
LncKCNQ1OT1 Promotes the Odontoblastic Differentiation of Dental Pulp Stem Cells via Regulating hsa-miR-153-3p/RUNX2 Axis
Abstract
:1. Introduction
2. Materials and Methods
2.1. DPSCs Culture
2.2. Cell Transfection
2.3. Cell Viability Assay
2.4. Odontoblastic Differentiation Culture
2.5. Quantitative Real-Time PCR (qRT-PCR)
2.6. Luciferase Reporter Gene Experiment
2.7. Alkaline Phosphatase (ALP) Activity Assay
2.8. Alizarin Red Staining
2.9. Western Blot Analysis
2.10. Statistical Analysis
3. Results
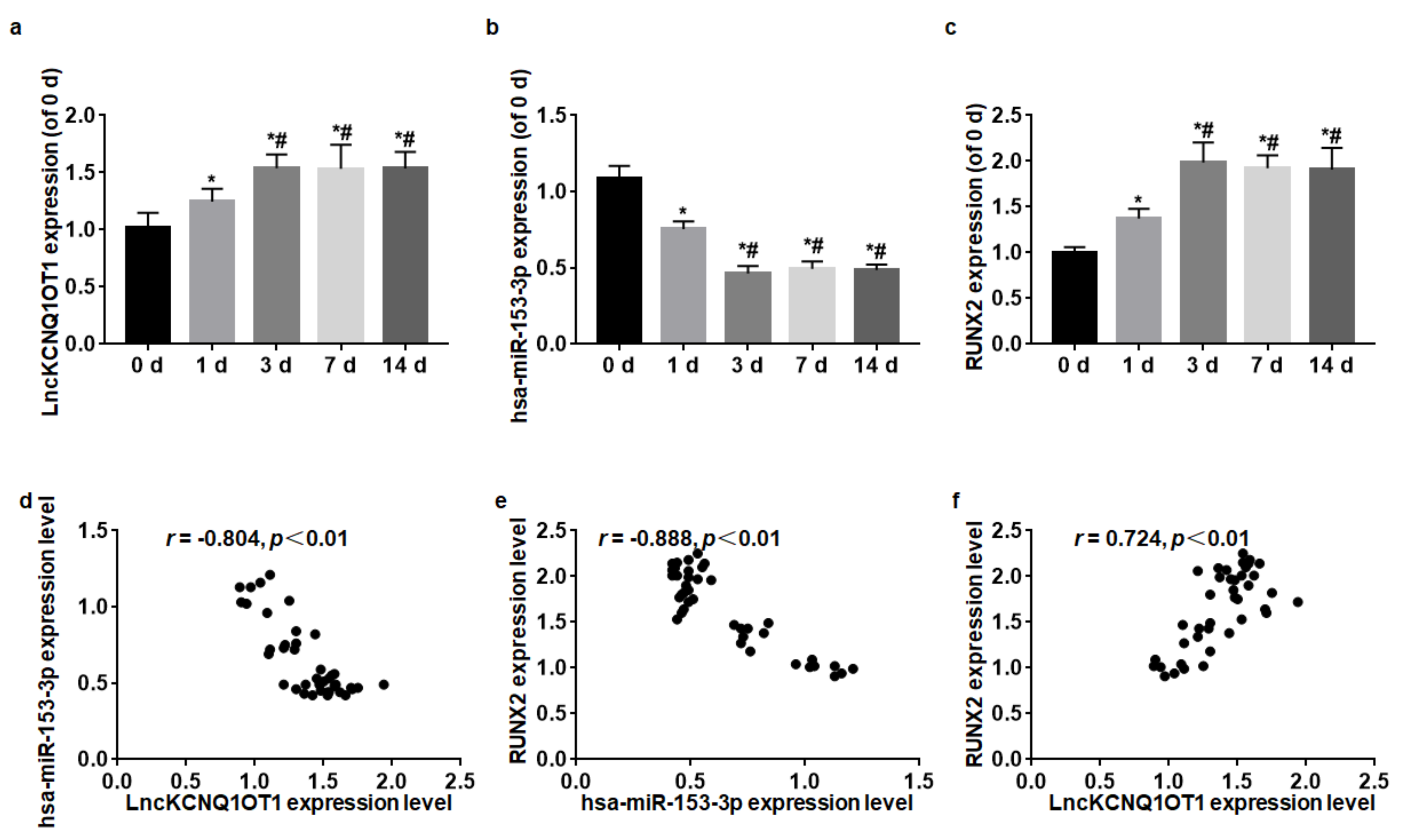
3.1. LncKCNQ1OT1, hsa-miR-153-3p, and RUNX2 Expression Levels during Odontoblastic Differentiation
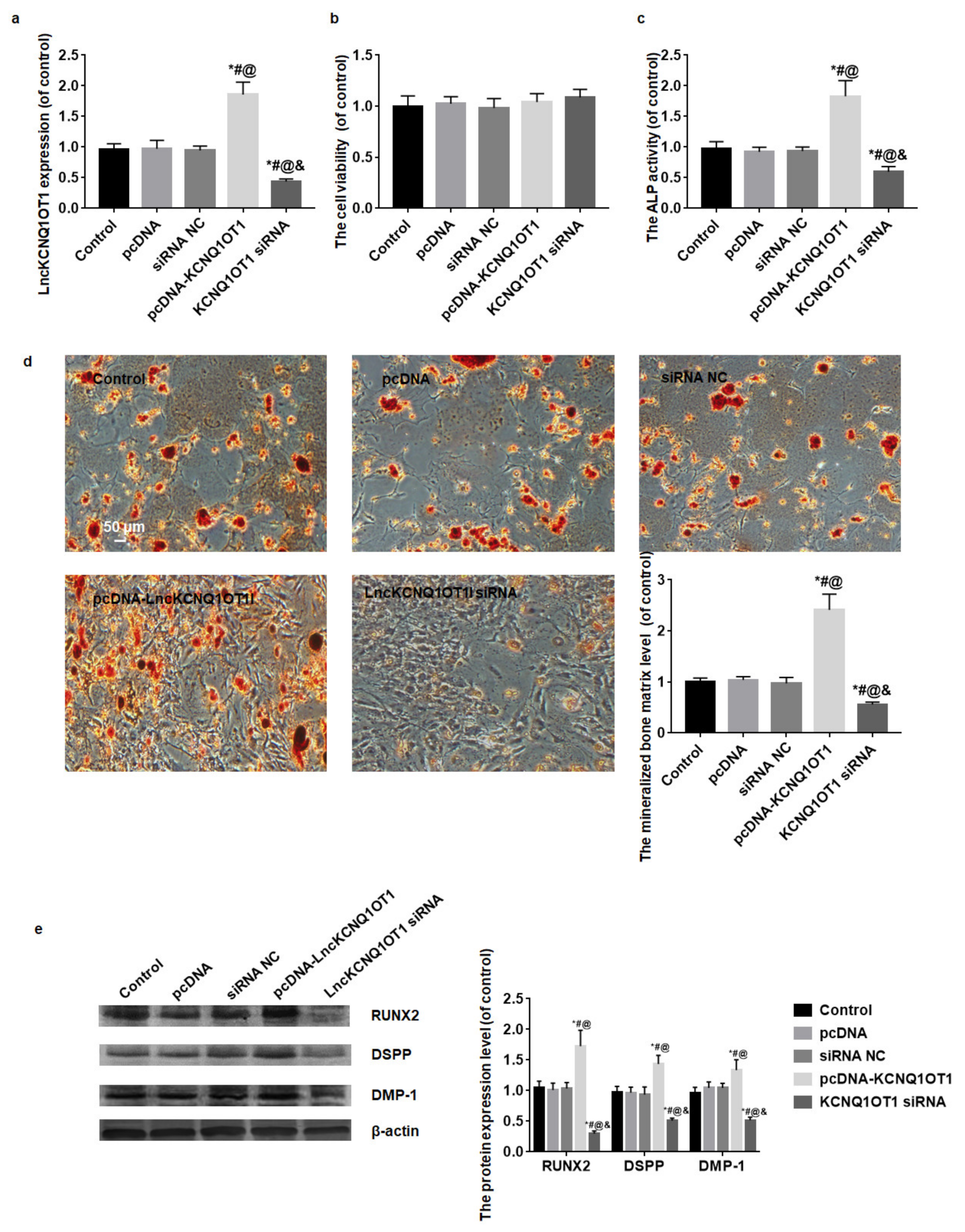
3.2. The Effect of LncKCNQ1OT1 on the Cell Viability and Odontoblastic Differentiation of DPSCs
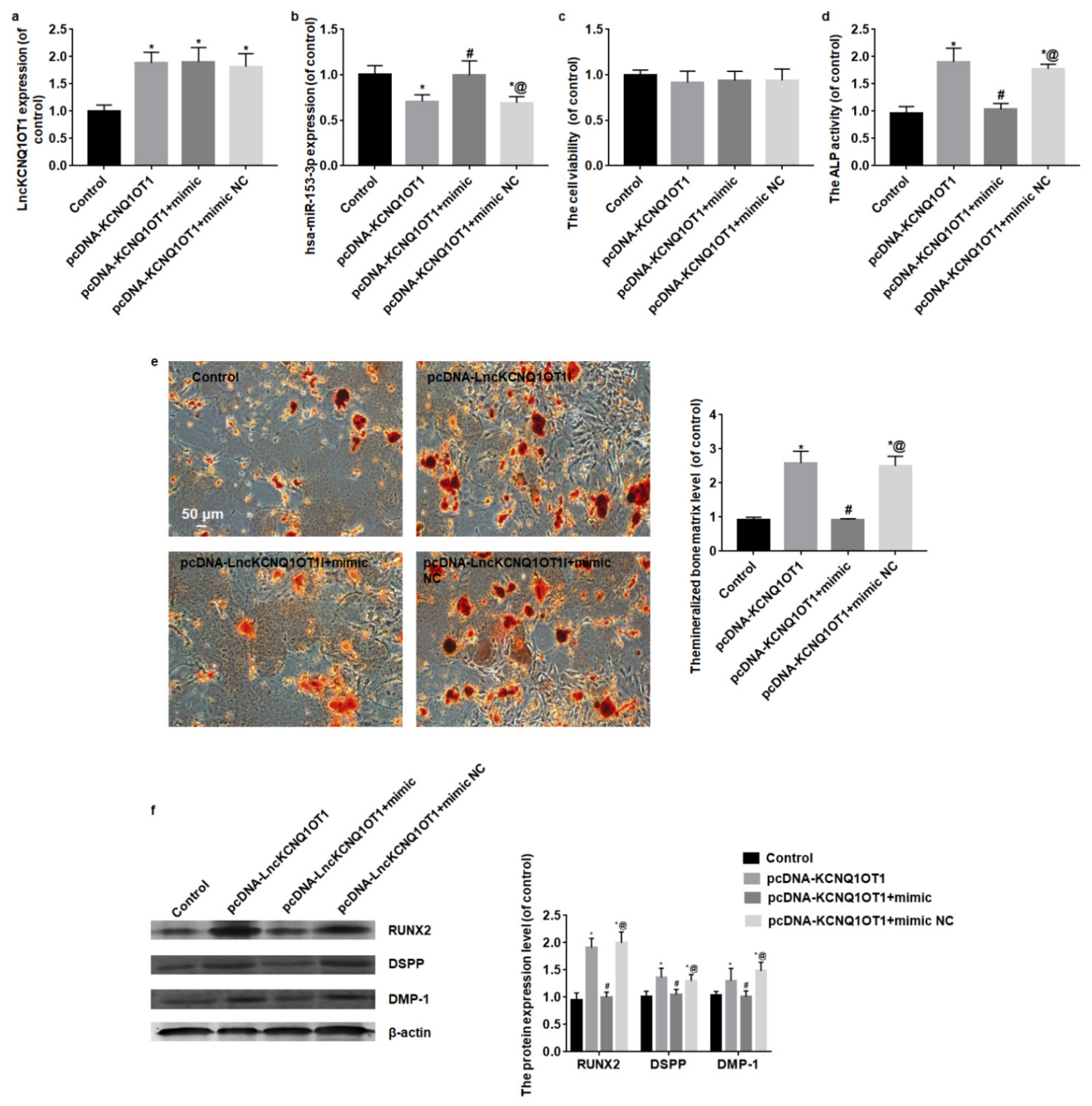
3.3. Overexpression of hsa-miR-153-3p Reverses the Promotion of Odontoblastic Differentiation Induced by LncKCNQ1OT1
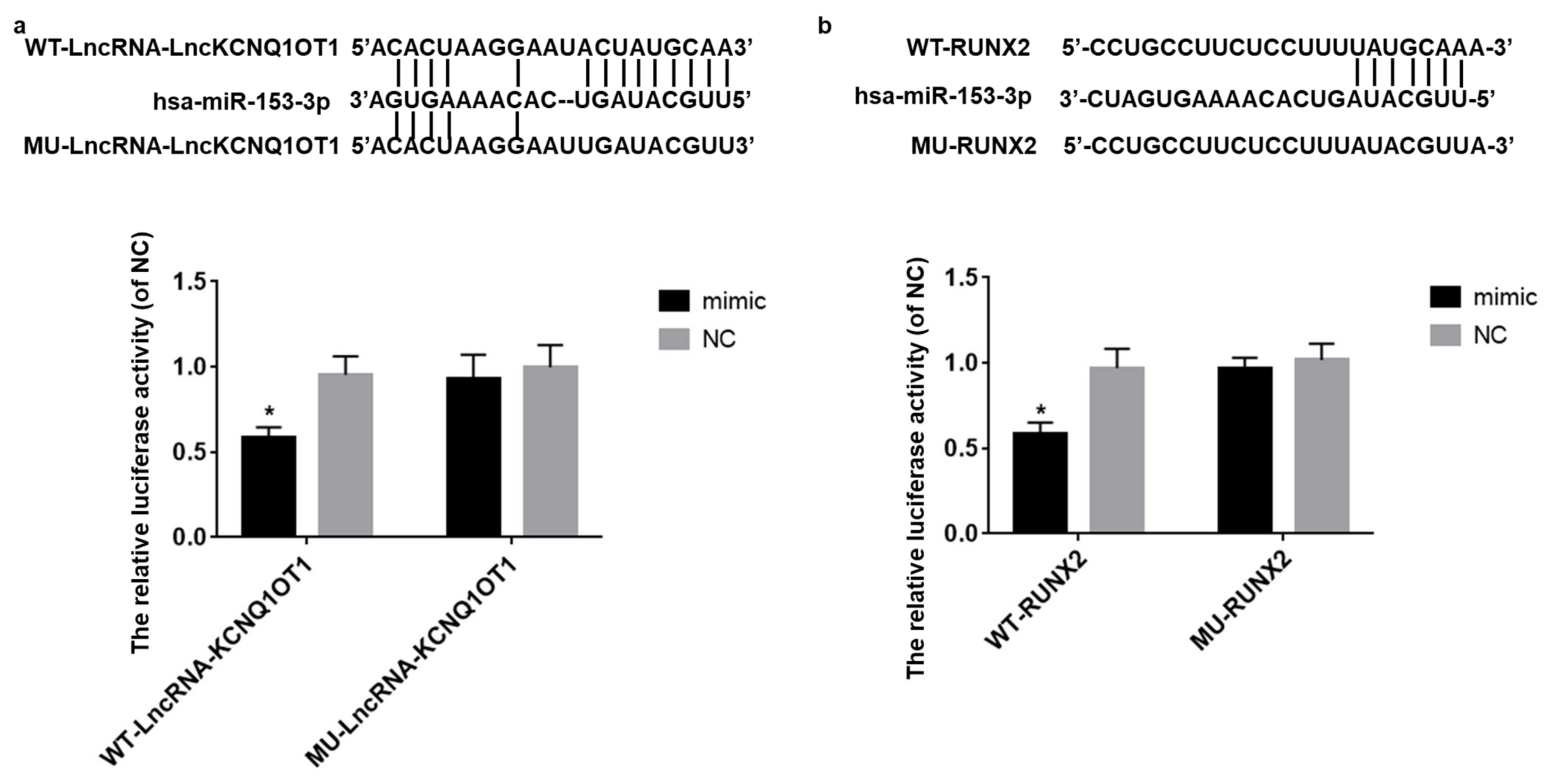
3.4. LncKCNQ1OT1 Act as a Sponge of hsa-miR-153-3p and hsa-miR-153-3p Target on RUNX2
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aydin, S.; Sahin, F. Stem Cells Derived from Dental Tissues. Adv. Exp. Med. Biol. 2019, 1144, 123–132. [Google Scholar] [PubMed]
- Nuti, N.; Corallo, C.; Chan, B.M.F.; Ferrari, M.; Gerami-Naini, B. Multipotent Differentiation of Human Dental Pulp Stem Cells: A Literature Review. Stem Cell Rev. Rep. 2016, 12, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Tatullo, M.; Marrelli, M.; Shakesheff, K.M.; White, L.J. Dental pulp stem cells: Function, isolation and applications in regenerative medicine. J. Tissue Eng. Regen. Med. 2015, 9, 1205–1216. [Google Scholar] [CrossRef] [PubMed]
