
Pregestational Exposure to T. gondii Produces Maternal Antibodies That Recognize Fetal Brain Mimotopes and Induces Neurochemical and Behavioral Dysfunction in the Offspring
 , , , , , , , ,
, , , , , , , ,  and
and 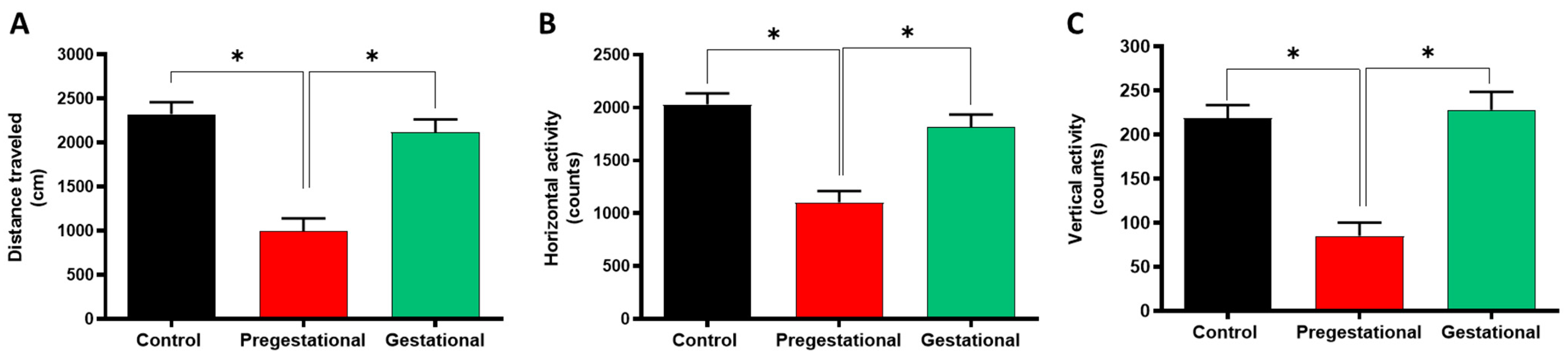{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Materials
2.3. Toxoplasma Gondii Lysate
2.4. Rat Immunization
2.5. Anti-T. gondii IgG Seropositivity
2.6. Behavioral and Cognitive Test
2.6.1. Locomotor Activity
2.6.2. Novel Object Recognition (NOR) Test
2.6.3. Crawley’s Test
2.7. Dopamine and Glutamate Determination
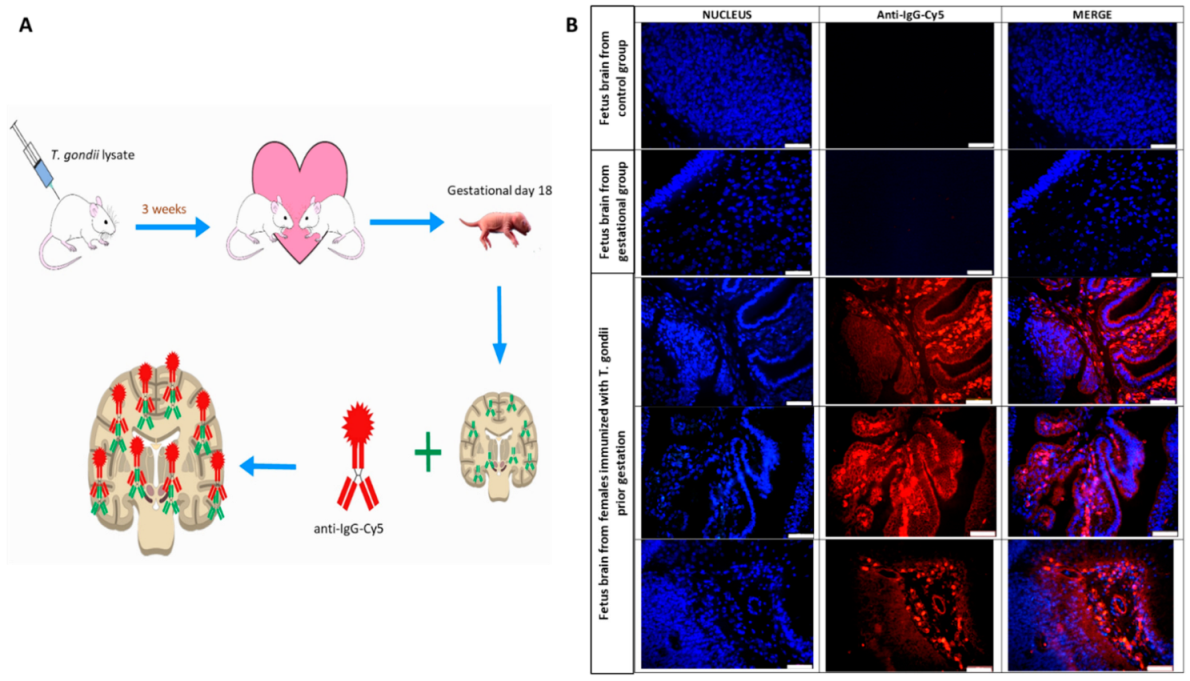
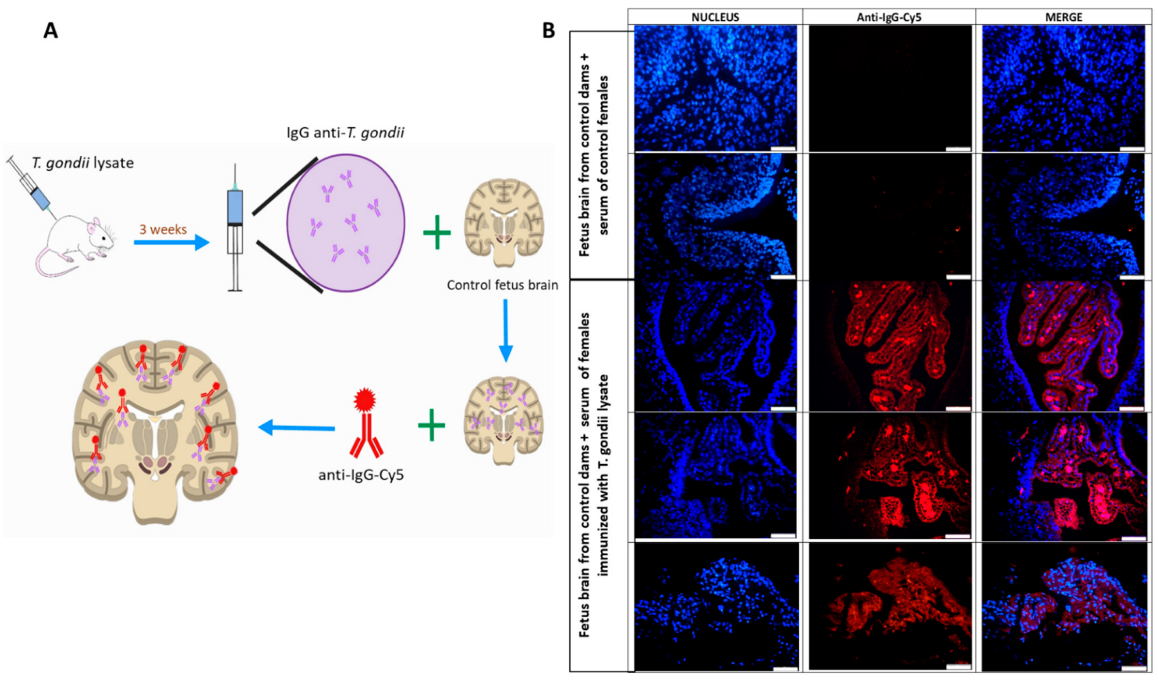
2.8. Maternal Pathogenic Antibodies Identification by Immunofluorescence
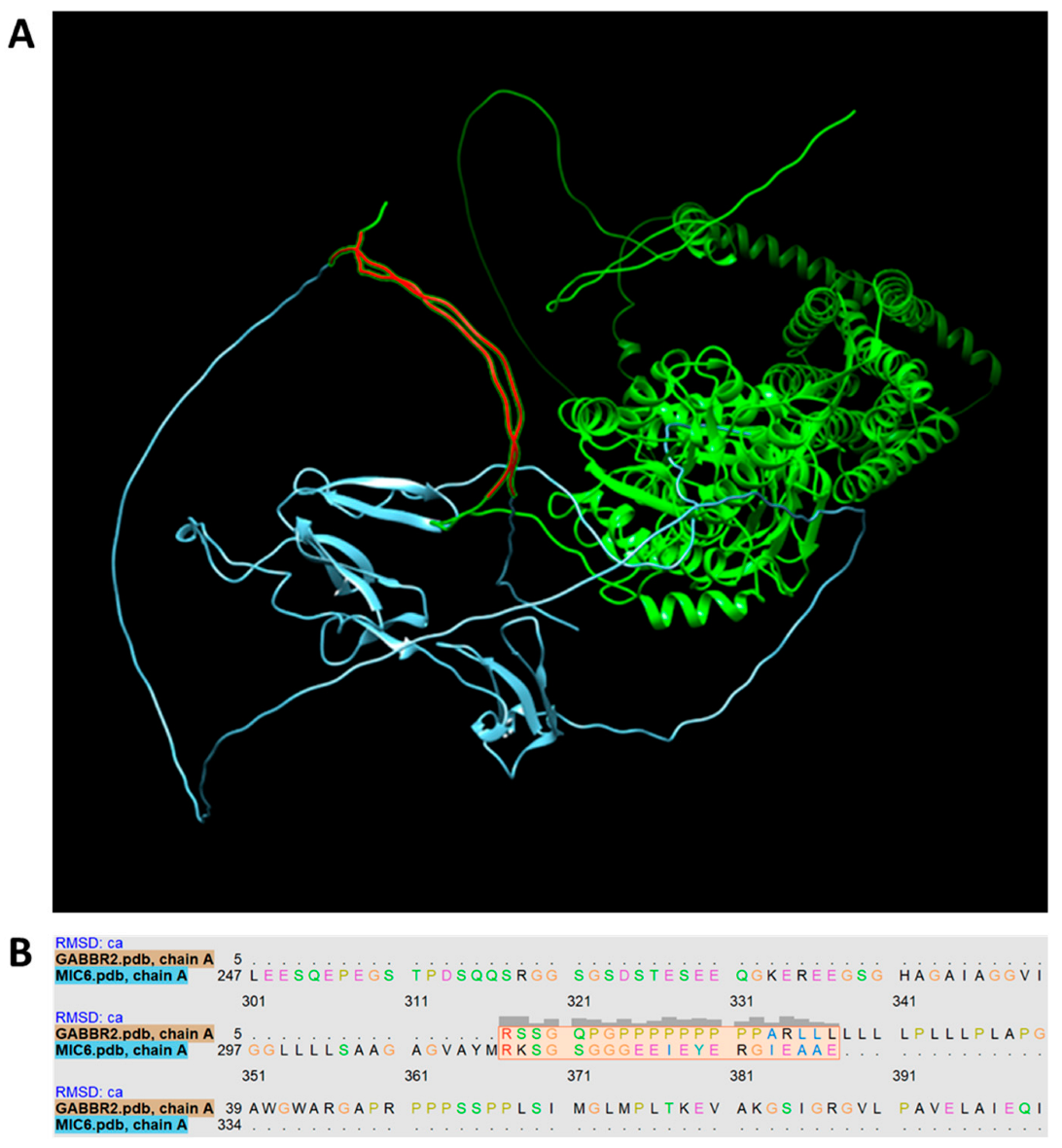
2.9. Protein Match between T. gondii and CNS
2.10. Statistical Analysis
3. Results
3.1. Pregestational T. gondii Lysate Administration Induces Locomotor Alterations in the Progeny
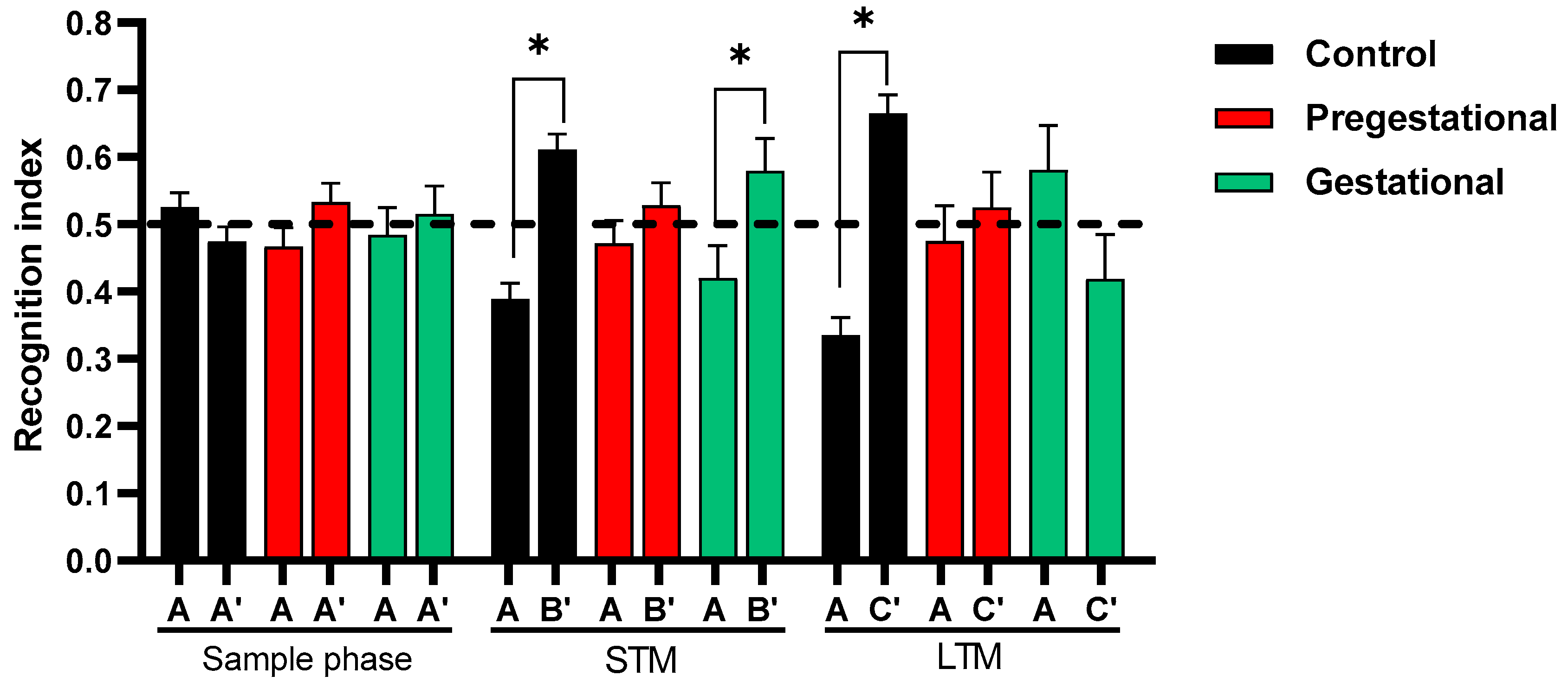
3.2. Cognitive Alteration Induced by the Maternal Immunization with T. gondii Lysate
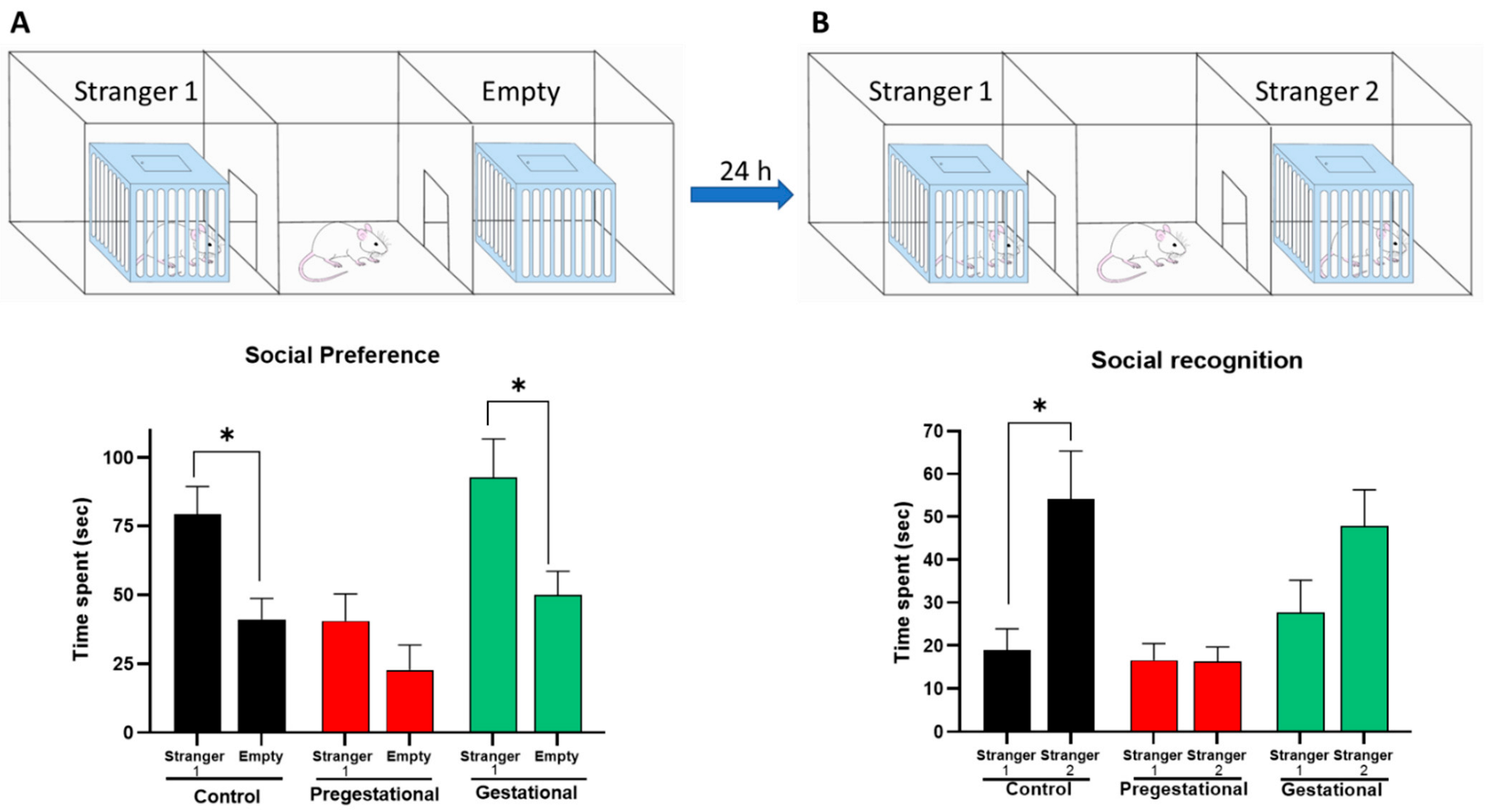
3.3. Social Behavior Alteration on the Progeny of Mothers Exposed to T. gondii Lysate
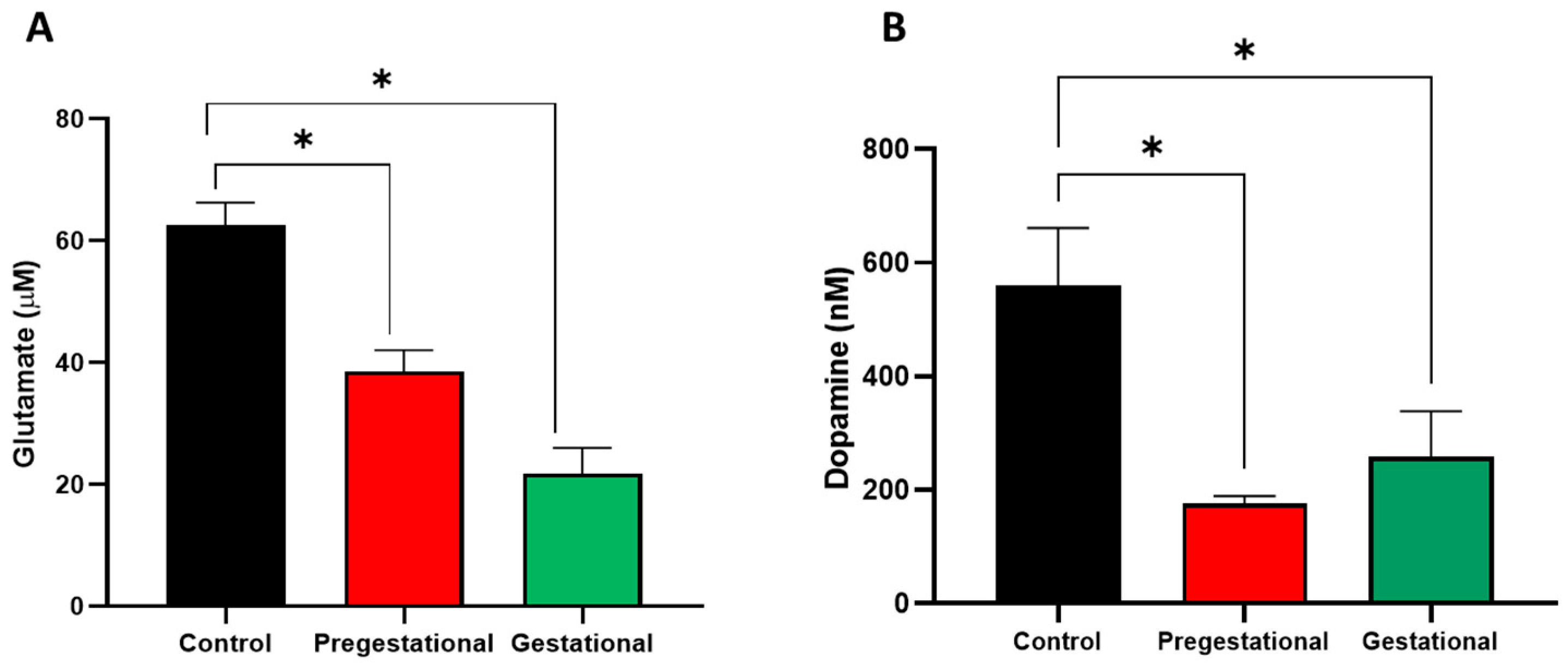
3.4. Neurochemical Alterations on the Progeny of Mothers Exposed to T. gondii Lysate
3.5. Formation of Pathogenic Antibodies Directed against Fetal Brain Proteins
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Matta, S.K.; Rinkenberger, N.; Dunay, I.R.; Sibley, L.D. Toxoplasma gondii infection and its implications within the central nervous system. Nat. Rev. Microbiol. 2021, 19, 467–480. [Google Scholar] [CrossRef] [PubMed]
- Fišar, Z. Biological hypotheses, risk factors, and biomarkers of schizophrenia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2022, 120, 110626. [Google Scholar] [CrossRef] [PubMed]
