
Define the Two Molecular Subtypes of Epithelioid Malignant Pleural Mesothelioma

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
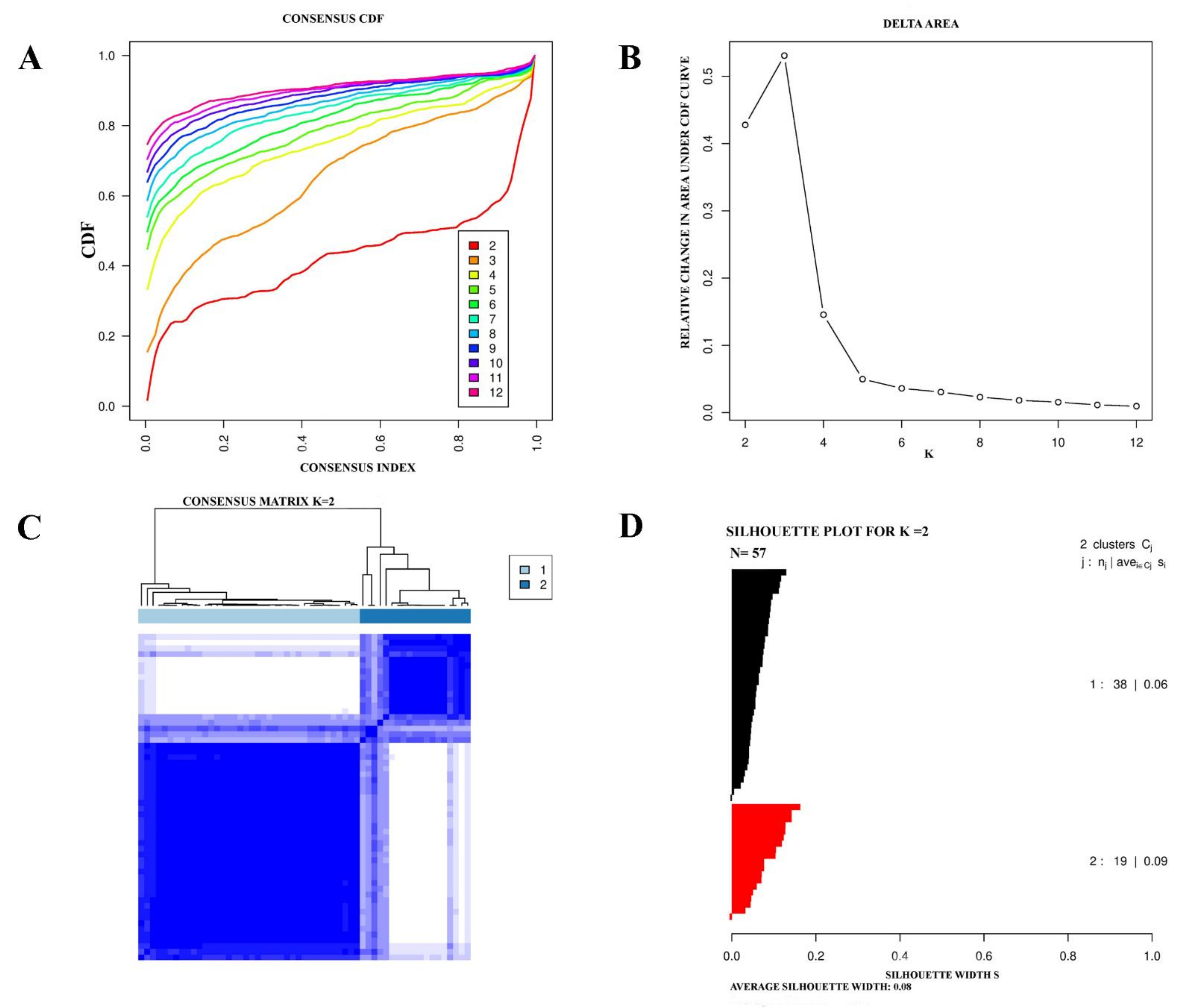
2.1. Determination and Validation of Molecular Subtypes of eMPM
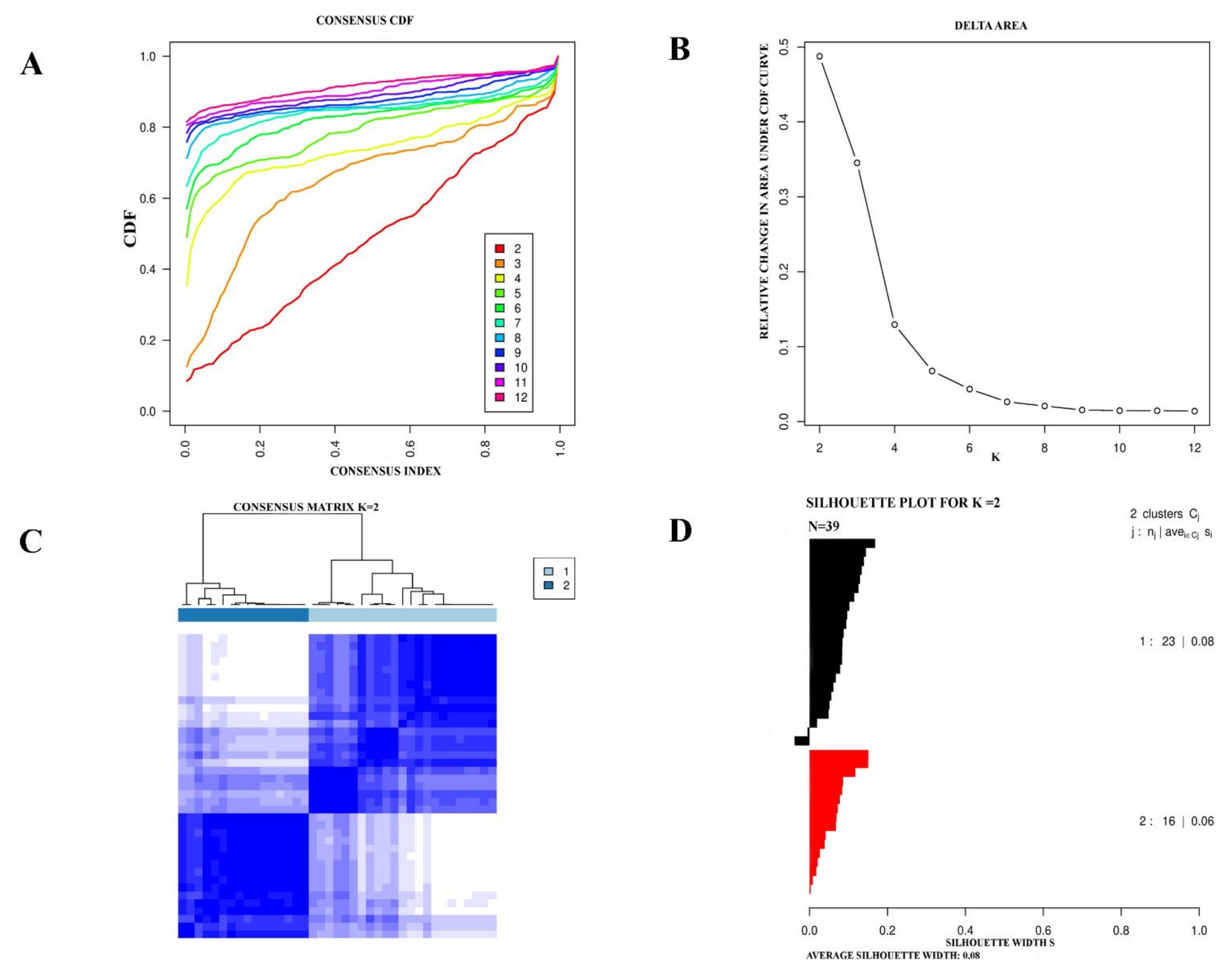
2.2. Reproducibility Measurement of eMPM Molecular Subtypes
2.3. Gene Ontology and Gene Set Enrichment (Gsea) Analysis
2.4. Statistical Analysis
3. Results
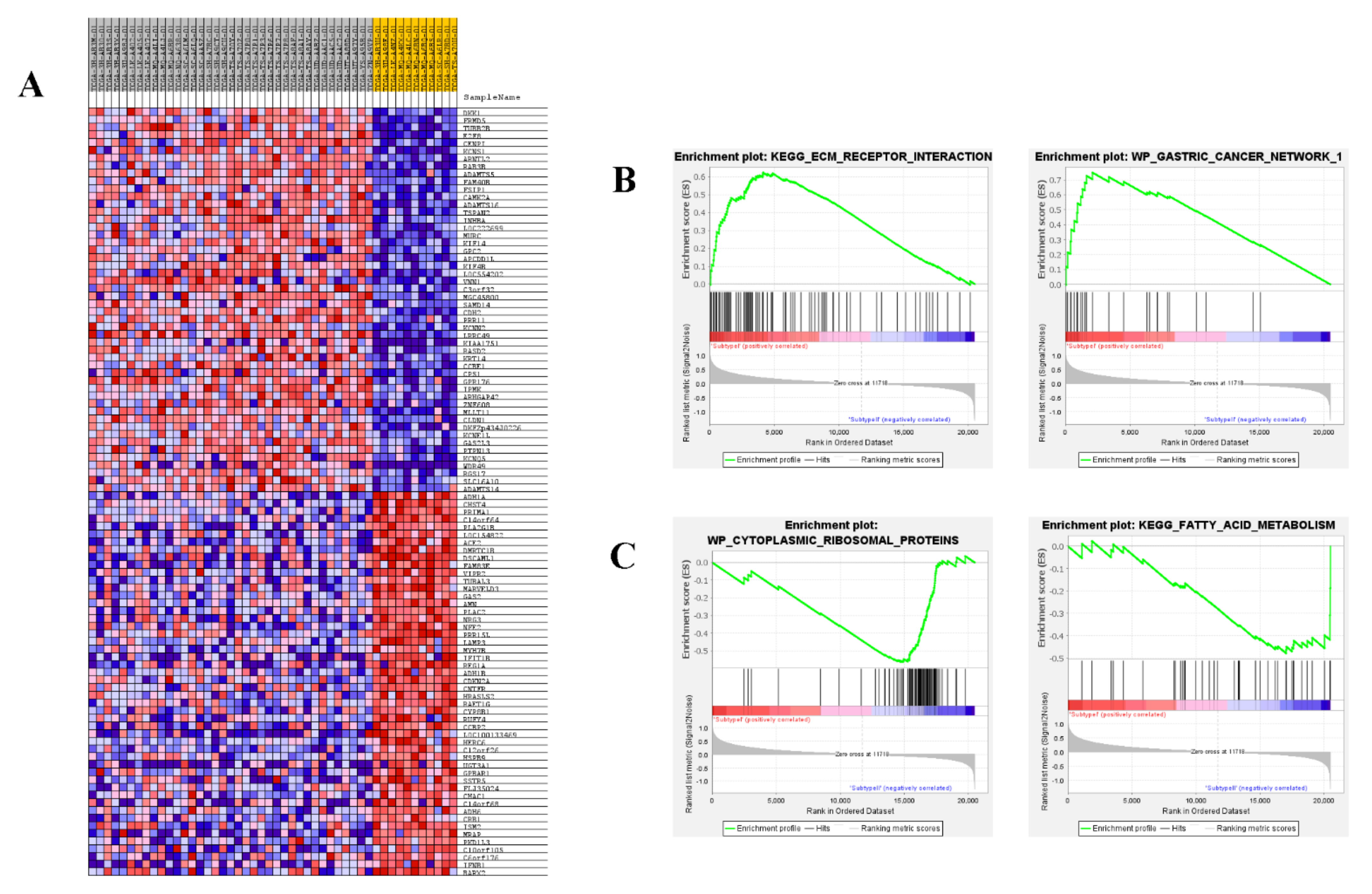
3.1. Consensus Clustering Identified Two Different Empm Molecular Subtypes
3.2. Validation of eMPM Molecular Subtypes by Independent Dataset
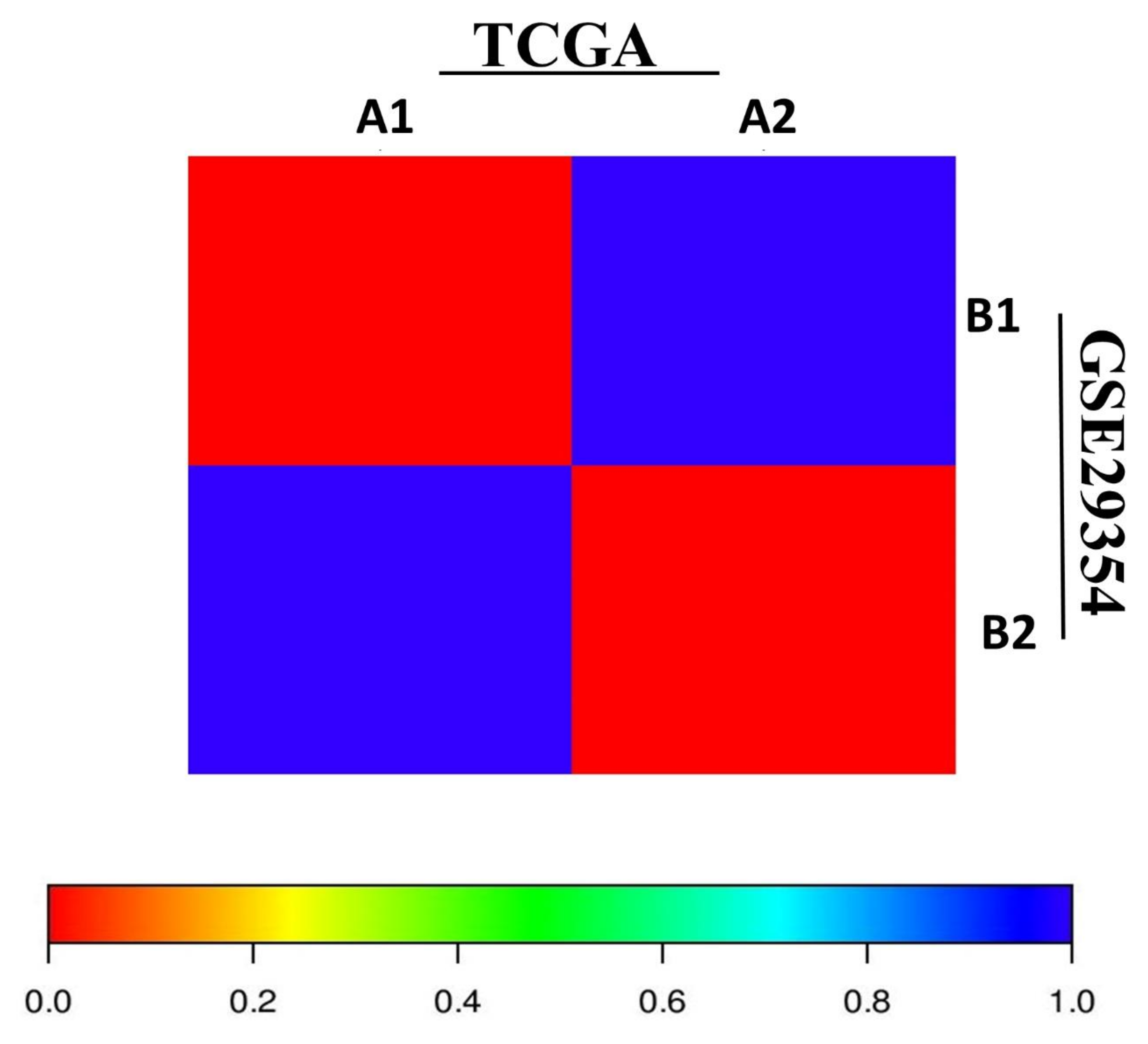
3.3. SubMap Analysis of Molecular Subtypes in Independent Empm Cohorts
3.4. Clinical Characteristics of eMPM Molecular Subtypes
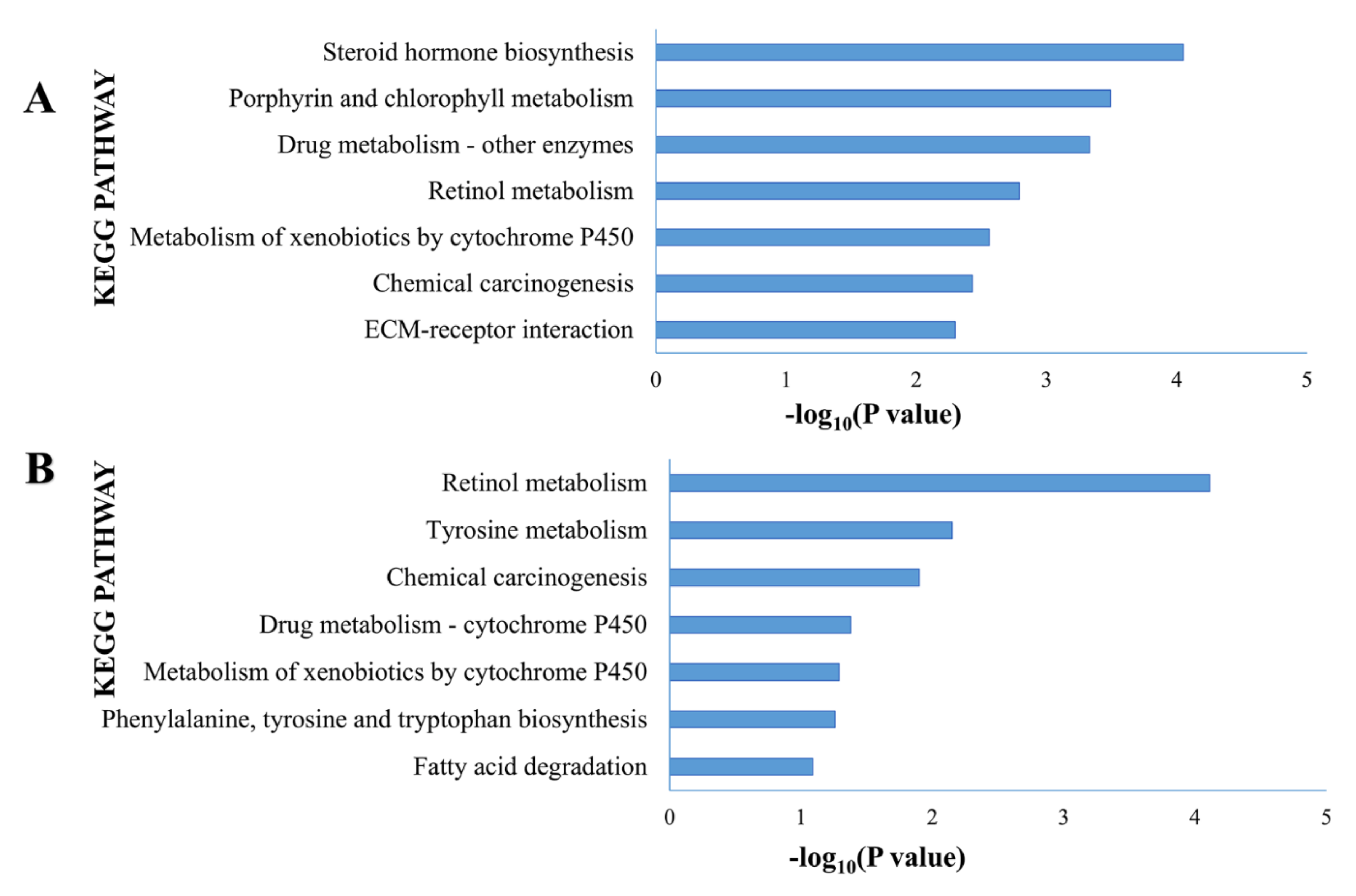
3.5. Functional Analysis of eMPM Subtype-Specific Genes
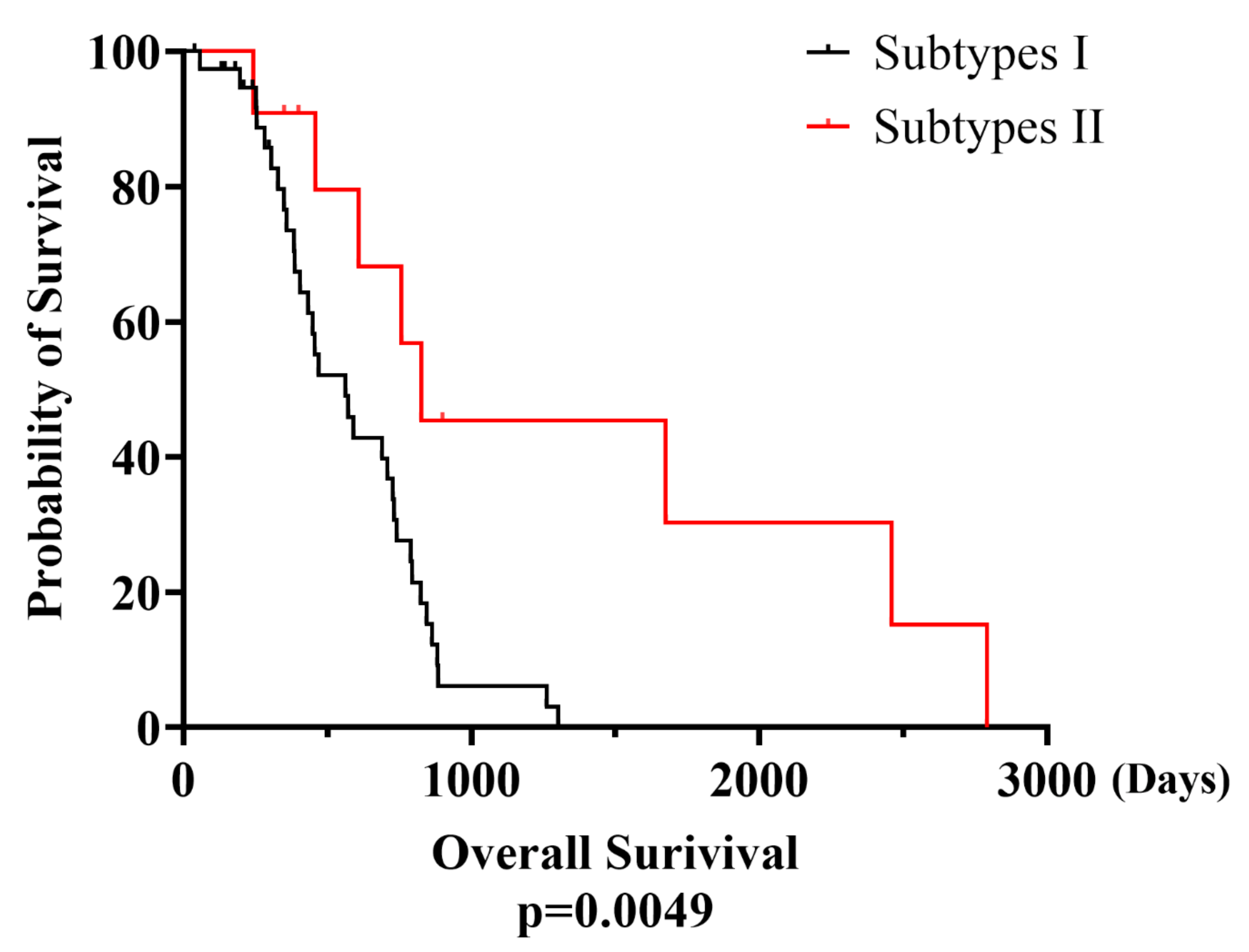
3.6. Clinical Implication of eMPM Subtyping
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ismail-Khan, R.; Robinson, L.A.; Williams, C.C., Jr.; Garrett, C.; Bepler, G.; Simon, G.R. Malignant pleural mesothelioma: A comprehensive review. Cancer Control. 2006, 13, 255–263. [Google Scholar] [CrossRef]
- Segura-González, M.; Urias-Rocha, J.; Castelán-Pedraza, J. Malignant Mesothelioma of the Tunica Vaginalis: A Rare Neoplasm—Case Report and Literature Review. Clin. Genitourin. Cancer 2015, 13, e401–e405. [Google Scholar] [CrossRef]
- Jasani, B.; Gibbs, A. Mesothelioma Not Associated With Asbestos Exposure. Arch. Pathol. Lab. Med. 2012, 136, 262–267. [Google Scholar] [CrossRef]
- Røe, O.D.; Stella, G. Malignant pleural mesothelioma: History, controversy and future of a manmade epidemic. Eur. Respir. Rev. 2015, 24, 115–131. [Google Scholar] [CrossRef]
- Gazdar, A.F.; Carbone, M. Molecular Pathogenesis of Malignant Mesothelioma and its Relationship to Simian Virus 40. Clin. Lung Cancer 2003, 5, 177–181. [Google Scholar] [CrossRef]
- Carbone, M. Simian virus 40 and human tumors: It is time to study mechanisms. J. Cell. Biochem. 2000, 76, 189–193. [Google Scholar] [CrossRef]
- Carbone, M.; Rizzo, P.; Pass, H. Simian virus 40: The link with human malignant mesothelioma is well established. Anticancer Res. 2000, 20, 875–877. [Google Scholar] [PubMed]
- Bolognesi, C.; Filiberti, R.; Neri, M.; Perrone, E.; Landini, E.; Canessa, P.A.; Simonassi, C.; Cerrano, P.G.; Mutti, L.; Puntoni, R. High frequency of micronuclei in peripheral blood lymphocytes as index of susceptibility to pleural malignant mesothe-lioma. Cancer Res. 2002, 62, 5418–5419. [Google Scholar]
- Jaklitsch, M.M.T.; Grondin, M.S.C.; Sugarbaker, M.D.J. Treatment of Malignant Mesothelioma. World J. Surg. 2001, 25, 210–217. [Google Scholar] [CrossRef]
- Robinson, B.W.; Lake, R.A. Advances in Malignant Mesothelioma. New Engl. J. Med. 2005, 353, 1591–1603. [Google Scholar] [CrossRef]
- Van Allen, E.M.; Wagle, N.; Stojanov, P.; Perrin, D.L.; Cibulskis, K.; Marlow, S.; Jane-Valbuena, J.; Friedrich, D.C.; Kryukov, G.; Carter, S.L.; et al. Whole-exome sequencing and clinical interpretation of formalin fixed, paraffin-embedded tumor samples to guide precision cancer medicine. Nat Med. 2014, 20, 682–688. [Google Scholar] [CrossRef] [PubMed]
- Meirson, T.; Pentimalli, F.; Cerza, F.; Baglio, G.; Gray, S.G.; Correale, P.; Krstic-Demonacos, M.; Markel, G.; Giordano, A.; Bomze, D.; et al. Comparison of 3 Randomized Clinical Trials of Frontline Therapies for Malignant Pleural Mesothelioma. JAMA Netw. Open. 2022, 5, e221490. [Google Scholar] [CrossRef]
- Messori, A.; Trippoli, S. Current treatments for inoperable mesothelioma: Indirect comparisons based on individual patient data reconstructed retrospectively from 4 trials. J. Chemother. 2022, 12, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Kerrigan, K.; Jo, Y.; Chipman, J.; Haaland, B.; Puri, S.; Akerley, W.; Patel, S. A Real-World Analysis of the Use of Systemic Therapy in Malignant Pleural Mesothelioma and the Differential Impacts on Overall Survival by Practice Pattern. JTO Clin. Res. Rep. 2022, 3, 100280. [Google Scholar] [CrossRef] [PubMed]
