
MiRNAs in Alcohol-Related Liver Diseases and Hepatocellular Carcinoma: A Step toward New Therapeutic Approaches?

Abstract
:Simple Summary
Abstract
1. Introduction
2. MicroRNAs


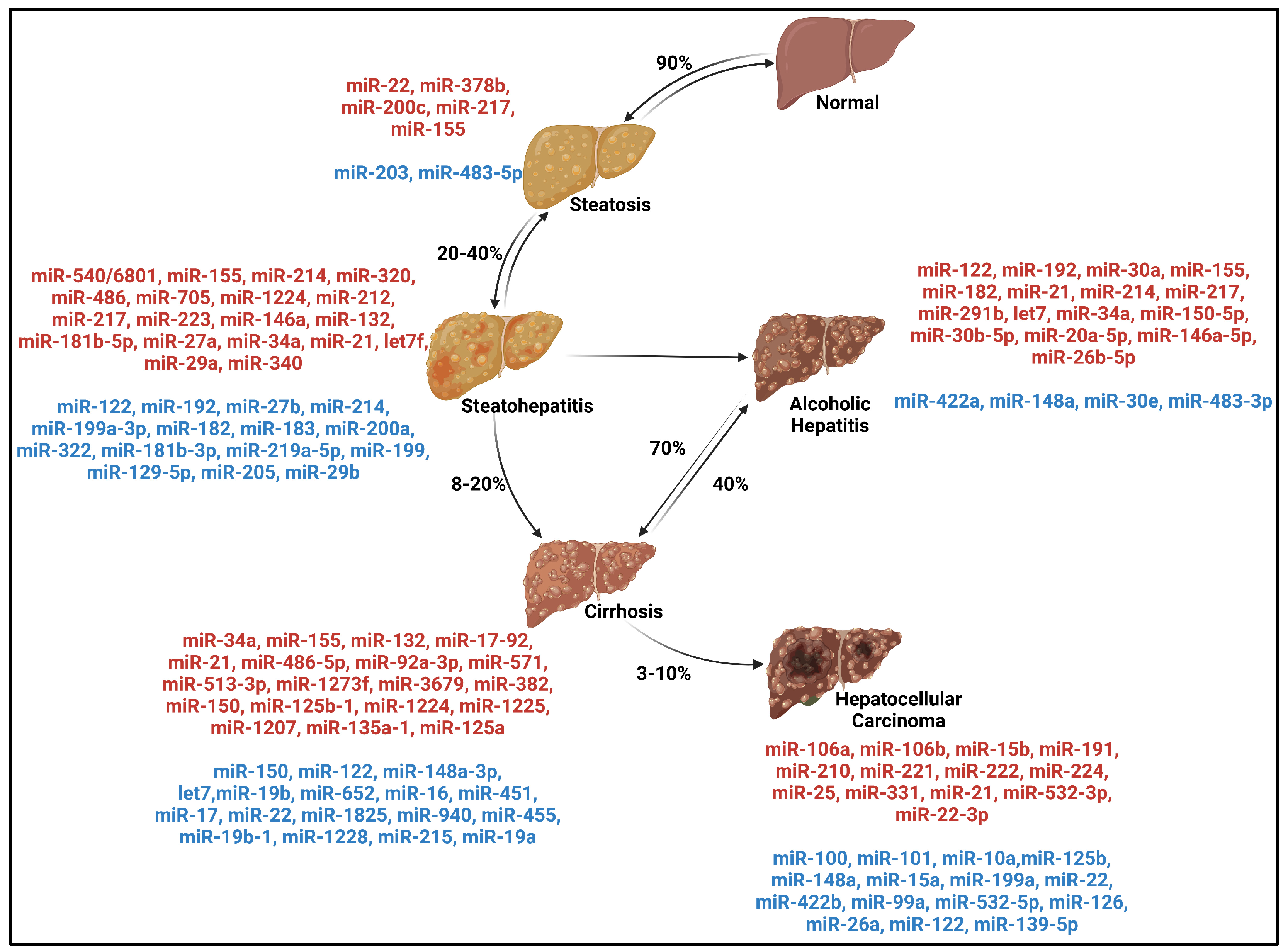{kind=link}
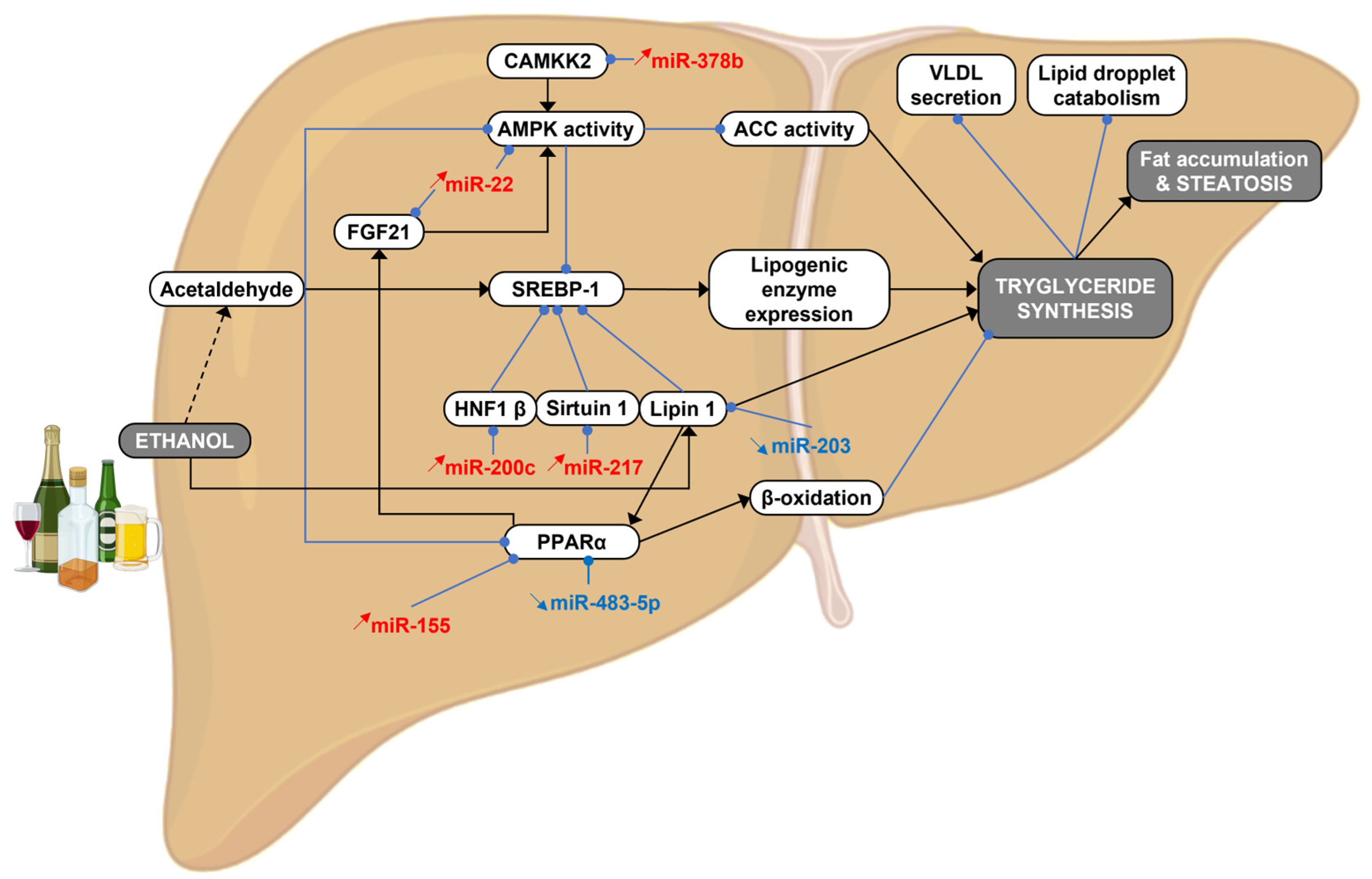{kind=link}
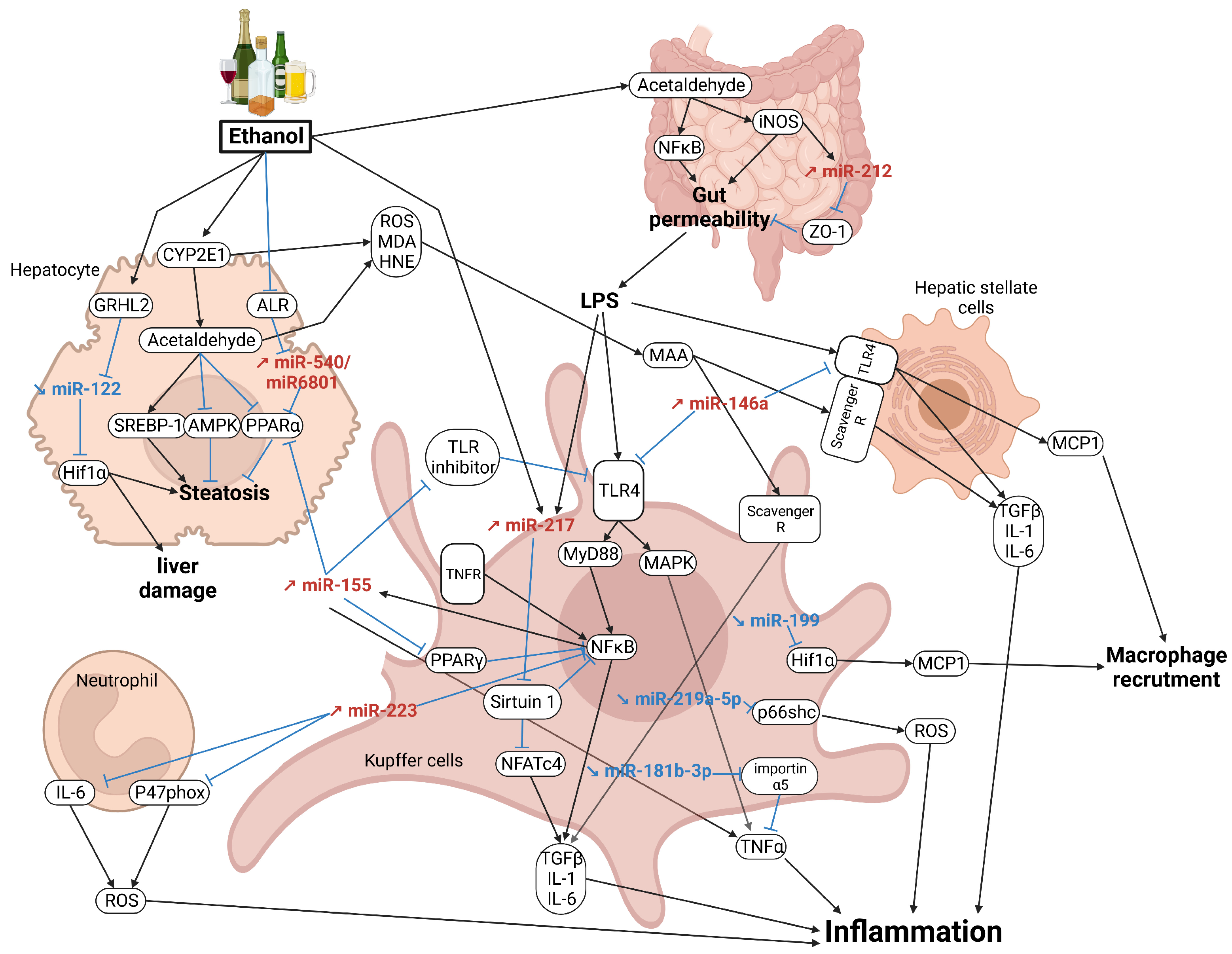{kind=link}
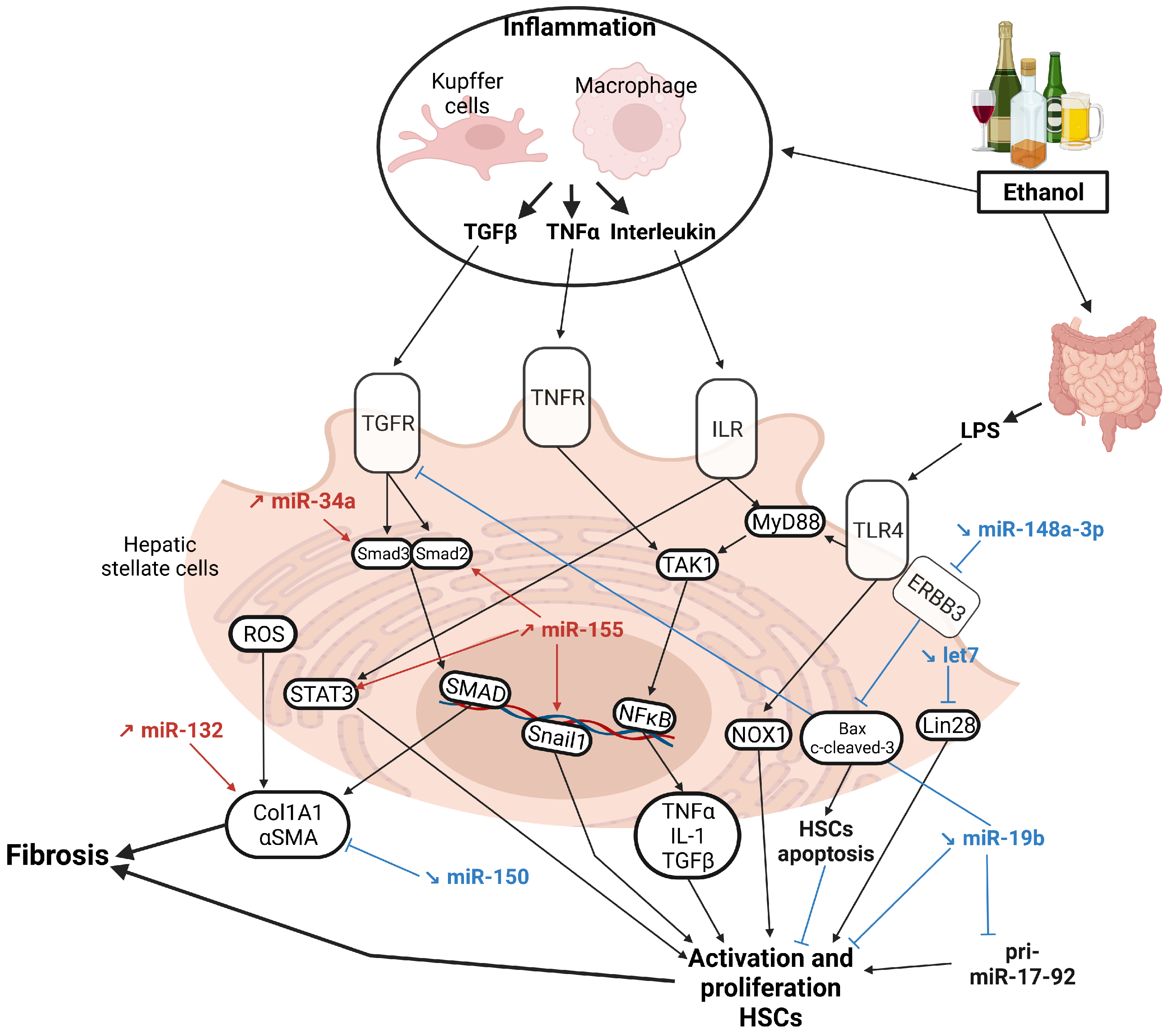{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MiRs | Expression | Function | Target | Model | Cell Types | Refs. |
|---|---|---|---|---|---|---|
| Alcohol-Related Steatosis | ||||||
| MiR-203 | Down | Decrease lipid accumulation | Lipin1 | AML12 | Hepatocyte | [37] |
| MiR-483-5p | Down | Steatosis cell proliferation | PPARα | Human Mice HepaRG | Hepatocyte | [38] |
| MiR-22 | Up | Steatosis | FGFR1 FGF21 IL6/JAK/STAT | Human Mice | Hepatocyte | [39,40,41] |
| MiR-378b | Up | Lipid accumulation | CAMKK2 | Mice Human Hepatocyte | Hepatocyte | [42] |
| MiR-200c | Up | Modulation of lipid homeostasis | Hnf1 Homeobox B | Mice | Liver | [43] |
| MiR-217 | Up | Inflammation Steatosis | Sirtuin-1 | Mice RAW 264.7 Kupffer cells | Hepatocyte Macrophage Kupffer cells | [44,45] |
| MiR-155 | Up | Promote liver steatosis Liver injury Inflammation Fibrosis | Snail1 Smad2 STAT3 PPARα TLR inhibitor PPARγ TNFα | Human Mice Raw 264.7 Hepa 1-6 | Kupffer cells Hepatocyte Hepatic stellate cells | [46,47,48,49,50] |
| Alcoholic Steatohepatitis | ||||||
| MiR-122 | Down | Protection against steatosis fibrosis | HiF1α TNFrsf13C | Human Mice RAW 264.7 Huh7 | Hepatocyte Kupffer cells Extracellular vesicles | [51,52,53,54] |
| MiR-192 | Down | Exosome induction | Rab27a Rab35 STX7 STX16 | Human | Hepatocyte Extracellular vesicles | [53,55] |
| MiR-27b | Down | Inflammation | LPS | Mice RAW 264.7 | Macrophage | [56] |
| MiR-214 | Down (mice) Up (human) | Liver fibrogenesis Induce oxidative stress | Gluthatione reductase | Human Mice Rat Bel7402 BRL | Hepatocyte | [40,57,58] |
| MiR-199a-3p | Down | n.a | n.a | Mice | n.a | [57] |
| MiR-182 | Down | Inflammation Apoptosis | Mcp-1 Ccl20 Cxcl5 Cxcl1 Bcl2 | Mice Human | Liver | [40,57] |
| MiR-183 | Down | Inflammatory | n.a | Mice | n.a | [57] |
| MiR-200a | Down | Disease severity | Gli2 | Mice | n.a | [57,59,60] |
| MiR-322 | Down | n.a | n.a | Mice | n.a | [57] |
| MiR-181b-3p | Down | Inflammatory | Importin α5 | Mice Rat | Kupffer cells | [61,62] |
| MiR-219a-5p | Down | Oxidative stress | P66shc | Rat AML-12 | Hepatocyte | [63] |
| MiR-199 | Down | Inflammation | Hif1α | Rat | Kupffer cells | [64] |
| MiR-129-5p | Down | Hepatic fibrosis Lipid metabolism | NEAT1 | Human Mice AML-12 | Hepatocyte | [65] |
| MiR-540 | Up | Hepatic steatosis Oxidative stress | PPARα ACOX1 | Mice | n.a | [66] |
| MiR-6801 | Up | Hepatic steatosis Oxidative stress | PPARα ACOX1 | Human | n.a | [66] |
| MiR-155 | Up | Promote liver steatosis Liver injury Inflammation fibrosis | Snail1 Smad2 STAT3 PPARα TLR inhibitor PPARγ TNFα | Human Mice Raw 264.7 Hepa 1-6 | Kupffer cells Hepatocyte Hepatic stellate cells | [46,47,48,49,50] |
| MiR-320 | Up | Inflammatory | n.a | Mice | n.a | [57] |
| MiR-486 | Up | Inflammatory | n.a | Mice | n.a | [57] |
| MiR-705 | Up | Inflammatory | n.a | Mice | n.a | [57] |
| MiR-1224 | Up | Inflammatory Tumor suppressor | n.a | Mice Human | Liver | [40,57] |
| MiR-212 | Up | Gut leakiness | ZO-1 | Mice | Gut epithelial cells | [67,68] |
| MiR-223 | Up | Inflammation Liver injury | IL-6 p47phox NFκB | Human Mice | Neutrophils Kupffer cells | [69,70,71] |
| MiR-146a | Up | Anti-inflammatory | TLR | Human Mice | Monocyte Kupffer cells | [72] |
| MiR-132 | Up | Inflammation Fibrosis | αSMA Collagen fibers Caspase 3 extracellular vesicles | Human Mice | Kupffer cells Hepatic stellate cells | [73,74] |
| MiR-181b-5p | Up | Oxidative stress Inflammation | PIAS1 | Rat | Hepatocyte | [75] |
| MiR-27a | Up | Fibrosis monocyte differentiation | ERK Sprouty 2 Nr1d2 CD206 | HumanHuh7.5 cells Monocytes | Kuppfer cells Monocytes Extracellular vesicles | [76,77,78] |
| MiR-34a | Up | Fibrosis Cellular senescence Mallory–Denk cell formation | Smad3 SIRT1 | Human Mice | Kuppfer cells Hepatocyte Hepatic stellate cells Mallory–Denk cells | [79,80,81,82,83] |
| MiR-21 | Up | Regulate hepatic cell survival, transformation, and remodel liver regeneration | VHL Fas ligand (TNF superfamily, member 6) (FASLG) and death receptor 5 (DR5) | Rat Human Mice | Hepatic stellate cells Kuppfer cells Hepatocyte | [40,84,85,86] |
| Let-7f | Up | Potential biomarkers Potential mediators of intercellular crossovers | n.a | Mice | Extracellular vesicles | [87] |