- Itoh, Y.; Sasaki, J.I.; Hashimoto, M.; Katata, C.; Hayashi, M.; Imazato, S. Pulp Regeneration by 3-dimensional Dental Pulp Stem Cell Constructs. J. Dent. Res. 2018, 97, 1137–1143. [Google Scholar] [CrossRef] [PubMed]
- Dong, Q.; Wang, Y.; Mohabatpour, F.; Zheng, L.; Papagerakis, S.; Chen, D.; Papagerakis, P. Dental Pulp Stem Cells: Isolation, Characterization, Expansion, and Odontoblast Differentiation for Tissue Engineering. Methods Mol. Biol. 2019, 1922, 91–101. [Google Scholar]
- Moonesi Rad, R.; Atila, D.; Akgün, E.E.; Evis, Z.; Keskin, D.; Tezcaner, A. Evaluation of human dental pulp stem cells behavior on a novel nanobiocomposite scaffold prepared for regenerative endodontics. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 100, 928–948. [Google Scholar] [CrossRef]
- Zhai, Q.; Dong, Z.; Wang, W.; Li, B.; Jin, Y. Dental stem cell and dental tissue regeneration. Front. Med. 2019, 13, 152–159. [Google Scholar] [CrossRef]
- Moussa, D.G.; Aparicio, C. Present and future of tissue engineering scaffolds for dentin-pulp complex regeneration. J. Tissue Eng. Regen. Med. 2019, 13, 58–75. [Google Scholar] [CrossRef] [Green Version]
- Cui, D.; Xiao, J.; Zhou, Y.; Zhou, X.; Liu, Y.; Peng, Y.; Yu, Y.; Li, H.; Zhou, X.; Yuan, Q.; et al. Epiregulin enhances odontoblastic differentiation of dental pulp stem cells via activating MAPK signalling pathway. Cell Prolif. 2019, 52, e12680. [Google Scholar] [CrossRef] [Green Version]
