, 288, 10093–10099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purushothaman, A.; Bandari, S.K.; Liu, J.; Mobley, J.A.; Brown, E.E.; Sanderson, R.D. Fibronectin on the surface of myeloma cell-derived exosomes mediates exosome-cell interactions. J. Biol. Chem. 2016, 291, 1652–1663. [Google Scholar] [CrossRef] [Green Version]
- Amin, R.; Tripathi, K.; Sanderson, R.D. Nuclear heparanase regulates chromatin remodeling, gene expression and PTEN tumor suppression function. Cells 2020, 9, 2038. [Google Scholar] [CrossRef] [PubMed]
- Vlodavsky, I.; Gross-Cohen, M.; Weissmann, M.; Ilan, N.; Sanderson, R.D. Opposing functions of heparanase-1 and heparanase-2 in cancer progression. Trends Biochem. Sci. 2018, 43, 18–31. [Google Scholar] [CrossRef]
- Daly, S.B.; Urquhart, J.E.; Hilton, E.; McKenzie, E.A.; Kammerer, R.A.; Lewis, M.; Kerr, B.; Stuart, H.; Donnai, D.; Long, D.A.; et al. Mutations in HPSE2 cause urofacial syndrome. Am. J. Hum. Genet. 2010, 86, 963–969. [Google Scholar] [CrossRef] [Green Version]
- Pang, J.; Zhang, S.; Yang, P.; Hawkins-Lee, B.; Zhong, J.; Zhang, Y.; Ochoa, B.; Agundez, J.A.G.; Voelckel, M.-A.; Fisher, R.B.; et al. Loss-of-function mutations in HPSE2 cause the autosomal recessive urofacial syndrome. Am. J. Hum. Genet. 2010, 86, 957–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuart, H.M.; Roberts, N.A.; Hilton, E.N.; McKenzie, E.A.; Daly, S.B.; Hadfield, K.D.; Rahal, J.S.; Gardiner, N.J.; Tanley, S.W.; Lewis, M.A.; et al. Urinary tract effects of HPSE2 mutations. J. Am. Soc. Nephrol. 2015, 26, 797–804. [Google Scholar] [CrossRef] [Green Version]
- Levy-Adam, F.; Cohen-Kaplan, V.; Shteingauz, A.; Gross, M.; Arvatz, G.; Naroditsky, I.; Ilan, N.; Doweck, I.; Vlodavsky, I. Heparanase-2 interacts with heparan sulfate with high affinity and inhibits heparanase activity. J. Biol. Chem. 2010, 285, 28010–28019. [Google Scholar] [CrossRef] [Green Version]
- Pinhal, M.A.S.; Melo, C.M.; Nader, H.B. The good and bad sides of heparanase-1 and heparanase-2. Adv. Exp. Med. Biol. 2020, 1221, 821–845. [Google Scholar] [CrossRef] [PubMed]
- Gross-Cohen, M.; Yanku, Y.; Kessler, O.; Barash, U.; Boyango, I.; Cid-Arregui, A.; Neufeld, G.; Ilan, N.; Vlodavsky, I. Heparanase 2 (Hpa2) attenuates tumor growth by inducing Sox2 expression. Matrix Biol. 2021, 99, 58–71. [Google Scholar] [CrossRef]
- Liu, J.; Knani, I.; Gross-Cohen, M.; Hu, J.; Wang, S.; Tang, L.; Ilan, N.; Yang, S.; Vlodavsky, I. Role of heparanase 2 (Hpa2) in gastric cancer. Neoplasia 2021, 23, 966–978. [Google Scholar] [CrossRef]
- Kayal, Y.; Singh, P.; Naroditsky, I.; Ilan, N.; Vlodavsky, I. Heparanase 2 (Hpa2) attenuates the growth of pancreatic carcinoma. Matrix Biol. 2021, 98, 21–31. [Google Scholar] [CrossRef]
- Knani, I.; Yanku, Y.; Gross-Cohen, M.; Ilan, N.; Vlodavsky, I. Heparanase 2 (Hpa2) attenuates the growth of human sarcoma. Matrix Biol. 2022, 113, 22–38. [Google Scholar] [CrossRef]
- Knani, I.; Singh, P.; Gross-Cohen, M.; Aviram, S.; Ilan, N.; Sanderson, R.D.; Aronheim, A.; Vlodavsky, I. Induction of heparanase 2 (Hpa2) expression by stress is mediated by ATF3. Matrix Biol. 2022, 105, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Ostrovsky, O.; Grushchenko-Polaq, A.H.; Beider, K.; Mayorov, M.; Canaani, J.; Shimoni, A.; Vlodavsky, I.; Nagler, A. Identification of strong intron enhancer in the heparanase gene: Effect of functional rs4693608 variant on HPSE enhancer activity in hematological and solid malignancies. Oncogenesis 2018, 7, 51–65. [Google Scholar] [CrossRef] [Green Version]
- Ostrovsky, O.; Baryakh, P.; Morgulis, Y.; Mayorov, M.; Bloom, N.; Beider, K.; Shimoni, A.; Vlodavsky, I.; Nagler, A. The HPSE gene insulator—A novel regulatory element that affects heparanase expression, stem cell mobilization and the risk of acute graft versus host disease. Cells 2021, 10, 2523. [Google Scholar] [CrossRef]