| MiR-29a | Up | Potential biomarkers Potential mediators of intercellular crossovers | n.a | Mice | Extracellular vesicles | [87] |
| MiR-340 | Up | Potential biomarkers Potential mediators of intercellular crossovers | n.a | Mice | Extracellular vesicles | [87] |
| MiR-205 | Down | Inflammation | Importinα5 | Mice | Kupffer cells | [88] |
| MiR-29b | Down | Inflammation | STAT3 | Mice RAW264.7 | Kupffer cells | [89] |
| MiR-217 | Up | Inflammation Steatosis | Sirtuin-1 | Mice RAW 264.7 Kupffer cells | Hepatocyte Macrophage Kupffer cells | [44,45] |
| Cirrhosis | ||||||
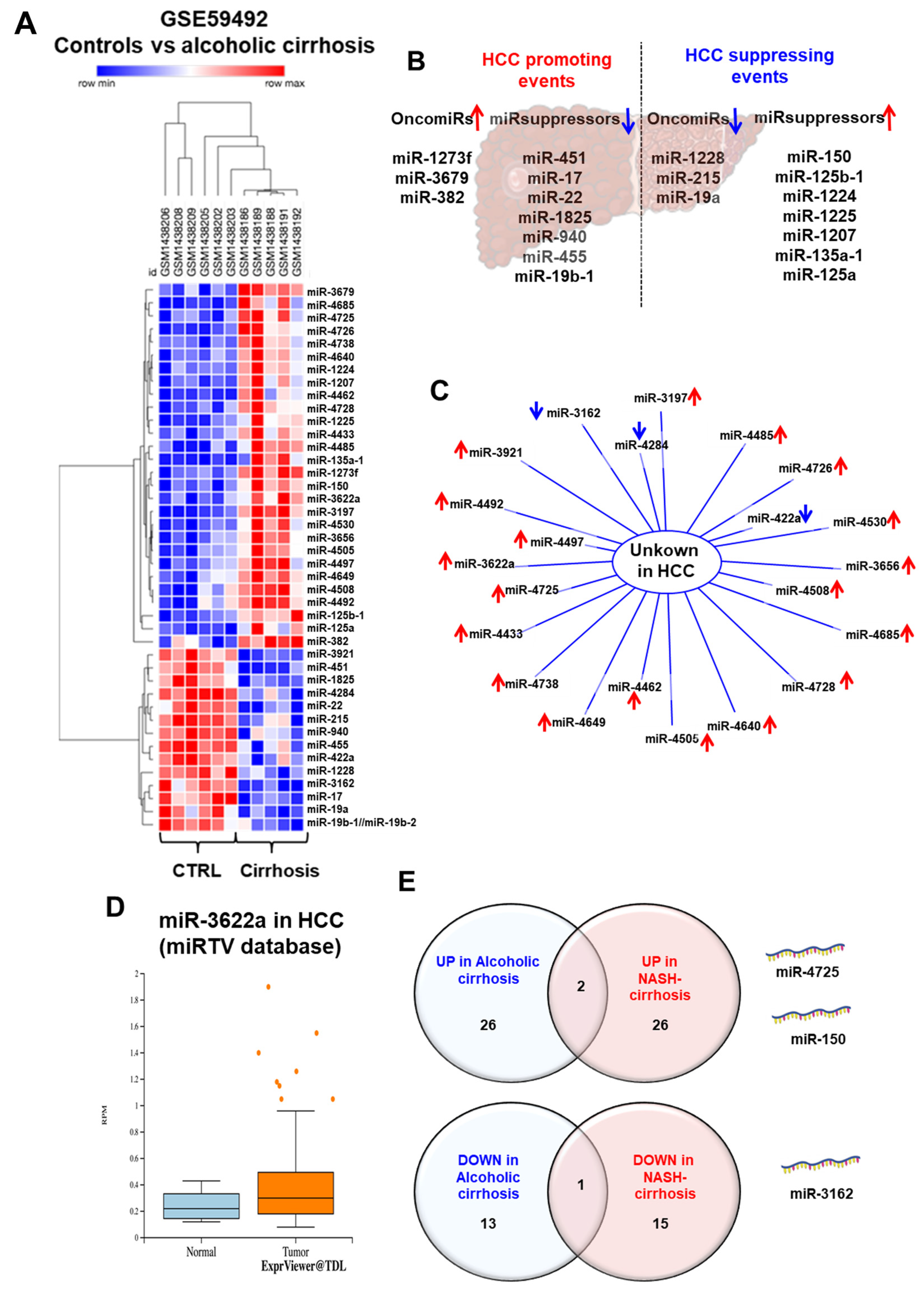
| MiR-150 | Up Down | Antifibrotic Tumor suppressor | αSMA Col1A1 | Human | Hepatic stellate cells | [40,90] |
| MiR-148a-3p | Down | Fibrosis | ERBB3 | Rat | Hepatic stellate cells | [91] |
| Let-7 | Down | Fibrosis Inflammatory | Lin28 TLR7 | Mice Human | Hepatic stellate cells | [92,93] |
| MiR-19b | Down | HSCs activation | Pri-miR-17-92 TGFβRII MeCP2 | Rat LX2 HepG2 | Hepatic stellate cells | [94] |
| MiR-652 | Down | n.a | n.a | Human | n.a | [95,96] |
| MiR-16 | Down | n.a | n.a | Human | Exosome | [97] |
| MiR-451 | Down | Tumor suppressor | n.a | Human | Liver | [40] |
| MiR-17 | Down | Tumor suppressor | n.a | Human | Liver | [40] |
| MiR-1825 | Down | Tumor suppressor | n.a | Human | Liver | [40] |
| MiR-940 | Down | Tumor suppressor | n.a | Human | Liver | [40] |
| MiR-455 | Down | Tumor suppressor | n.a | Human | Liver | [40] |
| MiR-19b-1 | Down | Tumor suppressor | n.a | Human | Liver | [40] |
| MiR-1228 | Down | OncomiR | n.a | Human | Liver | [40] |
| MiR-215 | Down | OncomiR | n.a | Human | Liver | [40] |
| MiR-19a | Down | OncomiR | n.a | Human | Liver | [40] |
| MiR-17-92 | Up | Fibrogenesis | n.a | n.a | Hepatic stellate cells | [94] |
| MiR-486-5p | Up | n.a | n.a | Human | n.a | [95,96] |
| MiR-92a-3p | Up | n.a | n.a | Human | n.a | [95,96] |
| MiR-571 | Up | n.a | CREBBP | Human | Hepatic stellate cells | [95,96,98] |
| MiR-513-3p | Up | n.a | n.a | Human | n.a | [95,96] |
| MiR-1273f | Up | OncomiR | n.a | Human | Liver | [40] |
| MiR-3679 | Up | OncomiR | n.a | Human | Liver | [40] |
| MiR-382 | Up | OncomiR | n.a | Human | Liver | [40] |
| MiR-125b-1 | Up | Tumor suppressor | n.a | Human | Liver | [40] |
| MiR-1225 | Up | Tumor suppressor | n.a | Human | Liver | [40] |
| MiR-1207 | Up | Tumor suppressor | n.a | Human | Liver | [40] |
| MiR-135a-1 | Up | Tumor suppressor | n.a | Human | Liver | [40] |
| MiR-125a | Up | Tumor suppressor | n.a | Human | Liver | [40] |
| MiR-22 | Down | Steatosis Tumor suppressor Deregulated pathways in HCC | FGFR1 FGF21 IL6/JAK/STAT | Human Mice | Hepatocyte | [39,40,41] |
| MiR-122 | Down | Protection against steatosis fibrosis | HiF1α TNFrsf13C | Human Mice RAW 264.7 Huh7 | Hepatocyte Kupffer cells Extracellular vesicles | [51,52,53,54] |
| MiR-155 | Up | Promote liver steatosis Liver injury Inflammation fibrosis | Snail1 Smad2 STAT3 PPARα TLR inhibitor PPARγ TNFα | Human Mice Raw 264.7 Hepa 1-6 | Kupffer cells Hepatocyte Hepatic stellate cells | [46,47,48,49,50] |
| MiR-1224 | Up | Inflammatory Tumor suppressor | n.a | Mice Human | n.a | [40,57] |
| MiR-132 | Up | Inflammation Fibrosis | αSMA Collagen fibers Caspase 3 extracellular vesicles | Human Mice | Kupffer cells Hepatic stellate cells | [73,74] |
| MiR-34a | Up | Fibrosis Cellular senescence Mallory–Denk cell formation | Smad3 SIRT1 | Human Mice | Kuppfer cells Hepatocyte Hepatic stellate cells Mallory–Denk cells | [79,80,81,82,83] |
| MiR-21 | Up | Regulate hepatic cell survival, transformation, and remodel liver regeneration | VHL Fas ligand (TNF superfamily, member 6) (FASLG) and death receptor 5 (DR5) | Rat Human Mice | Hepatic stellate cells Kuppfer cells Hepatocyte | [40,84,85,86] |
| Alcoholic Hepatitis | ||||||
| MiR-422a | Down | n.a | n.a | Human | n.a | [40] |
| MiR-30b-5p | Up | Associated mortality | n.a | Human | Extracellular vesicles | [99] |
| MiR-20a-5p | Up | Associated mortality | n.a | Human | Extracellular vesicles | [99] |
| MiR-26b-5p | Up | Associated mortality | n.a | Human | Extracellular vesicles | [99] |
| MiR-148a | Down | Anti-inflammatory Deregulated pathways in HCC | TXNIP Epigenetics TGFβ PI3K/AKT | Human Mice | Hepatocyte | [41,100] |
| MiR-30e | Down | Inflammation | UCP2 ATP H2O2 | Mice | n.a | [101] |
| MiR-483-3p | Down | Mallory–Denk cell formation | BRCA1 | Human | Mallory–Denk cells | [82] |
| MiR-146a-5p | Up | Associated mortality | n.a | Human | Extracellular vesicles | [99] |
| MiR-30a | Up | Autophagy | Beclin-1 | Human | Exosome | [53,102] |
| MiR-291b | Up | Inflammation | Tollip | Human Rat | Kupffer cells | [103] |
