
Salmonella Promotes Its Own Survival in B Cells by Inhibiting Autophagy

 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Salmonella Strains
2.2. Mice
2.3. Primary B-Cell Culture
2.4. Infection of B Cells with Salmonella
2.5. Viability Assays on Salmonella and B Cells
2.6. Western Blot Analysis
2.7. Statistical Analysis
3. Results
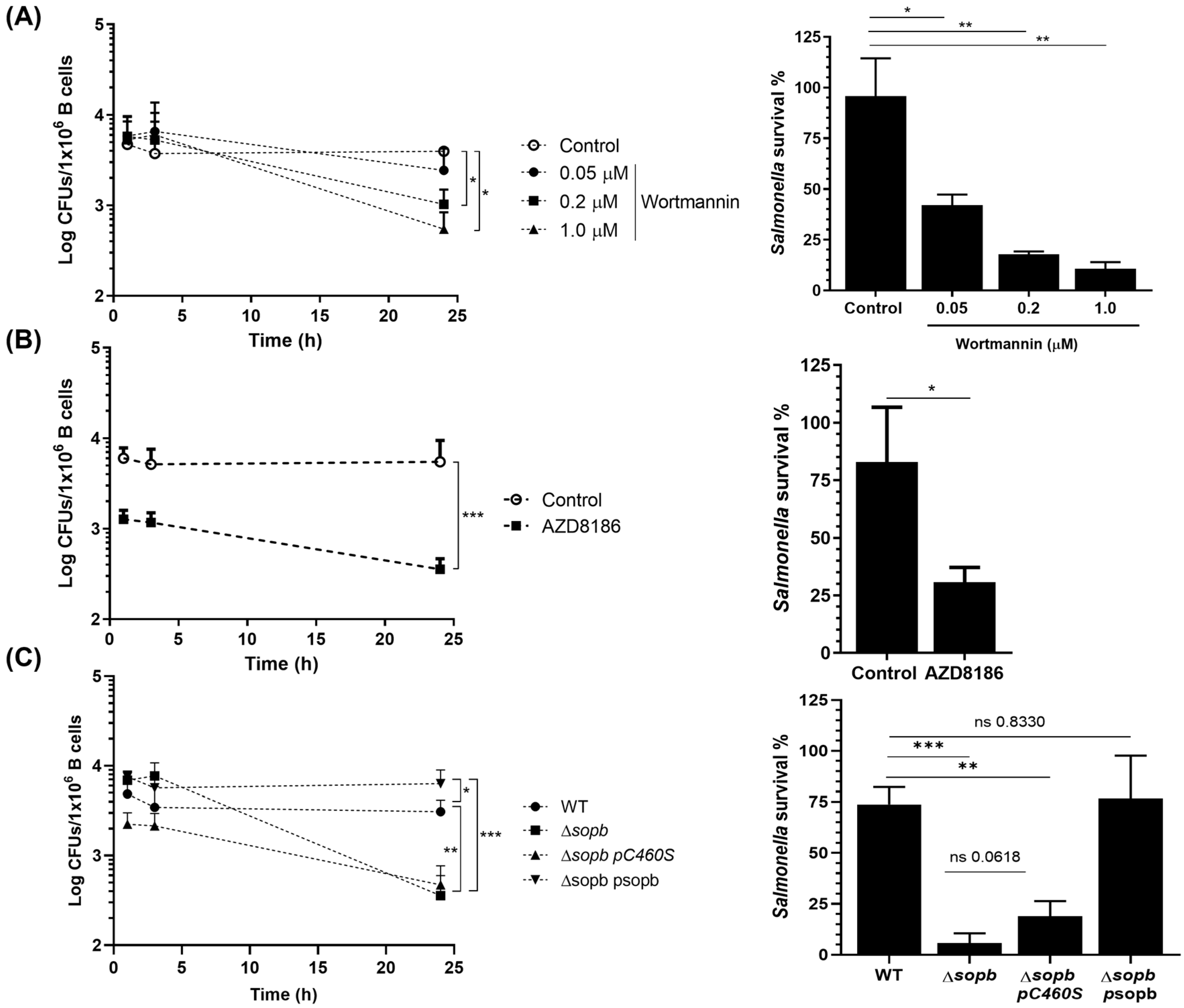
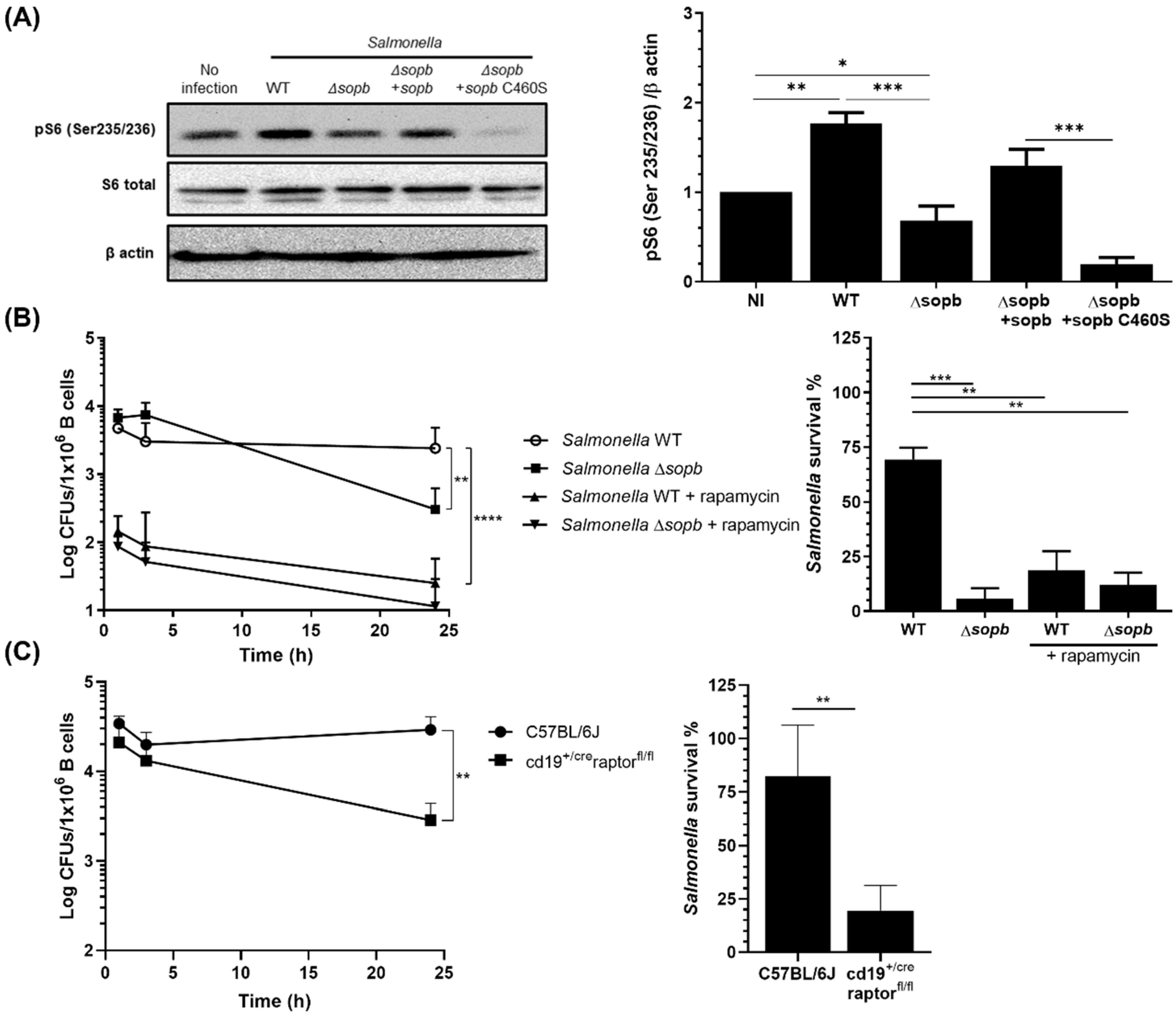
3.1. mTORC1 Activation by SopB Allows Salmonella Survival in B cells
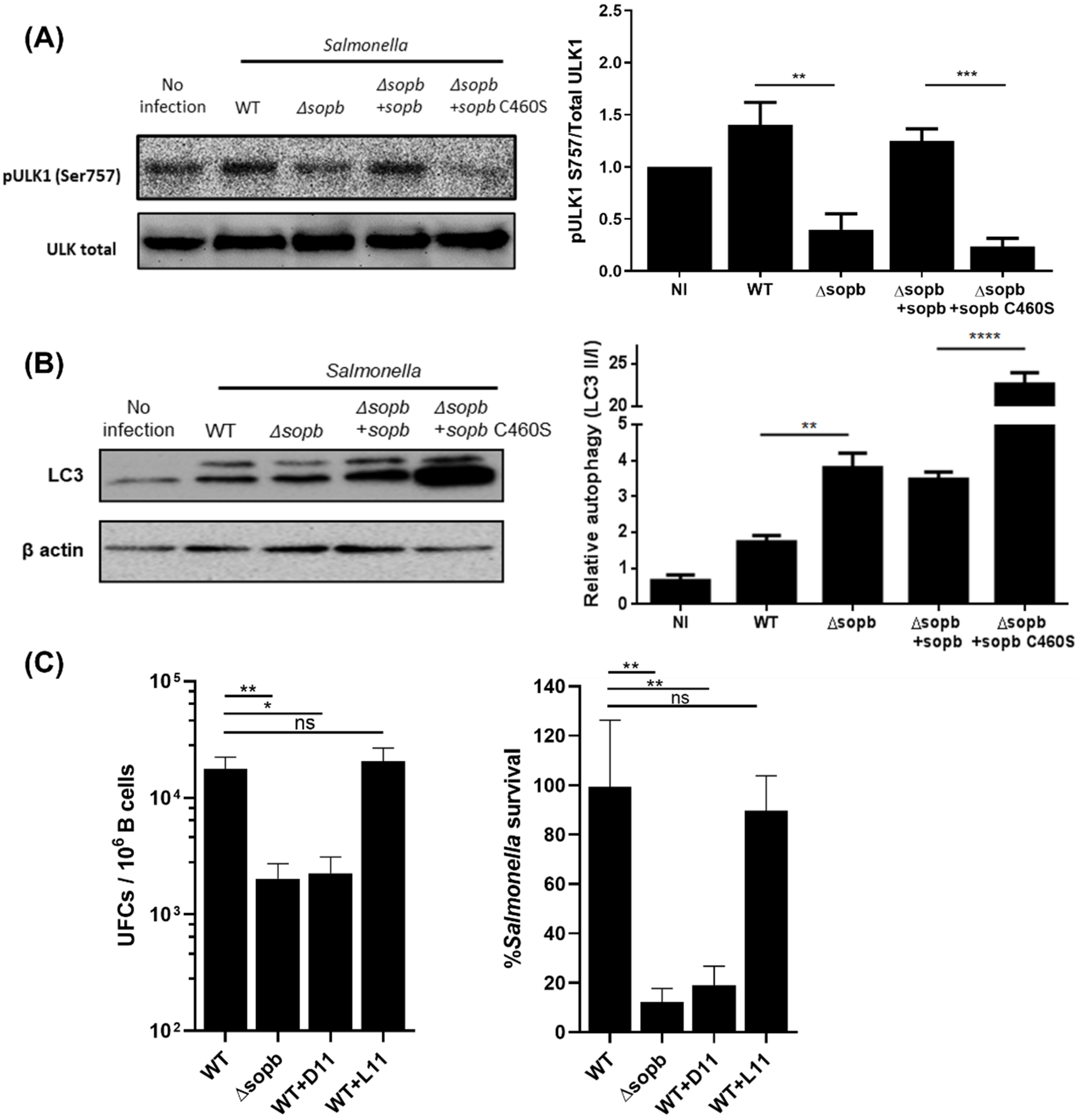