- Baumann, F.; Flores, E.; Napolitano, A.; Kanodia, S.; Taioli, E.; Pass, H.; Yang, H.; Carbone, M. Mesothelioma patients with germline BAP1 mutations have 7-fold improved long-term survival. Carcinogenesis 2015, 36, 76–81. [Google Scholar] [CrossRef]
- Ricciardi, S.; Cardillo, G.; Zirafa, C.C.; Carleo, F.; Facciolo, F.; Fontanini, G.; Mutti, L.; Melfi, F. Surgery for malignant pleural mesothelioma: An international guidelines review. J. Thorac. Dis. 2018, 10, S285–S292. [Google Scholar] [CrossRef]
- Bononi, A.; Napolitano, A.; Pass, H.I.; Yang, H.; Carbone, M. Latest developments in our understanding of the patho-genesis of mesothelioma and the design of targeted therapies. Expert Rev. Respir. Med. 2015, 9, 633–654. [Google Scholar] [CrossRef]
- Altomare, D.A.; Menges, C.W.; Pei, J.; Zhang, L.; Skele-Stump, K.L.; Carbone, M.; Kane, A.B.; Testa, J.R. Activated TNF-alpha/NF-kappaB signaling via down- regulation of Fas-associated factor 1 in asbestos-induced mesotheliomas from Arf knockout mice. Proc. Natl. Acad. Sci. USA 2009, 106, 3420–3425. [Google Scholar] [CrossRef]
- Hillegass, J.M.; Shukla, A.; Lathrop, S.A.; MacPherson, M.B.; Beuschel, S.L.; Butnor, K.J.; Testa, J.R.; Pass, H.I.; Carbone, M.; Steele, C.; et al. Inflammation precedes the development of human malignant meso-theliomas in a SCID mouse xenograft model. Ann. N. Y. Acad. Sci. 2010, 1203, 7–14. [Google Scholar] [CrossRef]
- Guo, G.; Chmielecki, J.; Goparaju, C.; Heguy, A.; Dolgalev, I.; Carbone, M.; Seepo, S.; Meyerson, M.; Pass, H.I. Whole-exome sequencing reveals frequent genetic alterations in BAP1, NF2, CDKN2A, and CUL1 in malignant pleural mesothelioma. Cancer Res. 2015, 75, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Bueno, R.; Stawiski, E.W.; Goldstein, L.D.; Durinck, S.; De Rienzo, A.; Modrusan, Z.; Gnad, F.; Nguyen, T.T.; Jaiswal, B.S.; Chirieac, L.R.; et al. Comprehensive genomic analysis of malignant pleural mesothelioma identifies recurrent mutations, gene fusions and splicing alterations. Nat. Genet. 2016, 48, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Nasu, M.; Emi, M.; Pastorino, S.; Tanji, M.; Powers, A.; Luk, H.; Baumann, F.; Zhang, Y.-A.; Gazdar, A.; Kanodia, S.; et al. High Incidence of Somatic BAP1 Alterations in Sporadic Malignant Mesothelioma. J. Thorac. Oncol. 2015, 10, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, Z.; Wan, Y.; Guan, D.; Ni, S.; Wang, L.; Zhang, Z.; Liu, J.; Liang, C.; Yu, Y.; Lu, A.; et al. A Loop-Based and AGO-Incorporated Virtual Screening Model Targeting AGO-Mediated miRNA–mRNA Interactions for Drug Discovery to Rescue Bone Phenotype in Genetically Modified Mice. Adv. Sci. 2020, 7, 1903451. [Google Scholar] [CrossRef]
- Travis, W.D.; Brambilla, E.; Burke, A.P.; Marx, A.; Nicholson, A.G. WHO Classification of Tumours of the Lung, Pleura, Thymus and Heart; International Agency for Research on Cancer (IARC): Lyon, France, 2015. [Google Scholar]
- Baas, P.; Fennell, D.; Kerr, K.M.; Van Schil, P.E.; Haas, R.L.; Peters, S. Malignant pleural mesothelioma: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2015, 26, v31–v39. [Google Scholar] [CrossRef] [PubMed]
- Husain, A.N.; Colby, T.; Ordonez, N.; Krausz, T.; Attanoos, R.; Beasley, M.B.; Borczuk, A.C.; Butnor, K.; Cagle, P.T.; Chirieac, L.R.; et al. Guidelines for Pathologic Diagnosis of Malignant Mesothelioma: 2012 Update of the Consensus Statement from the International Mesothelioma Interest Group. Arch. Pathol. Lab. Med. 2013, 137, 647–667. [Google Scholar] [CrossRef] [PubMed]
- Bertucci, F.; Finetti, P.; Rougemont, J.; Charafe-Jauffret, E.; Cervera, N.; Tarpin, C.; Nguyen, C.; Xerri, L.; Houlgatte, R.; Jacquemier, J.; et al. Gene Expression Profiling Identifies Molecular Subtypes of Inflammatory Breast Cancer. Cancer Res. 2005, 65, 2170–2178. [Google Scholar] [CrossRef] [PubMed]