| MiR-150-5p | Up | Cell death | CISH | Human | Liver | [104] |
| MiR-217 | Up | Inflammation Steatosis | Sirtuin-1 | Mice RAW 264.7 Kupffer cells | Hepatocyte Macrophage Kupffer cells | [44,45] |
| MiR-122 | Up | Protection against steatosis fibrosis | HiF1α TNFrsf13C | Human Mice RAW 264.7 Huh7 | Hepatocyte Kupffer cells Extracellular vesicles | [51,52,53,54] |
| MiR-192 | Up | Exosome induction | Rab27a Rab35 STX7 STX16 | Human | Hepatocyte Extracellular vesicles | [53,55] |
| MiR-214 | Up | Liver fibrogenesis Induce oxidative stress | Gluthatione reductase | Human Mice Rat Bel7402 BRL | Hepatocyte | [40,57,58] |
| MiR-182 | Up | Inflammation Apoptosis | Mcp-1 Ccl20 Cxcl5 Cxcl1 Bcl2 | Mice Human | Liver | [40,57] |
| MiR-155 | Up | Promote liver steatosis Liver injury Inflammation fibrosis | Snail1 Smad2 STAT3 PPARα TLR inhibitor PPARγ TNFα | Human Mice Raw 264.7 Hepa 1-6 | Kupffer cells Hepatocyte Hepatic stellate cells | [46,47,48,49,50] |
| MiR-34a | Up | Fibrosis Cellular senescence Mallory–Denk cells formation | Smad3 SIRT1 | Human Mice | Kuppfer cells Hepatocyte Hepatic stellate cells Mallory–Denk cells | [79,80,81,82,83] |
| MiR-21 | Up | Regulates hepatic cell survival, transformation, and remodeling Liver regeneration | VHL Fas ligand (TNF superfamily, member 6) (FASLG) and death receptor 5 (DR5) | Rat Human Mice | Hepatic stellate cells Kuppfer cells Hepatocyte | [40,84,85,86] |
| Let-7 | Up | Fibrosis Inflammatory | Lin28 TLR7 | Mice Human | Hepatic stellate cells | [92,93] |
| Hepatocellular carcinoma | ||||||
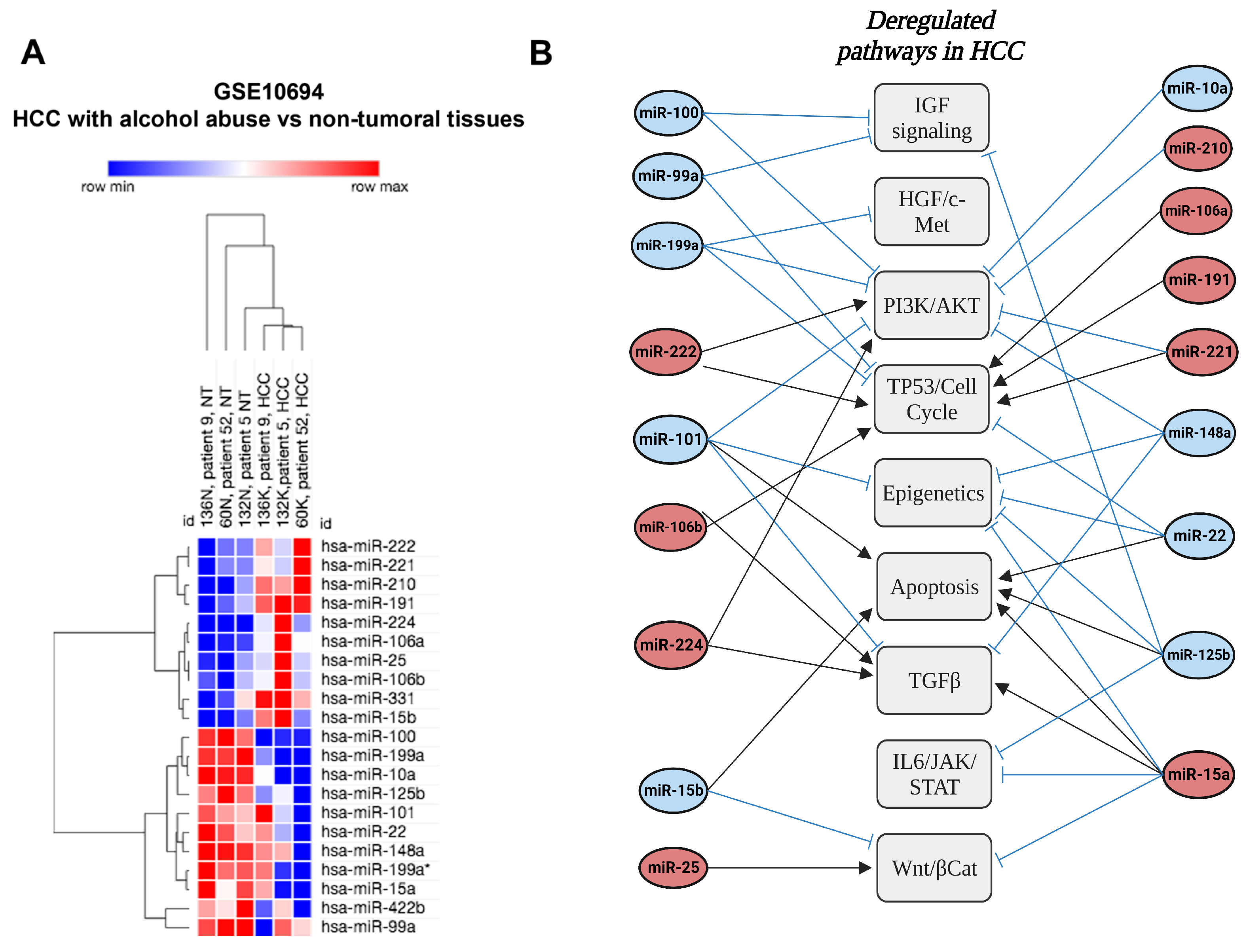
| MiR-100 | Down | Deregulated pathways in HCC | IGF signaling | Human | Liver | [41] |
| MiR-101 | Down | Deregulated pathways in HCC | Epigenetics TGFβ PI3K/AKT TP53/Cell cycle | Human | Liver | [41] |
| MiR-10a | Down | Deregulated pathways in HCC | MAPK Wnt/βCat | Human | Liver | [41] |
| MiR-125b | Down | Deregulated pathways in HCC | TP53/Cell cycle IL6/JAK/STAT IGF signaling | Human | Liver | [41] |
| MiR-15a | Down | Deregulated pathways in HCC | TGFβ | Human | Liver | [41] |
| MiR-199a | Down | Deregulated pathways in HCC | TGFβ | Human | Liver | [41] |
| MiR-422b | Down | Deregulated pathways in HCC | Human | Liver | [41] | |
| MiR-99a | Down | Deregulated pathways in HCC | IGF signaling | Human | Liver | [41] |
| MiR-139-5p | Down | Deregulated pathways in HCC | Human | Liver | [41] | |
| MiR-106a | Up | Deregulated pathways in HCC | Epigenetics | Human | Liver | [41] |
| MiR-106b | Up | Deregulated pathways in HCC | Epigenetics TGFβ | Human | Liver | [41] |
| MiR-15b | Up | Deregulated pathways in HCC | TP53/Cell cycle | Human | Liver | [41] |
| MiR-191 | Up | Deregulated pathways in HCC | Wnt/βCat NFκB TP53/Cell cycle | Human | Liver | [41] |
| MiR-210 | Up | Deregulated pathways in HCC | n.a | Human | Liver | [41] |
| MiR-221 | Up | Deregulated pathways in HCC | PI3K/AKT TP53/Cell cycle | Human | Liver | [41] |
| MiR-222 | Up | Deregulated pathways in HCC | Wnt/βCat PI3K/AKT | Human | Liver | [41] |
| MiR-224 | Up | Deregulated pathways in HCC | PI3K/AKT TP53/Cell cycle IL6/JAK/STAT | Human | Liver | [41] |
| MiR-25 | Up | Deregulated pathways in HCC | Wnt/βCat | Human | Liver | [41] |
| MiR-331 | Up | Deregulated pathways in HCC | n.a | Human | Liver | [41] |
| MiR-532-3p | Up | Promotes HCC cells migration, invasion, and proliferation | Protein tyrosine phosphatase receptor type T (PTPRT) | HCC specimens Hep3B HepG2 SMMC-7721 Huh7 MHCC-97 H | Hepatocyte | [105] |
| MiR-532-5p | Down | Promotes cell proliferation and metastasis | Chemokine (C-X-C motif) ligand 2 (CXCL2), X-ray Repair Cross Complementing 5 (XRCC5) | HEL7702 HEL7404 HCCLM3 SMMC7721 HepG2 PG5 MHCC97H Huh7 | Hepatocyte | [106,107] |
| MiR-22-3p | Up | Promotes HCC cells’ stemness and metastasis | Ten-eleven-translocation 2 (TET2) | HCC specimens Xenograft on BALB/C nude mice HCCLM3 | Cancer stem cells | [108] |
| MiR-126 | Down | Suppresses cell proliferation, invasion and migration | Epithelial Growth Factor Receptor (EGFR) | HCC specimens Hep3B MHCC97H Huh7 HCCLM3 | Hepatocyte Cancer stem cells | [109] |
| MiR-26a | Down | n.a | n.a | HCC specimens | n.a | [110] |
| MiR-22 | Down | Steatosis Tumor suppressor Deregulated pathways in HCC | FGFR1 FGF21 IL6/JAK/STAT | Human Mice | Hepatocyte | [39,40,41] |
| MiR-122 | Down | Protection against steatosis fibrosis | HiF1α TNFrsf13C | Human Mice RAW 264.7 Huh7 | Hepatocyte Kupffer cells Extracellular vesicles | [51,52,53,54] |
| MiR-21 | Up | Regulate hepatic cell survival, transformation, and remodel liver regeneration | VHL Fas ligand (TNF superfamily, member 6) (FASLG) and death receptor 5 (DR5) | Rat Human Mice | Hepatic stellate cells Kuppfer cells Hepatocyte | [40,84,85,86] |
