
Grade 2, 3 and Dedifferentiated Chondrosarcomas: A Comparative Study of Isocitrate Dehydrogenase-Mutant and Wild-Type Tumors with Implications for Prognosis and Therapy
 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Case Material
2.2. IDH Analysis
2.3. RNA Sequencing
2.4. RT-qPCR Analysis
2.5. Outcome Analysis
3. Results
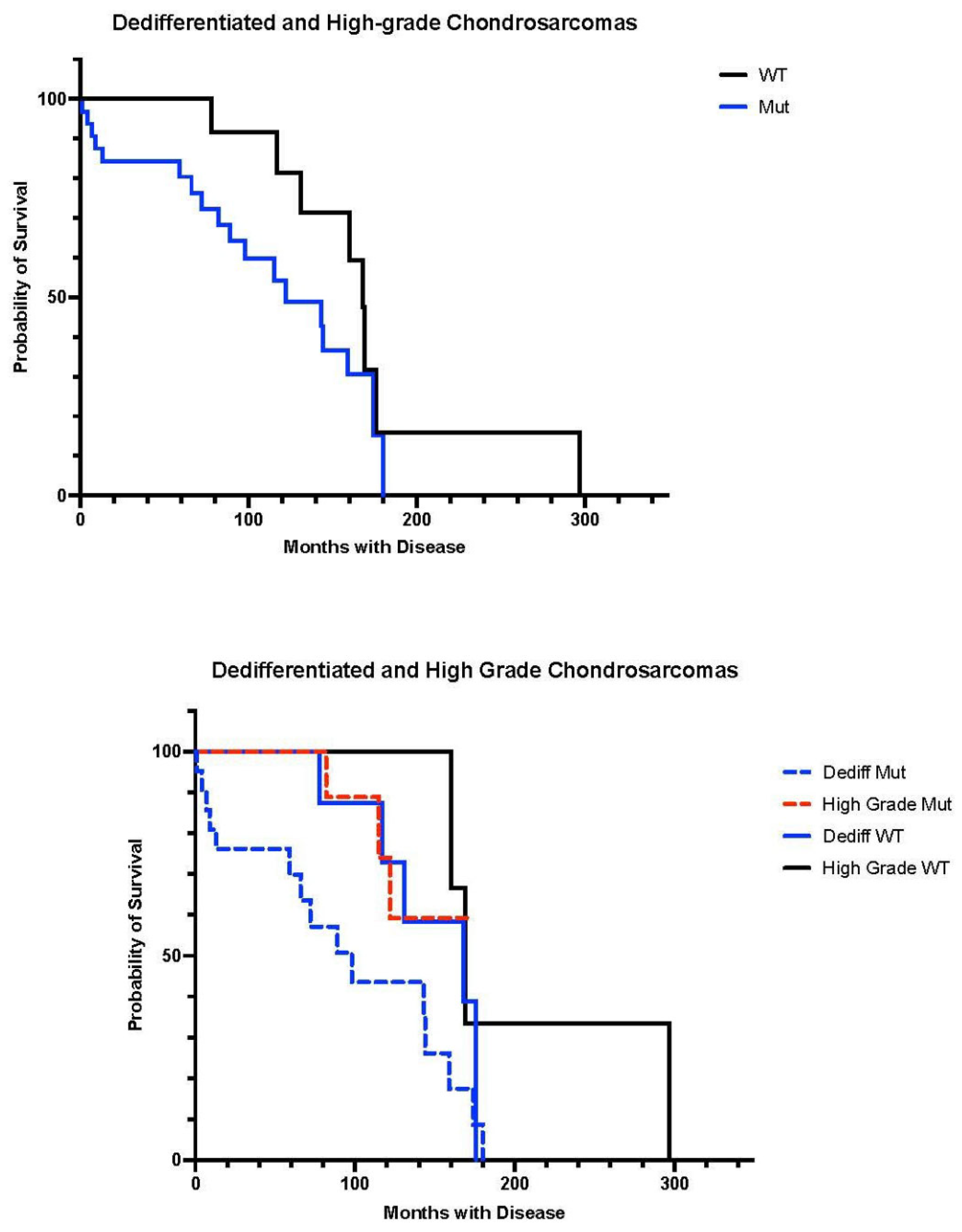
3.1. Patients
3.2. IDH Analysis
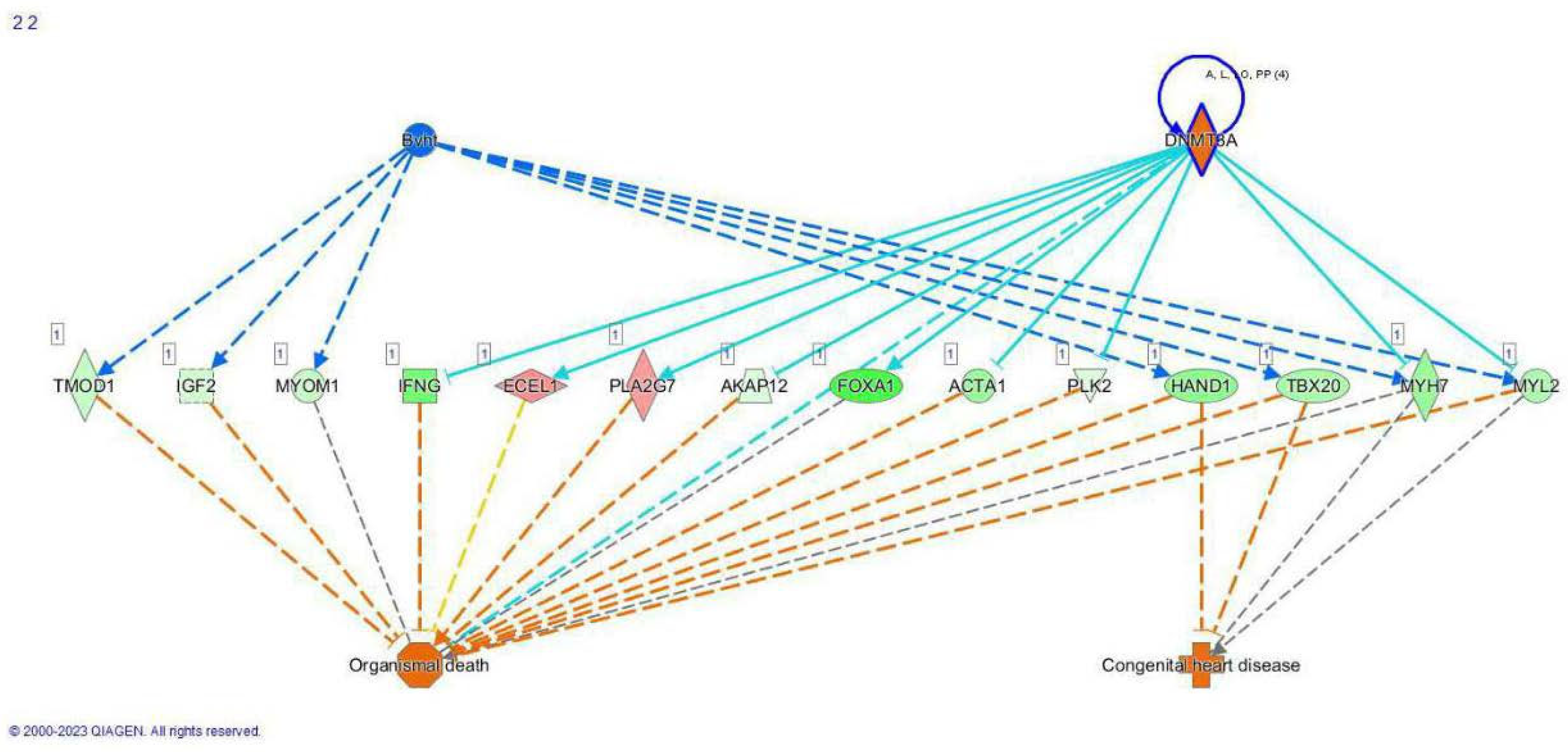
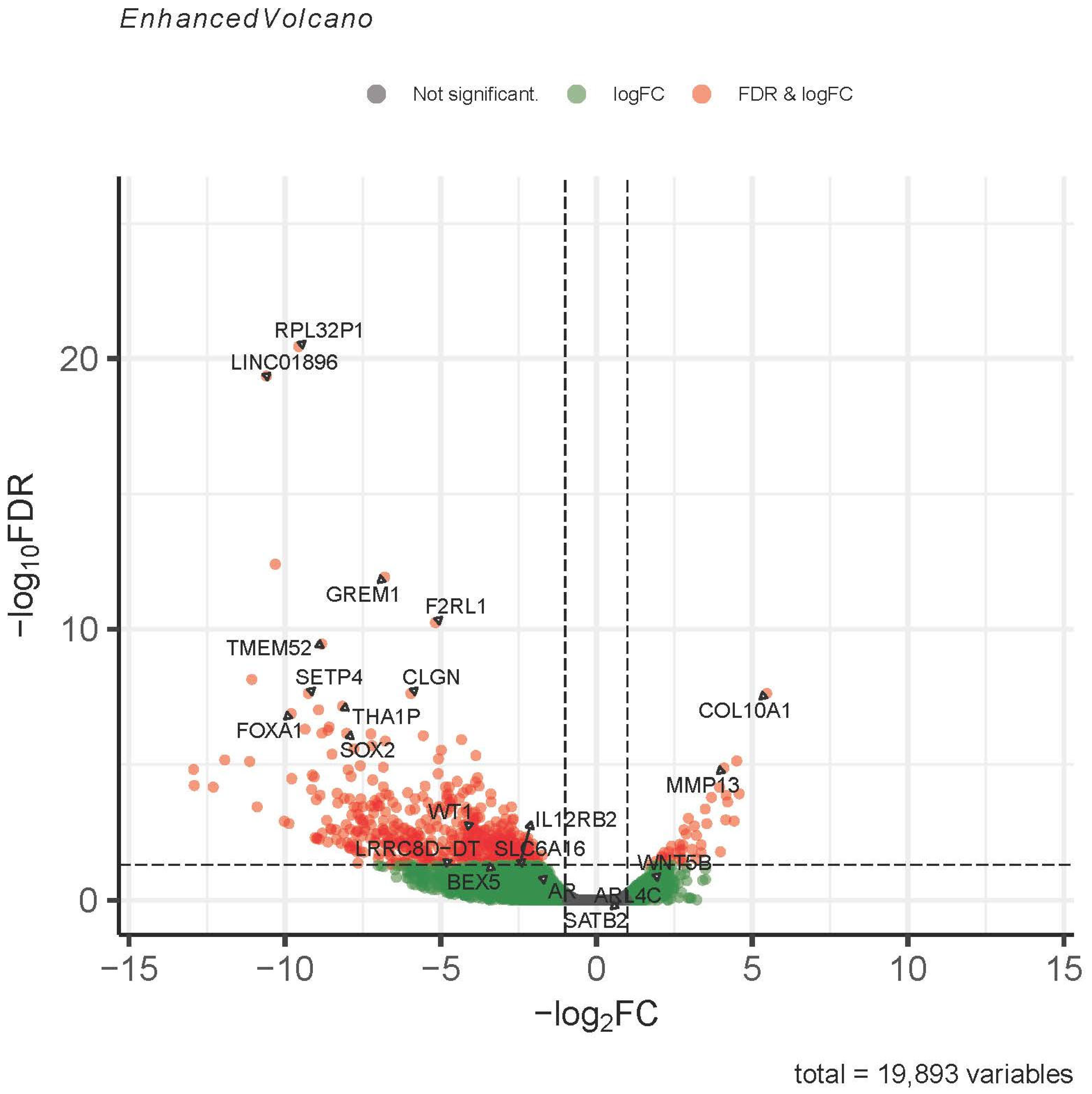
3.3. RNA Sequencing
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO Classification of Tumours Editorial Board. Soft tissue and bone tumours. In WHO Classification of Tumours Series, 5th ed.; IARC: Lyon, France, 2020; Volume 3. [Google Scholar]
- Nakagawa, M.; Sekimizu, M.; Endo, M.; Kobayashi, E.; Iwata, S.; Fukushima, S.; Yoshida, A.; Kitabayashi, I.; Ichikawa, H.; Kawai, A.; et al. Prognostic impact of IDH mutations in chondrosarcoma. J. Orthop. Sci. 2022, 27, 1315–1322. [Google Scholar] [CrossRef] [PubMed]
- Micaily, I.; Roche, M.; Ibrahim, M.; Martinez-Outschoorn, U.; Mallick, A. Metabolic pathways and targets in chondrosarcoma. Front. Oncol. 2021, 11, 772263. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Puviindran, V.; Puviindran, N.; Ding, X.; Shen, L.; Tang, Y.; Tsushima, H.; Yahara, Y.; Ban, G.; Zhang, G. Distinct roles of glutamine metabolism in benign and malignant cartilage tumors with IDH mutations. J. Bone Miner. Res. 2022, 37, 983–996. [Google Scholar] [CrossRef] [PubMed]
- Cross, W.; Lyskjaer, I.; Lesluyes, T.; Hargreaves, S.; Strobl, A.; Davies, C.; Waise, S.; Hames-Fathi, S.; Oukrif, D.; Ye, H.; et al. A genetic model for central chondrosarcoma evolution correlates with patient outcomes. Genome Med. 2022, 14, 99. [Google Scholar] [CrossRef] [PubMed]
- Miwa, S.; Yamamoto, N.; Hayashi, K.; Takeuchi, A.; Igarashi, K.; Tsuchiya, H. Therapeutic targets and emerging treatments in advanced chondrosarcoma. Int. J. Mol. Sci. 2022, 23, 1096. [Google Scholar] [CrossRef] [PubMed]