- Piper, M.; Beneyto, M.; Burne, T.H.; Eyles, D.W.; Lewis, D.A.; McGrath, J.J. The neurodevelopmental hypothesis of schizophrenia: Convergent clues from epidemiology and neuropathology. Psychiatr. Clin. N. Am. 2012, 35, 571–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichard, J.; Zimmer-Bensch, G. The Epigenome in Neurodevelopmental Disorders. Front. Neurosci. 2021, 15, 776809. [Google Scholar] [CrossRef] [PubMed]
- Pantelis, C.; Velakoulis, D.; McGorry, P.D.; Wood, S.J.; Suckling, J.; Phillips, L.J.; Yung, A.R.; Bullmore, E.T.; Brewer, W.; Soulsby, B.; et al. Neuroanatomical abnormalities before and after onset of psychosis: A cross-sectional and longitudinal MRI comparison. Lancet 2003, 361, 281–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pantelis, C.; Yucel, M.; Wood, S.J.; Velakoulis, D.; Sun, D.; Berger, G.; Stuart, G.W.; Yung, A.; Phillips, L.; McGorry, P.D. Structural brain imaging evidence for multiple pathological processes at different stages of brain development in schizophrenia. Schizophr. Bull. 2005, 31, 672–696. [Google Scholar] [CrossRef] [Green Version]
- Selemon, L.D.; Goldman-Rakic, P.S. The reduced neuropil hypothesis: A circuit based model of schizophrenia. Biol. Psychiatry 1999, 45, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.S.; Begg, M.D.; Gravenstein, S.; Schaefer, C.A.; Wyatt, R.J.; Bresnahan, M.; Babulas, V.P.; Susser, E.S. Serologic evidence of prenatal influenza in the etiology of schizophrenia. Arch. Gen. Psychiatry 2004, 61, 774–780. [Google Scholar] [CrossRef] [Green Version]