- Ostrovsky, O.; Korostishevsky, M.; Shafat, I.; Mayorov, M.; Ilan, N.; Vlodavsky, I.; Nagler, A. Inverse correlation between HPSE gene single nucleotide polymorphisms (SNPs) and heparanase expression: Possibility of multiple levels of heparanase regulation. J. Leukoc. Biol. 2009, 86, 445–455. [Google Scholar] [CrossRef] [Green Version]
- Ostrovsky, O.; Shimoni, A.; Baryakh, P.; Morgulis, Y.; Mayorov, M.; Beider, K.; Shteingauz, A.; Ilan, N.; Vlodavsky, I.; Nagler, A. Modification of heparanase gene expression in response to conditioning and LPS treatment: Strong correlation to rs4693608 SNP. J. Leukoc. Biol. 2014, 95, 677–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostrovsky, O.; Shimoni, A.; Rand, A.; Vlodavsky, I.; Nagler, A. Genetic variations in the heparanase gene (HPSE) associate with increased risk of GVHD following allogeneic stem cell transplantation: Effect of discrepancy between recipients and donors. Blood 2010, 115, 2319–2328. [Google Scholar] [CrossRef] [Green Version]
- Ostrovsky, O.; Beider, K.; Morgulis, Y.; Bloom, N.; Cid-Arregui, A.; Shimoni, A.; Vlodavsky, I.; Nagler, A. CMV seropositivity status increases heparanase SNPs regulatory activity, risk of acute GVHD and yield of CD34+ cell mobilization. Cells 2021, 10, 3489. [Google Scholar] [CrossRef]
- Ostrovsky, O.; Korostishevsky, M.; Levite, I.; Leiba, M.; Galski, H.; Vlodavsky, I.; Nagler, A. Association of heparanase gene (HPSE) single nucleotide polymorphisms with hematological malignancies. Leukemia 2007, 21, 2296–2303. [Google Scholar] [CrossRef] [PubMed]
- D’Agostino, M.; Cairns, D.A.; Lahuerta, J.J.; Wester, R.; Bertsch, U.; Waage, A.; Zamagni, E.; Mateos, M.V.; Dall’Olio, D.; van de Donk, N.W.C.J.; et al. Second revision of the International Staging System (R2-ISS) for overall survival in multiple myeloma: A European Myeloma Network (EMN) report within the HARMONY Project. J. Clin. Oncol. 2022, 40, 3406–3418. [Google Scholar] [CrossRef] [PubMed]
- Carlton, V.E.H.; Ireland, J.S.; Useche, F.; Feham, M. Functional single nucleotide polymorphism-based associated studies. Human Genom. 2006, 2, 391–402. [Google Scholar] [CrossRef] [Green Version]
- Walavalkar, K.; Notani, D. Beyond the coding genome: Non-coding mutations and cancer. Front. Biosci. 2020, 25, 1828–1838. [Google Scholar] [CrossRef]
- Fullard, J.F.; Rahman, S.; Roussos, P. Genetic variation in long-range enhancers. Curr. Top. Behav. Neurosci. 2019, 42, 35–50. [Google Scholar] [CrossRef]
- Matharu, N.; Ahituv, N. Modulating gene regulation to treat genetic disorders. Nat. Rev. Drug Discov. 2020, 19, 757–775. [Google Scholar] [CrossRef]
- Karamanos, N.K.; Piperigkou, Z.; Passi, A.; Gotte, M.; Rousselle, P.; Vlodavsky, I. Extracellular matrix-based cancer targeting. Trends Mol. Med. 2021, 27, 1000–1013. [Google Scholar] [CrossRef] [PubMed]
- Kiyan, Y.; Tkachuk, S.; Kurselis, K.; Shushakova, N.; Stahl, K.; Dawodu, D.; Kiyan, R.; Chichkov, B.; Haller, H. Heparanase-2 protects from LPS-mediated endothelial injury by inhibiting TLR4 signaling. Sci. Rep. 2019, 9, 13591. [Google Scholar] [CrossRef] [Green Version]
- Pape, T.; Hunkemoller, A.M.; Kümpers, P.; Haller, H.; David, S.; Stahl, K. Targeting the “sweet spot” in septic shock—A perspective on the endothelial glycocalyx regulating proteins heparanase-1 and -2. Matrix Biol. Plus 2021, 12, 100095. [Google Scholar] [CrossRef] [PubMed]
- Jayatilleke, K.M.; Hulett, M.D. Heparanase and the hallmarks of cancer. J. Transl. Med. 2020, 18, 453. [Google Scholar] [CrossRef]