- Cairns, R.; Mak, T. Oncogenic isocitrate dehydrogenase mutations: Mechanisms, models and clinical opportunities. Cancer Discov. 2013, 3, 730–741. [Google Scholar] [CrossRef] [PubMed]
- Molenaar, R.; Wilmink, J. IDH1/2 mutations in cancer stem cells and their implications for differentiation therapy. J. Histochem. Cytochem. 2022, 70, 83–97. [Google Scholar] [CrossRef]
- Bovee, J.; Cleton-Jansen, A.; Rosenberg, C.; Taminiau, A.; Cornelisse, C.J.; Hogendoorn, P. Molecular genetic characterization of both components of a dedifferentiated chondrosarcoma, with implications for its histogenesis. J. Pathol. 1999, 189, 454–462. [Google Scholar] [CrossRef]
- Nielsen, G.; Rosenberg, A. Diagnostic Pathology Bone, 2nd ed.; Elsevier: Philadelphia, PA, USA, 2017. [Google Scholar]
- Tap, W.; Villalobos, V.; Cole, G.; Burris, H.; Janku, F.; Mir, O.; Beeram, M.; Wagner, A.; Jiang, L.; Wu, B.; et al. Phase 1 study of the mutant IDH1 inhibitor Ivosidenib: Safety and clinical activity in patients with advanced chondrosarcoma. J. Clin. Oncol. 2020, 38, 1693–1701. [Google Scholar] [CrossRef]
- National Comprehensive Cancer Network. NCCN Clinical Practice Guidelines in Oncology, Bone Cancer, Version 1.2024; National Comprehensive Cancer Network: Plymouth Meeting, PA, USA, 2023. [Google Scholar]
- Mitchell, A.; Ayoub, K.; Mangham, D.; Grimer, R.; Carter, S.; Tillman, R. Experience in the treatment of dedifferentiated chondrosarcoma. J. Bone Jt. Surg. Br. 2000, 82, 55–61. [Google Scholar] [CrossRef]
- Italiano, A.; Mir, O.; Cioffi, A.; Palmerini, E.; Piperno-Neumann, S.; Perrin, C.; Chaigneau, L.; Penel, N.; Duffaud, F.; Kurtz, J.; et al. Advanced chondrosarcomas: Role of chemotherapy and survival. Ann. Oncol. 2013, 24, 2916–2922. [Google Scholar] [CrossRef] [PubMed]
