
Human Palatine Tonsils Are Linked to Alzheimer’s Disease through Function of Reservoir of Amyloid Beta Protein Associated with Bacterial Infection

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Background
2. Methods
2.1. Patients
2.2. Immunohistochemistry
2.3. Western Blots
2.4. Organoid Formation from Human Tonsillar Tissue
2.5. Treatment of Tonsil Organoids with S. aureus
2.6. Human Induced Pluripotent Stem Cell (iPSC) Culture
2.7. Generation of Human Brain Organoids and Treatment with S. aureus
2.8. Statistical and Reproducibility
3. Results
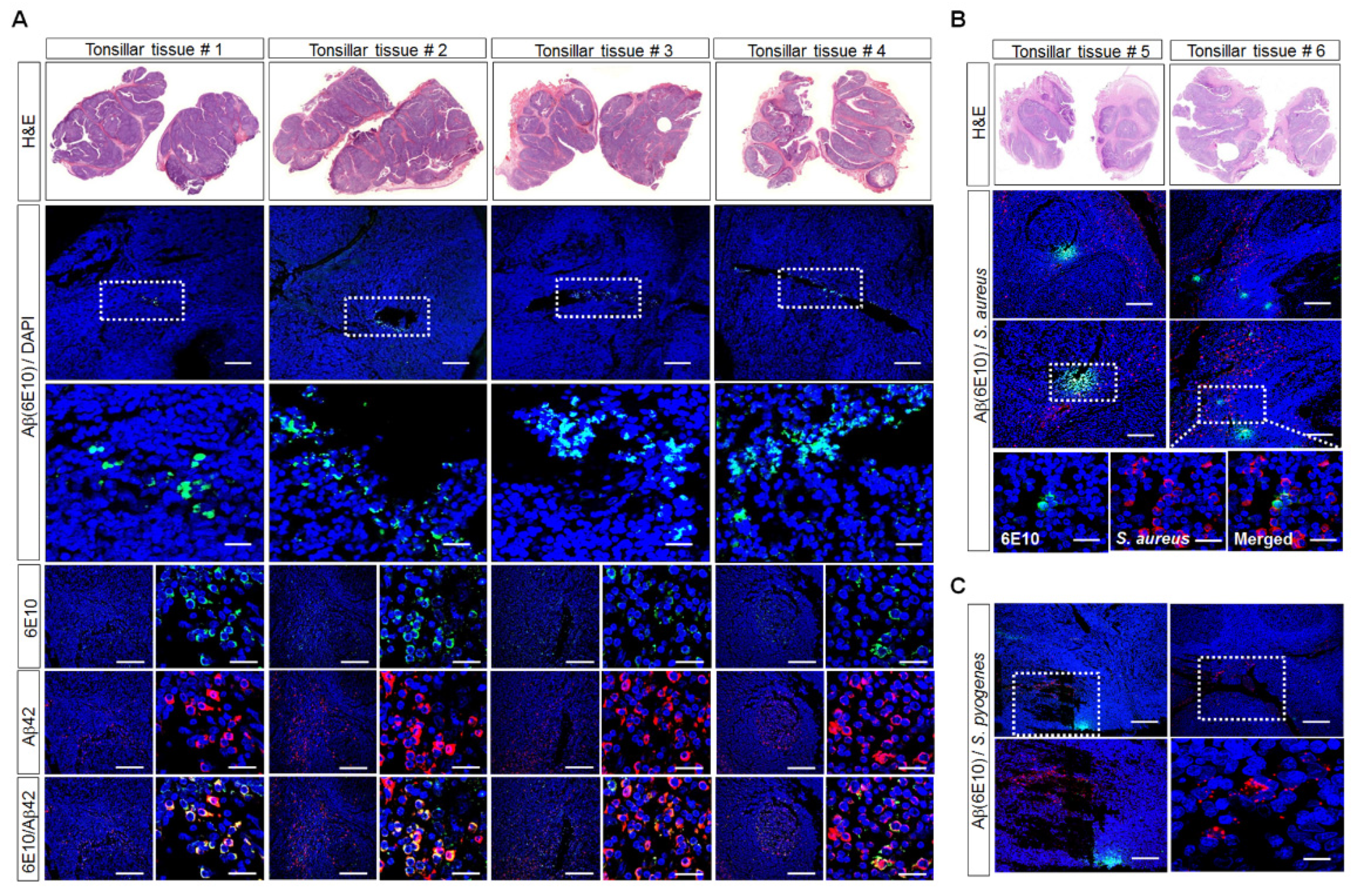
3.1. Aβ-Protein Deposition in Human Palatine-Tonsil Tissue
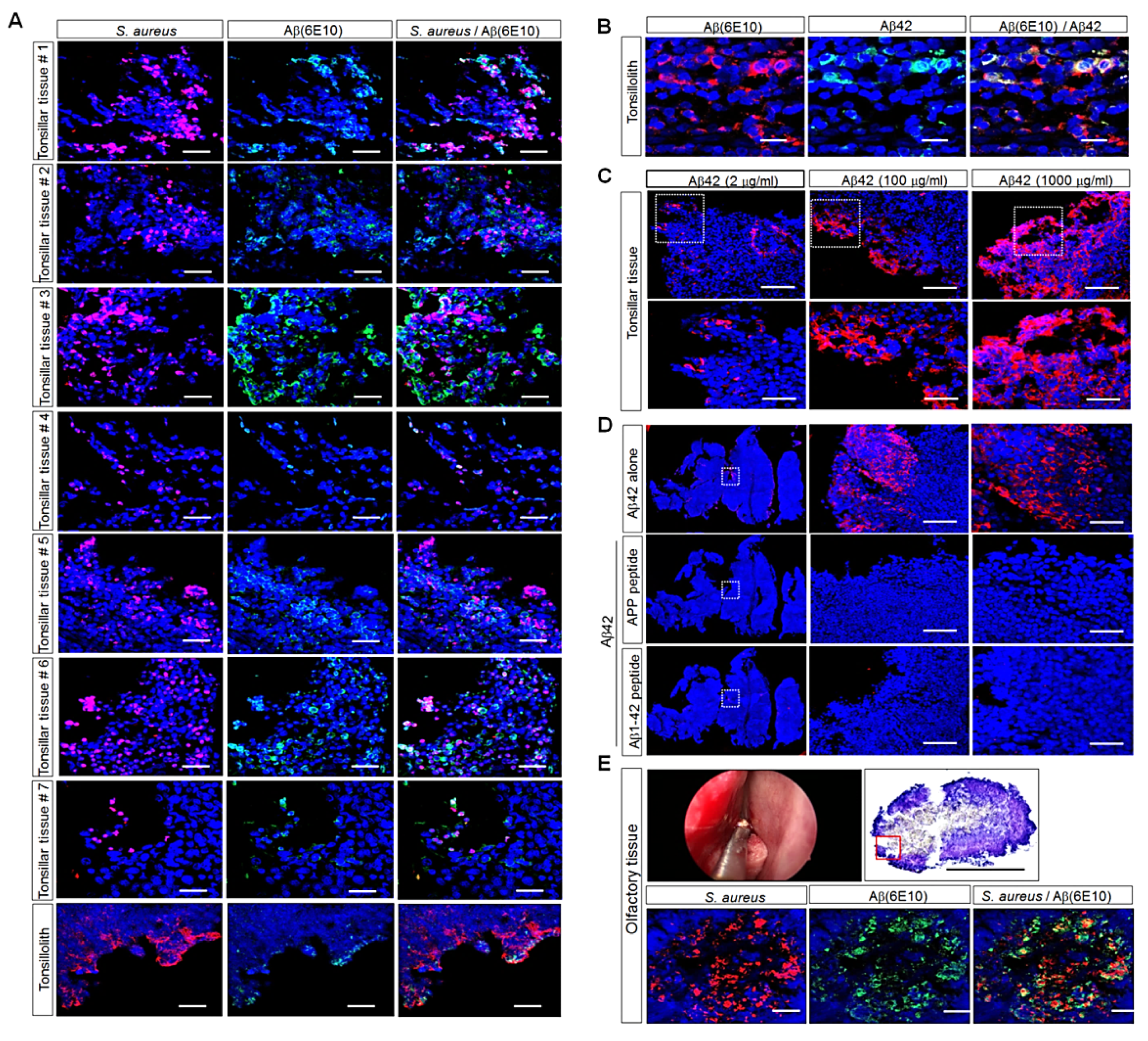
3.2. Detection of Aβ Deposition and S. aureus in Human Palatine-Tonsil Tissue
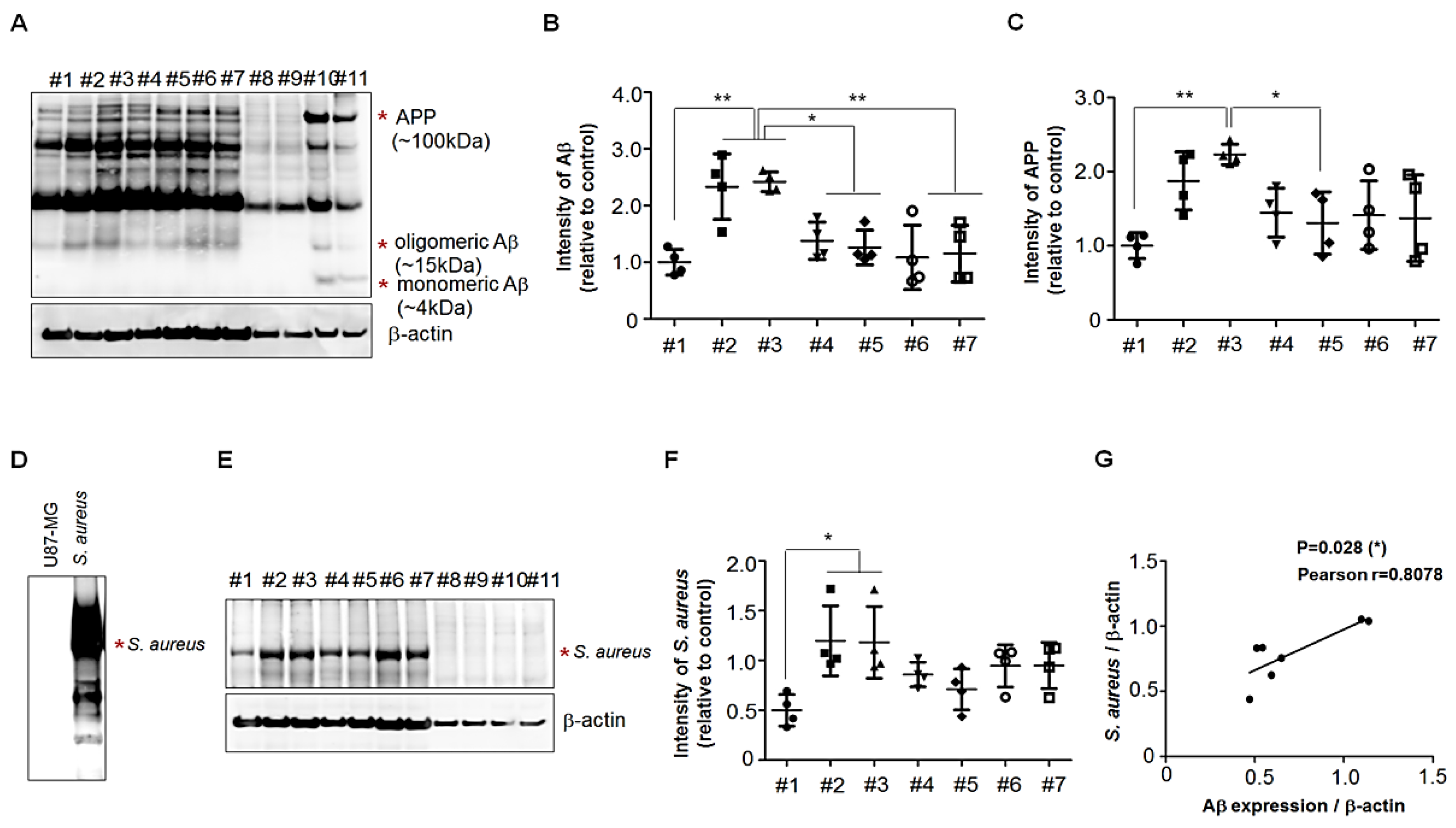
3.3. Presence of APP Fragments in Human Palatine-Tonsil Tissue
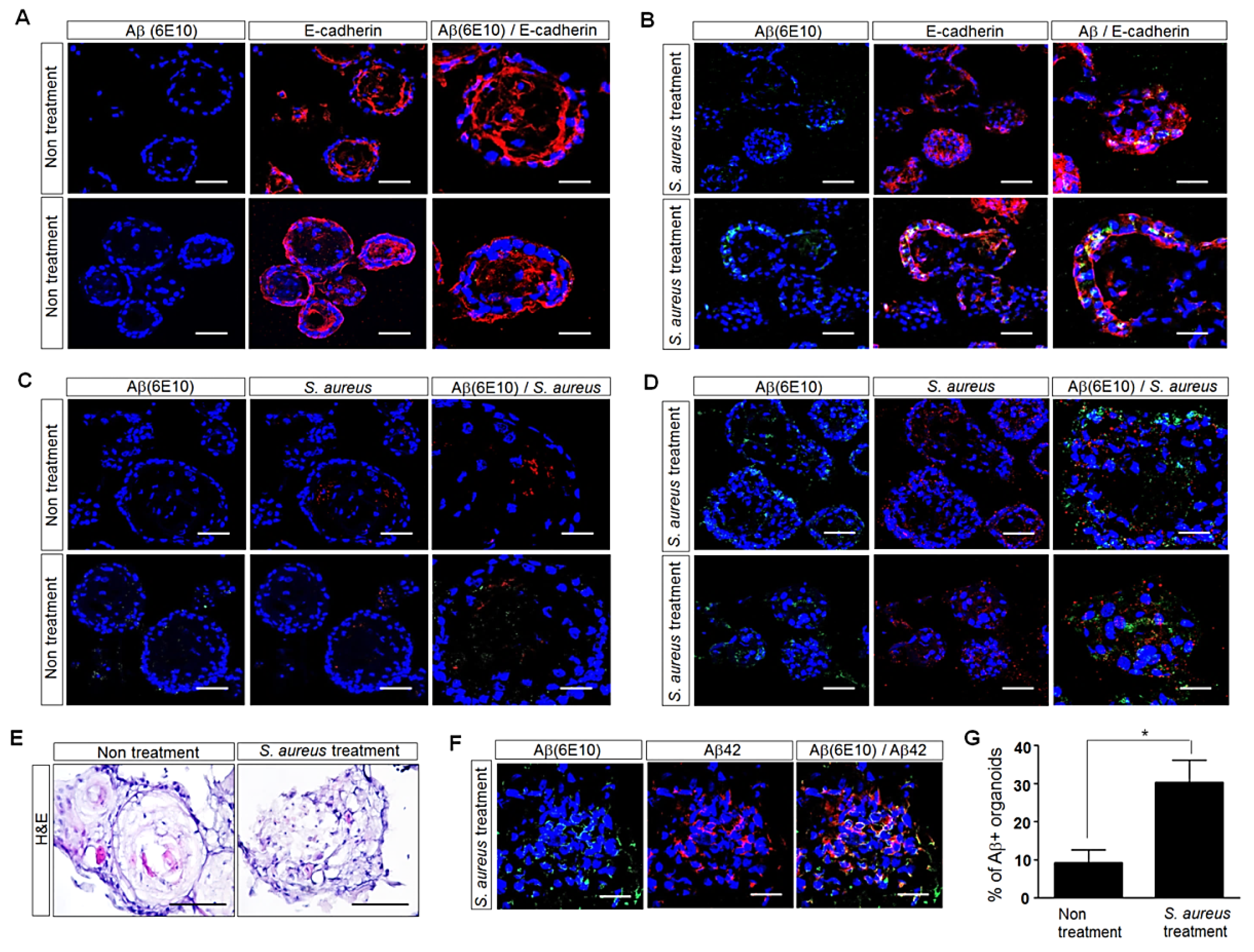
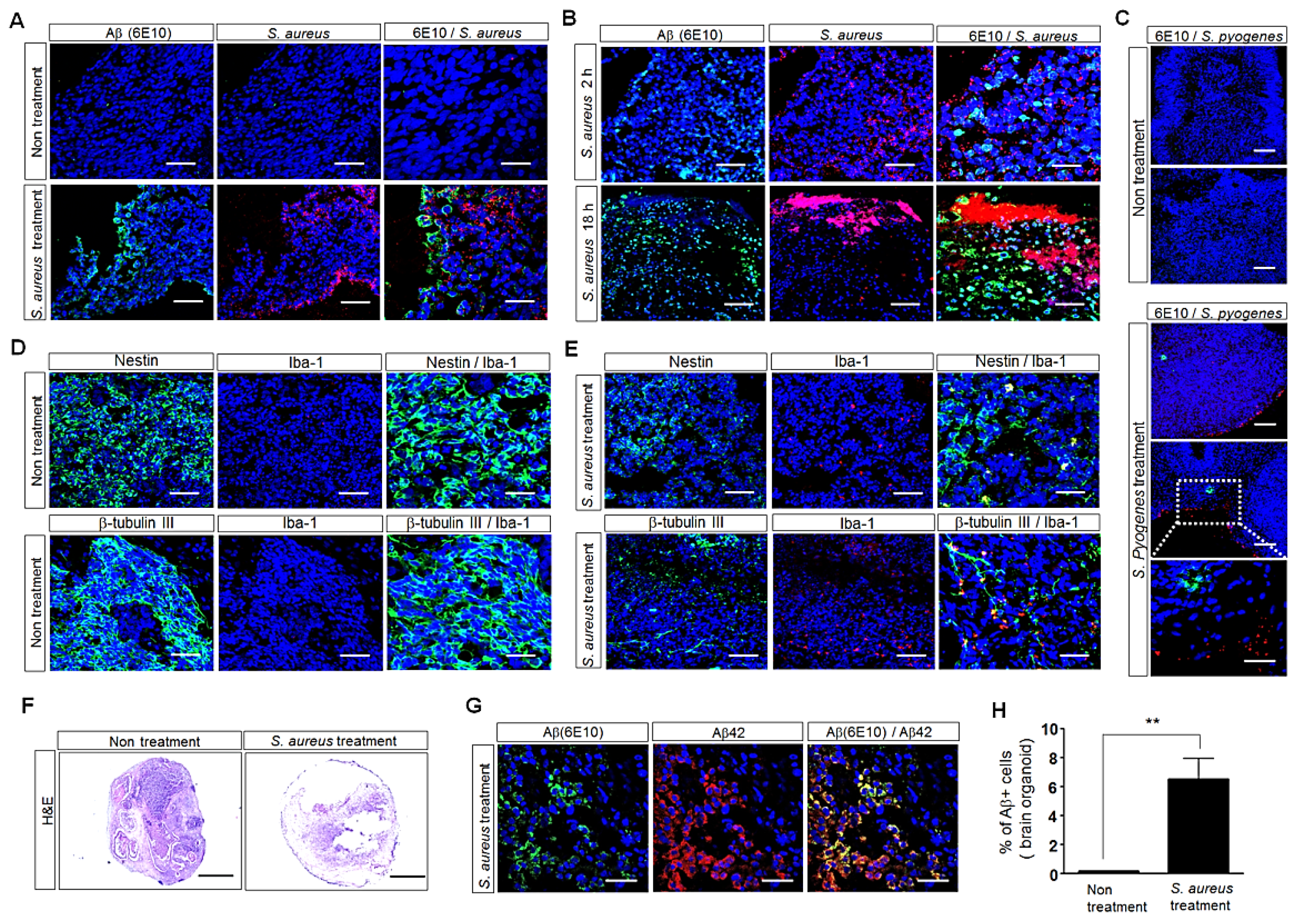
3.4. Influence of S. aureus on Aβ-Protein Expression in Human Palatine-Tonsil-Tissue-Derived Tonsil Organoids
3.5. Influence of S. aureus on Aβ-Protein Expression in Human iPSC (hiPSC)-Derived Human Brain Organoids
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Chow, V.W.; Mattson, M.P.; Wong, P.C.; Gleichmann, M. An Overview of APP Processing Enzymes and Products. NeuroMolecular Med. 2010, 12, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiltfang, J.; Esselmann, H.; Bibl, M.; Smirnov, A.; Otto, M.; Paul, S.; Kornhuber, J. Highly conserved and disease-specific patterns of carboxyterminally truncated Abeta peptides 1-37/38/39 in addition to 1-40/42 in Alzheimer’s disease and in patients with chronic neuroinflammation. J. Neurochem. 2002, 81, 481–496. [Google Scholar] [CrossRef] [PubMed]
- Sevalle, J.; Amoyel, A.; Robert, P.; Fournié-Zaluski, M.C.; Roques, B.; Checler, F. Aminopeptidase A contributes to the N-terminal truncation of amyloid beta-peptide. J. Neurochem. 2009, 109, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Araki, W.; Akiyama, H.; Tabira, T. Amino-truncated amyloid beta-peptide (Abeta5-40/42) produced from caspase-cleaved amyloid precursor protein is deposited in Alzheimer’s disease brain. FASEB J. 2004, 18, 1755–1757. [Google Scholar] [CrossRef]
