
The Mechanisms Underlying the Beneficial Impact of Aerobic Training on Cancer-Related Fatigue: A Conceptual Review
 , , and
, , and 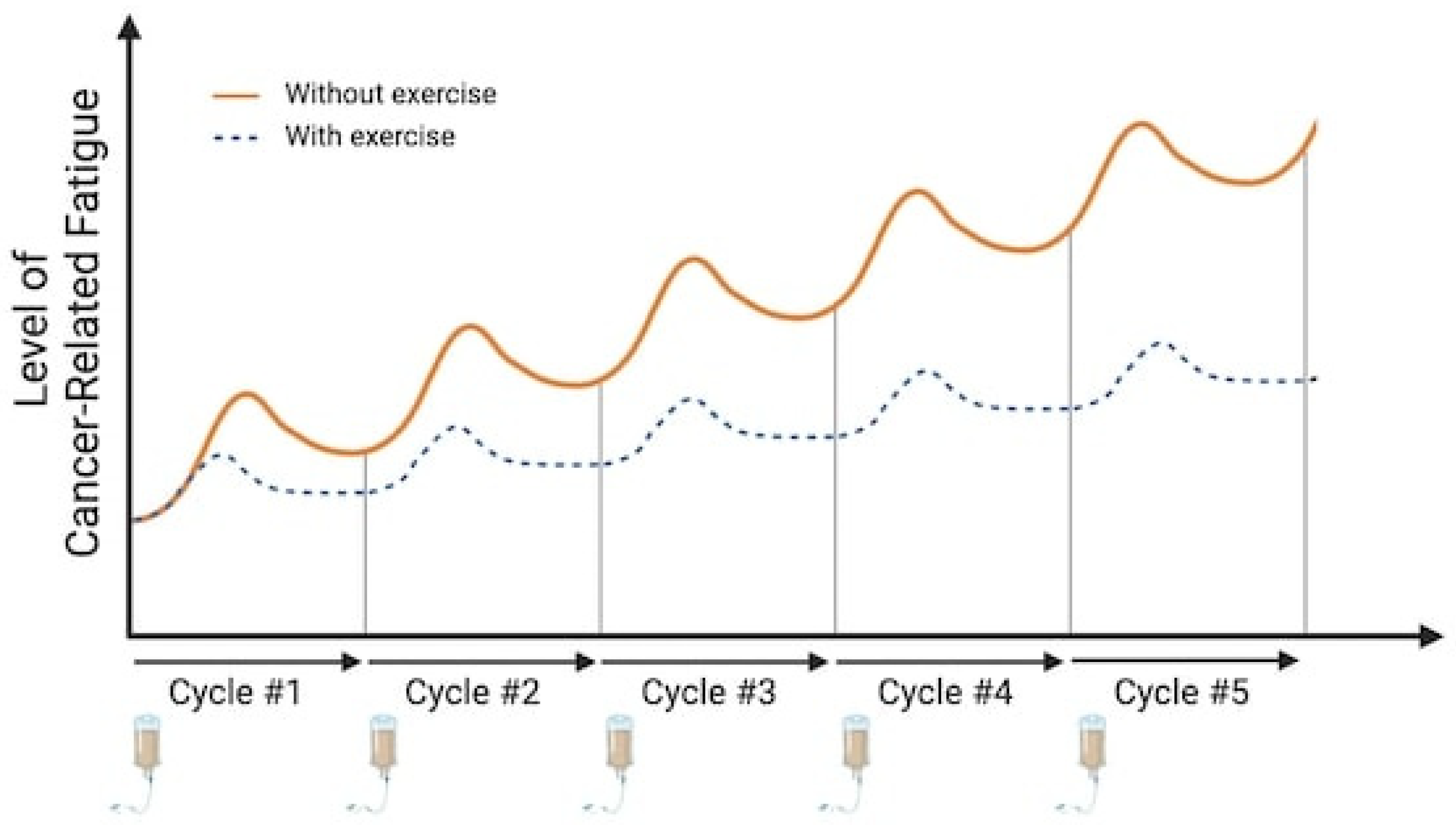{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. The Effects of Peripheral and Neural Inflammation on Cancer-Related Fatigue
2.1. Effect of Chemotherapeutic Treatments
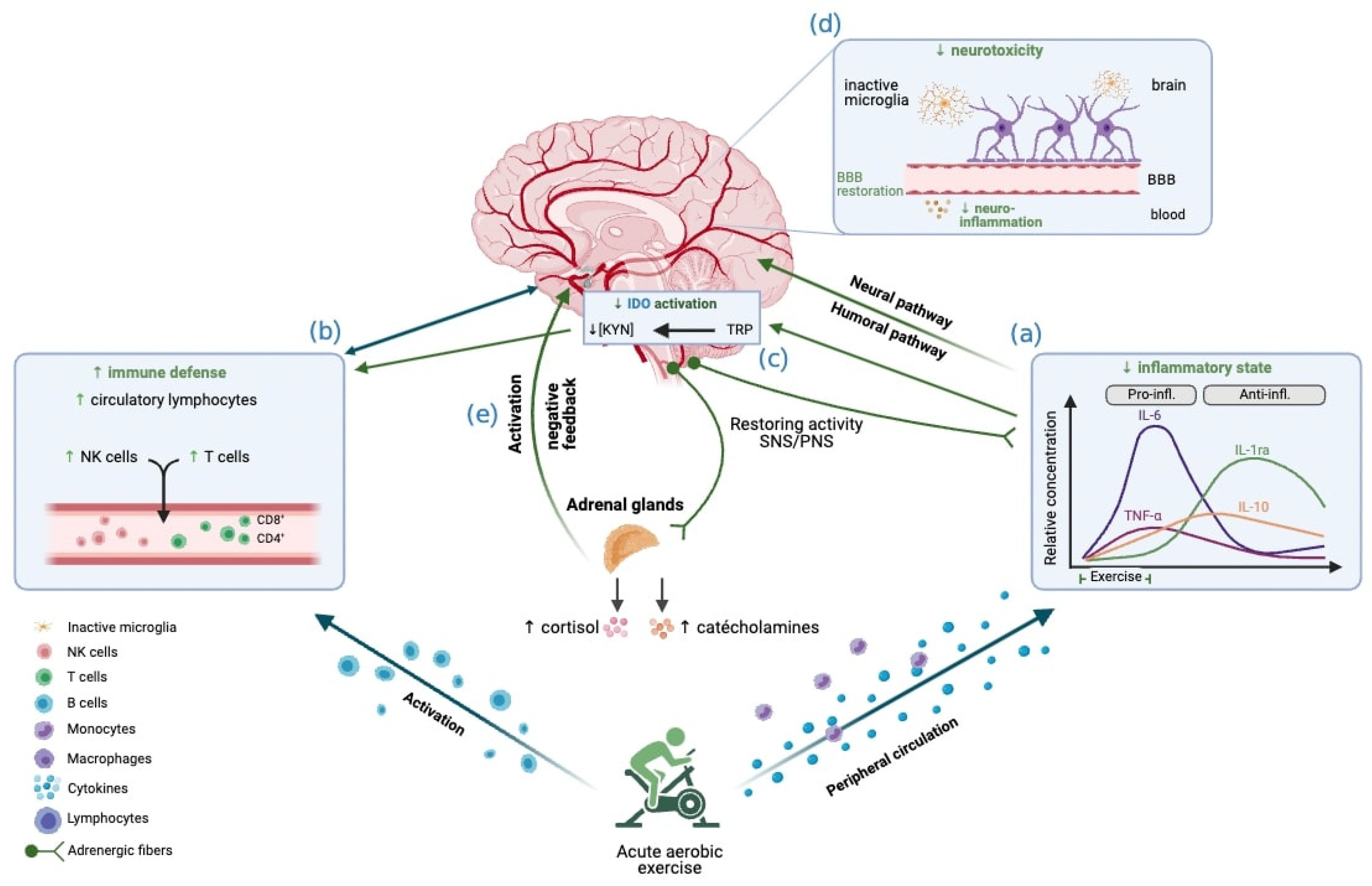
2.2. The Acute Effect of Aerobic Exercise on Peripheral and Neural Inflammation
3. Immune Function Dysregulation and Cancer-Related Fatigue
3.1. Immune Response after Chemotherapy
3.2. Acute Effect of Aerobic Exercise on Immune Markers
4. Neuroendocrine Alteration and Cancer-Related Fatigue
4.1. The Influence of Chemotherapeutic Agents on the Neuroendocrine System
4.2. The Acute Effect of Aerobic Exercise on the Neuroendocrine System
5. Physiological Variability in Exercise Response
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gerber, L.H. Cancer-Related Fatigue: Persistent, Pervasive, and Problematic. Phys. Med. Rehabil. Clin. N. Am. 2017, 28, 65–88. [Google Scholar] [CrossRef]
- Wang, X.S.; Woodruff, J.F. Cancer-Related and Treatment-Related Fatigue. Gynecol. Oncol. 2015, 136, 446–452. [Google Scholar] [CrossRef]
- Schmidt, M.E.; Hermann, S.; Arndt, V.; Steindorf, K. Prevalence and Severity of Long-Term Physical, Emotional, and Cognitive Fatigue across 15 Different Cancer Entities. Cancer Med. 2020, 9, 8053–8061. [Google Scholar] [CrossRef]
- Ma, Y.; He, B.; Jiang, M.; Yang, Y.; Wang, C.; Huang, C.; Han, L. Prevalence and Risk Factors of Cancer-Related Fatigue: A Systematic Review and Meta-Analysis. Int. J. Nurs. Stud. 2020, 111, 103707. [Google Scholar] [CrossRef]
- Jacobsen, P.B. Assessment of Fatigue in Cancer Patients. J. Natl. Cancer Inst. Monogr. 2004, 2004, 93–97. [Google Scholar] [CrossRef]
- Mock, V.; Atkinson, A.; Barsevick, A.; Cella, D.; Cimprich, B.; Cleeland, C.; Donnelly, J.; Eisenberger, M.A.; Escalante, C.; Hinds, P.; et al. NCCN Practice Guidelines for Cancer-Related Fatigue. Oncol. Williston Park N. Y. 2000, 14, 151–161. [Google Scholar]
- Husson, O.; Mols, F.; van de Poll-Franse, L.; de Vries, J.; Schep, G.; Thong, M.S.Y. Variation in Fatigue among 6011 (Long-Term) Cancer Survivors and a Normative Population: A Study from the Population-Based PROFILES Registry. Support. Care Cancer Off. J. Multinatl. Assoc. Support. Care Cancer 2015, 23, 2165–2174. [Google Scholar] [CrossRef]
- Reinertsen, K.V.; Cvancarova, M.; Loge, J.H.; Edvardsen, H.; Wist, E.; Fosså, S.D. Predictors and Course of Chronic Fatigue in Long-Term Breast Cancer Survivors. J. Cancer Surviv. Res. Pract. 2010, 4, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Arndt, V.; Koch-Gallenkamp, L.; Jansen, L.; Bertram, H.; Eberle, A.; Holleczek, B.; Schmid-Höpfner, S.; Waldmann, A.; Zeissig, S.R.; Brenner, H. Quality of Life in Long-Term and Very Long-Term Cancer Survivors versus Population Controls in Germany. Acta Oncol. Stockh. Swed. 2017, 56, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Ryan, J.L.; Carroll, J.K.; Ryan, E.P.; Mustian, K.M.; Fiscella, K.; Morrow, G.R. Mechanisms of Cancer-Related Fatigue. Oncologist 2007, 12 (Suppl. S1), 22–34. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Chu, S.; Gao, Y.; Ai, Q.; Liu, Y.; Li, X.; Chen, N. A Narrative Review of Cancer-Related Fatigue (CRF) and Its Possible Pathogenesis. Cells 2019, 8, 738. [Google Scholar] [CrossRef] [PubMed]