- Brown, A.S.; Cohen, P.; Greenwald, S.; Susser, E. Nonaffective psychosis after prenatal exposure to rubella. Am. J. Psychiatry 2000, 157, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.S.; Schaefer, C.A.; Quesenberry, C.P., Jr.; Liu, L.; Babulas, V.P.; Susser, E.S. Maternal exposure to toxoplasmosis and risk of schizophrenia in adult offspring. Am. J. Psychiatry 2005, 162, 767–773. [Google Scholar] [CrossRef] [PubMed]
- Buka, S.L.; Tsuang, M.T.; Torrey, E.F.; Klebanoff, M.A.; Bernstein, D.; Yolken, R.H. Maternal infections and subsequent psychosis among offspring. Arch. Gen. Psychiatry 2001, 58, 1032–1037. [Google Scholar] [CrossRef] [Green Version]
- Patterson, P.H. Immune involvement in schizophrenia and autism: Etiology, pathology and animal models. Behav. Brain Res. 2009, 204, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Urakubo, A.; Jarskog, L.F.; Lieberman, J.A.; Gilmore, J.H. Prenatal exposure to maternal infection alters cytokine expression in the placenta, amniotic fluid, and fetal brain. Schizophr. Res. 2001, 47, 27–36. [Google Scholar] [CrossRef]
- Samuelsson, A.M.; Jennische, E.; Hansson, H.A.; Holmang, A. Prenatal exposure to interleukin-6 results in inflammatory neurodegeneration in hippocampus with NMDA/GABA(A) dysregulation and impaired spatial learning. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290, R1345–R1356. [Google Scholar] [CrossRef] [Green Version]
- Cai, Z.; Pan, Z.L.; Pang, Y.; Evans, O.B.; Rhodes, P.G. Cytokine induction in fetal rat brains and brain injury in neonatal rats after maternal lipopolysaccharide administration. Pediatr. Res. 2000, 47, 64–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonakait, G.M. The effects of maternal inflammation on neuronal development: Possible mechanisms. Int. J. Dev. Neurosci. 2007, 25, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.S.; Susser, E.S. In utero infection and adult schizophrenia. Ment. Retard. Dev. Disabil. Res. Rev. 2002, 8, 51–57. [Google Scholar] [CrossRef]