- Bien, J.; Jefferson, T.; Causevic, M.; Jumpertz, T.; Munter, L.; Multhaup, G.; Weggen, S.; Becker-Pauly, C.; Pietrzik, C.U. The Metalloprotease Meprin β Generates Amino Terminal-truncated Amyloid β Peptide Species. J. Biol. Chem. 2012, 287, 33304–33313. [Google Scholar] [CrossRef] [Green Version]
- Haass, C. Take five--BACE and the gamma-secretase quartet conduct Alzheimer’s amyloid beta-peptide generation. EMBO J. 2004, 23, 483–488. [Google Scholar] [CrossRef]
- Balin, B.J.; Gérard, H.C.; Arking, E.J.; Appelt, D.M.; Branigan, P.J.; Abrams, J.T.; Whittum-Hudson, J.A.; Hudson, A.P. Identification and localization of Chlamydia pneumoniae in the Alzheimer’s brain. Med. Microbiol. Immunol. 1998, 187, 23–42. [Google Scholar] [CrossRef]
- Balin, B.; Hammond, C.; Little, C.S.; Appelt, D.; Hingley, S. Evidence for an infectious etiology in Alzheimer’s disease. In Advanced Understanding of Neurodegenerative Diseases; Chang, R.C.C., Ed.; InTech: Rijeka, Croatia, 2011; pp. 21–52. [Google Scholar]
- Bu, X.L.; Yao, X.Q.; Jiao, S.S.; Zeng, F.; Liu, Y.H.; Xiang, Y.; Wang, Y.J. A study on the association between infectious burden and Alzheimer’s disease. Eur. J. Neurol. 2015, 22, 1519–1525. [Google Scholar] [CrossRef]
- Gérard, H.C.; Dreses-Werringloer, U.; Wildt, K.S.; Deka, S.; Oszust, C.; Balin, B.J.; Hudson, A.P. Chlamydophila (Chlamydia) pneumoniae in the Alzheimer’s brain. FEMS Immunol. Med. Microbiol. 2006, 48, 355–366. [Google Scholar] [CrossRef] [Green Version]
- Little, C.S.; Joyce, T.A.; Hammond, C.J.; Matta, H.; Cahn, D.; Appelt, D.M.; Balin, B.J. Detection of bacterial antigens and Alzheimer’s disease-like pathology in the central nervous system of BALB/c mice following intranasal infection with a laboratory isolate of Chlamydia pneumoniae. Front. Aging Neurosci. 2014, 6, 304. [Google Scholar] [CrossRef] [Green Version]
- Surewaard, B.G.; Deniset, J.F.; Zemp, F.J.; Amrein, M.; Otto, M.; Conly, J.; Kubes, P. Identification and treatment of the Staphylococcus aureus reservoir in vivo. J. Exp. Med. 2016, 213, 1141–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decker, C.F. Pathogenesis of MRSA Infections. Disease-a-Month 2008, 54, 774–779. [Google Scholar] [CrossRef]