- LaVoy, E.C.P.; Fagundes, C.P.; Dantzer, R. Exercise, Inflammation, and Fatigue in Cancer Survivors. Exerc. Immunol. Rev. 2016, 22, 82–93. [Google Scholar] [PubMed]
- Barsevick, A.; Frost, M.; Zwinderman, A.; Hall, P.; Halyard, M.; GENEQOL Consortium. I’m so Tired: Biological and Genetic Mechanisms of Cancer-Related Fatigue. Qual. Life Res. Int. J. Qual. Life Asp. Treat. Care Rehabil. 2010, 19, 1419–1427. [Google Scholar] [CrossRef] [PubMed]
- Bower, J.E. Cancer-Related Fatigue—Mechanisms, Risk Factors, and Treatments. Nat. Rev. Clin. Oncol. 2014, 11, 597–609. [Google Scholar] [CrossRef] [PubMed]
- van Vulpen, J.K.; Sweegers, M.G.; Peeters, P.H.M.; Courneya, K.S.; Newton, R.U.; Aaronson, N.K.; Jacobsen, P.B.; Galvão, D.A.; Chinapaw, M.J.; Steindorf, K.; et al. Moderators of Exercise Effects on Cancer-Related Fatigue: A Meta-Analysis of Individual Patient Data. Med. Sci. Sports Exerc. 2020, 52, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Cramp, F.; Byron-Daniel, J. Exercise for the Management of Cancer-Related Fatigue in Adults. Cochrane Database Syst. Rev. 2012, 11, CD006145. [Google Scholar] [CrossRef]
- Fontvieille, A.; Parent-Roberge, H.; Maréchal, R.; Fülöp, T.; Riesco, E.; Pavic, M. Corrigendum to “Effects of a Mixed Exercise Program on Cancer Related-Fatigue and Health-Related Quality of Life in Oncogeriatric Patients: A Feasibility Study” [Journal of Geriatric Oncology 12 Issue 6 (2021) 915-921]. J. Geriatr. Oncol. 2022, 13, 1070. [Google Scholar] [CrossRef] [PubMed]
- Lipsett, A.; Barrett, S.; Haruna, F.; Mustian, K.; O’Donovan, A. The Impact of Exercise during Adjuvant Radiotherapy for Breast Cancer on Fatigue and Quality of Life: A Systematic Review and Meta-Analysis. Breast Edinb. Scotl. 2017, 32, 144–155. [Google Scholar] [CrossRef] [PubMed]
- Campbell, K.L.; Winters-Stone, K.M.; Wiskemann, J.; May, A.M.; Schwartz, A.L.; Courneya, K.S.; Zucker, D.S.; Matthews, C.E.; Ligibel, J.A.; Gerber, L.H.; et al. Exercise Guidelines for Cancer Survivors: Consensus Statement from International Multidisciplinary Roundtable. Med. Sci. Sports Exerc. 2019, 51, 2375–2390. [Google Scholar] [CrossRef]
- Mustian, K.M.; Cole, C.L.; Lin, P.J.; Asare, M.; Fung, C.; Janelsins, M.C.; Kamen, C.S.; Peppone, L.J.; Magnuson, A. Exercise Recommendations for the Management of Symptoms Clusters Resulting from Cancer and Cancer Treatments. In Seminars in Oncology Nursing; Elsevier: Amsterdam, The Netherlands, 2016; Volume 32, pp. 383–393. [Google Scholar]
- Farrell, C.; Turgeon, D.R. Normal Versus Chronic Adaptations to Aerobic Exercise. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Rusin, A.; Seymour, C.; Cocchetto, A.; Mothersill, C. Commonalities in the Features of Cancer and Chronic Fatigue Syndrome (CFS): Evidence for Stress-Induced Phenotype Instability? Int. J. Mol. Sci. 2022, 23, 691. [Google Scholar] [CrossRef]