- Dammann, O.; Leviton, A. Maternal intrauterine infection, cytokines, and brain damage in the preterm newborn. Pediatr. Res. 1997, 42, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Gilmore, J.H.; Fredrik Jarskog, L.; Vadlamudi, S.; Lauder, J.M. Prenatal infection and risk for schizophrenia: IL-1beta, IL-6, and TNFalpha inhibit cortical neuron dendrite development. Neuropsychopharmacology 2004, 29, 1221–1229. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, P.B.; Norgaard-Pedersen, B.; Waltoft, B.L.; Sorensen, T.L.; Hougaard, D.; Yolken, R.H. Early infections of Toxoplasma gondii and the later development of schizophrenia. Schizophr. Bull. 2007, 33, 741–744. [Google Scholar] [CrossRef]
- Martin, L.A.; Ashwood, P.; Braunschweig, D.; Cabanlit, M.; Van de Water, J.; Amaral, D.G. Stereotypies and hyperactivity in rhesus monkeys exposed to IgG from mothers of children with autism. Brain Behav. Immun. 2008, 22, 806–816. [Google Scholar] [CrossRef] [Green Version]
- Singer, H.S.; Morris, C.; Gause, C.; Pollard, M.; Zimmerman, A.W.; Pletnikov, M. Prenatal exposure to antibodies from mothers of children with autism produces neurobehavioral alterations: A pregnant dam mouse model. J. Neuroimmunol. 2009, 211, 39–48. [Google Scholar] [CrossRef]
- Braunschweig, D.; Golub, M.S.; Koenig, C.M.; Qi, L.; Pessah, I.N.; Van de Water, J.; Berman, R.F. Maternal autism-associated IgG antibodies delay development and produce anxiety in a mouse gestational transfer model. J. Neuroimmunol. 2012, 252, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.; Huerta, P.T.; Zhang, J.; Kowal, C.; Bertini, E.; Volpe, B.T.; Diamond, B. Neurotoxic autoantibodies mediate congenital cortical impairment of offspring in maternal lupus. Nat. Med. 2009, 15, 91–96. [Google Scholar] [CrossRef] [Green Version]