- Schuetze, S.; Bolejack, V.; Choy, E.; Ganjoo, K.; Staddon, A.; Chow, W.; Tawbi, H.; Samuels, B.; Patel, S.; von Mehren, M.; et al. Phase 2 study of dasatinib in patients with alveolar soft part sarcoma, chondrosarcoma, chordoma, epithelioid sarcoma, or solitary fibrous tumor. Cancer 2017, 123, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Chow, W.; Frankel, P.; Ruel, C.; Araujo, D.; Milhem, M.; Okuno, S.; Hartner, L.; Undevia, S.; Staddon, A. Results of a prospective phase 2 study of pazopanib in patients with surgically unresectable or metastatic chondrosarcoma. Cancer 2020, 126, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Horbinski, C.; Kofler, J.; Kelly, L.; Murdoch, G.; Nikiforova, M. Diagnostic use of IDH1/2 mutation analysis in routine clinical testing of formalin fixed, paraffin embedded glioma tissues. J. Neuropathol. Exp. Neurol. 2009, 68, 1319–1325. [Google Scholar] [CrossRef] [PubMed]
- Pirozzi, C.; Yan, H. The implications of IDH mutations for cancer development and therapy. Nat. Rev. Clin. Oncol. 2021, 18, 645–661. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Liu, Y.; Cai, S.; Qian, M.; Ding, J.; Larion, M.; Gilbert, M.; Yang, C. IDH mutation in glioma: Molecular mechanisms and potential therapeutic targets. Br. J. Cancer 2020, 122, 1580–1589. [Google Scholar] [CrossRef] [PubMed]
- WHO Classification of Tumours Editorial Board. Central Nervous System Tumours. In WHO Classification of Tumours Series, 5th ed.; IARC: Lyon, France, 2021; Volume 6. [Google Scholar]
- Tian, W.; Zhang, W.; Wang, Y.; Jin, R.; Wang, Y.; Guo, H.; Tang, Y.; Yao, X. Recent advances in IDH1 mutant inhibitor in cancer therapy. Front. Pharmacol. 2022, 13, 982424. [Google Scholar] [CrossRef] [PubMed]
- Nacev, B.; Sanchez-Vega, F.; Smith, S.; Antonescu, B.; Rosenbaum, E.; Shi, H.; Tang, C.; Socci, N.; Rana, S.; Gularte-Merida, R.; et al. Clinical sequencing of soft tissue and bone sarcomas delineates diverse genomic landscapes and potential therapeutic targets. Nat. Commun. 2022, 13, 3405. [Google Scholar] [CrossRef]