- Bower, J.E.; Lamkin, D.M. Inflammation and Cancer-Related Fatigue: Mechanisms, Contributing Factors, and Treatment Implications. Brain. Behav. Immun. 2013, 30, S48–S57. [Google Scholar] [CrossRef] [PubMed]
- Regan, J.; Feng, R.; Saligan, L. 4313 Impaired Natural Killer Cell Function May Be Associated with Cancer-Related Fatigue. J. Clin. Transl. Sci. 2020, 4, 8. [Google Scholar] [CrossRef]
- Francesco, F.; Alice, N.; Filippo, G.; Daniela, R.; Enrico, B.; Emanuela, A. Stress Axis in the Cancer Patient: Clinical Aspects and Management. Endocrines 2021, 2, 502–513. [Google Scholar] [CrossRef]
- Simó, M.; Navarro, X.; Yuste, V.J.; Bruna, J. Autonomic Nervous System and Cancer. Clin. Auton. Res. 2018, 28, 301–314. [Google Scholar] [CrossRef] [PubMed]
- O’Higgins, C.M.; Brady, B.; O’Connor, B.; Walsh, D.; Reilly, R.B. The Pathophysiology of Cancer-Related Fatigue: Current Controversies. Support. Care Cancer Off. J. Multinatl. Assoc. Support. Care Cancer 2018, 26, 3353–3364. [Google Scholar] [CrossRef]
- Bracci, L.; Schiavoni, G.; Sistigu, A.; Belardelli, F. Immune-Based Mechanisms of Cytotoxic Chemotherapy: Implications for the Design of Novel and Rationale-Based Combined Treatments against Cancer. Cell Death Differ. 2014, 21, 15–25. [Google Scholar] [CrossRef]
- Mantovani, A.; Barajon, I.; Garlanda, C. IL-1 and IL-1 Regulatory Pathways in Cancer Progression and Therapy. Immunol. Rev. 2018, 281, 57–61. [Google Scholar] [CrossRef]
- Vyas, D.; Laput, G.; Vyas, A.K. Chemotherapy-Enhanced Inflammation May Lead to the Failure of Therapy and Metastasis. OncoTargets Ther. 2014, 7, 1015–1023. [Google Scholar] [CrossRef] [PubMed]
- Collado-Hidalgo, A.; Bower, J.E.; Ganz, P.A.; Cole, S.W.; Irwin, M.R. Inflammatory Biomarkers for Persistent Fatigue in Breast Cancer Survivors. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2006, 12, 2759–2766. [Google Scholar] [CrossRef]
- Parent-Roberge, H.; Fontvieille, A.; Maréchal, R.; Wagner, R.; Fülöp, T.; Pavic, M.; Riesco, E. Effects of Combined Exercise Training on the Inflammatory Profile of Older Cancer Patients Treated with Systemic Therapy. Brain Behav. Immun. Health 2019, 2, 100016. [Google Scholar] [CrossRef] [PubMed]
- Raudonis, B.M.; Kelley, I.H.; Rowe, N.; Ellis, J. A Pilot Study of Proinflammatory Cytokines and Fatigue in Women with Breast Cancer During Chemotherapy. Cancer Nurs. 2017, 40, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Cruz, F.M.; Munhoz, B.A.; Alves, B.C.; Gehrke, F.S.; Fonseca, F.L.; Kuniyoshi, R.K.; Cubero, D.; Peppone, L.J.; del Giglio, A. Biomarkers of Fatigue Related to Adjuvant Chemotherapy for Breast Cancer: Evaluation of Plasma and Lymphocyte Expression. Clin. Transl. Med. 2015, 4, 4. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, Ø.; Laird, B.; Aass, N.; Lea, T.; Fayers, P.; Kaasa, S.; Klepstad, P. The Relationship between Pro-Inflammatory Cytokines and Pain, Appetite and Fatigue in Patients with Advanced Cancer. PLoS ONE 2017, 12, e0177620. [Google Scholar] [CrossRef]