| MiR-148a | Down | Anti-inflammatory Deregulated pathways in HCC | TXNIP Epigenetics TGFβ PI3K/AKT | Human Mice | Hepatocyte | [41,100] |
3. MicroRNAs in Alcohol-Induced Steatosis
3.1. Ethanol Metabolism
3.2. FGF21 and AMPKα Signaling
3.3. PPARα/γ Signaling
3.4. SREBP Signaling
3.5. Lipolysis in the Adipose Tissue
3.6. Alcohol-Related Steatosis as a Priming Event for Hepatocarcinogenesis?
4. MicroRNAs in Alcoholic Steatohepatitis (ASH)
4.1. Altered Gut–Liver Axis Toll-like Receptor Signaling
4.2. PPARα/γ Signaling
4.3. NFκB Signaling
4.4. Il-6/STAT3 Signaling
4.5. Oxidative Stress
4.6. Other Pathways
4.7. ASH as a Priming Event of Hepatocarcinogenesis
5. MicroRNAs in Alcohol-Associated Cirrhosis
5.1. HSCs Activation
5.2. Hepatocyte Proliferation
5.3. Other miRNAs with Poorly Characterized Functions
5.4. MiRNAs Fostering HCC Development
6. MicroRNAs in Alcoholic Hepatitis (AH)
6.1. Hippo/Yes-Associated Protein (YAP) Pathway Ductular Reaction
6.2. TLR and NFκB Signaling
6.3. Circulating microRNAs
6.4. Other miRNAs
7. MicroRNAs in Alcohol-Related Hepatocellular Carcinoma (HCC)
7.1. miRNAs with Oncogenic/Tumor Suppressive Functions in ALD-Related HCC
7.2. Other miRNAs with Poorly Defined Functions in ALD-Related HCC
| MiRs | Pathways | Model | Function | Target | Ref |
|---|---|---|---|---|---|
| MiR-100 | PI3K/AKT/mTOR IGF signaling | HCC cells from patients, Human HCC cell lines (SK-Hep1, MHCC97-L, SMMC-7721, HCCLM3, Huh7, Hep3B, and HepG2), | Tumor growth inhibition Apoptosis promotion Autophagy induction | Insulin-like growth factor 2 (IGF2), mammalian target of rapamycin (mTOR), and insulin like growth factor 1 receptor (IGF-1R) | [189,190] |
| MiR-101 | PI3K/AKT/mTOR, TGFβ, Epigenetics | HBV-related HCC tissue from patients, immortalized liver cell line L-02, and human HCC cell lines (HepG2, Hep3B, SMMC-7721, Huh7, MHCC-LM9) | Autophagy inhibition, Invasion and EMT inhibition, proapoptotic function, prevention of HCC progression | mTOR, EZH2, H3K27me3, EED, myeloid leukemia cell differentiation protein (Mcl-1), DNA methyltransferase 3A (DNMT3A), TGFβR1, Smad2 | [209,228,230,245,246,247,248,249] |
| MiR-106a | TP53/Cell cycle | Human HCC cell lines (HepG2 and Hep3B) | Apoptosis resistance, cell cycle progression and invasion | Tumor Protein P53 Inducible Nuclear Protein 1 (TP53INP1) and cyclin dependent kinase inhibitor 1A (CDKN1A) | [250] |
| MiR-106b | TP53/Cell cycle, TGFβ signaling | Tissue from patients, Human HCC cell lines (Hep3B, Huh7, HepG2, and Bel-7402) | Promote HCC cell proliferation and migration | Disabled homolog 2 (DAB2), SMAD Family Member 7 (SMAD7) | [207,209] |
| MiR-10a | PI3K/AKT/mTOR | HCC patients, human HCC cell lines (Huh7, HepG2, and PLC) | Cell proliferation inhibition chemosensibility | Musashi 1 (MSI1) | [251] |
| MiR-125b | IL6/JAK/STAT, IGF signaling, Apoptosis, epigenetics | Human HCC cell lines (MHCC97L, SMMC7721, HepG2, HL-7702), HCC tissue from patients | Promote apoptosis, induce cell senescence and invasion inhibition | IGF2, Mcl-1, Bcl-w, Interleukin (IL)-6, IL-6R, sirtuin 6 (SIRT6) and SIRT7 | [189,252,253,254] |
| MiR-148a | Epigenetics, PI3K/AKT, TGFβ signaling | Human HCC cell lines (MHCC97, Huh7, HepG2, SMMC-7721, and HCCLM3), normal liver cell line L02 | Cell proliferation inhibition Cell migration and invasion inhibition | DNA methyltransferase DNMT1, Death receptor-5 (DR-5), SMAD2 | [240,241,255] |
| MiR-15a | WNT/β-catenin, TGF-β signaling, epigenetics, JAK/STAT | HCC tissue from patients, Human HCC cell lines (HCC-LM3, Huh-7, CSQT2, HepG2, MHCC97H, and SMMC-7721), normal liver cell line THLE2, Tumor xenograft | Inhibition of HCC proliferation, migration and invasion. Promote apoptosis | O-linked N-acetylglucosamine (GlcNAc) transferase (OGT), Transforming Growth Factor Beta 1 (TGF-β1), SMAD7, WNT3A, signal transducer and activator of transcription 3 (STAT3) | [256,257,258,259,260] |
| MiR-15b | Apoptosis, WNT/β-catenin | HCC patients, Human HCC cell lines (HepG2, Huh7, Hep3B, MHCC-97L and MHCC-97H) | Cell proliferation inhibition Promote apoptosis | WNT3A, B-cell lymphoma 2 (BCL-2) | [261] |
| MiR-191 | TP53/Cell cycle | HCC tissue from patients, Hep3B and HepG2 cell lines | Cell cycle progression and cell proliferation | ZO-1-associated Y-box factor (ZONAB)/cyclinD1 | [262] |
| MiR-199a | HGF/c-Met, TP53/Cell cycle, PI3K/AKT/mTOR | Human HCC cell lines (Huh7, HepG2, SNU182, PLC/PRF/5, Hep3B, SNU423, and SNU449) | Inhibition of cell proliferation, cell cycle arrest, apoptosis induction | CD44, mTOR, c-Met, zinc-fingers and homeoboxes-1 (ZHX1) | [263,264,265] |