- Gordon, R.J.; Lowy, F.D. Pathogenesis of methicillin-resistant Staphylococcus aureus infection. Clin. Infect. Dis. 2008, 46 (Suppl. S5), S350–S359. [Google Scholar] [CrossRef] [Green Version]
- Lowy, F.D. Staphylococcus aureus infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef]
- Diep, B.A.; Otto, M. The role of virulence determinants in community-associated MRSA pathogenesis. Trends Microbiol. 2008, 16, 361–369. [Google Scholar] [CrossRef] [Green Version]
- Jacobsson, G.; Gustafsson, E.; Andersson, R. Outcome for invasive Staphylococcus aureus infections. Eur. J. Clin. Microbiol. Infect. Dis. 2008, 27, 839–848. [Google Scholar] [CrossRef] [PubMed]
- Naber, C.K. Staphylococcus aureus bacteremia: Epidemiology, pathophysiology, and management strategies. Clin. Infect. Dis. 2009, 48 (Suppl. S4), S231–S237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bloch, O.; Papadopoulos, M.C.; Manley, G.T.; Verkman, A.S. Aquaporin-4 gene deletion in mice increases focal edema associated with staphylococcal brain abscess. J. Neurochem. 2005, 95, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, M.; Benfield, T.L.; Skinhoej, P.; Jensen, A.G. Haematogenous Staphylococcus aureus meningitis. A 10-year nationwide study of 96 consecutive cases. BMC Infect. Dis. 2006, 6, 49. [Google Scholar] [CrossRef] [Green Version]
- Vartzelis, G.; Theodoridou, M.; Daikos, G.L.; Dellagrammaticas, H.; Syriopoulou, V.P. Brain abscesses complicating Staphylococcus aureus sepsis in a premature infant. Infection 2005, 33, 36–38. [Google Scholar] [CrossRef]
- Spitzer, P.; Condic, M.; Herrmann, M.; Oberstein, T.J.; Scharin-Mehlmann, M.; Gilbert, D.F.; Maler, J.M. Amyloidogenic amyloid-β-peptide variants induce microbial agglutination and exert antimicrobial activity. Sci. Rep. 2016, 6, 32228. [Google Scholar] [CrossRef] [PubMed]