- Sorlie, T.; Perou, C.M.; Tibshirani, R.; Aas, T.; Geisler, S.; Johnsen, H.; Hastie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; et al. Gene expression patterns of breast carcinomas distinguish tumor subclasses with clinical implications. Proc. Natl. Acad. Sci. USA 2001, 98, 10869–10874. [Google Scholar] [CrossRef]
- Lei, Z.; Tan, I.; Das, K.; Deng, N.; Zouridis, H.; Pattison, S.; Chua, C.; Feng, Z.; Guan, Y.K.; Ooi, C.-H.; et al. Identification of Molecular Subtypes of Gastric Cancer With Different Responses to PI3-Kinase Inhibitors and 5-Fluorouracil. Gastroenterology 2013, 145, 554–565. [Google Scholar] [CrossRef] [PubMed]
- Cristescu, R.; Lee, J.; Nebozhyn, M.; Kim, K.-M.; Ting, J.C.; Wong, S.S.; Liu, J.; Yue, Y.G.; Wang, J.; Yu, K.; et al. Molecular analysis of gastric cancer identifies subtypes associated with distinct clinical outcomes. Nat. Med. 2015, 21, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Saddozai, U.A.K.; Wang, F.; Akbar, M.U.; Zhang, L.; An, Y.; Zhu, W.; Xie, L.; Li, Y.; Ji, X.; Guo, X. Identification of Clinical Relevant Molecular Subtypes of Pheochromocytoma. Front. Endocrinol. 2021, 12, 607. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Yan, Z.; Lv, J.; Xin, J.; Dang, Y.; Sun, X.; An, Y.; Qi, Y.; Jiang, Q.; Zhu, W.; et al. Gene Expression Profiling Reveals Distinct Molecular Subtypes of Esophageal Squamous Cell Carcinoma in Asian Populations. Neoplasia 2019, 21, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Jo, V.Y.; Mills, A.M.; Zhu, S.X.; Lee, C.-H.; Espinosa, I.; Nucci, M.R.; Varma, S.; Forgó, E.; Hastie, T.; et al. Clinically Relevant Molecular Subtypes in Leiomyosarcoma. Clin. Cancer Res. 2015, 21, 3501–3511. [Google Scholar] [CrossRef] [PubMed]
- Goldhirsch, A.; Wood, W.C.; Coates, A.S.; Gelber, R.D.; Thürlimann, B.; Senn, H.J. Strategies for subtypes–dealing with the diversity of breast cancer: Highlights of the St. Gallen international expert consensus on the primary therapy of early breast cancer. Ann. Oncol. 2011, 22, 1736–1747. [Google Scholar] [CrossRef]
- Wilkerson, M.D.; Hayes, D.N. ConsensusClusterPlus: A class discovery tool with confidence assessments and item tracking. Bioinformatics 2010, 26, 1572–1573. [Google Scholar] [CrossRef] [PubMed]
- Rousseeuw, P.J. Silhouettes: A graphical aid to the interpretation and validation of cluster analysis. J. Comput. Appl. Math. 1987, 20, 53–65. [Google Scholar] [CrossRef]
- Hoshida, Y.; Brunet, J.-P.; Tamayo, P.; Golub, T.R.; Mesirov, J.P. Subclass Mapping: Identifying Common Subtypes in Independent Disease Data Sets. PLoS ONE 2007, 2, e1195. [Google Scholar] [CrossRef]
- Tusher, V.G.; Tibshirani, R.; Chu, G. Significance analysis of microarrays applied to the ionizing radiation response. Proc. Natl. Acad. Sci. USA 2001, 98, 5116–5121. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Tibshirani, R. Finding consistent patterns: A nonparametric approach for identifying differential expression in RNA-Seq data. Stat. Methods Med. Res. 2011, 22, 519–536. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef]
- Moroni, M.; Veronese, S.; Benvenuti, S.; Marrapese, G.; Sartore-Bianchi, A.; Di Nicolantonio, F.; Gambacorta, M.; Siena, S.; Bardelli, A. Gene copy number for epidermal growth factor receptor (EGFR) and clinical response to antiEGFR treatment in colorectal cancer: A cohort study. Lancet Oncol. 2005, 6, 279–286. [Google Scholar] [CrossRef]
- Lehmann, B.D.; Bauer, J.A.; Chen, X.; Sanders, M.E.; Chakravarthy, A.B.; Shyr, Y.; Pietenpol, J.A. Identification of human triple-negative breast cancer subtypes and preclinical models for selection of targeted therapies. J. Clin. Investig. 2011, 121, 2750–2767. [Google Scholar] [CrossRef] [PubMed]