- Guo, C.; Pirozzi, C.; Lopez, G.; Yan, H. Isocitrate dehydrogenase mutations in gliomas: Mechanisms, biomarkers and therapeutic target. Curr. Opin. Neurol. 2011, 24, 648–652. [Google Scholar] [CrossRef]
- Nicolle, R.; Ayadi, M.; Gomez-Brouchet, A.; Armenoult, L.; Banneau, G.; Elarouci, N.; Tallegas, M.; Decouvelaere, A.; Aubert, S.; Redini, F.; et al. Integrated molecular characterization of chondrosarcoma reveals critical determinants of disease progression. Nat. Commun. 2019, 10, 4622. [Google Scholar] [CrossRef]
- Venneker, S.; Kruisselbrink, A.; Baranski, Z.; Palubeckaite, I.; Briaire-de Bruijn, I.; Oosting, J.; French, P.; Danen, E.; Bovee, J. Beyond the influence of IDH mutations: Exploring epigenetic vulnerabilities in chondrosarcoma. Cancers 2020, 12, 3589. [Google Scholar] [CrossRef] [PubMed]
- Amary, M.; Bacsi, K.; Maggiani, F.; Damato, S.; Halai, D.; Berisha, F.; Pollock, R.; O’Donnell, P.; Grigoriadis, A.; Diss, T.; et al. IDH1 and IDH2 mutations are frequent events in central chondrosarcoma and central and periosteal chondromas but not in other mesenchymal tumours. J. Pathol. 2011, 224, 334–343. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, N.; Wong, D.; Lum, A.; Lin, J.; Ho, J.; Lee, C.; Yip, S. Characterization of isocitrate dehydrogenase 1/isocitrate dehydrogenase 2 gene mutation and the d-2-hydroxyglutarate oncometabolite level in dedifferentiated chondrosarcoma. Histopathology 2020, 76, 722–730. [Google Scholar] [CrossRef] [PubMed]
- Stoeckl, S.; Lindner, G.; Li, S.; Schuster, P.; Haferkamp, S.; Wagner, F.; Prodinger, P.; Multhoff, G.; Boxberg, M.; Hillman, A.; et al. SOX9 knockout induces polyploidy and changes sensitivity to tumor treatment strategies in a chondrosarcoma cell line. Int. J. Mol. Sci. 2020, 21, 7627. [Google Scholar] [CrossRef] [PubMed]