- Sonaimuthu, P.; Ching, X.T.; Fong, M.Y.; Kalyanasundaram, R.; Lau, Y.L. Induction of Protective Immunity against Toxoplasmosis in BALB/c Mice Vaccinated with Toxoplasma gondii Rhoptry-1. Front. Microbiol. 2016, 7, 808. [Google Scholar] [CrossRef] [Green Version]
- Tyebji, S.; Seizova, S.; Garnham, A.L.; Hannan, A.J.; Tonkin, C.J. Impaired social behaviour and molecular mediators of associated neural circuits during chronic Toxoplasma gondii infection in female mice. Brain Behav. Immun. 2019, 80, 88–108. [Google Scholar] [CrossRef]
- Kaidanovich-Beilin, O.; Lipina, T.; Vukobradovic, I.; Roder, J.; Woodgett, J.R. Assessment of social interaction behaviors. J. Vis. Exp. 2011, 48, e2473. [Google Scholar] [CrossRef]
- Mednick, S.A.; Machon, R.A.; Huttunen, M.O.; Bonett, D. Adult schizophrenia following prenatal exposure to an influenza epidemic. Arch. Gen. Psychiatry 1988, 45, 189–192. [Google Scholar] [CrossRef]
- Brown, A.S. Prenatal infection as a risk factor for schizophrenia. Schizophr. Bull. 2006, 32, 200–202. [Google Scholar] [CrossRef] [Green Version]
- Brown, A.S. Epidemiologic studies of exposure to prenatal infection and risk of schizophrenia and autism. Dev. Neurobiol. 2012, 72, 1272–1276. [Google Scholar] [CrossRef]
- Mortensen, P.B.; Norgaard-Pedersen, B.; Waltoft, B.L.; Sorensen, T.L.; Hougaard, D.; Torrey, E.F.; Yolken, R.H. Toxoplasma gondii as a risk factor for early-onset schizophrenia: Analysis of filter paper blood samples obtained at birth. Biol. Psychiatry 2007, 61, 688–693. [Google Scholar] [CrossRef] [PubMed]
- Malek, A.; Sager, R.; Kuhn, P.; Nicolaides, K.H.; Schneider, H. Evolution of maternofetal transport of immunoglobulins during human pregnancy. Am. J. Reprod. Immunol. 1996, 36, 248–255. [Google Scholar] [CrossRef]