- Zhong, J.; Tu, X.; Kong, Y.; Guo, L.; Li, B.; Zhong, W.; Cheng, Y.; Jiang, Y.; Jiang, Q. LncRNA H19 promotes odontoblastic differentiation of human dental pulp stem cells by regulating miR-140-5p and BMP-2/FGF9. Stem Cell Res. Ther. 2020, 11, 202. [Google Scholar] [CrossRef]
- Zhang, X.; Ning, T.; Wang, H.; Xu, S.; Yu, H.; Luo, X.; Hao, C.; Wu, B.; Ma, D. Stathmin regulates the proliferation and odontoblastic/osteogenic differentiation of human dental pulp stem cells through Wnt/beta-catenin signaling pathway. J. Proteomics 2019, 202, 103364. [Google Scholar] [CrossRef]
- Yu, H.; Zhang, X.; Song, W.; Pan, T.; Wang, H.; Ning, T.; Wei, Q.; Xu, H.H.K.; Wu, B.; Ma, D. Effects of 3-dimensional Bioprinting Alginate/Gelatin Hydrogel Scaffold Extract on Proliferation and Differentiation of Human Dental Pulp Stem Cells. J. Endod. 2019, 45, 706–715. [Google Scholar] [CrossRef]
- Lu, X.; Chen, X.; Xing, J.; Lian, M.; Huang, D.; Lu, Y.; Feng, G.; Feng, X. miR-140-5p regulates the odontoblastic differentiation of dental pulp stem cells via the Wnt1/beta-catenin signaling pathway. Stem Cell Res. Ther. 2019, 10, 226. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.; Zhang, Q.; Yu, S.; Lin, Y.; Li, S.; Liu, H.; Chen, Z. miR-3065-5p regulates mouse odontoblastic differentiation partially through bone morphogenetic protein receptor type II. Biochem. Biophys. Res. Commun. 2018, 495, 493–498. [Google Scholar] [CrossRef]