- Blumer, M. Bone tissue and histological and molecular events during development of the long bones. Ann. Anat. 2021, 235, 151704. [Google Scholar] [CrossRef] [PubMed]
- Iseulys, R.; Gomez-Brouchet, A.; Bouvier, C.; Du Bouexic, G.; Karanian, M.; Blay, J.; Dutour, A. The immune landscape of chondrosarcoma reveals an immunosuppressive environment in the dedifferentiated subtypes and exposes CSFR1 + macrophages as a promising therapeutic target. J. Bone Oncol. 2020, 20, 100271. [Google Scholar] [PubMed]
- Tiet, T.; Hopyan, S.; Nadesan, P.; Gokgoz, N.; Poon, R.; Lin, A.; Yan, T.; Andrulis, I.; Alman, B.; Wunder, J. Constitutive hedgehog signaling in chondrosarcoma up-regulates tumor cell proliferation. Am. J. Pathol. 2006, 168, 321–330. [Google Scholar] [CrossRef]
- Meijer, D.; de Jong, D.; Pansuriya, T.C.; van den Akker, B.; Picci, P.; Szuhai, K.; Bovee, J. Genetic characterization of mesenchymal, clear cell and dedifferentiated chondrosarcoma. Genes Chromosomes Cancer 2012, 51, 899–909. [Google Scholar] [CrossRef]
- Gadelha, R.; Machado, C.; de Pinho Pessoa, F.; Pantoja, L.; Barreto, I.; Ribeiro, R.; de Moraes Filho, M.; de Moraes, M.; Khayat, A.; Moreira-Nunes, C. The role of WRAP53 in cell homeostasis and carcinogenesis onset. Curr. Issues Mol. Biol. 2022, 44, 5498–5514. [Google Scholar] [CrossRef]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef]
- Liu, Z.; Shen, F.; Wang, H.; Li, A.; Wang, J.; Du, L.; Liu, B.; Zhang, B.; Lian, X.; Pang, B.; et al. Abnormally high expression of HOXA2 as an independent factor for poor prognosis in glioma patients. Cell Cycle 2020, 19, 1632. [Google Scholar] [CrossRef] [PubMed]
- De Pagter-Holthuizen, P.; Jansen, M.; van der Kammen, R.; van Schaik, F.; Sussenback, J. Differential expression of the human insulin-like growth factor II gene. Characterization of the IGF-II mRNAs and an mRNA encoding a putative IGF-II associated protein. Biochem. Biophys. Acta 1988, 950, 282–295. [Google Scholar] [CrossRef] [PubMed]