- Paez, J.G.; Jänne, P.A.; Lee, J.C.; Tracy, S.; Greulich, H.; Gabriel, S.; Herman, P.; Kaye, F.J.; Lindeman, N.; Boggon, T.J.; et al. EGFR Mutations in Lung Cancer: Correlation with Clinical Response to Gefitinib Therapy. Science 2004, 304, 1497–1500. [Google Scholar] [CrossRef] [PubMed]
- Shavelle, R.; Vavra-Musser, K.; Lee, J.; Brooks, J. Life Expectancy in Pleural and Peritoneal Mesothelioma. Lung Cancer Int. 2017, 2017, 2782590. [Google Scholar] [CrossRef]
- Navin, N.; Kendall, J.; Troge, J.; Andrews, P.; Rodgers, L.; McIndoo, J.; Cook, K.; Stepansky, A.; Levy, D.; Esposito, D.; et al. Tumor evolution inferred by single cell sequencing. Nature 2011, 472, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Gerlinger, M.; Rowan, A.J.; Horswell, S.; Larkin, J.; Endesfelder, D.; Gronroos, E.; Martinez, P.; Matthews, N.; Stewart, A.; Tarpey, P.; et al. Intratumor heterogeneity and branched evolution revealed by mul-tiregion sequencing. N. Engl. J. Med. 2012, 366, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Piccart-Gebhart, M.J.; Procter, M.; Leyland-Jones, B.; Goldhirsch, A.; Untch, M.; Smith, I.; Gianni, L.; Baselga, J.; Bell, R.; Jack-isch, C.; et al. Trastuzumab after Ad-juvant Chemotherapy in HER2-Positive Breast Cancer. New Engl. J. Med. 2005, 353, 1659–1672. [Google Scholar] [CrossRef]
- de Reyniès, A.; Jaurand, M.-C.; Renier, A.; Couchy, G.; Hysi, I.; Elarouci, N.; Galateau-Sallé, F.; Copin, M.-C.; Hofman, P.; Cazes, A.; et al. Molecular Classification of Malignant Pleural Mesothelioma: Identification of a Poor Prognosis Subgroup Linked to the Epithelial-to-Mesenchymal Transition. Clin. Cancer Res. 2014, 20, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- De Santi, C.; Melaiu, O.; Bonotti, A.; Cascione, L.; Di Leva, G.; Foddis, R.; Cristaudo, A.; Lucchi, M.; Mora, M.; Truini, A.; et al. Deregulation of miRNAs in malignant pleural mesothelioma is associated with prognosis and suggests an alteration of cell metabolism. Sci. Rep. 2017, 7, 3140. [Google Scholar] [CrossRef] [PubMed]
- Ranzato, E.; Martinotti, S.; Magnelli, V.; Murer, B.; Biffo, S.; Mutti, L.; Burlando, B. Epigallocatechin-3-gallate induces mesothelioma cell deathviaH2O2−dependent T-type Ca2+channel opening. J. Cell Mol. Med. 2012, 16, 2667–2678. [Google Scholar] [CrossRef] [PubMed]
- Manente, A.G.; Valenti, D.; Pinton, G.; Jithesh, P.V.; Daga, A.; Rossi, L.; Gray, S.; O’Byrne, K.J.; Fennell, D.A.; Vacca, R.A.; et al. Estrogen receptor β activation impairs mitochondrial oxidative metabolism and affects malignant mesothelioma cell growth in vitro and in vivo. Oncogenesis 2013, 2, e72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, B.; Wu, W.; Li, Y.; Hoppe, D.; Stannek, P.; Glinka, A.; Niehrs, C. LDL-receptor-related protein 6 is a receptor for Dickkopf proteins. Nature 2001, 411, 321–325. [Google Scholar] [CrossRef]
- Zhu, G.; Song, J.; Chen, W.; Yuan, D.; Wang, W.; Chen, X.; Liu, H.; Su, H.; Zhu, J. Expression and Role of Dickkopf-1 (Dkk1) in Tumors: From the Cells to the Patients. Cancer Manag. Res. 2021, 13, 659–675. [Google Scholar] [CrossRef]
- Yu, B.; Yang, X.; Xu, Y.; Yao, G.; Shu, H.; Lin, B.; Hood, L.; Wang, H.; Yang, S.; Gu, J.; et al. Elevated expression of DKK1 is associated with cytoplasmic/nuclear β-catenin accumulation and poor prognosis in hepatocellular carcinomas. J. Hepatol. 2009, 50, 948–957. [Google Scholar] [CrossRef] [PubMed]
- Tung, E.K.-K.; Mak, C.K.-M.; Fatima, S.; Lo, R.C.-L.; Zhao, H.; Zhang, C.; Dai, H.; Poon, R.T.-P.; Yuen, M.-F.; Lai, C.-L.; et al. Clinicopathological and prognostic significance of serum and tissue Dickkopf-1 levels in human hepatocellular carcinoma. Liver Int. 2011, 31, 1494–1504. [Google Scholar] [CrossRef] [PubMed]
- Sato, N.; Yamabuki, T.; Takano, A.; Koinuma, J.; Aragaki, M.; Masuda, K.; Ishikawa, N.; Kohno, N.; Ito, H.; Miyamoto, M.; et al. Wnt Inhibitor Dickkopf-1 as a Target for Passive Cancer Immunotherapy. Cancer Res. 2010, 70, 5326–5336. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.J.; Wan, X.Z.; Yu, B. Expression of Dickkopf (Dkk1) in liver cancer and in multiple human cancer cell lines. Zhongliu 2006, 26, 1109–1112. [Google Scholar]