- Xu, Z.; Zhang, X.; Chang, H.; Kong, Y.; Ni, Y.; Liu, R.; Zhang, X.; Hu, Y.; Yang, Z.; Hou, M.; et al. Rescue of maternal immune activation-induced behavioral abnormalities in adult mouse offspring by pathogen-activated maternal Treg cells. Nat. Neurosci. 2021, 24, 818–830. [Google Scholar] [CrossRef]
- Meyer, U.; Feldon, J. To poly(I:C) or not to poly(I:C): Advancing preclinical schizophrenia research through the use of prenatal immune activation models. Neuropharmacology 2012, 62, 1308–1321. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Smith, S.E.; Malkova, N.; Tse, D.; Su, Y.; Patterson, P.H. Activation of the maternal immune system alters cerebellar development in the offspring. Brain Behav. Immun. 2009, 23, 116–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bake, S.; Friedman, J.A.; Sohrabji, F. Reproductive age-related changes in the blood brain barrier: Expression of IgG and tight junction proteins. Microvasc. Res. 2009, 78, 413–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, G.H.; Yuan, Z.G.; Zhou, D.H.; He, X.H.; Liu, M.M.; Yan, C.; Yin, C.C.; He, Y.; Lin, R.Q.; Zhu, X.Q. Toxoplasma gondii microneme protein 6 (MIC6) is a potential vaccine candidate against toxoplasmosis in mice. Vaccine 2009, 27, 6570–6574. [Google Scholar] [CrossRef]
- Xu, X.P.; Liu, W.G.; Xu, Q.M.; Zhu, X.Q.; Chen, J. Evaluation of immune protection against Toxoplasma gondii infection in mice induced by a multi-antigenic DNA vaccine containing TgGRA24, TgGRA25 and TgMIC6. Parasite 2019, 26, 58. [Google Scholar] [CrossRef] [Green Version]
- Coyle, J.T.; Konopaske, G.T. Chapter 58-The Neurochemistry of Schizophrenia. In Basic Neurochemistry, 8th ed.; Brady, S.T., Siegel, G.J., Albers, R.W., Price, D.L., Eds.; Academic Press: Cambridge, MA, USA, 2012; pp. 1000–1011. [Google Scholar] [CrossRef]