- Ilan, N.; Bhattacharya, U.; Barash, U.; Boyango, I.; Yanku, Y.; Gross-Cohen, M.; Vlodavsky, I. Heparanase—The massage comes in different flavors. Adv. Exp. Med. Biol. 2020, 1221, 253–283. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Macleod, V.; Bendre, M.; Huang, Y.; Theus, A.M.; Miao, H.Q.; Kussie, P.; Yaccoby, S.; Epstein, J.; Suva, L.J.; et al. Heparanase promotes the spontaneous metastasis of myeloma cells to bone. Blood 2005, 105, 1303–1309. [Google Scholar] [CrossRef]
- Purushothaman, A.; Chen, L.; Yang, Y.; Sanderson, R.D. Heparanase stimulation of protease expression implicates it as a master regulator of the aggressive tumor phenotype in myeloma. J. Biol. Chem. 2008, 283, 32628–32636. [Google Scholar] [CrossRef] [Green Version]
- Andersen, N.F.; Vogel, U.; Klausen, T.W.; Gimsing, P.; Gregersen, H.; Abildgaard, N.; Vangsted, A.J. Polymorphisms in the heparanase gene in multiple myeloma associated with bone morbidity and survival. Eur. J. Haematol. 2015, 94, 60–66. [Google Scholar] [CrossRef] [Green Version]
- Bansal, R.; Rakshit, S.; Kumar, S. Extramedullary disease in multiple myeloma. Blood Cancer J. 2021, 11, 161. [Google Scholar] [CrossRef]
- Alsayed, Y.; Ngo, H.; Runnels, J.; Leleu, X.; Singha, U.K.; Pitsillides, C.M.; Spencer, J.A.; Kimlinger, T.; Ghobrial, J.M.; Jia, X.; et al. Mechanisms of regulation of CXCR4/SDF-1 (CXCL12)-dependent migration and homing in multiple myeloma. Blood 2007, 109, 2708–2717. [Google Scholar] [CrossRef]
- Beider, K.; Rosenberg, E.; Bitner, H.; Shimoni, A.; Leiba, M.; Koren-Michowitz, M.; Ribakovsky, E.; Klein, S.; Olam, D.; Weiss, L.; et al. The sphingosine-1-phosphate modulator FTY720 targets multiple myeloma via the CXCR4/CXCL12 pathway. Clin. Cancer Res. 2017, 23, 1733–1747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blade, J.; Fernandez de Larrea, C.; Rosinol, L.; Cibeira, M.T.; Jimenez, R.; Powles, R. Soft-tissue plasmacytomas in multiple myeloma: Incidence, mechanisms of extramedullary spread, and treatment approach. J. Clin. Oncol. 2011, 29, 3805–3812. [Google Scholar] [CrossRef]
- Roccaro, A.M.; Mishima, Y.; Sacco, A.; Moschetta, M.; Tai, Y.-T.; Shi, J.; Zhang, Y.; Reagan, M.R.; Huynh, D.; Kawano, Y.; et al. CXCR4 regulates extra-medullary myeloma through epithelial-mesenchymal-transition-like transcriptional activation. Cell Rep. 2015, 12, 622–635. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.L.; Gutter-Kapon, L.; Ilan, N.; Batool, T.; Singh, K.; Digre, A.; Luo, Z.; Sandler, S.; Shaked, Y.; Sanderson, R.D.; et al. Significance of host heparanase in promoting tumor growth and metastasis. Matrix Biol. 2020, 93, 25–40. [Google Scholar] [CrossRef]
- Mayfosh, A.J.; Nguyen, T.K.; Hulett, M.D. The heparanase regulatory network in health and disease. Int. J. Mol. Sci. 2021, 22, 11096. [Google Scholar] [CrossRef] [PubMed]
- He, Y.Q.; Sutcliffe, E.L.; Bunting, K.L.; Li, J.; Goodall, K.J.; Poon, I.K.; Hulett, M.D.; Freeman, C.; McInnes, R.L.; Taya, T.; et al. The endoglycosidase heparanase enters the nucleus of T lymphocytes and modulates H3 methylation at actively transcribed genes via the interplay with key chromatin modifying enzymes. Transcription 2012, 3, 130–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ullah, T.R. The role of CXCR4 in multiple myeloma: Cells’ journey from bone marrow to beyond. J. Bone Oncol. 2019, 17, 100253. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Huang, J.; Zhang, P.; Luo, J.; Cheng, P.; Miao, L.; Lai, Y. Integrated analysis of hub gene expression in multiple myeloma. J. BUON 2021, 26, 2040–2052. [Google Scholar]
- Giuliani, N.; Bonomini, S.; Romagnani, P.; Lazzaretti, M.; Morandi, F.; Colla, S.; Tagliaferri, S.; Lasagni, L.; Annunziato, F.; Crugnola, M.; et al. CXCR3 and its binding chemokines in myeloma cells: Expression of isoforms and potential relationships with myeloma cell proliferation and survival. Haematologica 2006, 91, 1489–1497. [Google Scholar]