- Qin, W.X.; Zhang, H.T.; Yu, Y.J. Uses of DKK-1 protein in diagnosis of cancer. Patent WO/2007/104181A1, 2007. [Google Scholar]
- Kim, J.; Hu, Z.; Cai, L.; Li, K.; Choi, E.; Faubert, B.; Bezwada, D.; Rodriguez-Canales, J.; Villalobos, P.; Lin, Y.-F.; et al. CPS1 maintains pyrimidine pools and DNA synthesis in KRAS/LKB1-mutant lung cancer cells. Nature 2017, 546, 168–172. [Google Scholar] [CrossRef] [PubMed]
- Pham-Danis, C.; Gehrke, S.; Danis, E.; Rozhok, A.I.; Daniels, M.W.; Gao, D.; Collins, C.; Di Paola, J.T.; D’Alessandro, A.; DeGregori, J. Urea Cycle Sustains Cellular Energetics upon EGFR Inhibition in EGFR-Mutant NSCLC. Mol. Cancer Res. 2019, 17, 1351–1364. [Google Scholar] [CrossRef]
- Çeliktas, M.; Tanaka, I.; Chandra Tripathi, S.; Fahrmann, J.F.; Aguilar-Bonavides, C.; Villalobos, P.; Delgado, O.; Dhillon, D.; Dennison, J.B..; Ostrin, E.J.; et al. Role of CPS1 in Cell Growth, Metabolism and Prognosis in LKB1-Inactivated Lung Adeno-carcinoma. J. Natl. Cancer Inst. 2017, 109, 1–9. [Google Scholar]
- Yao, S.; Nguyen, T.-V.; Rolfe, A.; Agrawal, A.A.; Ke, J.; Peng, S.; Colombo, F.; Yu, S.; Bouchard, P.; Wu, J.; et al. Small Molecule Inhibition of CPS1 Activity through an Allosteric Pocket. Cell Chem. Biol. 2020, 27, 259–268.e5. [Google Scholar] [CrossRef] [PubMed]
- Nagelkerke, A.; Bussink, J.; Mujcic, H.; Wouters, B.G.; Lehmann, S.; Sweep, F.C.; Span, P.N. Hypoxia stimulates migration of breast cancer cells via the PERK/ATF4/LAMP3-arm of the unfolded protein response. Breast Cancer Res. 2013, 15, R2. [Google Scholar] [CrossRef] [PubMed]
- Nagelkerke, A.; Mujcic, H.; Bussink, J.; Wouters, B.G.; Van Laarhoven, H.W.M.; Sweep, F.C.G.J.; Span, P.N. Hypoxic regulation and prognostic value of LAMP3 expression in breast cancer. Cancer 2011, 117, 3670–3681. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.; Chen, Y.; Liu, D.; Li, F.; Li, X.; Jia, W. High Expression of LAMP3 Is a Novel Biomarker of Poor Prognosis in Patients with Esophageal Squamous Cell Carcinoma. Int. J. Mol. Sci. 2015, 16, 17655–17667. [Google Scholar] [CrossRef]
- Liu, K.; He, Q.; Liao, G.; Han, J. Identification of critical genes and gene interaction networks that mediate osteosarcoma metastasis to the lungs. Exp. Ther. Med. 2015, 10, 1796–1806. [Google Scholar] [CrossRef]
- Grosso, S.; Marini, A.; Gyuraszova, K.; Voorde, J.V.; Sfakianos, A.; Garland, G.D.; Tenor, A.R.; Mordue, R.; Chernova, T.; Morone, N.; et al. The pathogenesis of mesothelioma is driven by a dysregulated translatome. Nat. Commun. 2021, 12, 1–17. [Google Scholar] [CrossRef]
- Bendris, N.; Lemmers, B.; Blanchard, J.M. Cell cycle, cytoskeleton dynamics and beyond: The many functions of cyclins and CDK inhibitors. Cell Cycle 2015, 14, 1786–1798. [Google Scholar] [CrossRef]
- Sapoznik, S.; Aviel-Ronen, S.; Bahar-Shany, K.; Zadok, O.; Levanon, K. CCNE1 expression in high grade serous carcinoma does not correlate with chemoresistance. Oncotarget 2017, 8, 62240–62247. [Google Scholar] [CrossRef]
- Fu, Y.-P.; Kohaar, I.; Moore, L.E.; Lenz, P.; Figueroa, J.D.; Tang, W.; Porter-Gill, P.; Chatterjee, N.; Scott-Johnson, A.; Garcia-Closas, M.; et al. The 19q12 Bladder Cancer GWAS Signal: Association with Cyclin E Function and Aggressive Disease. Cancer Res. 2014, 74, 5808–5818. [Google Scholar] [CrossRef]
- Pontoriero, A.; Iatì, G.; Aiello, D.; Pergolizzi, S. Stereotactic Radiotherapy in the Retreatment of Recurrent Cervical Cancers, Assessment of Toxicity, and Treatment Response. Technol. Cancer Res. Treat. 2016, 15, 759–765. [Google Scholar] [CrossRef]
- Bani-Hani, K.E.; Almasri, N.M.; Khader, Y.S.; Sheyab, F.M.; Karam, H.N. Combined Evaluation of Expressions of Cyclin E and p53 Proteins as Prognostic Factors for Patients with Gastric Cancer. Clin. Cancer Res. 2005, 11, 1447–1453. [Google Scholar] [CrossRef] [PubMed]