- Zautner, A.E.; Krause, M.; Stropahl, G.; Holtfreter, S.; Frickmann, H.; Maletzki, C.; Podbielski, A. Intracellular persisting Staphylococcus aureus is the major pathogen in recurrent tonsillitis. PLoS ONE 2010, 5, e9452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nave, H.; Gebert, A.; Pabst, R. Morphology and immunology of the human palatine tonsil. Anat. Embryol. 2001, 204, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Kurnatowski, P.; Putyński, L.; Łapienis, M.; Kowalska, B. Physical and emotional disturbances in children with adenotonsillar hypertrophy. J. Laryngol. Otol. 2007, 122, 931–935. [Google Scholar] [CrossRef] [PubMed]
- Erdag, T.K.; Ecevit, M.C.; Guneri, E.A.; Dogan, E.; Ikiz, A.O.; Sutay, S. Pathologic evaluation of routine tonsillectomy and adenoidectomy specimens in the pediatric population: Is it really necessary? Int. J. Pediatr. Otorhinolaryngol. 2005, 69, 1321–1325. [Google Scholar] [CrossRef]
- Kim, H.K.; Kim, H.; Lee, M.K.; Choi, W.H.; Jang, Y.; Shin, J.S.; Yoo, J. Generation of tonsil organoids as an ex vivo model for SARS-CoV-2 infection. BioRxiv. 2020. [Google Scholar] [CrossRef]
- Kim, Y.; Park, N.; Rim, Y.A.; Nam, Y.; Jung, H.; Lee, K.; Ju, J.H. Establishment of a complex skin structure via layered co-culture of keratinocytes and fibroblasts derived from induced pluripotent stem cells. Stem Cell Res. Ther. 2018, 9, 217. [Google Scholar] [CrossRef] [Green Version]
- Österlund, A.; Popa, R.; Nikkilä, T.; Scheynius, A.; Engstrand, L. Intracellular Reservoir of Streptococcus pyogenes In Vivo: A Possible Explanation for Recurrent Pharyngotonsillitis. Laryngoscope 1997, 107, 640–647. [Google Scholar] [CrossRef]
- Grant, M.K.O.; Handoko, M.; Rozga, M.; Brinkmalm, G.; Portelius, E.; Blennow, K.; Ashe, K.H.; Zahs, K.R.; Liu, P. Human cerebrospinal fluid 6E10-immunoreactive protein species contain amyloid precursor protein fragments. PLoS ONE 2019, 14, e0212815. [Google Scholar]
- Hansson, O.; Zetterberg, H.; Vanmechelen, E.; Vanderstichele, H.; Andreasson, U.; Londos, E.; Blennow, K. Evaluation of plasma Aβ40 and Aβ42 as predictors of conversion to Alzheimer’s disease in patients with mild cognitive impairment. Neurobiol. Aging 2008, 31, 357–367. [Google Scholar] [CrossRef]