- Cannon, W.B. Bodily Changes in Pain, Hunger, Fear and Rage; Appleton-Crofts: New York, NY, USA, 1929. [Google Scholar]
- Goligorsky, M.S. The concept of ce.ellular “fight-or-flight” reaction to stress. Am. J. Physiol. Renal. Physiol. 2001, 280, F551–F561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carpenter, M.B. The autonomic nervous system. In Human Neuroanatomy, 7th ed.; The William & Wilkins Co.: Baltimore, MD, USA, 1976; pp. 191–212. [Google Scholar]
- Hanoun, M.; Maryanovich, M.; Arnal-Estape, A.; Frenette, P.S. Neural regulation of hematopoiesis, inflammation and cancer. Neuron 2015, 86, 360–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roucourt, B.; Meeussen, S.; Bao, J.; Zimmermann, P.; David, G. Heparanase activates the syndecan-syntenin-ALIX exosome pathway. Cell Res. 2015, 25, 412–428. [Google Scholar] [CrossRef] [Green Version]
- Ruan, J.; Trotter, T.N.; Nan, L.; Luo, R.; Javed, A.; Sanderson, R.D.; Suva, L.J.; Yang, Y. Heparanase inhibits osteoblastogenesis and shifts bone marrow progenitor cell fate in myeloma bone disease. Bone 2013, 57, 10–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Variable | Characteristic | No. of Cases (%) |
|---|---|---|
| median—60 | ||
| Age (years) | range—27–90 | |
| <60 | 128 (46.0) | |
| ≥60 | 150 (54.0) | |
| Gender | male | 174 (62.6) |
| female | 104 (37.4) | |
| MM | multiple myeloma | 231(83.1) |
| SMM | smoldering multiple myeloma | 28 (10.1) |
| MGUS | monoclonal gammopathy | 19 (6.8) |
| ISS (international staging system) | I | 39 (14.0) |
| II | 66 (23.7) | |
| III | 46 (16.5) | |
| NA | 127 (45.7) | |
| Beta-2-microglobulin (µg/dL) | median—4.1 | range—1.2–14.8 |
| Creatinine (mg/dL) | median—1.04 | range—0.5–20.0 |
| Albumin (g/dL) | median—3.9 | range—1.9–5.2 |
| Calcium (mg/dL) | median—9.4 | range—7.4–16.2 |
| Hemoglobin (g/dL) | median—11.4 | range—5.2–16.5 |
| Bone morbidity | primary | 149 (69.0) |
| secondary | 13 (6.0) | |
| without bone disease | 54 (25.0) | |
| Extramedullary disease | primary | 50 (23.1) |
| secondary | 39 (18.1) | |
| without EMD | 127 (58.8) | |
| Renal failure | with renal failure | 57 (26.3) |
| without renal failure | 160 (73.7) |
| SNPs/ Genotypes and Alleles | Stage I | Stage II | Stage III | χ2/ p-Value |
|---|---|---|---|---|
| Number/ Incidence | Number/ Incidence | Number/ Incidence | ||
| rs4693608 AA AG GG A G | 6 (15.4%) 23 (59.0%) 10 (25.6%) 35 (44.9%) 43 (55.1%) | 27 (40.9%) 31 (47.0%) 8 (12.0%) 85 (64.4%) 47 (35.6%) | 15 (32.6%) 24 (52.2%) 7 (15.2%) 54 (58.7%) 38 (41.3%) | I − III: 3.86, 0.15 I − II + III: 7.62, 0.022 I + II − III: 0.089, 0.96 I − III: 3.23, 0.072 I − II + III: 6.99, 0.0082 I + II − III: 0.063, 0.8 |
| rs4693084 GG GT TT G T | 17 (43.6%) 18 (46.2%) 4 (10.3 %) 52 (66.7%) 26 (33.3%) | 44 (66.7%) 19 (28.8%) 3 (4.5%) 107 (81.1%) 25 (18.9%) | 30 (65.2%) 13 (28.3%) 3 (6.5%) 73 (79.3%) 19 (20.7%) | I − III: 4.0, 0.14 I − II + III: 6.175, 0.046 I + II − III: 0.74, 0.69 I − III: 3.49, 0.062 I − II + III: 6.09, 0.014 I + II − III: 0.47, 0.49 |
| rs4426765 AA AC CC A C | 25 (64.1%) 8 (20.5%) 6 (15.4%) 58 (74.4%) 20 (25.6%) | 33 (50.0%) 29 (43.9%) 4 (6.1%) 95 (72.0%) 37 (28.0%) | 29 (63.0%) 16 (34.8%) 1 (2.2%) 74 (80.4%) 18 (19.6%) | I − III: 6.0, 0.05 I − II + III: 8.31, 0.016 I + II − III: 2.71, 0.26 I − III: 0.9, 0.34 I − II + III: 0.037, 0.85 I + II − III: 1.97, 0.16 |
| rs28649799 AA AG GG A G | 30 (76.9%) 9 (23.1%) 0 69 (88.5%) 9 (11.5%) | 58 (87.9%) 7 (10.6%) 1 (1.5%) 123 (93.2%) 9 (6.8%) | 35 (76.1%) 9 (19.6%) 2 (4.3%) 79 (85.9%) 13 (14.1%) | I − III: 1.82, 0.4 I − II + III: 2.53, 0.28 I + II − III: 2.45, 0.29 I − III: 0.25, 0.62 I − II + III: 0.19, 0.67 I + II − III: 2.15, 0.14 |