| MiR-210 | PI3K/AKT | Human HCC cell lines (HepG2, MHCC-97H and HuH7) | Promote proliferation and invasion Inhibition of apoptosis | PI3K, AKT, mTOR | [223] |
| MiR-22 | Epigenetics, TP53/Cell Cycle | HCC tissue from patients, Human HCC cell line PLC/PRF/5 and MHCC97L | Induction of apoptosis Cell proliferation inhibition | X-linked IAP (XIAP), Histone deacetylase 4 (HDAC4), Cyclin-dependent kinase inhibitor 1A (CDKN1A) | [266,267,268] |
| MiR-221 | PI3K/AKT/mTOR TP53/Cell cycle | HCC patients, HCC cell lines (PLC/PRF/5, Huh7, HepG2, SNU-449, SNU398, SNU-423 and SK-Hep-1) | Cell proliferation Cell cycle progression | CD44, CDKN1B/p27, CDKN1C/p57 DNA damage-inducible transcript 4 (DDIT4) | [228,229,230] |
| MiR-222 | PI3K/AKT/mTOR, TP53/Cell Cycle | Human HCC cell lines (HepG2, Hep3B, HKCI-4, and HKCI-9) | Cell proliferation, Migration, and invasion and inhibits apoptosis | p27 protein phosphatase 2A subunit B (PPP2R2A) | [269,270] |
| MiR-224 | PI3K/AKT/mTOR, TGFβ signaling | HCC tissue from patients, Human HCC cell lines (HepG2) | Cell proliferation | Protein Phosphatase 2 Scaffold Subunit Abeta (PPP2R1B), SMAD4 | [271,272,273] |
| MiR-25 | WNT/β-catenin | Human HCC cell lines (HCCLM3 and Huh7) | cell proliferation, migration and invasion | PTEN | [274] |
| MiR-99a | IGF signaling, TP53/cell cycle Epigenetics | HCC tissue from patients, Human HCC cell lines (Hep2G SMMC-7721, Huh7, and Hep3B) | Cell proliferation and invasion inhibition, block cell cycle | IGF1R mTOR AGO2 | [275,276,277] |
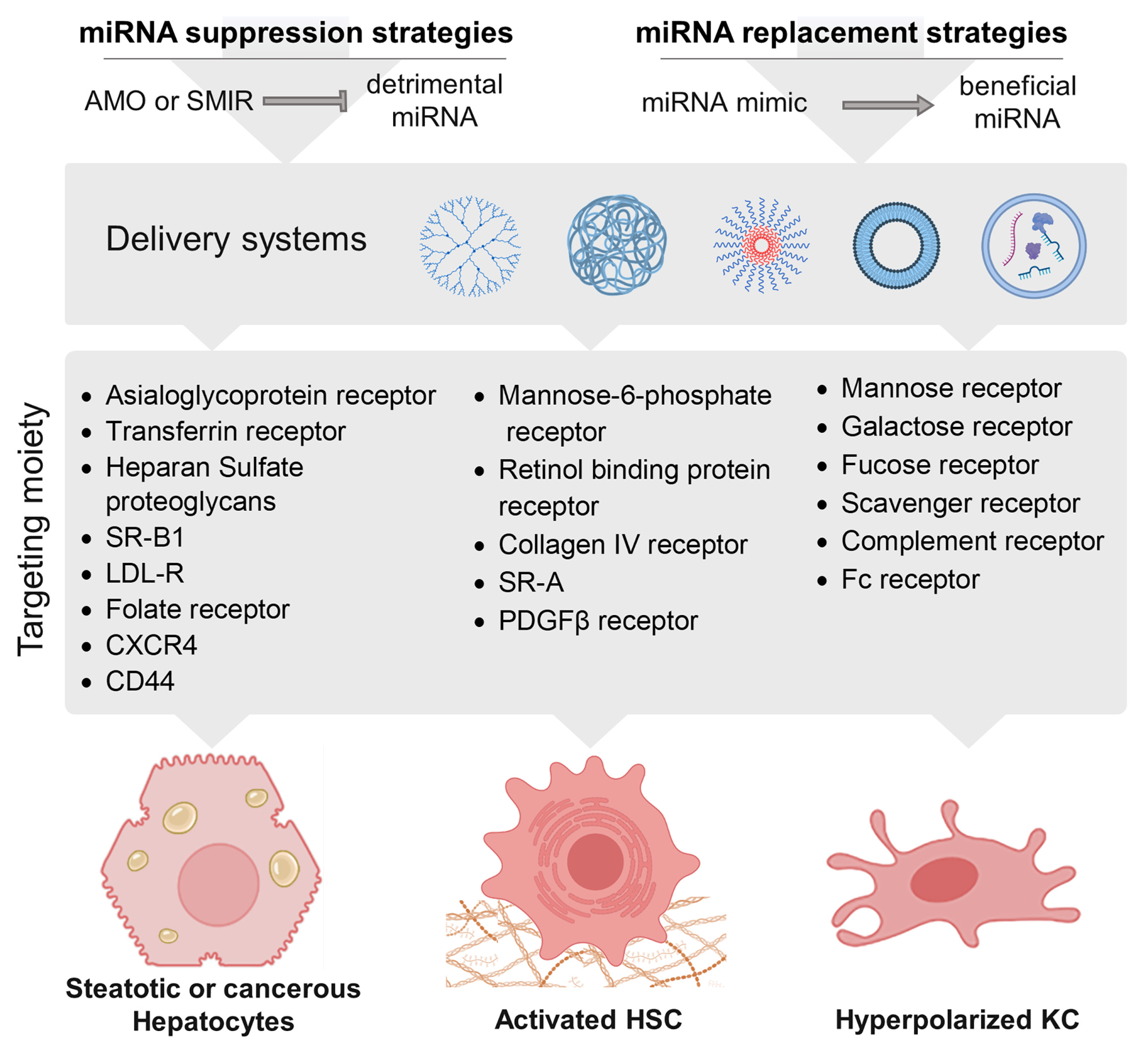
8. Therapeutical Strategies against ALD/HCC-Related miRNAs
8.1. A Myriad of Strategies to Target miRNAs
8.2. Therapeutic targeting of miRNAs in ALD
8.2.1. Steatosis
8.2.2. ASH
8.2.3. Cirrhosis
8.2.4. HCC
8.3. Therapeutic Approaches Targeting miRNAs in Clinical Trials and Future Perspectives
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Malnick, S.D.H.; Alin, P.; Somin, M.; Neuman, M.G. Fatty Liver Disease-Alcoholic and Non-Alcoholic: Similar but Different. Int. J. Mol. Sci. 2022, 23, 16226. [Google Scholar] [CrossRef] [PubMed]
- Devarbhavi, H.; Asrani, S.K.; Arab, J.P.; Nartey, Y.A.; Pose, E.; Kamath, P.S. Global Burden of Liver Disease: 2023 Update. J. Hepatol. 2023, 79, 516–537. [Google Scholar] [CrossRef] [PubMed]
- Macpherson, I.; Abeysekera, K.W.M.; Harris, R.; Mansour, D.; McPherson, S.; Rowe, I.; Rosenberg, W.; Dillon, J.F.; Yeoman, A. Identification of Liver Disease: Why and How. Frontline Gastroenterol. 2022, 13, 367–373. [Google Scholar] [CrossRef]
- Nassir, F.; Rector, R.S.; Hammoud, G.M.; Ibdah, J.A. Pathogenesis and Prevention of Hepatic Steatosis. Gastroenterol. Hepatol. 2015, 11, 167–175. [Google Scholar]
- Idilman, I.S.; Ozdeniz, I.; Karcaaltincaba, M. Hepatic Steatosis: Etiology, Patterns, and Quantification. Semin. Ultrasound CT MRI 2016, 37, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Feldstein, A.E.; Gores, G.J. Apoptosis in Alcoholic and Nonalcoholic Steatohepatitis. Front. Biosci. J. Virtual Libr. 2005, 10, 3093–3099. [Google Scholar] [CrossRef]
- Allameh, A.; Niayesh-Mehr, R.; Aliarab, A.; Sebastiani, G.; Pantopoulos, K. Oxidative Stress in Liver Pathophysiology and Disease. Antioxid. Basel Switz. 2023, 12, 1653. [Google Scholar] [CrossRef]
- Mehal, W.; Imaeda, A. Cell Death and Fibrogenesis. Semin. Liver Dis. 2010, 30, 226–231. [Google Scholar] [CrossRef]
- Lackner, C.; Tiniakos, D. Fibrosis and Alcohol-Related Liver Disease. J. Hepatol. 2019, 70, 294–304. [Google Scholar] [CrossRef]
- Tarao, K.; Nozaki, A.; Ikeda, T.; Sato, A.; Komatsu, H.; Komatsu, T.; Taguri, M.; Tanaka, K. Real Impact of Liver Cirrhosis on the Development of Hepatocellular Carcinoma in Various Liver Diseases-Meta-Analytic Assessment. Cancer Med. 2019, 8, 1054–1065. [Google Scholar] [CrossRef]
- Philips, C.A.; Augustine, P.; Yerol, P.K.; Rajesh, S.; Mahadevan, P. Severe Alcoholic Hepatitis: Current Perspectives. Hepatic Med. Evid. Res. 2019, 11, 97–108. [Google Scholar] [CrossRef]
- Ganne-Carrié, N.; Nahon, P. Hepatocellular Carcinoma in the Setting of Alcohol-Related Liver Disease. J. Hepatol. 2019, 70, 284–293. [Google Scholar] [CrossRef] [PubMed]
- Bosch, F.X.; Ribes, J.; Díaz, M.; Cléries, R. Primary Liver Cancer: Worldwide Incidence and Trends. Gastroenterology 2004, 127, S5–S16. [Google Scholar] [CrossRef] [PubMed]
- Bruix, J.; Gores, G.J.; Mazzaferro, V. Hepatocellular Carcinoma: Clinical Frontiers and Perspectives. Gut 2014, 63, 844–855. [Google Scholar] [CrossRef] [PubMed]