- Kuo, Y.M.; Kokjohn, T.A.; Watson, M.D.; Woods, A.S.; Cotter, R.J.; Sue, L.I.; Roher, A.E. Elevated Aβ42 in skeletal muscle of Alzheimer disease patients suggests peripheral alterations of AβPP metabolism. Am. J. Pathol. 2000, 156, 797–805. [Google Scholar] [CrossRef]
- Li, Q.X.; Whyte, S.; Tanner, J.E.; Evin, G.; Beyreuther, K.; Masters, C.L. Secretion of Alzheimer’s disease Aβ amyloid peptide by activated human platelets. Lab. Invest. 1998, 78, 461–469. [Google Scholar] [PubMed]
- Van Nostrand, W.E.; Melchor, J.P. Disruption of pathologic amyloid beta-protein fibril assembly on the surface of cultured human cerebrovascular smooth muscle cells. Amyloid 2001, 8, 20–27. [Google Scholar] [PubMed]
- Selkoe, D.J.; Podlisny, M.B.; Joachim, C.L.; Vickers, E.A.; Lee, G.; Fritz, L.C.; Oltersdorf, T. Beta-amyloid precursor protein of Alzheimer disease occurs as 110- to 135-kilodalton membrane-associated proteins in neural and nonneural tissues. Proc. Natl. Acad. Sci. USA 1988, 85, 7341–7345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catteruccia, N.; Willingale-Theune, J.; Bunke, D.; Prior, R.; Masters, C.L.; Crisanti, A.; Beyreuther, K. Ultrastructural localization of the putative precursors of the A4 amyloid protein associated with Alzheimer’s disease. Am. J. Pathol. 1990, 137, 19–26. [Google Scholar]
- Sandbrink, R.; Masters, C.L.; Beyreuther, K. Beta A4-amyloid protein precursor mRNA isoforms without exon 15 are ubiquitously expressed in rat tissues including brain, but not in neurons. J. Biol. Chem. 1994, 269, 1510–1517. [Google Scholar] [CrossRef]
- Bu, X.L.; Xiang, Y.; Jin, W.S.; Wang, J.; Shen, L.L.; Huang, Z.L.; Wang, Y.J. Blood-derived amyloid-β protein induces Alzheimer’s disease pathologies. Mol. Psychiatry 2018, 23, 1948–1956. [Google Scholar] [CrossRef]
- Zeng, F.; Liu, Y.; Huang, W.; Qing, H.; Kadowaki, T.; Kashiwazaki, H.; Wu, Z. Receptor for advanced glycation end products up-regulation in cerebral endothelial cells mediates cerebrovascular-related amyloid β accumulation after Porphyromonas gingivalis infection. J. Neurochem. 2021, 158, 724–736. [Google Scholar] [CrossRef]