- Małkiewicz, M.A.; Szarmach, A.; Sabisz, A.; Cubała, W.J.; Szurowska, E.; Winklewski, P.J. Blood-Brain Barrier Permeability and Physical Exercise. J. Neuroinflamm. 2019, 16, 15. [Google Scholar] [CrossRef]
- Michael, B.D.; Griffiths, M.J.; Granerod, J.; Brown, D.; Keir, G.; Wnęk, M.; Cox, D.J.; Vidyasagar, R.; Borrow, R.; Parkes, L.M.; et al. The Interleukin-1 Balance During Encephalitis Is Associated with Clinical Severity, Blood-Brain Barrier Permeability, Neuroimaging Changes, and Disease Outcome. J. Infect. Dis. 2016, 213, 1651–1660. [Google Scholar] [CrossRef] [PubMed]
- Roerink, M.E.; van der Schaaf, M.E.; Dinarello, C.A.; Knoop, H.; van der Meer, J.W.M. Interleukin-1 as a Mediator of Fatigue in Disease: A Narrative Review. J. Neuroinflamm. 2017, 14, 16. [Google Scholar] [CrossRef]
- Wardill, H.R.; Mander, K.A.; Van Sebille, Y.Z.A.; Gibson, R.J.; Logan, R.M.; Bowen, J.M.; Sonis, S.T. Cytokine-Mediated Blood Brain Barrier Disruption as a Conduit for Cancer/Chemotherapy-Associated Neurotoxicity and Cognitive Dysfunction. Int. J. Cancer 2016, 139, 2635–2645. [Google Scholar] [CrossRef]
- Weiss, N.; Miller, F.; Cazaubon, S.; Couraud, P.-O. The Blood-Brain Barrier in Brain Homeostasis and Neurological Diseases. Biochim. Biophys. Acta 2009, 1788, 842–857. [Google Scholar] [CrossRef]
- Capuron, L.; Miller, A.H. Immune System to Brain Signaling: Neuropsychopharmacological Implications. Pharmacol. Ther. 2011, 130, 226–238. [Google Scholar] [CrossRef]
- Dobryakova, E.; Genova, H.M.; DeLuca, J.; Wylie, G.R. The Dopamine Imbalance Hypothesis of Fatigue in Multiple Sclerosis and Other Neurological Disorders. Front. Neurol. 2015, 6, 52. [Google Scholar] [CrossRef]
- Dietrich, J.; Prust, M.; Kaiser, J. Chemotherapy, Cognitive Impairment and Hippocampal Toxicity. Neuroscience 2015, 309, 224–232. [Google Scholar] [CrossRef]
- Hamid, A.A.; Pettibone, J.R.; Mabrouk, O.S.; Hetrick, V.L.; Schmidt, R.; Vander Weele, C.M.; Kennedy, R.T.; Aragona, B.J.; Berke, J.D. Mesolimbic Dopamine Signals the Value of Work. Nat. Neurosci. 2016, 19, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Petersen, A.M.W.; Pedersen, B.K. The Anti-Inflammatory Effect of Exercise. J. Appl. Physiol. 2005, 98, 1154–1162. [Google Scholar] [CrossRef] [PubMed]
- Windsor, M.T.; Bailey, T.G.; Perissiou, M.; Meital, L.; Golledge, J.; Russell, F.D.; Askew, C.D. Cytokine Responses to Acute Exercise in Healthy Older Adults: The Effect of Cardiorespiratory Fitness. Front. Physiol. 2018, 9, 203. [Google Scholar] [CrossRef] [PubMed]
- Fischer, C.P. Interleukin-6 in Acute Exercise and Training: What Is the Biological Relevance? Exerc. Immunol. Rev. 2006, 12, 6–33. [Google Scholar]
- Nash, D.; Hughes, M.G.; Butcher, L.; Aicheler, R.; Smith, P.; Cullen, T.; Webb, R. IL-6 Signaling in Acute Exercise and Chronic Training: Potential Consequences for Health and Athletic Performance. Scand. J. Med. Sci. Sports 2023, 33, 4–19. [Google Scholar] [CrossRef]
- Antunes, B.M.; Campos, E.Z.; Dos Santos, R.V.T.; Rosa-Neto, J.C.; Franchini, E.; Bishop, N.C.; Lira, F.S. Anti-Inflammatory Response to Acute Exercise Is Related with Intensity and Physical Fitness. J. Cell. Biochem. 2019, 120, 5333–5342. [Google Scholar] [CrossRef]