| rs4364254 TT TC CC T C | 20 (51.3%) 15 (38.5%) 4 (10.3%) 55 (70.5%) 23 (29.5%) | 33 (50.0%) 26 (37.9%) 8 (12.1%) 91 (68.9%) 41 (31.1%) | 23 (50.0%) 18 (39.1%) 5 (10.9%) 64 (69.6%) 28 (30.4%) | I − III: 0.017, 0.99 I − II + III: 0.059, 0.97 I + II − III: 0.008, 0.99 I − III: 0.018, 0.89 I − II + III: 0.047, 0.83 I + II − III: 0, 1 |
| SNPs/ Genotypes and Alleles | Total EMD | Paraskeletal Disease | ||||
|---|---|---|---|---|---|---|
| Yes Number/ Incidence | No Number/ Incidence | χ2/ p-Value | Yes Number/ Incidence | No Number/ Incidence | χ2/ p-Value | |
| rs4693608 | ||||||
| AA | 9 (18.0%) | 59 (36.4%) | 11.98, | 8 (18.6%) | 59 (36.4%) | 8.07, |
| AG | 25 (50.0%) | 82 (50.6%) | 0.003 | 23 (53.5%) | 82 (50.6%) | 0.018 |
| GG | 16 (32.0%) | 21 (13%) | 12 (27.9%) | 21 (13.0%) | ||
| A | 43 (43.0%) | 200 (61.7%) | 10.96, | 39 (45.3%) | 200 (61.7%) | 7.5, |
| G | 57 (57.0%) | 124 (38.3%) | 0.001 | 47 (54.7%) | 124 (38.3%) | 0.006 |
| rs4693084 | ||||||
| GG | 25 (50.0%) | 107 (66.5%) | 8.65, | 22 (51.2%) | 107 (66.5%) | 5.76, |
| GT | 18 (36.0%) | 48 (29.8%) | 0.013 | 16 (37.2%) | 48 (29.8%) | 0.056 |
| TT | 7 (14.0%) | 6 (3.7%) | 5 (11.6%) | 6 (3.7%) | ||
| G | 68 (68.0%) | 262 (81.1%) | 7.996, | 60 (69.8%) | 262 (81.1%) | 5.49, |
| T | 32 (32.0%) | 60 (18.6%) | 0.005 | 26 (30.2%) | 60 (18.6%) | 0.019 |
| rs4426765 | ||||||
| AA | 23 (46.0%) | 103 (63.6%) | 4.94, | 18 (41.9%) | 103 (63.6%) | 6.77, |
| AC | 22 (44.0%) | 49 (30.2%) | 0.085 | 20 (46.5%) | 49 (30.2%) | 0.034 |
| CC | 5 (10.0%) | 10 (6.2%) | 5 (11.6%) | 10 (6.2%) | ||
| A | 68 (68.0%) | 255 (78.7%) | 4.82, | 56 (65.1%) | 255 (78.7%) | 6.85, |
| C | 32 (32.0%) | 69 (21.3%) | 0.028 | 30 (34.9%) | 69 (21.3%) | 0.009 |
| rs28649799 | ||||||
| AA | 35 (70.0%) | 135 (83.3%) | 4.33, | 32 (74.4%) | 135 (83.3%) | 3.44, |
| AG | 13 (26.0%) | 24 (14.8%) | 0.12 | 11 (25.6%) | 24 (14.8%) | 0.18 |
| GG | 2 (4.0%) | 3 (1.9%) | 0 | 3 (1.9%) | ||
| A | 83 (83.0%) | 294 (90.7%) | 4.65, | 75 (87.2%) | 294 (90.7%) | 0.94, |
| G | 17 (17.0%) | 30 (9.3%) | 0.031 | 11 (12.8%) | 30 (9.3%) | 0.33 |
| rs4364254 | ||||||
| TT | 17 (34.0%) | 97 (59.9%) | 11.42, | 16 (37.2%) | 97 (59.9%) | 7.92, |
| TC | 23 (46.0%) | 51 (31.5%) | 0.003 | 19 (44.2%) | 51 (31.5%) | 0.019 |
| CC | 10 (20.0%) | 14 (8.6%) | 8 (18.6%) | 14 (8.6%) | ||
| T | 57 (57.0%) | 245 (75.6%) | 12.92, | 51 (59.3%) | 245 (75.6%) | 9.01, |
| C | 43 (43.0%) | 79 (24.4%) | 0.00032 | 35 (40.7%) | 79 (24.4%) | 0.003 |
| rs4693608 | ||||||
| rs4364254 | ||||||
| HR | 9 (18.0%) | 59 (36.4%) | 17.99, | 8 (18.6%) | 59 (36.4%) | 14.32, |
| MR | 19 (38.0%) | 77 (47.5%) | 0.00012 | 17 (39.5%) | 77 (47.5%) | 0.00078 |
| LR | 22 (44.0%) | 26 (16.1%) | Yc: 16.036, 0.00033 | 18 (41.9%) | 26 (16.1%) | Yc: 12.472, 0.002 |
| SNPs/ Genotypes and Alleles | Total EMD | Paraskeletal Disease | ||||
|---|---|---|---|---|---|---|
| Yes Number/ Incidence | No Number/ Incidence | χ2/ p-Value | Yes Number/ Incidence | No Number/ Incidence | χ2/ p-Value | |
| rs2040022 | ||||||
| AA | 19 (38%) | 57 (35.2%) | NS | 15 (34.9%) | 57 (35.2%) | NS |
| AG | 21 (42%) | 81 (50.0%) | 19 (44.2%) | 81 (50.0%) | ||
| GG | 10 (20%) | 24 (14.8%) | 9 (20.9%) | 24 (14.8%) | ||
| A | 59 (59%) | 195 (60.2%) | NS | 49 (57.0%) | 195 (60.2%) | NS |
| G | 41 (41%) | 129 (39.8%) | 37 (43.0%) | 129 (39.8%) | ||
| rs658225 | ||||||
| AA | 32 (64%) | 110 (67.9%) | NS | 26 (60.5%) | 110 (67.9%) | NS |
| AG | 15 (30%) | 50 (30.9%) | 15 (34.9%) | 50 (30.9%) | ||
| GG | 3 (6%) | 2 (1.2%) | 2 (4.7%) | 2 (1.2%) | ||
| A | 79 (79%) | 270 (83.3%) | NS | 67 (77.9%) | 270 (83.3%) | NS |
| G | 21 (21%) | 54 (16.7%) | 19 (22.1%) | 54 (6.7%) | ||