- Wang, M.J.; Yi, S.; Han, J.-Y.; Park, S.Y.; Jang, J.-W.; Chun, I.K.; Kim, S.E.; Lee, B.S.; Kim, G.J.; Yu, J.S.; et al. Oligomeric forms of amyloid-β protein in plasma as a potential blood-based biomarker for Alzheimer’s disease. Alzheimer’s Res. Ther. 2017, 9, 98. [Google Scholar] [CrossRef] [Green Version]
- Mielke, M.M.; Hagen, C.E.; Xu, J.; Chai, X.; Vemuri, P.; Lowe, V.J.; Airey, D.C.; Knopman, D.S.; Roberts, R.O.; Machulda, M.M.; et al. Plasma phospho-tau181 increases with Alzheimer’s disease clinical severity and is associated with tau- and amyloid-positron emission tomography. Alzheimer’s Dement. 2018, 14, 989–997. [Google Scholar] [CrossRef]
- Olsson, B.; Lautner, R.; Andreasson, U.; Öhrfelt, A.; Portelius, E.; Bjerke, M.; Zetterberg, H. CSF and blood biomarkers for the diagnosis of Alzheimer’s disease: A systematic review and meta-analysis. Lancet Neurol. 2016, 15, 673–684. [Google Scholar] [CrossRef]
- Park, J.-C.; Han, S.-H.; Cho, H.J.; Byun, M.S.; Yi, D.; Choe, Y.M.; Kang, S.; Jung, E.S.; Won, S.J.; Kim, E.H.; et al. Chemically treated plasma Aβ is a potential blood-based biomarker for screening cerebral amyloid deposition. Alzheimer’s Res. Ther. 2017, 9, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, D.R.; Kern, D.W.; Wroblewski, K.E.; McClintock, M.K.; Dale, W.; Pinto, J.M. Olfactory dysfunction predicts subsequent dementia in older U.S. adults. J. Am. Geriatr. Soc. 2018, 66, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Rao, X.; Gao, Y.; Wang, J.; Xu, F. Amyloid-β Deposition and Olfactory Dysfunction in an Alzheimer’s Disease Model. J. Alzheimer’s Dis. 2013, 37, 699–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kameshima, N.; Yanagisawa, D.; Tooyama, I. β-Amyloid peptide (1–40) in the brain reaches the nasal cavity via a non-blood pathway. Neurosci. Res. 2013, 76, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Lee, S.-M.; Cho, S.; Kang, J.-H.; Minn, Y.-K.; Park, H.; Choi, S.H. Amyloid beta in nasal secretions may be a potential biomarker of Alzheimer’s disease. Sci. Rep. 2019, 9, 4966. [Google Scholar] [CrossRef] [PubMed]
- Gysin, C. Indications of pediatric tonsillectomy. ORL J. Otorhinolaryngol. Relat. Spec. 2013, 75, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Jeong, J.H.; Lee, D.W.; Ryu, R.A.; Lee, Y.S.; Lee, S.H.; Kang, J.O.; Tae, K. Bacteriologic comparison of tonsil core in recurrent tonsillitis and tonsillar hypertrophy. Laryngoscope 2007, 117, 2146–2151. [Google Scholar] [CrossRef]