- Mee-inta, O.; Zhao, Z.-W.; Kuo, Y.-M. Physical Exercise Inhibits Inflammation and Microglial Activation. Cells 2019, 8, 691. [Google Scholar] [CrossRef]
- Cerqueira, É.; Marinho, D.A.; Neiva, H.P.; Lourenço, O. Inflammatory Effects of High and Moderate Intensity Exercise—A Systematic Review. Front. Physiol. 2020, 10, 1550. [Google Scholar] [CrossRef]
- Cianciulli, A.; Dragone, T.; Calvello, R.; Porro, C.; Trotta, T.; Lofrumento, D.D.; Panaro, M.A. Lofrumento IL-10 Plays a Pivotal Role in Anti-Inflammatory Effects of Resveratrol in Activated Microglia Cells. Int. Immunopharmacol. 2015, 24, 369–376. [Google Scholar] [CrossRef]
- Souza, P.S.; Gonçalves, E.D.; Pedroso, G.S.; Farias, H.R.; Junqueira, S.C.; Marcon, R.; Tuon, T.; Cola, M.; Silveira, P.C.L.; Santos, A.R.; et al. Physical Exercise Attenuates Experimental Autoimmune Encephalomyelitis by Inhibiting Peripheral Immune Response and Blood-Brain Barrier Disruption. Mol. Neurobiol. 2017, 54, 4723–4737. [Google Scholar] [CrossRef]
- Miggelbrink, A.M.; Jackson, J.D.; Lorrey, S.J.; Srinivasan, E.S.; Waibl-Polania, J.; Wilkinson, D.S.; Fecci, P.E. CD4 T-Cell Exhaustion: Does It Exist and What Are Its Roles in Cancer? Clin. Cancer Res. 2021, 27, 5742–5752. [Google Scholar] [CrossRef]
- Dolina, J.S.; Van Braeckel-Budimir, N.; Thomas, G.D.; Salek-Ardakani, S. CD8+ T Cell Exhaustion in Cancer. Front. Immunol. 2021, 12, 715234. [Google Scholar] [CrossRef]
- Mandarano, A.H.; Maya, J.; Giloteaux, L.; Peterson, D.L.; Maynard, M.; Gottschalk, C.G.; Hanson, M.R. Myalgic Encephalomyelitis/Chronic Fatigue Syndrome Patients Exhibit Altered T Cell Metabolism and Cytokine Associations. J. Clin. Investig. 2020, 130, 1491–1505. [Google Scholar] [CrossRef] [PubMed]
- Brenu, E.W.; van Driel, M.L.; Staines, D.R.; Ashton, K.J.; Hardcastle, S.L.; Keane, J.; Tajouri, L.; Peterson, D.; Ramos, S.B.; Marshall-Gradisnik, S.M. Longitudinal Investigation of Natural Killer Cells and Cytokines in Chronic Fatigue Syndrome/Myalgic Encephalomyelitis. J. Transl. Med. 2012, 10, 88. [Google Scholar] [CrossRef] [PubMed]
- Ojo-Amaize, E.A.; Conley, E.J.; Peter, J.B. Decreased Natural Killer Cell Activity Is Associated with Severity of Chronic Fatigue Immune Dysfunction Syndrome. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 1994, 18 (Suppl. S1), S157–S159. [Google Scholar] [CrossRef] [PubMed]
- Sand, K.L.; Flatebo, T.; Andersen, M.B.; Maghazachi, A.A. Effects of Exercise on Leukocytosis and Blood Hemostasis in 800 Healthy Young Females and Males. World J. Exp. Med. 2013, 3, 11–20. [Google Scholar] [CrossRef]
- Campbell, J.P.; Riddell, N.E.; Burns, V.E.; Turner, M.; van Zanten, J.J.C.S.V.; Drayson, M.T.; Bosch, J.A. Acute Exercise Mobilises CD8+ T Lymphocytes Exhibiting an Effector-Memory Phenotype. Brain. Behav. Immun. 2009, 23, 767–775. [Google Scholar] [CrossRef]