- Palmini, G.; Marini, F.; Brandi, M. What is new in the miRNA world regarding osteosarcoma and chondrosarcoma? Molecules 2017, 22, 417. [Google Scholar] [CrossRef] [PubMed]
- Florke Gee, R.; Chen, H.; Lee, A.; Daly, C.; Wilander, B.; Tacer, K.; Potts, P. Emerging roles of the MAGE protein family in stress response pathways. J. Biol. Chem. 2020, 295, 16121–16155. [Google Scholar] [CrossRef]
- Kimura, K.; Matsumoto, S.; Harada, T.; Morii, E.; Nagatomo, I.; Shintani, Y.; Kikuchi, A. ARL4C is associated with initiation and progression of lung adenocarcinoma and represents a therapeutic target. Cancer Sci. 2020, 111, 951–961. [Google Scholar] [CrossRef]
- Available online: www.genecards.org (accessed on 1 September 2023).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IDH | ||||||
|---|---|---|---|---|---|---|
| Case | Age | Sex | Site | Size cm | Grade | Status |
| 1 | 37 | M | Arm | 22.5 | dd | Mut |
| 2 | 81 | M | Prox femur | 17.6 | dd | Mut |
| 3 | 82 | F | Thorax | 24.1 | 2 | WT |
| 4 | 57 | M | Pelvis | 9.8 | 3 | Mut |
| 5 | 70 | M | Sternum | - | 2 | WT |
| 6 | 85 | F | Humerus | - | dd | Mut |
| 7 | 72 | M | Femur | - | dd | Mut |
| 8 | 83 | F | Humerus | 7.2 | dd | - |
| 9 | 77 | M | Humerus | 9.2 | dd | Mut |
| 10 | 56 | M | Femur | - | 2 | Mut |
| Scapula | ||||||
| 11 | 62 | F | Chest | 5.8 | dd | Mut |
| 12 | 44 | M | Prox femur | 21.2 | dd | Mut |
| 13 | 76 | M | Chest wall | 12.5 | 3 | WT |
| 14 | 65 | M | Pelvis | 6.3 | 3 | - |
| 15 | 76 | M | Pelvis | - | 2 | Mut |
| 16 | 91 | M | Chest wall | 10 | dd | WT |
| 17 | 58 | M | Femur | - | 2 | Mut |
| 18 | 38 | M | Neck | - | 3 | Mut |
| 19 | 56 | F | Femur | 33 | dd | Mut |
| 20 | 74 | M | Pelvis | 13.2 | dd | WT |
| 21 | 61 | F | Pelvis | - | 3 | Mut |
| 22 | 84 | F | Hand | - | 2 | Mut |
| 23 | 54 | F | Femur | 9 | 2 | - |