- Etemadmoghadam, D.; Au-Yeung, G.; Wall, M.; Mitchell, C.; Kansara, M.; Loehrer, E.; Batzios, C.; George, J.; Ftouni, S.; Weir, B.A.; et al. Resistance to CDK2 Inhibitors Is Associated with Selection of Polyploid Cells in CCNE1-Amplified Ovarian Cancer. Clin. Cancer Res. 2013, 19, 5960–5971. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, N.; Nakayama, K.; Shamima, Y.; Ishikawa, M.; Katagiri, A.; Iida, K.; Miyazaki, K. Gene amplification CCNE1 is related to poor survival and potential therapeutic target in ovarian cancer. Cancer 2010, 116, 2621–2634. [Google Scholar] [CrossRef]
- Yang, L.; Fang, D.; Chen, H.; Lu, Y.; Dong, Z.; Ding, H.-F.; Jing, Q.; Su, S.-B.; Huang, S. Cyclin-dependent kinase 2 is an ideal target for ovary tumors with elevated cyclin E1 expression. Oncotarget 2015, 6, 20801–20812. [Google Scholar] [CrossRef]
- Etemadmoghadam, D.; Weir, B.A.; Au-Yeung, G.; Alsop, K.; Mitchell, G.; George, J.; Davis, S.; D’Andrea, A.D.; Simpson, K.; Hahn, W.C.; et al. Synthetic lethality between CCNE1 amplification and loss of BRCA1. Proc. Natl. Acad. Sci. USA 2013, 110, 19489–19494. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.K.; LaPlant, B.; Chng, W.J.; Zonder, J.; Callander, N.; Fonseca, R.; Fruth, B.; Roy, V.; Erlichman, C.; Stewart, A.K. Dinaciclib, a novel CDK inhibitor, demonstrates encouraging single-agent activity in patients with relapsed multiple mye-loma. Blood 2015, 125, 443–448. [Google Scholar] [CrossRef]
- Mitri, Z.; Karakas, C.; Wei, C.; Briones, B.; Simmons, H.; Ibrahim, N.; Alvarez, R.; Murray, J.L.; Keyomarsi, K.; Moulder, S. A phase 1 study with dose expansion of the CDK inhibitor dinaciclib (SCH 727965) in combination with epirubicin in patients with metastatic triple negative breast cancer. Investig. New Drugs 2015, 33, 890–894. [Google Scholar] [CrossRef]
- Wood, L.D.; Calhoun, E.S.; Silliman, N.; Ptak, J.; Szabo, S.; Powell, S.M.; Riggins, G.J.; Wang, T.L.; Yan, H.; Gazdar, A.; et al. Somatic mutations of GUCY2F, EPHA3, and NTRK3 in human cancers. Hum. Mutat. 2006, 27, 1060–1061. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, W.; Qian, J.; Wu, J.; Jiang, L.; Ling, C. R-spondin family members as novel biomarkers and prognostic factors in lung cancer. Oncol. Lett. 2019, 18, 4008–4015. [Google Scholar] [CrossRef]
- Davies, H.; Hunter, C.; Smith, R.; Stephens, P.; Greenman, C.; Bignell, G.; Teague, J.; Butler, A.; Edkins, S.; Stevens, C.; et al. Somatic Mutations of the Protein Kinase Gene Family in Human Lung Cancer. Cancer Res. 2005, 65, 7591–7595. [Google Scholar] [CrossRef]
- Wang, K.; Yuen, S.T.; Xu, J.; Lee, S.P.; Yan, H.H.N.; Shi, S.T.; Siu, H.C.; Deng, S.; Chu, K.M.; Law, S.; et al. Whole-genome sequencing and comprehensive molecular profiling identify new driver mutations in gastric cancer. Nat. Genet. 2014, 46, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.; Sun, W.; Wang, X.; Xu, X.; Li, M.; Huang, D.; Xu, E.; Lai, M.; Zhang, H. RNF43 frameshift mutations contribute to tumourigenesis in right-sided colon cancer. Pathol. Res. Pr. 2019, 215, 152453. [Google Scholar] [CrossRef] [PubMed]
- Giannakis, M.; Hodis, E.; Mu, X.J.; Yamauchi, M.; Rosenbluh, J.; Cibulskis, K.; Saksena, G.; Lawrence, M.S.; Qian, Z.R.; Nishihara, R.; et al. RNF43 is frequently mutated in colorectal and endometrial cancers. Nat. Genet. 2014, 46, 1264–1266. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Hao, H.-X.; Growney, J.D.; Woolfenden, S.; Bottiglio, C.; Ng, N.; Lu, B.; Hsieh, M.H.; Bagdasarian, L.; Meyer, R.; et al. Inactivating mutations of RNF43 confer Wnt dependency in pancreatic ductal adenocarcinoma. Proc. Natl. Acad. Sci. USA 2013, 110, 12649–12654. [Google Scholar] [CrossRef] [PubMed]