- Brenu, E.W.; Hardcastle, S.L.; Atkinson, G.M.; van Driel, M.L.; Kreijkamp-Kaspers, S.; Ashton, K.J.; Staines, D.R.; Marshall-Gradisnik, S.M. Natural Killer Cells in Patients with Severe Chronic Fatigue Syndrome. Autoimmun. Highlights 2013, 4, 69–80. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, S.; Zhang, B.; Qiao, L.; Zhang, Y.; Zhang, Y. T Cell Dysfunction and Exhaustion in Cancer. Front. Cell Dev. Biol. 2020, 8, 17. [Google Scholar] [CrossRef]
- Mathes, S.; Mester, J.; Bloch, W.; Wahl, P. Impact of High-Intensity and High-Volume Exercise on Short-Term Perturbations in the Circulating Fraction of Different Cell Types. J. Sports Med. Phys. Fitness 2017, 57, 130–137. [Google Scholar] [CrossRef]
- Dorneles, G.P.; Haddad, D.O.; Fagundes, V.O.; Vargas, B.K.; Kloecker, A.; Romão, P.R.T.; Peres, A. High Intensity Interval Exercise Decreases IL-8 and Enhances the Immunomodulatory Cytokine Interleukin-10 in Lean and Overweight-Obese Individuals. Cytokine 2016, 77, 1–9. [Google Scholar] [CrossRef]
- Parent-Roberge, H.; Fontvieille, A.; Tai, L.H.; Pavic, M.; Fülöp, T.; Riesco, E. Acute Nk Cells Response to Different Aerobic Exercise Modalities in Metastatic Cancer Patients Undergoing Chemotherapy. Med. Sci. Sports Exerc. 2023, 55, 479. [Google Scholar] [CrossRef]
- Bigley, A.B.; Rezvani, K.; Chew, C.; Sekine, T.; Pistillo, M.; Crucian, B.; Bollard, C.M.; Simpson, R.J. Acute Exercise Preferentially Redeploys NK-Cells with a Highly-Differentiated Phenotype and Augments Cytotoxicity against Lymphoma and Multiple Myeloma Target Cells. Brain. Behav. Immun. 2014, 39, 160–171. [Google Scholar] [CrossRef]
- Coletta, A.M.; Agha, N.H.; Baker, F.L.; Niemiro, G.M.; Mylabathula, P.L.; Brewster, A.M.; Bevers, T.B.; Fuentes-Mattei, E.; Basen-Engquist, K.; Gilchrist, S.C.; et al. The Impact of High-Intensity Interval Exercise Training on NK-Cell Function and Circulating Myokines for Breast Cancer Prevention among Women at High Risk for Breast Cancer. Breast Cancer Res. Treat. 2021, 187, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Saligan, L.N.; Olson, K.; Filler, K.; Larkin, D.; Cramp, F.; Yennurajalingam, S.; Sriram, Y.; Escalante, C.P.; del Giglio, A.; Kober, K.M.; et al. The Biology of Cancer-Related Fatigue: A Review of the Literature. Support. Care Cancer Off. J. Multinatl. Assoc. Support. Care Cancer 2015, 23, 2461–2478. [Google Scholar] [CrossRef]
- Olivas-Aguirre, M.; Torres-López, L.; Pottosin, I.; Dobrovinskaya, O. Overcoming Glucocorticoid Resistance in Acute Lymphoblastic Leukemia: Repurposed Drugs Can Improve the Protocol. Front. Oncol. 2021, 11, 617937. [Google Scholar] [CrossRef]
- Bower, J.E.; Ganz, P.A.; Dickerson, S.S.; Petersen, L.; Aziz, N.; Fahey, J.L. Diurnal Cortisol Rhythm and Fatigue in Breast Cancer Survivors. Psychoneuroendocrinology 2005, 30, 92–100. [Google Scholar] [CrossRef]
- Dinsen, S.; Baslund, B.; Klose, M.; Rasmussen, A.K.; Friis-Hansen, L.; Hilsted, L.; Feldt-Rasmussen, U. Why Glucocorticoid Withdrawal May Sometimes Be as Dangerous as the Treatment Itself. Eur. J. Intern. Med. 2013, 24, 714–720. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Miller, B.J.; Stefanek, M.E.; Miller, A.H. Inflammation-Induced Activation of the Indoleamine 2,3-Dioxygenase Pathway: Relevance to Cancer-Related Fatigue. Cancer 2015, 121, 2129–2136. [Google Scholar] [CrossRef]