| 24 | 79 | M | Prox femur | 17.5 | dd | Mut |
| 25 | 70 | F | Femur | - | 2 | Mut |
| 26 | 72 | F | Rib | 7 | 2 | WT |
| 27 | 62 | M | Pelvis | 9 | dd | Mut |
| 28 | 72 | F | Pelvis | 13.5 | 2 | - |
| 29 | 67 | M | Femur | 27 | dd | WT |
| 30 | 74 | M | Sacrum | 6.2 | dd | - |
| 31 | 57 | M | Humerus | 9.7 | 2 | - |
| 32 | 71 | M | Chest wall | 4.5 | 2 | - |
| 33 | 88 | F | Humerus | - | 2 | Mut |
| 34 | 14 | F | Pelvis | 6 | 2 | - |
| 35 | 62 | F | Chest wall | - | 2 | - |
| 36 | 83 | F | Femur | 6 | 3 | Mut |
| 37 | 80 | M | Pelvis | - | dd | WT |
| 38 | 63 | F | Talus | 11 | dd | - |
| 39 | 70 | M | Humerus | 7 | dd | - |
| 40 | 18 | F | Femur | - | 2 | - |
| 41 | 74 | M | Chest wall | 2 | 2 | - |
| 42 | 83 | M | Rib | 6 | dd | WT |
| 43 | 88 | M | Pelvis | 5.1 | 2 | - |
| 44 | 64 | M | Pelvis | 10.6 | dd | Mut |
| 45 | 65 | F | Femur | 16 | dd | - |
| 46 | 66 | M | Sternum | 8 | 2 | - |
| 47 | 70 | F | Humerus | - | dd | Mut |
| 48 | 50 | M | Femur | 5.5 | 2 | Mut |
| 49 | 88 | F | Femur | 11 | dd | Mut |
| 50 | 51 | M | Scapula | - | 2 | WT |
| 51 | 34 | F | Humerus | 5.5 | 2 | - |
| 52 | 49 | F | Thigh | - | 3 | - |
| 53 | 43 | M | Pelvis | 25 | 2 | WT |
| 54 | 74 | F | Femur | - | dd | WT |
| 55 | 56 | M | Pelvis | 18 | 2 | - |
| 56 | 84 | M | Pelvis | 9 | 2 | Mut |
| 57 | 64 | M | Femur | 11.5 | dd | WT |
| 58 | 82 | F | Femur | 3.5 | 2 | WT |
| 59 | 68 | M | Pelvis | 5.5 | 2 | Mut |
| 60 | 70 | M | Tibia | 9.5 | dd | WT |
| 61 | 67 | F | Humerus | 14 | dd | Mut |
| 62 | 56 | M | Femur | 10 | dd | Mut |
| 63 | 71 | F | Femur | 20.5 | dd | Mut |
| 64 | 32 | M | Pelvis | 18 | dd | Mut |
| 65 | 74 | F | Femur | 6 | dd | - |
| 66 | 49 | M | Pelvis | 10.5 | dd | - |
| 67 | 75 | M | Humerus | 9.5 | dd | - |
| 68 | 57 | M | Pelvis | 12 | dd | Mut |
| 69 | 56 | F | Pelvis | - | dd | Mut |
| DEG Higher in IDH WT | DEG Higher in IDH Mut |
|---|---|
| GREM1 | COL10A1 |