- Pimpin, L.; Cortez-Pinto, H.; Negro, F.; Corbould, E.; Lazarus, J.V.; Webber, L.; Sheron, N. EASL HEPAHEALTH Steering Committee Burden of Liver Disease in Europe: Epidemiology and Analysis of Risk Factors to Identify Prevention Policies. J. Hepatol. 2018, 69, 718–735. [Google Scholar] [CrossRef] [PubMed]
- Morgan, T.R.; Mandayam, S.; Jamal, M.M. Alcohol and Hepatocellular Carcinoma. Gastroenterology 2004, 127, S87–S96. [Google Scholar] [CrossRef] [PubMed]
- Loomba, R.; Yang, H.-I.; Su, J.; Brenner, D.; Barrett-Connor, E.; Iloeje, U.; Chen, C.-J. Synergism between Obesity and Alcohol in Increasing the Risk of Hepatocellular Carcinoma: A Prospective Cohort Study. Am. J. Epidemiol. 2013, 177, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.-M.; Govindarajan, S.; Arakawa, K.; Yu, M.C. Synergism of Alcohol, Diabetes, and Viral Hepatitis on the Risk of Hepatocellular Carcinoma in Blacks and Whites in the U.S. Cancer 2004, 101, 1009–1017. [Google Scholar] [CrossRef] [PubMed]
- Amonker, S.; Houshmand, A.; Hinkson, A.; Rowe, I.; Parker, R. Prevalence of Alcohol-Associated Liver Disease: A Systematic Review and Meta-Analysis. Hepatol. Commun. 2023, 7, e0133. [Google Scholar] [CrossRef]
- Lu, H. Narrative Review: Glucocorticoids in Alcoholic Hepatitis-Benefits, Side Effects, and Mechanisms. J. Xenobiotics 2022, 12, 266–288. [Google Scholar] [CrossRef]
- Foncea, C.G.; Sporea, I.; Lupușoru, R.; Moga, T.V.; Bende, F.; Șirli, R.; Popescu, A. Day-4 Lille Score Is a Good Prognostic Factor and Early Predictor in Assessing Therapy Response in Patients with Liver Cirrhosis and Severe Alcoholic Hepatitis. J. Clin. Med. 2021, 10, 2338. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, N.; Shor, J.; Szabo, G. Alcoholic Hepatitis: A Review. Alcohol Alcohol. Oxf. Oxfs. 2019, 54, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Bataller, R.; Brenner, D.A. Liver Fibrosis. J. Clin. Investig. 2005, 115, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Montemurro, N.; Ricciardi, L.; Scerrati, A.; Ippolito, G.; Lofrese, G.; Trungu, S.; Stoccoro, A. The Potential Role of Dysregulated MiRNAs in Adolescent Idiopathic Scoliosis and 22q11.2 Deletion Syndrome. J. Pers. Med. 2022, 12, 1925. [Google Scholar] [CrossRef] [PubMed]
- Chimenti, C.; Magnocavallo, M.; Vetta, G.; Alfarano, M.; Manguso, G.; Ajmone, F.; Ballatore, F.; Costantino, J.; Ciaramella, P.; Severino, P.; et al. The Role of MicroRNA in the Myocarditis: A Small Actor for a Great Role. Curr. Cardiol. Rep. 2023, 25, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Dutta, A. MicroRNAs in Cancer. Annu. Rev. Pathol. 2009, 4, 199–227. [Google Scholar] [CrossRef] [PubMed]
- Pekarek, L.; Torres-Carranza, D.; Fraile-Martinez, O.; García-Montero, C.; Pekarek, T.; Saez, M.A.; Rueda-Correa, F.; Pimentel-Martinez, C.; Guijarro, L.G.; Diaz-Pedrero, R.; et al. An Overview of the Role of MicroRNAs on Carcinogenesis: A Focus on Cell Cycle, Angiogenesis and Metastasis. Int. J. Mol. Sci. 2023, 24, 7268. [Google Scholar] [CrossRef]
- Fang, Z.; Dou, G.; Wang, L. MicroRNAs in the Pathogenesis of Nonalcoholic Fatty Liver Disease. Int. J. Biol. Sci. 2021, 17, 1851–1863. [Google Scholar] [CrossRef]
- Hochreuter, M.Y.; Dall, M.; Treebak, J.T.; Barrès, R. MicroRNAs in Non-Alcoholic Fatty Liver Disease: Progress and Perspectives. Mol. Metab. 2022, 65, 101581. [Google Scholar] [CrossRef]
- Valinezhad Orang, A.; Safaralizadeh, R.; Kazemzadeh-Bavili, M. Mechanisms of MiRNA-Mediated Gene Regulation from Common Downregulation to MRNA-Specific Upregulation. Int. J. Genom. 2014, 2014, 970607. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. Elegans Heterochronic Gene Lin-4 Encodes Small RNAs with Antisense Complementarity to Lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- An, X.; Sarmiento, C.; Tan, T.; Zhu, H. Regulation of Multidrug Resistance by MicroRNAs in Anti-Cancer Therapy. Acta Pharm. Sin. B 2017, 7, 38–51. [Google Scholar] [CrossRef] [PubMed]
- Sobolewski, C.; Dubuquoy, L.; Legrand, N. MicroRNAs, Tristetraprolin Family Members and HuR: A Complex Interplay Controlling Cancer-Related Processes. Cancers 2022, 14, 3516. [Google Scholar] [CrossRef] [PubMed]
- Wilczynska, A.; Bushell, M. The Complexity of MiRNA-Mediated Repression. Cell Death Differ. 2015, 22, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, K.; Pimienta, M.; Seki, E. Alcoholic Liver Disease: A Current Molecular and Clinical Perspective. Liver Res. 2018, 2, 161–172. [Google Scholar] [CrossRef]
- Cheng, X.-Y.; Liu, J.-D.; Lu, X.-Y.; Yan, X.; Huang, C.; Meng, X.-M.; Li, J. MiR-203 Inhibits Alcohol-Induced Hepatic Steatosis by Targeting Lipin1. Front. Pharmacol. 2018, 9, 275. [Google Scholar] [CrossRef]