- Johnson, T.S.; McGaha, T.; Munn, D.H. Chemo-Immunotherapy: Impact of Indoleamine 2,3-Dioxygenase in Defining Immunogenic versus Tolerogenic Cell Death in the Tumor Microenvironment. Adv. Exp. Med. Biol. 2017, 1036, 91–104. [Google Scholar] [CrossRef]
- Lanser, L.; Kink, P.; Egger, E.M.; Willenbacher, W.; Fuchs, D.; Weiss, G.; Kurz, K. Inflammation-Induced Tryptophan Breakdown Is Related with Anemia, Fatigue, and Depression in Cancer. Front. Immunol. 2020, 11, 249. [Google Scholar] [CrossRef]
- Heisler, L.K.; Pronchuk, N.; Nonogaki, K.; Zhou, L.; Raber, J.; Tung, L.; Yeo, G.S.H.; O’Rahilly, S.; Colmers, W.F.; Elmquist, J.K.; et al. Serotonin Activates the Hypothalamic-Pituitary-Adrenal Axis via Serotonin 2C Receptor Stimulation. J. Neurosci. Off. J. Soc. Neurosci. 2007, 27, 6956–6964. [Google Scholar] [CrossRef]
- Neefjes, E.C.W.; van der Vorst, M.J.D.L.; Blauwhoff-Buskermolen, S.; Verheul, H.M.W. Aiming for a Better Understanding and Management of Cancer-Related Fatigue. Oncologist 2013, 18, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.P.; Franco, N.F.; Varney, B.; Sundaram, G.; Brown, D.A.; de Bie, J.; Lim, C.K.; Guillemin, G.J.; Brew, B.J. Expression of the Kynurenine Pathway in Human Peripheral Blood Mononuclear Cells: Implications for Inflammatory and Neurodegenerative Disease. PLoS ONE 2015, 10, e0131389. [Google Scholar] [CrossRef]
- Frumento, G.; Rotondo, R.; Tonetti, M.; Damonte, G.; Benatti, U.; Ferrara, G.B. Tryptophan-Derived Catabolites Are Responsible for Inhibition of T and Natural Killer Cell Proliferation Induced by Indoleamine 2,3-Dioxygenase. J. Exp. Med. 2002, 196, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Evans, E.S.; Hackney, A.C.; Pebole, M.M.; McMurray, R.G.; Muss, H.B.; Deal, A.M.; Battaglini, C.L. Adrenal Hormone and Metabolic Biomarker Responses to 30 Min of Intermittent Cycling Exercise in Breast Cancer Survivors. Int. J. Sports Med. 2016, 37, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Evans, E.S.; Little, J.; McNeill, K.T.; Poole, C.; Bailey, S.P. Hypothalamic-Pituitary-Adrenal Axis Responses to an Acute Bout of Moderate Intensity Aerobic Exercise in Breast Cancer Survivors. FASEB J. 2019, 33, 534.3. [Google Scholar] [CrossRef]
- Duclos, M.; Gouarne, C.; Bonnemaison, D. Acute and Chronic Effects of Exercise on Tissue Sensitivity to Glucocorticoids. J. Appl. Physiol. 2003, 94, 869–875. [Google Scholar] [CrossRef]
- Chaouloff, F. Effects of Acute Physical Exercise on Central Serotonergic Systems. Med. Sci. Sports Exerc. 1997, 29, 58–62. [Google Scholar] [CrossRef]
- Basso, J.C.; Suzuki, W.A. The Effects of Acute Exercise on Mood, Cognition, Neurophysiology, and Neurochemical Pathways: A Review. Brain Plast. 2017, 2, 127–152. [Google Scholar] [CrossRef]
- Kurz, K.; Fiegl, M.; Holzner, B.; Giesinger, J.; Pircher, M.; Weiss, G.; Denz, H.A.; Fuchs, D. Fatigue in Patients with Lung Cancer Is Related with Accelerated Tryptophan Breakdown. PLoS ONE 2012, 7, e36956. [Google Scholar] [CrossRef]