| rs7081116 | ||||||
| AA | 30 (60%) | 99 (61.9%) | NS | 25 (58.1%) | 99 (61.9%) | NS |
| AG | 17 (34%) | 55 (34.4%) | 15 (34.9%) | 55 (34.4%) | ||
| GG | 3 (6%) | 6 (3.8%) | 3 (7.0%) | 6 (3.8%) | ||
| A | 77 (77%) | 253 (79.1%) | NS | 65 (75.6%) | 253 (79.1%) | NS |
| G | 23 (23%) | 67 (20.9%) | 21 (24.4%) | 67 (20.9%) | ||
| rs861399 | ||||||
| TT | 36 (72%) | 111 (68.5%) | NS | 31 (72.1%) | 111 (68.5%) | NS |
| TC | 13 (26%) | 49 (30.2%) | 11 (25.6%) | 49 (30.2%) | ||
| CC | 2 (1%) | 2 (1.2%) | 1 (2.3%) | 2 (1.2%) | ||
| T | 85 (85%) | 271 (83.6%) | NS | 73 (84.9%) | 271 (83.6%) | NS |
| C | 15 (15%) | 53 (16.4%) | 13 (15.1%) | 53 (16.4%) | ||
| rs7076702 | ||||||
| CC | 29 (58%) | 95 (58.6%) | NS | 24 (55.8%) | 95 (58.6%) | NS |
| CG | 18 (36%) | 59 (36.4%) | 16 (37.2%) | 59 (36.4%) | ||
| GG | 3 (6%) | 8 (4.9%) | 3 (7.0%) | 8 (4.9%) | ||
| C | 76 (76%) | 249 (76.9%) | NS | 64 (74.4%) | 249 (76.9%) | NS |
| G | 24 (24%) | 75 (23.1%) | 22 (25.6%) | 75 (23.1%) | ||
| rs475711 | ||||||
| CC | 28 (56%) | 101 (62.3%) | NS | 23 (53.5%) | 101 (62.3%) | NS |
| CT | 19 (38%) | 56 (34.6%) | 17 (39.5%) | 56 (34.6%) | ||
| TT | 3 (6%) | 5 (3.1%) | 3 (7.0%) | 5 (3.1%) | ||
| C | 75 (75%) | 258 (79.6%) | NS | 63 (73.3%) | 258 (79.6%) | NS |
| T | 25 (25%) | 66 (20.4%) | 23 (26.7%) | 66 (20.4%) | ||
| rs657442 | ||||||
| CC | 50 (100%) | 140 (86.4%) | 0.0065 | 43 (100%) | 140 (86.4%) | 0.011 |
| CA | 0 | 21 (13.0%) | 0 | 21 (13.0%) | ||
| AA | 0 | 1 (0.6%) | 0 | 1 (0.6%) | ||
| C | 100 (100%) | 301 (92.9%) | 0.0038 | 86 (100%) | 301 (92.9%) | 0.0066 |
| A | 0 | 23 (7.1%) | 0 | 23 (7.1%) | ||
| rs57455933 | ||||||
| AA | 26 (52%) | 75 (46.3%) | NS | 23 (53.5%) | 75 (46.3%) | NS |
| AT | 21 (42%) | 68 (42.0%) | 18 (41.7%) | 68 (42.0%) | ||
| TT | 3 (6%) | 19 (11.7%) | 2 (4.7%) | 19 (11.7%) | ||
| A | 73 (73%) | 218 (67.3%) | NS | 64 (74.4%) | 218 (67.3%) | NS |
| T | 27 (27%) | 106 (32.7%) | 22 (25.6%) | 106 (32.7%) | ||
| rs621644 | ||||||
| GG | 25 (50%) | 85 (52.8%) | NS | 22 (51.2%) | 85 (52.8%) | NS |
| GA | 22 (44%) | 65 (40.4%) | 19 (44.2%) | 65 (40.4%) | ||
| AA | 3 (6%) | 11 (6.8%) | 2 (4.7%) | 11 (6.8%) | ||
| G | 72 (72%) | 235 (73%) | NS | 63 (73.3%) | 235 (73%) | NS |
| A | 28 (28%) | 87 (27%) | 23 (26.7%) | 87 (27%) | ||
| rs1998756 | ||||||
| CC | 20 (40%) | 74 (45.7%) | NS | 18 (41.9%) | 74 (45.7%) | NS |
| CT | 24 (48%) | 71 (43.8%) | 20 (46.5%) | 71 (43.8%) | ||
| TT | 6 (12%) | 17 (10.5%) | 5 (11.6%) | 17 (10.5%) | ||
| C | 64 (64%) | 219 (67.6%) | NS | 56 (65.1%) | 219 (67.6%) | NS |
| T | 36 (36%) | 105 (32.4%) | 30 (34.9%) | 105 (32.4%) | ||
| rs754585 | ||||||
| CC | 10 (20%) | 39 (24.1%) | NS | 8 (18.6%) | 39 (24.1%) | NS |
| CT | 29 (58%) | 77 (47.5%) | 27 (62.8%) | 77 (47.5%) | ||
| TT | 11 (22%) | 46 (28.4%) | 8 (18.6%) | 46 (28.4%) | ||
| C | 49 (49%) | 155 (47.8%) | NS | 43 (50.0%) | 155 (47.8%) | NS |
| T | 51 (51%) | 169 (52.2%) | 43 (50.0%) | 169 (52.2%) | ||
| Genotypes | Yes Number/Incidence | No Number/Incidence | χ2/ p-Value |
|---|---|---|---|
| HR-CC | 9 (18.0%) | 50 (30.9%) | |
| HR-CA | 0 | 8 (4.9%) | |
| HR-AA | 0 | 1 (0.6%) | |
| MR-CC | 19 (38.0%) | 69 (42.6%) | |
| MR-CA | 0 | 8 (4.9%) | |
| MR-AA | 0 | 0 | |
| LR-CC | 22 (44.0%) | 21 (13.0) | |
| LR-CA | 0 | 5 (3.1%) | |
| LR-AA | 0 | 0 | |
| Group A1 Group A2 Group A3 | 9 (18.0%) 19 (38.0%) 22 (44.0%) | 67 (41.4%) 74 (45.7%) 21 (12.9%) | χ2df2 = 22.47, p = 0.0000049 Yates’ χ2 = 22.18, Yates’ p = 0.000015 |
| Group B1 Group B2 Group B3 | 9 (18.0%) 19 (38.0%) 22 (44.0%) | 72 (44.4%) 69 (42.6%) 21 (13.0%) | χ2df2 = 25.33, p = 0.0000036 Yates’ χ2 = 23.065, Yates’ p = 0.0000098 |
| SNPs/ Genotypes and Alleles | Total EMD | Paraskeletal Disease | ||||