- Niture, S.; Gadi, S.; Qi, Q.; Gyamfi, M.A.; Varghese, R.S.; Rios-Colon, L.; Chimeh, U.; Ressom, H.W.; Kumar, D. MicroRNA-483-5p Inhibits Hepatocellular Carcinoma Cell Proliferation, Cell Steatosis, and Fibrosis by Targeting PPARα and TIMP2. Cancers 2023, 15, 1715. [Google Scholar] [CrossRef]
- Hu, Y.; Liu, H.-X.; Jena, P.K.; Sheng, L.; Ali, M.R.; Wan, Y.-J.Y. MiR-22 Inhibition Reduces Hepatic Steatosis via FGF21 and FGFR1 Induction. JHEP Rep. Innov. Hepatol. 2020, 2, 100093. [Google Scholar] [CrossRef]
- Blaya, D.; Coll, M.; Rodrigo-Torres, D.; Vila-Casadesús, M.; Altamirano, J.; Llopis, M.; Graupera, I.; Perea, L.; Aguilar-Bravo, B.; Díaz, A.; et al. Integrative MicroRNA Profiling in Alcoholic Hepatitis Reveals a Role for MicroRNA-182 in Liver Injury and Inflammation. Gut 2016, 65, 1535–1545. [Google Scholar] [CrossRef]
- Li, W.; Xie, L.; He, X.; Li, J.; Tu, K.; Wei, L.; Wu, J.; Guo, Y.; Ma, X.; Zhang, P.; et al. Diagnostic and Prognostic Implications of MicroRNAs in Human Hepatocellular Carcinoma. Int. J. Cancer 2008, 123, 1616–1622. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-Z.; Lu, J.; Li, Y.-Y.; Zhong, Y.-J.; Yang, C.-F.; Zhang, Y.; Huang, L.-H.; Huang, S.-M.; Li, Q.-R.; Wu, D.; et al. MicroRNA-378b Regulates Ethanol-Induced Hepatic Steatosis by Targeting CaMKK2 to Mediate Lipid Metabolism. Bioengineered 2021, 12, 12659–12676. [Google Scholar] [CrossRef] [PubMed]
- Mostofa, M.G.; Tran, M.; Gilling, S.; Lee, G.; Fraher, O.; Jin, L.; Kang, H.; Park, Y.-K.; Lee, J.-Y.; Wang, L.; et al. MicroRNA-200c Coordinates HNF1 Homeobox B and Apolipoprotein O Functions to Modulate Lipid Homeostasis in Alcoholic Fatty Liver Disease. J. Biol. Chem. 2022, 298, 101966. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Hu, M.; Zhang, R.; Shen, Z.; Flatow, L.; You, M. MicroRNA-217 Promotes Ethanol-Induced Fat Accumulation in Hepatocytes by down-Regulating SIRT1. J. Biol. Chem. 2012, 287, 9817–9826. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Liang, X.; Jogasuria, A.; Davidson, N.O.; You, M. MiR-217 Regulates Ethanol-Induced Hepatic Inflammation by Disrupting Sirtuin 1-Lipin-1 Signaling. Am. J. Pathol. 2015, 185, 1286–1296. [Google Scholar] [CrossRef] [PubMed]
- Bala, S.; Marcos, M.; Kodys, K.; Csak, T.; Catalano, D.; Mandrekar, P.; Szabo, G. Up-Regulation of MicroRNA-155 in Macrophages Contributes to Increased Tumor Necrosis Factor {alpha} (TNF{alpha}) Production via Increased MRNA Half-Life in Alcoholic Liver Disease. J. Biol. Chem. 2011, 286, 1436–1444. [Google Scholar] [CrossRef]
- Hartmann, P.; Tacke, F. Tiny RNA with Great Effects: MiR-155 in Alcoholic Liver Disease. J. Hepatol. 2016, 64, 1214–1216. [Google Scholar] [CrossRef]
- Bala, S.; Csak, T.; Saha, B.; Zatsiorsky, J.; Kodys, K.; Catalano, D.; Satishchandran, A.; Szabo, G. The Pro-Inflammatory Effects of MiR-155 Promote Liver Fibrosis and Alcohol-Induced Steatohepatitis. J. Hepatol. 2016, 64, 1378–1387. [Google Scholar] [CrossRef]
- Mandrekar, P.; Ambade, A.; Lim, A.; Szabo, G.; Catalano, D. An Essential Role for Monocyte Chemoattractant Protein-1 in Alcoholic Liver Injury: Regulation of Proinflammatory Cytokines and Hepatic Steatosis in Mice. Hepatol. Baltim. Md 2011, 54, 2185–2197. [Google Scholar] [CrossRef]
- Rachakonda, V.; Bataller, R.; Duarte-Rojo, A. Recent Advances in Alcoholic Hepatitis. F1000Research 2020, 9, 97. [Google Scholar] [CrossRef]
- Lu, X.; Liu, Y.; Xuan, W.; Ye, J.; Yao, H.; Huang, C.; Li, J. Circ_1639 Induces Cells Inflammation Responses by Sponging MiR-122 and Regulating TNFRSF13C Expression in Alcoholic Liver Disease. Toxicol. Lett. 2019, 314, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Satishchandran, A.; Ambade, A.; Rao, S.; Hsueh, Y.-C.; Iracheta-Vellve, A.; Tornai, D.; Lowe, P.; Gyongyosi, B.; Li, J.; Catalano, D.; et al. MicroRNA 122, Regulated by GRLH2, Protects Livers of Mice and Patients From Ethanol-Induced Liver Disease. Gastroenterology 2018, 154, 238–252.e7. [Google Scholar] [CrossRef] [PubMed]
- Momen-Heravi, F.; Saha, B.; Kodys, K.; Catalano, D.; Satishchandran, A.; Szabo, G. Increased Number of Circulating Exosomes and Their MicroRNA Cargos Are Potential Novel Biomarkers in Alcoholic Hepatitis. J. Transl. Med. 2015, 13, 261. [Google Scholar] [CrossRef] [PubMed]
- Ambade, A.; Satishchandran, A.; Szabo, G. Alcoholic Hepatitis Accelerates Early Hepatobiliary Cancer by Increasing Stemness and MiR-122-Mediated HIF-1α Activation. Sci. Rep. 2016, 6, 21340. [Google Scholar] [CrossRef]
- Bala, S.; Babuta, M.; Catalano, D.; Saiju, A.; Szabo, G. Alcohol Promotes Exosome Biogenesis and Release via Modulating Rabs and MiR-192 Expression in Human Hepatocytes. Front. Cell Dev. Biol. 2021, 9, 787356. [Google Scholar] [CrossRef]