- Lyon, D.E.; Walter, J.M.; Starkweather, A.R.; Schubert, C.M.; McCain, N.L. Tryptophan Degradation in Women with Breast Cancer: A Pilot Study. BMC Res. Notes 2011, 4, 156. [Google Scholar] [CrossRef]
- Metcalfe, A.J.; Koliamitra, C.; Javelle, F.; Bloch, W.; Zimmer, P. Acute and Chronic Effects of Exercise on the Kynurenine Pathway in Humans—A Brief Review and Future Perspectives. Physiol. Behav. 2018, 194, 583–587. [Google Scholar] [CrossRef]
- Joisten, N.; Walzik, D.; Metcalfe, A.J.; Bloch, W.; Zimmer, P. Physical Exercise as Kynurenine Pathway Modulator in Chronic Diseases: Implications for Immune and Energy Homeostasis. Int. J. Tryptophan Res. 2020, 13, 1178646920938688. [Google Scholar] [CrossRef] [PubMed]
- Agudelo, L.Z.; Femenía, T.; Orhan, F.; Porsmyr-Palmertz, M.; Goiny, M.; Martinez-Redondo, V.; Correia, J.C.; Izadi, M.; Bhat, M.; Schuppe-Koistinen, I.; et al. Skeletal Muscle PGC-1α1 Modulates Kynurenine Metabolism and Mediates Resilience to Stress-Induced Depression. Cell 2014, 159, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Joisten, N.; Kummerhoff, F.; Koliamitra, C.; Schenk, A.; Walzik, D.; Hardt, L.; Knoop, A.; Thevis, M.; Kiesl, D.; Metcalfe, A.J.; et al. Exercise and the Kynurenine Pathway: Current State of Knowledge and Results from a Randomized Cross-over Study Comparing Acute Effects of Endurance and Resistance Training. Exerc. Immunol. Rev. 2020, 26, 24–42. [Google Scholar] [PubMed]
- Cormie, P.; Lamb, S.; Newton, R.; Valentine, L.; McKiernan, S.; Spry, N.; Joseph, D.; Taaffe, D.; Galvao, D. One Size Doesn’t Fit All: Responders and Nonresponders to Exercise Interventions for Cancer Patients and Survivors. Asia-Pac. J. Clin. Oncol. 2017, 13, 85–86. [Google Scholar]
- Bouchard, C.; Rankinen, T. Individual Differences in Response to Regular Physical Activity. Med. Sci. Sports Exerc. 2001, 33, S446–S451; discussion S452–S453. [Google Scholar] [CrossRef]
- Morishita, S.; Hamaue, Y.; Fukushima, T.; Tanaka, T.; Fu, J.B.; Nakano, J. Effect of Exercise on Mortality and Recurrence in Patients with Cancer: A Systematic Review and Meta-Analysis. Integr. Cancer Ther. 2020, 19, 1534735420917462. [Google Scholar] [CrossRef]
- Cormie, P.; Zopf, E.M.; Zhang, X.; Schmitz, K.H. The Impact of Exercise on Cancer Mortality, Recurrence, and Treatment-Related Adverse Effects. Epidemiol. Rev. 2017, 39, 71–92. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fontvieille, A.; Parent-Roberge, H.; Fülöp, T.; Pavic, M.; Riesco, E. The Mechanisms Underlying the Beneficial Impact of Aerobic Training on Cancer-Related Fatigue: A Conceptual Review. Cancers 2024, 16, 990. https://doi.org/10.3390/cancers16050990
Fontvieille A, Parent-Roberge H, Fülöp T, Pavic M, Riesco E. The Mechanisms Underlying the Beneficial Impact of Aerobic Training on Cancer-Related Fatigue: A Conceptual Review. Cancers. 2024; 16(5):990. https://doi.org/10.3390/cancers16050990
Chicago/Turabian StyleFontvieille, Adeline, Hugo Parent-Roberge, Tamás Fülöp, Michel Pavic, and Eléonor Riesco. 2024. "The Mechanisms Underlying the Beneficial Impact of Aerobic Training on Cancer-Related Fatigue: A Conceptual Review" Cancers 16, no. 5: 990. https://doi.org/10.3390/cancers16050990