- Skoulakis, C.; Tigiroglou, E.; Gkarelis, K.; Klapsa, D.; Damani, A.; Papadakis, C.; Petinaki, E. Level of Streptococcus pyogenes in patients with recurrent tonsillitis and tonsillar hypertrophy. Scand. J. Infect. Dis. 2008, 40, 899–903. [Google Scholar] [CrossRef]
- Van Staaij, B.K.; Van Den Akker, E.H.; De Haas Van Dorsser, E.H.; Fleer, A.; Hoes, A.W.; Schilder, A.G. Does the tonsillar surface flora differ in children with and without tonsillar disease? Acta Otolaryngol. 2003, 123, 873–878. [Google Scholar] [CrossRef]
- Stjernquist-Desatnik, A.; Holst, E. Tonsillar microbial flora: Comparison of recurrent tonsillitis and normal tonsils. Acta Otolaryngol. 1999, 119, 102–106. [Google Scholar] [PubMed]
- Kheirandish-Gozal, L.; Philby, M.F.; Alonso-Álvarez, M.L.; Terán-Santos, J.; Gozal, D. Biomarkers of Alzheimer disease in children with obstructive sleep apnea: Effect of adenotonsillectomy. Sleep 2016, 39, 1225–1232. [Google Scholar] [CrossRef] [Green Version]
- Sakono, M.; Zako, T. Amyloid oligomers: Formation and toxicity of Abeta oligomers. FEBS J. 2010, 277, 1348–1358. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.L.; Serpell, L.C. Membrane and surface interactions of Alzheimer’s Aβ peptide—Insights into the mechanism of cytotoxicity. FEBS J. 2011, 278, 3905–3917. [Google Scholar] [CrossRef] [PubMed]
- De Chiara, G.; Marcocci, M.E.; Sgarbanti, R.; Civitelli, L.; Ripoli, C.; Piacentini, R.; Garaci, E.; Grassi, C.; Palamara, A.T. Infectious Agents and Neurodegeneration. Mol. Neurobiol. 2012, 46, 614–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, S.A.; Harris, E.A. Herpes Simplex Virus Type 1 and Other Pathogens are Key Causative Factors in Sporadic Alzheimer’s Disease. J. Alzheimer’s Dis. 2015, 48, 319–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miklossy, J. Emerging roles of pathogens in Alzheimer disease. Expert Rev. Mol. Med. 2011, 13, e30. [Google Scholar] [CrossRef] [Green Version]