- Ryland, G.L.; Hunter, S.M.; A Doyle, M.; Rowley, S.M.; Christie, M.; E Allan, P.; Bowtell, D.D.; Gorringe, K.L.; Campbell, I.G. RNF43 is a tumour suppressor gene mutated in mucinous tumours of the ovary. J. Pathol. 2012, 229, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Talpaz, M.; Shah, N.P.; Kantarjian, H.; Donato, N.; Nicoll, J.; Paquette, R.; Cortes, J.; O’Brien, S.; Nicaise, C.; Bleickardt, E.; et al. Dasatinib in Imatinib-Resistant Philadelphia Chromosome–Positive Leukemias. New Engl. J. Med. 2006, 354, 2531–2541. [Google Scholar] [CrossRef]
- Keating, G.M. Dasatinib: A Review in Chronic Myeloid Leukaemia and Ph+ Acute Lymphoblastic Leukaemia. Drugs 2016, 77, 85–96. [Google Scholar] [CrossRef]
- Proffitt, K.D.; Madan, B.; Ke, Z.; Pendharkar, V.; Ding, L.; Lee, M.A.; Hannoush, R.N.; Virshup, D.M. Pharmacological Inhibition of the Wnt Acyltransferase PORCN Prevents Growth of WNT-Driven Mammary Cancer. Cancer Res. 2013, 73, 502–507. [Google Scholar] [CrossRef]
- Aguilera, K.Y.; Dawson, D.W. WNT Ligand Dependencies in Pancreatic Cancer. Front. Cell Dev. Biol. 2021, 9, 671022. [Google Scholar] [CrossRef]
- Liu, J.; Pan, S.; Hsieh, M.H.; Ng, N.; Sun, F.; Wang, T.; Kasibhatla, S.; Schuller, A.G.; Li, A.G.; Cheng, D.; et al. Targeting Wnt-driven cancer through the inhibition of Porcupine by LGK974. Proc. Natl. Acad. Sci. USA 2013, 110, 20224–20229. [Google Scholar] [CrossRef]
- Madan, B.; Ke, Z.; Harmston, N.; Ho, S.Y.; O Frois, A.; Alam, J.; Jeyaraj, D.; Pendharkar, V.; Ghosh, K.; Virshup, I.H.; et al. Wnt addiction of genetically defined cancers reversed by PORCN inhibition. Oncogene 2015, 35, 2197–2207. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Cao, J.; Zhang, N.; Tu, M.; Xu, F.; Wei, S.; Chen, X.; Xu, Y. Identification of RSPO2 Fusion Mutations and Target Therapy Using a Porcupine Inhibitor. Sci. Rep. 2018, 8, 14244. [Google Scholar] [CrossRef] [PubMed]
- Bhamra, I.; Adams, N.; Armer, R.; Bingham, M.; McKeever, H.; Phillips, C.; Thompson, B.; Woodcock, S. Novel porcupine (PORCN) inhibitor RXC004: Evaluation in models of RNF43 loss of function cancers. J. Clin. Oncol. 2017, 35, e14094. [Google Scholar] [CrossRef]
- Xie, X.; You, M.; Meng, E.; Wang, S.; Niu, B.; Huang, W. Complete and durable response to crizotinib in a patient with malignant pleural mesothelioma harboring CD74-ROS1 fusion. J. Cancer Res. Clin. Oncol. 2022, 148, 2561–12566. [Google Scholar] [CrossRef]
- Witkiewicz, A.K.; Knudsen, K.E.; Dicker, A.P.; Knudsen, E.S. The meaning of p16(ink4a) expression in tumors: Functional sig-nificance, clinical associations and future developments. Cell Cycle. 2011, 10, 2497–2503. [Google Scholar] [CrossRef]
- Kobayashi, N.; Toyooka, S.; Yanai, H.; Soh, J.; Fujimoto, N.; Yamamoto, H.; Ichihara, S.; Kimura, K.; Ichimura, K.; Sano, Y.; et al. Frequent p16 inactivation by homozygous deletion or methylation is associated with a poor prognosis in Japanese patients with pleural mesothelioma. Lung Cancer 2008, 62, 120–125. [Google Scholar] [CrossRef]
- Bahnassy, A.A.; Zekri, A.-R.; Abou-Bakr, A.A.; El-Deftar, M.M.; El Bastawisy, A.; Sakr, M.A.; El-Sherif, G.M.; Gaafar, R.M. Aberrant expression of cell cycle regulatory genes predicts overall and disease free survival in malignant pleural mesothelioma patients. Exp. Mol. Pathol. 2012, 93, 154–161. [Google Scholar] [CrossRef]
- Chung, C.T.; Santos, G.D.C.; Hwang, D.M.; Ludkovski, O.; Pintilie, M.; Squire, J.A.; Tsao, M.S. FISH assay development for the detection of p16/CDKN2A deletion in malignant pleural mesothelioma. J. Clin. Pathol. 2010, 63, 630–634. [Google Scholar] [CrossRef] [PubMed]
- Dacic, S.; Kothmaier, H.; Land, S.; Shuai, Y.; Halbwedl, I.; Morbini, P.; Murer, B.; Comin, C.; Galateau-Salle, F.; Demirag, F.; et al. Prognostic significance of p16/cdkn2a loss in pleural malignant mesotheliomas. Virchows Arch. 2008, 453, 627–635. [Google Scholar] [CrossRef]
- Lopez-Rios, F.; Chuai, S.; Flores, R.; Shimizu, S.; Ohno, T.; Wakahara, K.; Illei, P.B.; Hussain, S.; Krug, L.; Zakowski, M.F.; et al. Global gene ex-pression profiling of pleural mesotheliomas: Overexpression of Aurora kinases and P16/CDKN2A deletion as prognostic factors and critical evaluation of microarray-based prognostic prediction. Cancer Res. 2006, 66, 2970–2979. [Google Scholar] [CrossRef]