- Anderson, G.; Maes, M. Schizophrenia: Linking prenatal infection to cytokines, the tryptophan catabolite (TRYCAT) pathway, NMDA receptor hypofunction, neurodevelopment and neuroprogression. Prog. Neuropsychopharmacol. Biol. Psychiatry 2013, 42, 5–19. [Google Scholar] [CrossRef]
- Konopaske, G.T.; Coyle, J.T. Chapter 46-The neurobiology of schizophrenia∗. In Neurobiology of Brain Disorders, 2nd ed.; Zigmond, M.J., Wiley, C.A., Chesselet, M.-F., Eds.; Academic Press: Cambridge, MA, USA, 2023; pp. 843–860. [Google Scholar] [CrossRef]
- Bressan, R.A.; Crippa, J.A. The role of dopamine in reward and pleasure behaviour--review of data from preclinical research. Acta Psychiatr. Scand. Suppl. 2005, 427, 14–21. [Google Scholar] [CrossRef]
- Luby, E.D.; Cohen, B.D.; Rosenbaum, G.; Gottlieb, J.S.; Kelley, R. Study of a new schizophrenomimetic drug; sernyl. AMA Arch. Neurol. Psychiatry 1959, 81, 363–369. [Google Scholar] [CrossRef]
- Anis, N.A.; Berry, S.C.; Burton, N.R.; Lodge, D. The dissociative anaesthetics, ketamine and phencyclidine, selectively reduce excitation of central mammalian neurones by N-methyl-aspartate. Br. J. Pharmacol. 1983, 79, 565–575. [Google Scholar] [CrossRef] [Green Version]
- Coyle, J.T. Glutamate and schizophrenia: Beyond the dopamine hypothesis. Cell. Mol. Neurobiol. 2006, 26, 365–384. [Google Scholar] [CrossRef]
- Coyle, J.T.; Basu, A.; Benneyworth, M.; Balu, D.; Konopaske, G. Glutamatergic synaptic dysregulation in schizophrenia: Therapeutic implications. Handb. Exp. Pharmacol. 2012, 213, 267–295. [Google Scholar] [CrossRef] [Green Version]
- Schwarcz, R.; Hunter, C.A. Toxoplasma gondii and schizophrenia: Linkage through astrocyte-derived kynurenic acid? Schizophr. Bull. 2007, 33, 652–653. [Google Scholar] [CrossRef] [Green Version]