|---|---|---|---|---|---|---|
| Yes Number/ Incidence | No Number/ Incidence | χ2/ p-Value | Yes Number/ Incidence | No Number/ Incidence | χ2/ p-Value | |
| rs2040022 | ||||||
| AA | 15 (40.5%) | 41 (34.2%) | NS | 9 (32.1%) | 41 (34.2%) | NS |
| AG | 16 (43.2%) | 61 (50.8%) | 14 (50.0%) | 61 (50.8%) | ||
| GG | 6 (16.2%) | 18 (15.0%) | 5 (17.9%) | 18 (15.0%) | ||
| A | 46 (62.1%) | 143 (59.6%) | NS | 32 (57.1%) | 143 (59.6%) | NS |
| G | 28 (37.8%) | 97 (40.4%) | 24 (42.9%) | 97 (40.4%) | ||
| rs658225 | ||||||
| AA | 31 (83.8%) | 77 (64.2%) | 5.24, | 23 (82.1%) | 77 (64.2%) | 3.5, |
| AG | 6 (16.2%) | 41 (34.2%) | 0.073 | 5 (17.9%) | 41 (34.2%) | 0.17 |
| GG | 0 | 2 (1.7%) | 0 | 2 (1.7%) | ||
| A | 68 (91.9%) | 195 (81.3%) | 4.71, | 51 (91.1%) | 195 (81.3%) | 3.12, |
| G | 6 (8.1%) | 45 (18.7%) | 0.03 | 5 (8.9%) | 45 (18.7%) | 0.077 |
| rs7081116 | ||||||
| AA | 24 (64.9%) | 72 (61.0%) | NS | 16 (57.1%) | 72 (61.0%) | NS |
| AG | 11 (29.7%) | 42 (35.6%) | 11 (39.3%) | 42 (35.6%) | ||
| GG | 2 (5.4%) | 4 (3.4%) | 1 (3.6%) | 4 (3.4%) | ||
| A | 59 (79.7%) | 186 (78.8%) | NS | 43 (76.8%) | 186 (78.8%) | NS |
| G | 15 (20.3%) | 50 (21.2%) | 13 (23.2%) | 50 (21.2%) | ||
| rs861399 | ||||||
| TT | 25 (67.6%) | 82 (68.3%) | NS | 17 (60.7%) | 82 (68.3%) | NS |
| TC | 11 (29.7%) | 37 (30.8%) | 11 (39.3%) | 37 (30.8%) | ||
| CC | 1 (2.7%) | 1 (0.8%) | 0 | 1 (0.8%) | ||
| T | 61 (82.4%) | 201 (83.7%) | NS | 45 (80.4%) | 201 (83.7%) | NS |
| C | 13 (17.6%) | 39 (16.3%) | 11 (19.6%) | 39 (16.3%) | ||
| rs7076702 | ||||||
| CC | 24 (64.9%) | 67 (55.8%) | NS | 16 (57.1%) | 67 (55.8%) | NS |
| CG | 11 (29.7%) | 47 (39.2%) | 11 (39.3%) | 47 (39.2%) | ||
| GG | 2 (5.4%) | 6 (5.0%) | 1 (3.6%) | 6 (5.0%) | ||
| C | 59 (79.7%) | 181 (75.4%) | NS | 43 (76.8%) | 181 (75.4%) | NS |
| G | 15 (20.3%) | 59 (24.6%) | 13 (23.2%) | 59 (24.6%) | ||
| rs475711 | ||||||
| CC | 23 (62.2%) | 75 (62.5%) | NS | 16 (57.1%) | 75 (62.5%) | NS |
| CT | 13 (35.1%) | 41 (34.2%) | 12 (42.9%) | 41 (34.2%) | ||
| TT | 1 (2.7%) | 4 (3.3%) | 0 | 4 (3.3%) | ||
| C | 59 (79.7%) | 191 (79.6%) | NS | 44 (78.6%) | 191 (79.6%) | NS |
| T | 15 (20.3%) | 49 (20.4%) | 12 (21.4%) | 49 (20.4%) | ||
| rs657442 | ||||||
| CC | 34 (91.9%) | 102 (85.0%) | NS | 26 (92.9%) | 102 (85.0%) | NS |
| CA | 3 (8.1%) | 17 (14.2%) | 2 (7.1%) | 17 (14.2%) | ||
| AA | 0 | 1 (0.8%) | 0 | 1 (0.8%) | ||
| C | 71 (95.9%) | 221 (92.1%) | NS | 54 (96.4%) | 221 (92.1%) | NS |
| A | 3 (4.1%) | 19 (7.9%) | 2 (3.6%) | 19 (7.9%) | ||
| rs57455933 | ||||||
| AA | 20 (54.1%) | 53 (44.2%) | NS | 14 (50.0%) | 53 (44.2%) | NS |
| AT | 11 (29.7%) | 54 (45.0%) | 8 (28.6%) | 54 (45.0%) | ||
| TT | 6 (16.2%) | 13 (10.8%) | 6 (21.4%) | 13 (10.8%) | ||
| A | 51 (68.9%) | 160 (66.7%) | NS | 36 (64.3%) | 160 (66.7%) | NS |
| T | 23 (31.1%) | 106 (33.3%) | 20 (35.7%) | 106 (33.3%) | ||
| rs621644 | ||||||
| GG | 20 (54.1%) | 62 (52.1%) | 7.16, | 13 (46.4%) | 62 (52.1%) | 9.9, |
| GA | 11 (29.7%) | 52 (43.7%) | 0.028 | 9 (32.1%) | 52 (43.7%) | 0.007 |
| AA | 6 (16.2%) | 5 (4.2%) | 6 (21.4%) | 5 (4.2%) | ||
| G | 51 (68.9%) | 176 (73.9%) | NS | 35 (62.5%) | 176 (73.9%) | 2.93, |
| A | 23 (31.1%) | 62 (26.1%) | 21 (37.5%) | 62 (26.1%) | 0.087 | |
| rs1998756 | ||||||
| CC | 18 (48.6%) | 54 (45.0%) | NS | 14 (50.0%) | 54 (45.0%) | NS |
| CT | 15 (40.5%) | 54 (45.0%) | 11 (39.3%) | 54 (45.0%) | ||
| TT | 4 (10.8%) | 12 (10.0%) | 3 (10.7%) | 12 (10.0%) | ||
| C | 51 (68.9%) | 162 (67.5%) | NS | 39 (69.6%) | 162 (67.5%) | NS |
| T | 23 (31.1%) | 78 (32.5%) | 17 (30.4%) | 78 (32.5%) | ||
| rs754585 | ||||||
| CC | 8 (21.6%) | 30 (25.0%) | 7.24, | 6 (21.4%) | 30 (25.0%) | 5.42, |
| CT | 24 (64.9%) | 50 (41.7%) | 0.027 | 18 (64.3%) | 50 (41.7%) | 0.067 |