- Li, Q.; Huang, L. MiR-148a-3p Regulates the Invasion and Odontoblastic Differentiation of Human Dental Pulp Stem Cells via the Wnt1/beta-Catenin Pathway. Int. J. Stem Cells 2021, 4, 9822. [Google Scholar]
- Marques, T.M.; Gama-Carvalho, M. Network Approaches to Study Endogenous RNA Competition and Its Impact on Tissue-Specific microRNA Functions. Biomolecules 2022, 12, 1972. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, W.; Zhu, W.; Dong, J.; Cheng, Y.; Yin, Z.; Shen, F. Mechanisms and Functions of Long Non-Coding RNAs at Multiple Regulatory Levels. Int. J. Mol. Sci. 2019, 20, 276. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Zhang, K.; Qiu, W.; Luo, Y.; Pan, Y.; Li, J.; Yang, Y.; Wu, B.; Fang, F. Genome-wide identification of long noncoding RNAs and their competing endogenous RNA networks involved in the odontogenic differentiation of human dental pulp stem cells. Stem Cell Res. Ther. 2020, 11, 114. [Google Scholar] [CrossRef] [Green Version]
- Fang, F.; Zhang, K.; Chen, Z.; Wu, B. Noncoding RNAs: New insights into the odontogenic differentiation of dental tissue-derived mesenchymal stem cells. Stem Cell Res. Ther. 2019, 10, 297. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Xu, S.; Dao, J.; Gan, Z.; Zeng, X. Differential expression of lncRNA/miRNA/mRNA and their related functional networks during the osteogenic/odontogenic differentiation of dental pulp stem cells. J. Cell Physiol. 2020, 235, 3350–3361. [Google Scholar] [CrossRef]