- Alonso, R.; Pisa, D.; Aguado, B.; Carrasco, L. Identification of Fungal Species in Brain Tissue from Alzheimer’s Disease by Next-Generation Sequencing. J. Alzheimer’s Dis. 2017, 58, 55–67. [Google Scholar] [CrossRef]
- Kumar, D.K.V.; Choi, S.H.; Washicosky, K.J.; Eimer, W.A.; Tucker, S.; Ghofrani, J.; Moir, R.D. Amyloid-β peptide protects against microbial infection in mouse and worm models of Alzheimer’s disease. Sci. Transl. Med. 2016, 8, 340–372. [Google Scholar] [CrossRef] [Green Version]
- Soscia, S.J.; Kirby, J.E.; Washicosky, K.J.; Tucker, S.M.; Ingelsson, M.; Hyman, B.; Burton, M.A.; Goldstein, L.E.; Duong, A.; Tanzi, R.E.; et al. The Alzheimer’s disease-associated amyloid beta-protein is an antimicrobial peptide. PLoS ONE 2010, 5, e9505. [Google Scholar] [CrossRef]
- Kühnel, T.S.; Reichert, T.E. Trauma of the midface. GMS Curr. Top. Otorhinolaryngol. Head Neck Surg 2015, 14, 1–45. [Google Scholar]
- Erturk, M.; Kayalioglu, G.; Govsa, F.; Varol, T.; Ozgur, T. The cranio-orbital foramen, the groove on the lateral wall of the human orbit, and the orbital branch of the middle meningeal artery. Clin. Anat. 2004, 18, 10–14. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, J.Y.; Lee, J.E.; Kim, H.K.; Park, Y.-J.; Jeon, J.H.; Park, S.-A.; Lee, N.; Lee, I.H.; Kim, D.H.; Yang, S.H.; et al. Human Palatine Tonsils Are Linked to Alzheimer’s Disease through Function of Reservoir of Amyloid Beta Protein Associated with Bacterial Infection. Cells 2022, 11, 2285. https://doi.org/10.3390/cells11152285
Lim JY, Lee JE, Kim HK, Park Y-J, Jeon JH, Park S-A, Lee N, Lee IH, Kim DH, Yang SH, et al. Human Palatine Tonsils Are Linked to Alzheimer’s Disease through Function of Reservoir of Amyloid Beta Protein Associated with Bacterial Infection. Cells. 2022; 11(15):2285. https://doi.org/10.3390/cells11152285
Chicago/Turabian StyleLim, Jung Yeon, Jung Eun Lee, Han Kyung Kim, Yeon-Joon Park, Jung Ho Jeon, Soon-A. Park, Naeun Lee, Il Hwan Lee, Do Hyun Kim, Seung Ho Yang, and et al. 2022. "Human Palatine Tonsils Are Linked to Alzheimer’s Disease through Function of Reservoir of Amyloid Beta Protein Associated with Bacterial Infection" Cells 11, no. 15: 2285. https://doi.org/10.3390/cells11152285