| TT | 5 (13.5%) | 40 (33.3%) | 4 (14.3%) | 40 (33.3%) | ||
| C | 40 (54.1%) | 155 (45.8%) | NS | 30 (53.6%) | 155 (45.8%) | NS |
| T | 34 (45.9%) | 169 (54.2%) | 26 (46.4%) | 169 (54.2%) | ||
| Genotypes | Secondary EMD Number/Incidence | Total EMD Number/Incidence | No Number/Incidence |
|---|---|---|---|
| AA-CC | 8 (21.1%) | 13 (14.9%) | 22 (18.3%) |
| AA-CT | 20 (52.6%) | 40 (46.0%) | 28 (23.3%) |
| AA-TT | 4 (10.5%) | 10 (11.5%) | 27 (22.5%) |
| AG-CC | 1 (2.6%) | 4 (4.6%) | 8 (6.7%) |
| AG-CT | 4 (10.5%) | 12 (13.8%) | 22 (18.3%) |
| AG-TT | 1 (2.6%) | 5 (5.8%) | 11 (9.2%) |
| GG-CC | 0 | 1 (1.1) | 0 |
| GG-CT | 0 | 1 (1.1) | 0 |
| GG-TT | 0 | 1 (1.1) | 2 (1.7%) |
| SNPs | Genotypes and Alleles | Yes Number/Incidence | No Number/Incidence | χ2/ p-Value |
|---|---|---|---|---|
| rs2040022 | AA | 56 (35.4%) | 17 (32.1%) | NS NS |
| AG | 78 (49.4%) | 26 (49.0%) | ||
| GG | 24 (15.2%) | 10 (18.9%) | ||
| A | 190 (60.1%) | 60 (56.6%) | ||
| G | 126 (39.9%) | 46 (43.4%) | ||
| rs658225 | AA | 109 (69.0%) | 34 (64.2%) | NS NS |
| AG | 44 (27.8%) | 19 (35.8%) | ||
| GG | 5 (3.2%) | 0 | ||
| A | 262 (82.9%) | 87 (82.1%) | ||
| G | 54 (17.1%) | 19 (17.9%) | ||
| rs7081116 | AA | 91 (58.3%) | 35 (66.0%) | NS NS |
| AG | 56 (35.9%) | 18 (34.0%) | ||
| GG | 9 (5.8%) | 0 | ||
| A | 238 (76.3%) | 88 (83.0%) | ||
| G | 74 (23.7%) | 18 (17.0%) | ||
| rs861399 | TT | 104 (65.8%) | 40 (75.5%) | NS NS |
| TC | 51 (32.3%) | 13 (24.5%) | ||
| CC | 3 (1.9%) | 0 | ||
| T | 259 (82.0%) | 93 (87.3%) | ||
| C | 57 (18.0%) | 13 (12.3%) | ||
| rs7076702 | CC | 87 (55.1%) | 34 (64.2%) | NS NS |
| CG | 61 (38.6%) | 18 (34.0%) | ||
| GG | 10 (6.3%) | 1 (1.9%) | ||
| C | 235 (74.4%) | 86 (81.1%) | ||
| G | 81 (25.6%) | 20 (18.9%) | ||
| rs475711 | CC | 90 (57.0%) | 36 (67.9%) | NS NS |
| CT | 60 (38.0%) | 17 (32.1%) | ||
| TT | 8 (5.0%) | 0 | ||
| C | 240 (76.0%) | 89 (84.0%) | ||
| T | 76 (24.0%) | 17 (16.0%) | ||
| rs657442 | CC | 149 (94.3%) | 41 (77.4%) | 13.74, |
| CA | 9 (5.7%) | 11 (20.8%) | 0.001 | |
| AA | 0 | 1 (1.9%) | ||
| C | 307 (97.2%) | 93 (87.7%) | 14.24, | |
| A | 9 (2.8%) | 13 (12.3%) | 0.00016 | |
| rs57455933 | AA | 77 (48.7%) | 24 (45.3%) | |
| AT | 67 (42.4%) | 20 (37.7%) | NS | |
| TT | 14 (8.9%) | 9 (17.0%) | ||
| A | 221 (69.9%) | 68 (64.2%) | ||
| T | 95 (30.1%) | 38 (35.4%) | NS | |
| rs621644 | GG | 81 (51.7%) | 28 (52.8%) | |
| GA | 64 (40.8%) | 22 (41.5%) | NS | |
| AA | 12 (7.6%) | 3 (5.7%) | ||
| G | 226 (72.0%) | 78 (73.6%) | ||
| A | 88 (28.0%) | 28 (26.4%) | NS | |
| rs1998756 | CC | 74 (46.8%) | 20 (37.7%) | |
| CT | 66 (41.8%) | 27 (51.0%) | NS | |
| TT | 18 (11.4%) | 6 (11.3%) | ||
| C | 214 (67.7%) | 67 (63.2%) | ||
| T | 102 (32.3%) | 39 (36.8%) | NS | |
| rs754585 | CC | 37 (23.4%) | 13 (24.5%) | |
| CT | 80 (50.6%) | 25 (47.2%) | NS | |
| TT | 41 (25.9%) | 15 (28.3%) | ||
| C | 154 (48.7%) | 51 (48.1%) | ||
| T | 162 (51.3%) | 55 (51.9%) | NS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ostrovsky, O.; Beider, K.; Magen, H.; Leiba, M.; Sanderson, R.D.; Vlodavsky, I.; Nagler, A. Effect of HPSE and HPSE2 SNPs on the Risk of Developing Primary Paraskeletal Multiple Myeloma. Cells 2023, 12, 913. https://doi.org/10.3390/cells12060913
Ostrovsky O, Beider K, Magen H, Leiba M, Sanderson RD, Vlodavsky I, Nagler A. Effect of HPSE and HPSE2 SNPs on the Risk of Developing Primary Paraskeletal Multiple Myeloma. Cells. 2023; 12(6):913. https://doi.org/10.3390/cells12060913
Chicago/Turabian StyleOstrovsky, Olga, Katia Beider, Hila Magen, Merav Leiba, Ralph D. Sanderson, Israel Vlodavsky, and Arnon Nagler. 2023. "Effect of HPSE and HPSE2 SNPs on the Risk of Developing Primary Paraskeletal Multiple Myeloma" Cells 12, no. 6: 913. https://doi.org/10.3390/cells12060913