- Liu, M.; Chen, L.; Wu, J.; Lin, Z.; Huang, S. Long noncoding RNA MEG3 expressed in human dental pulp regulates LPS-Induced inflammation and odontogenic differentiation in pulpitis. Exp. Cell Res. 2021, 400, 112495. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Song, Z.; Huang, S.; Wang, R.; Qin, W.; Guo, J.; Lin, Z. lncRNA DANCR suppresses odontoblast-like differentiation of human dental pulp cells by inhibiting wnt/beta-catenin pathway. Cell Tissue Res. 2016, 364, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Camilleri, S.; McDonald, F. Runx2 and dental development. Eur. J. Oral Sci. 2006, 114, 361–373. [Google Scholar] [CrossRef]
- Han, N.; Zheng, Y.; Li, R.; Li, X.; Zhou, M.; Niu, Y.; Zhang, Q. beta-catenin enhances odontoblastic differentiation of dental pulp cells through activation of Runx2. PLoS ONE 2014, 9, e88890. [Google Scholar]
- Qin, X.; Jiang, Q.; Komori, H.; Sakane, C.; Fukuyama, R.; Matsuo, Y.; Ito, K.; Miyazaki, T.; Komori, T. Runt-related transcription factor-2 (Runx2) is required for bone matrix protein gene expression in committed osteoblasts in mice. J. Bone Miner. Res. 2021, 1, 27111. [Google Scholar] [CrossRef]
- Wang, C.G.; Liao, Z.; Xiao, H.; Liu, H.; Hu, Y.-H.; Liao, Q.-D.; Zhong, D. LncRNA KCNQ1OT1 promoted BMP2 expression to regulate osteogenic differentiation by sponging miRNA-214. Exp. Mol. Pathol. 2019, 107, 77–84. [Google Scholar] [CrossRef]
- Jiang, H.; Jia, P. MiR-153-3p inhibits osteogenic differentiation of periodontal ligament stem cells through KDM6A-induced demethylation of H3K27me3. J. Periodontal Res. 2021, 56, 379–387. [Google Scholar] [CrossRef]