- Schwarcz, R.; Rassoulpour, A.; Wu, H.Q.; Medoff, D.; Tamminga, C.A.; Roberts, R.C. Increased cortical kynurenate content in schizophrenia. Biol. Psychiatry 2001, 50, 521–530. [Google Scholar] [CrossRef]
- Sathyasaikumar, K.V.; Stachowski, E.K.; Wonodi, I.; Roberts, R.C.; Rassoulpour, A.; McMahon, R.P.; Schwarcz, R. Impaired kynurenine pathway metabolism in the prefrontal cortex of individuals with schizophrenia. Schizophr. Bull. 2011, 37, 1147–1156. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romero Núñez, E.; Blanco Ayala, T.; Vázquez Cervantes, G.I.; Roldán-Roldán, G.; González Esquivel, D.F.; Muñiz-Hernández, S.; Salazar, A.; Méndez Armenta, M.; Gómez-Manzo, S.; González-Conchillos, H.; et al. Pregestational Exposure to T. gondii Produces Maternal Antibodies That Recognize Fetal Brain Mimotopes and Induces Neurochemical and Behavioral Dysfunction in the Offspring. Cells 2022, 11, 3819. https://doi.org/10.3390/cells11233819
Romero Núñez E, Blanco Ayala T, Vázquez Cervantes GI, Roldán-Roldán G, González Esquivel DF, Muñiz-Hernández S, Salazar A, Méndez Armenta M, Gómez-Manzo S, González-Conchillos H, et al. Pregestational Exposure to T. gondii Produces Maternal Antibodies That Recognize Fetal Brain Mimotopes and Induces Neurochemical and Behavioral Dysfunction in the Offspring. Cells. 2022; 11(23):3819. https://doi.org/10.3390/cells11233819
Chicago/Turabian StyleRomero Núñez, Eunice, Tonali Blanco Ayala, Gustavo Ignacio Vázquez Cervantes, Gabriel Roldán-Roldán, Dinora Fabiola González Esquivel, Saé Muñiz-Hernández, Alelí Salazar, Maricela Méndez Armenta, Saúl Gómez-Manzo, Hugo González-Conchillos, and et al. 2022. "Pregestational Exposure to T. gondii Produces Maternal Antibodies That Recognize Fetal Brain Mimotopes and Induces Neurochemical and Behavioral Dysfunction in the Offspring" Cells 11, no. 23: 3819. https://doi.org/10.3390/cells11233819