3.2. SopB-Mediated Activation of mTORC1 Inhibits Autophagy in B Cells
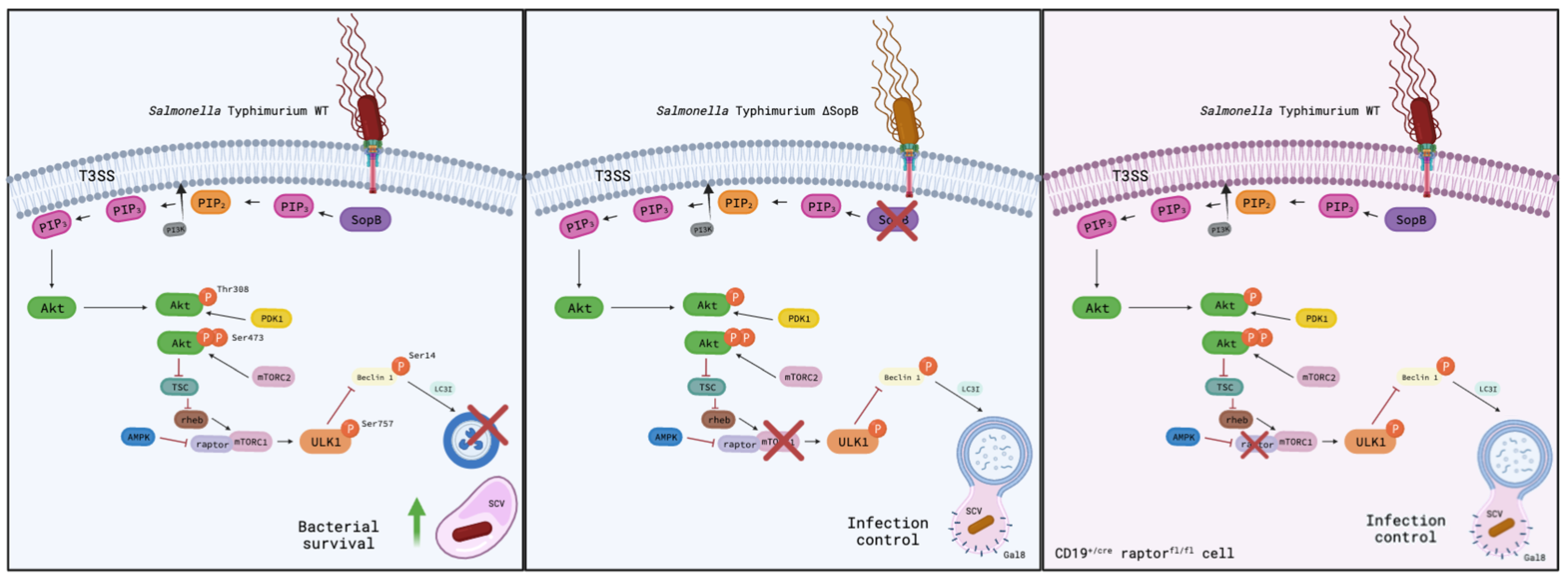
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Haraga, A.; Ohlson, M.B.; Miller, S.I. Salmonellae interplay with host cells. Nat. Rev. Microbiol. 2008, 6, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Drug Resistant Nontyphoidal Salmonella. 2019; p. 2. Available online: www.wou.edu/las/natsci_math/biology/boomer/Bio440/emerging2002/Salmonella2 (accessed on 8 December 2021).