| TMEM52 | MMP13 |
| FOXA1 | HHIP |
| ALX Homeobox 1, 3 | IBSP |
| HOXA2 | COL26A1 |
| WT1 | WRAP53 |
| BMP7 | TP53 |
| SOX2 | |
| IGF2 | |
| MAGEA1 | |
| NES | |
| ERG | |
| FGF18 | |
| DKK2 |
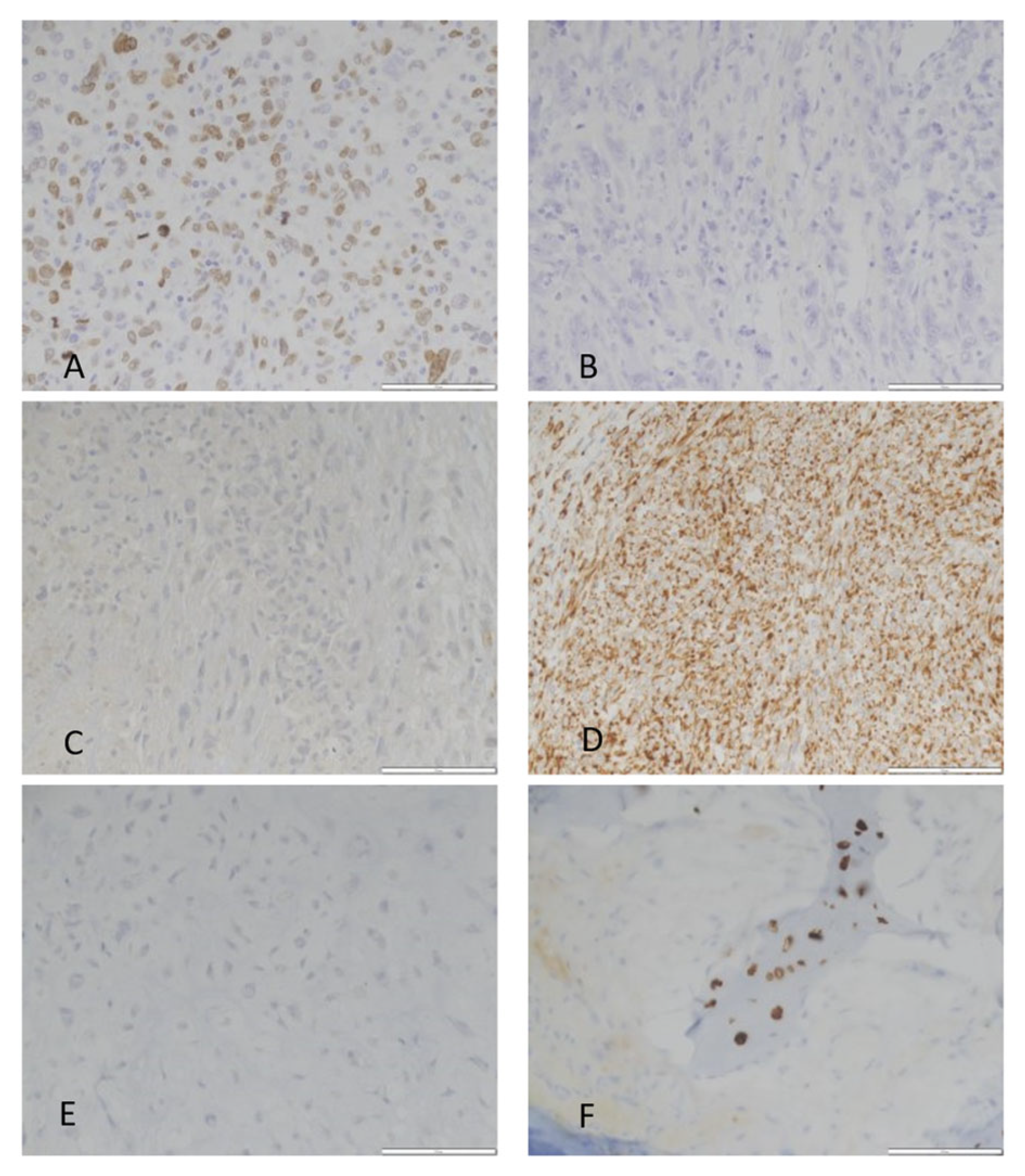
| Case | COL10A1 | MMP13 | SATB2 | IDH Mutation | Grade |
|---|---|---|---|---|---|
| 9 | Expressed | Expressed | Expressed | IDH1 R132S | Dedifferentiated |
| 6 | Expressed | Low Expression | Expressed | IDH2 R172S | Dedifferentiated |
| 17 | Expressed | Expressed | Expressed | IDH1 R132L | 2 |
| 4 | High Expression | High Expression | High Expression | IDH1 R132C | 3 |
| 2 | High Expression | High Expression | High Expression | IDH1 R132C | Dedifferentiated |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schoedel, K.; Heim, T.; Duensing, A.; Lohse, I.; Presutti, L.; Belayneh, R.; Bhogal, S.; Singh-Varma, A.; Chang, A.; Chandran, U.; et al. Grade 2, 3 and Dedifferentiated Chondrosarcomas: A Comparative Study of Isocitrate Dehydrogenase-Mutant and Wild-Type Tumors with Implications for Prognosis and Therapy. Cancers 2024, 16, 247. https://doi.org/10.3390/cancers16020247
Schoedel K, Heim T, Duensing A, Lohse I, Presutti L, Belayneh R, Bhogal S, Singh-Varma A, Chang A, Chandran U, et al. Grade 2, 3 and Dedifferentiated Chondrosarcomas: A Comparative Study of Isocitrate Dehydrogenase-Mutant and Wild-Type Tumors with Implications for Prognosis and Therapy. Cancers. 2024; 16(2):247. https://doi.org/10.3390/cancers16020247
Chicago/Turabian StyleSchoedel, Karen, Tanya Heim, Anette Duensing, Ines Lohse, Laura Presutti, Rebekah Belayneh, Sumail Bhogal, Anya Singh-Varma, Alexander Chang, Uma Chandran, and et al. 2024. "Grade 2, 3 and Dedifferentiated Chondrosarcomas: A Comparative Study of Isocitrate Dehydrogenase-Mutant and Wild-Type Tumors with Implications for Prognosis and Therapy" Cancers 16, no. 2: 247. https://doi.org/10.3390/cancers16020247