- Takeda, M.; Kasai, T.; Enomoto, Y.; Takano, M.; Morita, K.; Kadota, E.; Nonomura, A. 9p21 deletion in the diagnosis of ma-lignant mesothelioma, using fluorescence in situ hybridization analysis. Pathol. Int. 2010, 60, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Yao, Y.; Yan, S.; Gao, R.; Lu, W.; He, W. Chiral Protein Supraparticles for Tumor Suppression and Synergistic Immunotherapy: An Enabling Strategy for Bioactive Supramolecular Chirality Construction. Nano Lett. 2020, 20, 5844–5852. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.S.; Crown, J.P.; Lang, I.; Boer, K.; Bondarenko, I.M.; Kulyk, S.O.; Ettl, J.; Patel, R.; Pinter, T.; Schmidt, M.; et al. The cyclin-dependent kinase 4/6 inhibitor palbociclib in combination with letrozole versus letrozole alone as first-line treatment of oestrogen receptor-positive, HER2-negative, advanced breast cancer (PALOMA-1/TRIO-18): A randomised phase 2 study. Lancet Oncol. 2015, 16, 25–35. [Google Scholar] [CrossRef]
- Dickson, M.A.; Schwartz, G.K.; Keohan, M.L.; D’Angelo, S.P.; Gounder, M.M.; Chi, P.; Antonescu, C.R.; Landa, J.; Qin, L.-X.; Crago, A.M.; et al. Progression-Free Survival Among Patients With Well-Differentiated or Dedifferentiated Liposarcoma Treated With CDK4 Inhibitor Palbociclib. JAMA Oncol. 2016, 2, 937–940. [Google Scholar] [CrossRef] [PubMed]
- Kuiper, J.L.; Ronden, M.I.; Becker, A.; Heideman, D.A.; Van Hengel, P.; Ylstra, B.; Thunnissen, E.; Smit, E.F. Trans-formation to a squamous cell carcinoma phenotype of an EGFR-mutated NSCLC patient after treatment with an EGFR-tyrosine kinase inhibitor. J. Clin. Pathol. 2015, 68, 320–321. [Google Scholar] [CrossRef] [PubMed]
- Gopalan, P.K.; Pinder, M.C.; Chiappori, A.; Ivey, A.M.; Gordillo Villegas, A.; Kaye, F.J. A phase II clinical trial of the CDK 4/6 inhibitor palbociclib (PD 0332991) in previously treated, advanced non-small cell lung cancer (NSCLC) patients with inac-tivated CDKN2A. J. Clin. Oncol. 2014, 32, 8077. [Google Scholar] [CrossRef]
- Levin, P.A.; Mayer, M.; Hoskin, S.; Sailors, J.; Oliver, D.H.; Gerber, D.E. Histologic transformation from adenocar-cinoma to squamous cell carcinoma as a mechanism of resistance to EGFR inhibition. J. Thorac. Oncol. 2015, 10, e86–e88. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subtype I | Subtype II | |
|---|---|---|
| 1979 gene sets | 1568 | 411 |
| FDR < 25% | 175 | 68 |
| Nominal p-value < 5% | 245 | 81 |
| Nominal p-value < 1% | 141 | 47 |
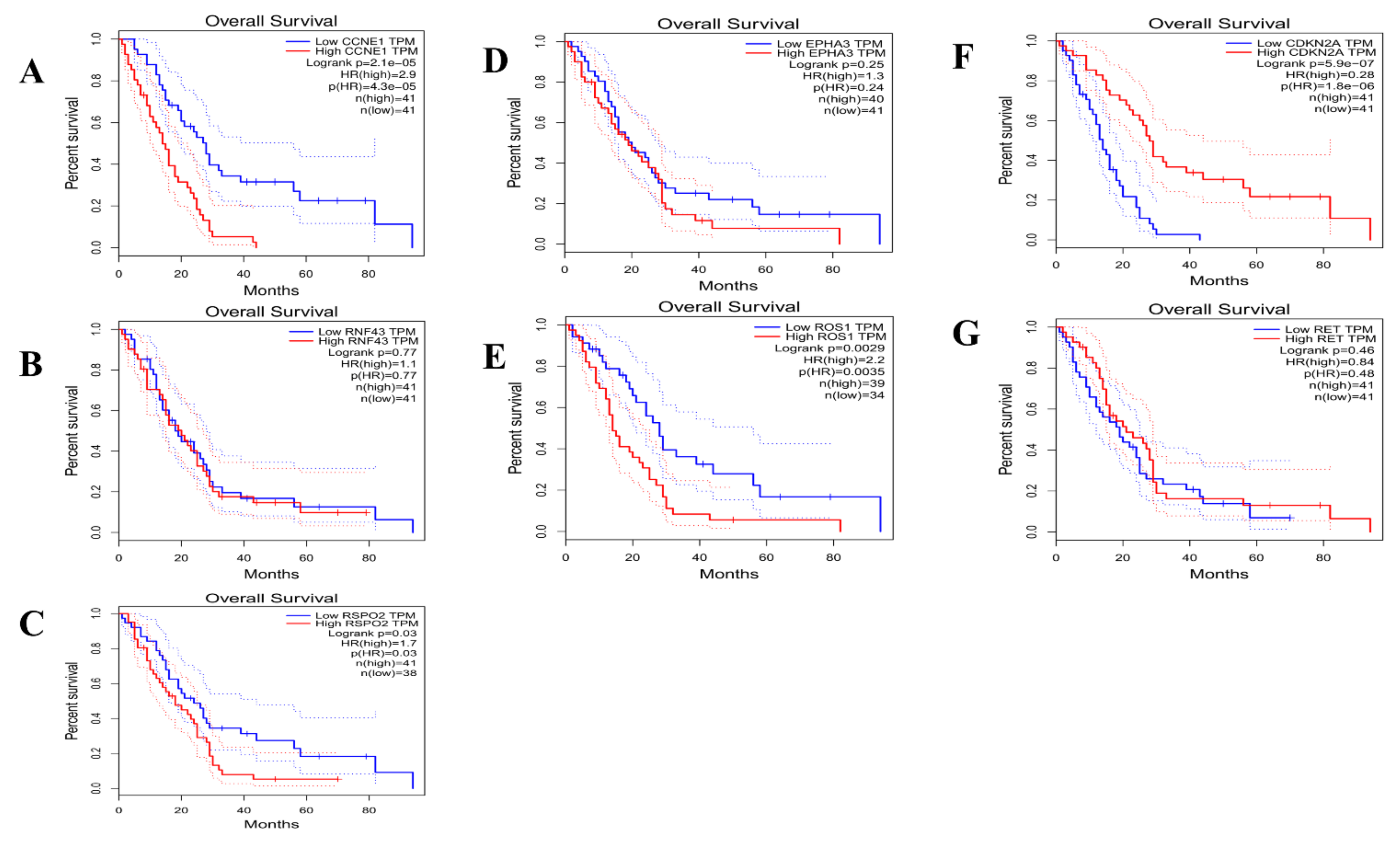
| Gene Overexpressed | Examples of Potential Therapeutic Agents | |
|---|---|---|
| CCNE1 | CDK2 inhibitor | |
| EPHA3 | Dasatinib, Ephrin inhibitors | |
| Subtype I | RNF43 | Porcupine inhibitors |
| ROS1 | Crizotinib | |
| RSPO2 | WNT inhibitors | |
| Subtype II | CDKN2A | CDK4/6 inhibitors |
| RET | Sorafenib, vandetinib, RET inhibitors | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saddozai, U.A.K.; Wang, F.; Khattak, S.; Akbar, M.U.; Badar, M.; Khan, N.H.; Zhang, L.; Zhu, W.; Xie, L.; Li, Y.; et al. Define the Two Molecular Subtypes of Epithelioid Malignant Pleural Mesothelioma. Cells 2022, 11, 2924. https://doi.org/10.3390/cells11182924
Saddozai UAK, Wang F, Khattak S, Akbar MU, Badar M, Khan NH, Zhang L, Zhu W, Xie L, Li Y, et al. Define the Two Molecular Subtypes of Epithelioid Malignant Pleural Mesothelioma. Cells. 2022; 11(18):2924. https://doi.org/10.3390/cells11182924
Chicago/Turabian StyleSaddozai, Umair Ali Khan, Fengling Wang, Saadullah Khattak, Muhammad Usman Akbar, Muhammad Badar, Nazeer Hussain Khan, Lu Zhang, Wan Zhu, Longxiang Xie, Yongqiang Li, and et al. 2022. "Define the Two Molecular Subtypes of Epithelioid Malignant Pleural Mesothelioma" Cells 11, no. 18: 2924. https://doi.org/10.3390/cells11182924