- Levine, M.M.; Black, R.; Lanata, C. Precise Estimation of the Numbers of Chronic Carriers of Salmonella typhi in Santiago, Chile, an Endemic Area. J. Infect. Dis. 1982, 146, 724–726. [Google Scholar] [CrossRef] [PubMed]
- Hornick, R.B.; Greisman, S.E.; Woodward, T.E.; DuPont, H.L.; Dawkins, A.T.; Snyder, M.J. Typhoid Fever: Pathogenesis and Immunologic Control. N. Engl. J. Med. 1970, 283, 686–691. [Google Scholar] [CrossRef] [PubMed]
- Dongol, S.; Thompson, C.N.; Clare, S.; Nga, T.V.T.; Duy, P.T.; Karkey, A.; Arjyal, A.; Koirala, S.; Khatri, N.S.; Maskey, P.; et al. The Microbiological and Clinical Characteristics of Invasive Salmonella in Gallbladders from Cholecystectomy Patients in Kathmandu, Nepal. PLoS ONE 2012, 7, e47342. [Google Scholar] [CrossRef] [Green Version]
- Brigo, N.; Pfeifhofer-Obermair, C.; Tymoszuk, P.; Demetz, E.; Engl, S.; Barros-Pinkelnig, M.; Dichtl, S.; Fischer, C.; Valente De Souza, L.; Petzer, V.; et al. Cytokine-Mediated Regulation of ARG1 in Macrophages and Its Impact on the Control of Salmonella enterica Serovar Typhimurium Infection. Cells 2021, 10, 1823. [Google Scholar] [CrossRef]
- Rosales-Reyes, R.; Alpuche-Aranda, C.; Ramírez-Aguilar, M.D.L.L.; Castro-Eguiluz, A.D.; Ortiz-Navarrete, V. Survival of Salmonella enterica Serovar Typhimurium within Late Endosomal-Lysosomal Compartments of B Lymphocytes Is Associated with the Inability To Use the Vacuolar Alternative Major Histocompatibility Complex Class I Antigen-Processing Pathway. Infect. Immun. 2005, 73, 3937–3944. [Google Scholar] [CrossRef] [Green Version]
- Castro-Eguiluz, D.; Pelayo, R.; Rosales-Garcia, V.; Rosales-Reyes, R.; Alpuche-Aranda, C.; Ortiz-Navarrete, V. B cell precursors are targets for Salmonella infection. Microb. Pathog. 2009, 47, 52–56. [Google Scholar] [CrossRef]
- Alvarez, M.I.; Glover, L.C.; Luo, P.; Wang, L.; Theusch, E.; Oehlers, S.H.; Walton, E.M.; Tram, T.T.B.; Kuang, Y.-L.; Rotter, J.I.; et al. Human genetic variation in VAC14 regulates Salmonella invasion and typhoid fever through modulation of cholesterol. Proc. Natl. Acad. Sci. USA 2017, 114, E7746–E7755. [Google Scholar] [CrossRef] [Green Version]
- Patel, J.C.; Galán, J.E. Differential activation and function of Rho GTPases during Salmonella–host cell interactions. J. Cell Biol. 2006, 175, 453–463. [Google Scholar] [CrossRef] [Green Version]
- Burkinshaw, B.J.; Prehna, G.; Worrall, L.J.; Strynadka, N.C. Structure of Salmonella Effector Protein SopB N-terminal Domain in Complex with Host Rho GTPase Cdc42. J. Biol. Chem. 2012, 287, 13348–13355. [Google Scholar] [CrossRef] [Green Version]
- Knodler, L.; Finlay, B.B.; Steele-Mortimer, O. The Salmonella Effector Protein SopB Protects Epithelial Cells from Apoptosis by Sustained Activation of Akt. J. Biol. Chem. 2005, 280, 9058–9064. [Google Scholar] [CrossRef] [Green Version]
- Perez-Lopez, A.; Rosales-Reyes, R.; Alpuche-Aranda, C.M.; Ortiz-Navarrete, V. Salmonella Downregulates Nod-like Receptor Family CARD Domain Containing Protein 4 Expression To Promote Its Survival in B Cells by Preventing Inflammasome Activation and Cell Death. J. Immunol. 2013, 190, 1201–1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Gil, A.; Galán-Enríquez, C.S.; Pérez-López, A.; Nava, P.; Alpuche-Aranda, C.; Ortiz-Navarrete, V. SopB activates the Akt-YAP pathway to promote Salmonella survival within B cells. Virulence 2018, 9, 1390–1402. [Google Scholar] [CrossRef] [Green Version]