- Xia, W.; Han, X.; Wang, L. E26 transformation-specific 1 is implicated in the inhibition of osteogenic differentiation induced by chronic high glucose by directly regulating Runx2 expression. J. Biomed. Res. 2021, 36, 39–47. [Google Scholar] [CrossRef]
- D’Souza, R.N.; Aberg, T.; Gaikwad, J.; Cavender, A.; Owen, M.; Karsenty, G.; Thesleff, I. Cbfa1 is required for epithelial-mesenchymal interactions regulating tooth development in mice. Development 1999, 126, 2911–2920. [Google Scholar] [CrossRef]
- Jiang, H.; Sodek, J.; Karsenty, G.; Thomas, H.; Ranly, D.; Chen, J. Expression of core binding factor Osf2/Cbfa-1 and bone sialoprotein in tooth development. Mech. Dev. 1999, 81, 169–173. [Google Scholar] [CrossRef]
- Tu, S.; Wu, J.; Chen, L.; Tian, Y.; Qin, W.; Huang, S.; Wang, R.; Lin, Z.; Song, Z. LncRNA CALB2 sponges miR-30b-3p to promote odontoblast differentiation of human dental pulp stem cells via up-regulating RUNX2. Cell Signal. 2020, 73, 109695. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Liu, H.; Lin, H.; Yuan, G.; Zhang, L.; Chen, Z. MicroRNA-338-3p promotes differentiation of mDPC6T into odontoblast-like cells by targeting Runx2. Mol. Cell Biochem. 2013, 377, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Zhan, F.L.; Liu, X.Y.; Wang, X.B. The Role of MicroRNA-143-5p in the Differentiation of Dental Pulp Stem Cells into Odontoblasts by Targeting Runx2 via the OPG/RANKL Signaling Pathway. J. Cell Biochem. 2018, 119, 536–546. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Chen, Y.; Zhang, X.; Lu, X.; Hong, J.; Guo, X.; Zhou, D. Knockdown of lncRNA KCNQ1OT1 suppresses the adipogenic and osteogenic differentiation of tendon stem cell via downregulating miR-138 target genes PPARgamma and RUNX2. Cell Cycle 2018, 17, 2374–2385. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences (5′-3′) |