- Castrejón-Jiménez, N.S.; Leyva-Paredes, K.; Hernández-González, J.C.; Luna-Herrera, J.; García-Pérez, B.E. The role of autophagy in bacterial infections. Biosci. Trends 2015, 9, 149–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogawa, M.; Yoshimori, T.; Suzuki, T.; Sagara, H.; Mizushima, N.; Sasakawa, C. Escape of Intracellular Shigella from Autophagy. Science 2005, 307, 727–731. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kundu, M.; Viollet, B.; Guan, K.-L. AMPK and mTOR regulate autophagy through direct phosphorylation of Ulk1. Nat. Cell Biol. 2011, 13, 132–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starokadomskyy, P.; Dmytruk, K.V. A bird’s-eye view of autophagy. Autophagy 2013, 9, 1121–1126. [Google Scholar] [CrossRef] [Green Version]
- Meley, D.; Bauvy, C.; Houben-Weerts, J.H.; Dubbelhuis, P.F.; Helmond, M.T.; Codogno, P.; Meijer, A.J. AMP-activated Protein Kinase and the Regulation of Autophagic Proteolysis. J. Biol. Chem. 2006, 281, 34870–34879. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.P.; Brown, J.R.; Sag, D.; Zhang, L.; Suttles, J. Adenosine 5′-Monophosphate–Activated Protein Kinase Regulates IL-10–Mediated Anti-Inflammatory Signaling Pathways in Macrophages. J. Immunol. 2014, 194, 584–594. [Google Scholar] [CrossRef] [Green Version]
- Birmingham, C.L.; Canadien, V.; Kaniuk, N.A.; Steinberg, B.E.; Higgins, D.E.; Brumell, J.H. Listeriolysin O allows Listeria monocytogenes replication in macrophage vacuoles. Nature 2008, 451, 350–354. [Google Scholar] [CrossRef]
- Huang, D.; Bao, L. Mycobacterium tuberculosis EspB protein suppresses interferon-γ-induced autophagy in murine macrophages. J. Microbiol. Immunol. Infect. 2015, 49, 859–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kayath, C.A.; Hussey, S.; El Hajjami, N.; Nagra, K.; Philpott, D.; Allaoui, A. Escape of intracellular Shigella from autophagy requires binding to cholesterol through the type III effector, IcsB. Microbes Infect. 2010, 12, 956–966. [Google Scholar] [CrossRef] [PubMed]
- Miao, E.; Brittnacher, M.; Haraga, A.; Jeng, R.L.; Welch, M.D.; Miller, S.I. Salmonella effectors translocated across the vacuolar membrane interact with the actin cytoskeleton. Mol. Microbiol. 2003, 48, 401–415. [Google Scholar] [CrossRef] [PubMed]
- Knodler, L.A.; Winfree, S.; Drecktrah, D.; Ireland, R.; Steele-Mortimer, O. Ubiquitination of the bacterial inositol phosphatase, SopB, regulates its biological activity at the plasma membrane. Cell. Microbiol. 2009, 11, 1652–1670. [Google Scholar] [CrossRef] [Green Version]
- Finn, C.E.; Chong, A.; Cooper, K.G.; Starr, T.; Steele-Mortimer, O. A second wave of Salmonella T3SS1 activity prolongs the lifespan of infected epithelial cells. PLOS Pathog. 2017, 13, e1006354. [Google Scholar] [CrossRef]
- Rosales-Reyes, R.; Pérez-López, A.; Sánchez-Gómez, C.; Hernández-Mote, R.R.; Castro-Eguiluz, D.; Ortiz-Navarrete, V.; Alpuche-Aranda, C.M. Salmonella infects B cells by macropinocytosis and formation of spacious phagosomes but does not induce pyroptosis in favor of its survival. Microb. Pathog. 2012, 52, 367–374. [Google Scholar] [CrossRef]
- Zhou, L.; Li, Y.; Gao, S.; Yuan, H.; Zuo, L.; Wu, C.; Huang, R.; Wu, S. Salmonella spvC Gene Inhibits Autophagy of Host Cells and Suppresses NLRP3 as Well as NLRC4. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef]
- Owen, K.A.; Meyer, C.B.; Bouton, A.H.; Casanova, J.E. Activation of Focal Adhesion Kinase by Salmonella Suppresses Autophagy via an Akt/mTOR Signaling Pathway and Promotes Bacterial Survival in Macrophages. PLOS Pathog. 2014, 10, e1004159. [Google Scholar] [CrossRef] [Green Version]
- Scheidel, J.; Amstein, L.; Ackermann, J.; Dikic, I.; Koch, I. In Silico Knockout Studies of Xenophagic Capturing of Salmonella. PLOS Comput. Biol. 2016, 12, e1005200. [Google Scholar] [CrossRef] [Green Version]
- Kreibich, S.; Emmenlauer, M.; Fredlund, J.; Rämö, P.; Münz, C.; Dehio, C.; Enninga, J.; Hardt, W.-D. Autophagy Proteins Promote Repair of Endosomal Membranes Damaged by the Salmonella Type Three Secretion System 1. Cell Host Microbe 2015, 18, 527–537. [Google Scholar] [CrossRef] [Green Version]
- Cemma, M.; Kim, P.K.; Brumell, J.H. The ubiquitin-binding adaptor proteins p62/SQSTM1 and NDP52 are recruited independently to bacteria-associated microdomains to target Salmonella to the autophagy pathway. Autophagy 2011, 7, 341–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauthe, M.; Orhon, I.; Rocchi, C.; Zhou, X.; Luhr, M.; Hijlkema, K.-J.; Coppes, R.P.; Engedal, N.; Mari, M.; Reggiori, F. Chloroquine inhibits autophagic flux by decreasing autophagosome-lysosome fusion. Autophagy 2018, 14, 1435–1455. [Google Scholar] [CrossRef] [PubMed]
- Fedele, A.O.; Proud, C.G. Chloroquine and bafilomycin A mimic lysosomal storage disorders and impair mTORC1 signalling. Biosci. Rep. 2020, 40, BSR20200905. [Google Scholar] [CrossRef] [Green Version]
- Tattoli, I.; Sorbara, M.T.; Vuckovic, D.; Ling, A.; Soares, F.; Carneiro, L.; Yang, C.; Emili, A.; Philpott, D.J.; Girardin, S.E. Amino Acid Starvation Induced by Invasive Bacterial Pathogens Triggers an Innate Host Defense Program. Cell Host Microbe 2012, 11, 563–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luis, L.-B.; Ana, G.-T.; Carlos, G.-E.; Abraham, G.-G.; Iris, E.-G.; Martha, M.-L.; Vianney, O.-N. Salmonella Promotes Its Own Survival in B Cells by Inhibiting Autophagy. Cells 2022, 11, 2061. https://doi.org/10.3390/cells11132061
Luis L-B, Ana G-T, Carlos G-E, Abraham G-G, Iris E-G, Martha M-L, Vianney O-N. Salmonella Promotes Its Own Survival in B Cells by Inhibiting Autophagy. Cells. 2022; 11(13):2061. https://doi.org/10.3390/cells11132061
Chicago/Turabian StyleLuis, Lopez-Bailon, Gonzalez-Telona Ana, Galán-Enríquez Carlos, García-Gil Abraham, Estrada-García Iris, Moreno-Lafont Martha, and Ortiz-Navarrete Vianney. 2022. "Salmonella Promotes Its Own Survival in B Cells by Inhibiting Autophagy" Cells 11, no. 13: 2061. https://doi.org/10.3390/cells11132061