|---|---|
| LncKCNQ1OT1 | Forward: ACTCACTCACTCACTCACT |
| Reverse: CTGGCTCCTTCTATCACATT | |
| hsa-miR-153-3p | Forward: ACACTCCAGCTGGGTTGCATAGTCACAAA |
| Reverse: CAGTGCGTGTCGTGGAGT | |
| RUNX2 | Forward: TGCCACCTCTGACTTCTGC |
| Reverse: GATGAAATGCCTGGGAACTG | |
| U6 | Forward: GTGCTCGCTTCGGCAGCACAT |
| Reverse: TACCTTGCGAAGTGCTTAAAC | |
| GAPDH | Forward: AGGTGAAGGTCGGAGTCAAC |
| Reverse: CGCTCCTGGAAGATGGTGAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, X.; Zhang, J.; Lu, Y.; Xing, J.; Lian, M.; Feng, G.; Huang, D.; Wang, C.; Shen, N.; Feng, X. LncKCNQ1OT1 Promotes the Odontoblastic Differentiation of Dental Pulp Stem Cells via Regulating hsa-miR-153-3p/RUNX2 Axis. Cells 2022, 11, 3506. https://doi.org/10.3390/cells11213506
Lu X, Zhang J, Lu Y, Xing J, Lian M, Feng G, Huang D, Wang C, Shen N, Feng X. LncKCNQ1OT1 Promotes the Odontoblastic Differentiation of Dental Pulp Stem Cells via Regulating hsa-miR-153-3p/RUNX2 Axis. Cells. 2022; 11(21):3506. https://doi.org/10.3390/cells11213506
Chicago/Turabian StyleLu, Xiaohui, Jiawen Zhang, Yuanzhou Lu, Jing Xing, Min Lian, Guijuan Feng, Dan Huang, Chenfei Wang, Nimei Shen, and Xingmei Feng. 2022. "LncKCNQ1OT1 Promotes the Odontoblastic Differentiation of Dental Pulp Stem Cells via Regulating hsa-miR-153-3p/RUNX2 Axis" Cells 11, no. 21: 3506. https://doi.org/10.3390/cells11213506