- Thulasingam, S.; Massilamany, C.; Gangaplara, A.; Dai, H.; Yarbaeva, S.; Subramaniam, S.; Riethoven, J.-J.; Eudy, J.; Lou, M.; Reddy, J. MiR-27b*, an Oxidative Stress-Responsive MicroRNA Modulates Nuclear Factor-KB Pathway in RAW 264.7 Cells. Mol. Cell. Biochem. 2011, 352, 181–188. [Google Scholar] [CrossRef]
- Dolganiuc, A.; Petrasek, J.; Kodys, K.; Catalano, D.; Mandrekar, P.; Velayudham, A.; Szabo, G. MicroRNA Expression Profile in Lieber-DeCarli Diet-Induced Alcoholic and Methionine Choline Deficient Diet-Induced Nonalcoholic Steatohepatitis Models in Mice. Alcohol. Clin. Exp. Res. 2009, 33, 1704–1710. [Google Scholar] [CrossRef]
- Dong, X.; Liu, H.; Chen, F.; Li, D.; Zhao, Y. MiR-214 Promotes the Alcohol-Induced Oxidative Stress via down-Regulation of Glutathione Reductase and Cytochrome P450 Oxidoreductase in Liver Cells. Alcohol. Clin. Exp. Res. 2014, 38, 68–77. [Google Scholar] [CrossRef]
- Yu, F.; Zheng, Y.; Hong, W.; Chen, B.; Dong, P.; Zheng, J. MicroRNA-200a Suppresses Epithelial-to-mesenchymal Transition in Rat Hepatic Stellate Cells via GLI Family Zinc Finger 2. Mol. Med. Rep. 2015, 12, 8121–8128. [Google Scholar] [CrossRef]
- Katoh, Y.; Katoh, M. Hedgehog Signaling, Epithelial-to-Mesenchymal Transition and MiRNA (Review). Int. J. Mol. Med. 2008, 22, 271–275. [Google Scholar] [CrossRef]
- Saikia, P.; Bellos, D.; McMullen, M.R.; Pollard, K.A.; de la Motte, C.; Nagy, L.E. MicroRNA 181b-3p and Its Target Importin A5 Regulate Toll-like Receptor 4 Signaling in Kupffer Cells and Liver Injury in Mice in Response to Ethanol. Hepatol. Baltim. Md 2017, 66, 602–615. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Gao, B. A Small Specific-Sized Hyaluronic Acid Ameliorates Alcoholic Liver Disease by Targeting a Small RNA: New Hope for Therapy? Hepatol. Baltim. Md 2017, 66, 321–323. [Google Scholar] [CrossRef]
- Fu, R.; Zhou, J.; Wang, R.; Sun, R.; Feng, D.; Wang, Z.; Zhao, Y.; Lv, L.; Tian, X.; Yao, J. Protocatechuic Acid-Mediated MiR-219a-5p Activation Inhibits the P66shc Oxidant Pathway to Alleviate Alcoholic Liver Injury. Oxid. Med. Cell. Longev. 2019, 2019, 3527809. [Google Scholar] [CrossRef] [PubMed]
- Yeligar, S.; Tsukamoto, H.; Kalra, V.K. Ethanol-Induced Expression of ET-1 and ET-BR in Liver Sinusoidal Endothelial Cells and Human Endothelial Cells Involves Hypoxia-Inducible Factor-1alpha and MicrorNA-199. J. Immunol. Baltim. Md 1950 2009, 183, 5232–5243. [Google Scholar] [CrossRef]
- Ye, J.; Lin, Y.; Yu, Y.; Sun, D. LncRNA NEAT1/MicroRNA-129-5p/SOCS2 Axis Regulates Liver Fibrosis in Alcoholic Steatohepatitis. J. Transl. Med. 2020, 18, 445. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Rani, R.; Karns, R.; Gandhi, C.R. Augmenter of Liver Regeneration Protein Deficiency Promotes Hepatic Steatosis by Inducing Oxidative Stress and MicroRNA-540 Expression. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2019, 33, 3825–3840. [Google Scholar] [CrossRef]
- Tang, Y.; Zhang, L.; Forsyth, C.B.; Shaikh, M.; Song, S.; Keshavarzian, A. The Role of MiR-212 and INOS in Alcohol-Induced Intestinal Barrier Dysfunction and Steatohepatitis. Alcohol. Clin. Exp. Res. 2015, 39, 1632–1641. [Google Scholar] [CrossRef]
- Tang, Y.; Banan, A.; Forsyth, C.B.; Fields, J.Z.; Lau, C.K.; Zhang, L.J.; Keshavarzian, A. Effect of Alcohol on MiR-212 Expression in Intestinal Epithelial Cells and Its Potential Role in Alcoholic Liver Disease. Alcohol. Clin. Exp. Res. 2008, 32, 355–364. [Google Scholar] [CrossRef]
- Ye, D.; Zhang, T.; Lou, G.; Liu, Y. Role of MiR-223 in the Pathophysiology of Liver Diseases. Exp. Mol. Med. 2018, 50, 1–12. [Google Scholar] [CrossRef]
- Gu, J.; Xu, H.; Chen, Y.; Li, N.; Hou, X. MiR-223 as a Regulator and Therapeutic Target in Liver Diseases. Front. Immunol. 2022, 13, 860661. [Google Scholar] [CrossRef]
- Li, M.; He, Y.; Zhou, Z.; Ramirez, T.; Gao, Y.; Gao, Y.; Ross, R.A.; Cao, H.; Cai, Y.; Xu, M.; et al. MicroRNA-223 Ameliorates Alcoholic Liver Injury by Inhibiting the IL-6-P47phox-Oxidative Stress Pathway in Neutrophils. Gut 2017, 66, 705–715. [Google Scholar] [CrossRef] [PubMed]
- Slevin, E.; Baiocchi, L.; Wu, N.; Ekser, B.; Sato, K.; Lin, E.; Ceci, L.; Chen, L.; Lorenzo, S.R.; Xu, W.; et al. Kupffer Cells: Inflammation Pathways and Cell-Cell Interactions in Alcohol-Associated Liver Disease. Am. J. Pathol. 2020, 190, 2185–2193. [Google Scholar] [CrossRef] [PubMed]
- Bala, S.; Szabo, G. MicroRNA Signature in Alcoholic Liver Disease. Int. J. Hepatol. 2012, 2012, 498232. [Google Scholar] [CrossRef] [PubMed]
- Momen-Heravi, F.; Catalano, D.; Talis, A.; Szabo, G.; Bala, S. Protective Effect of LNA-Anti-MiR-132 Therapy on Liver Fibrosis in Mice. Mol. Ther. Nucleic Acids 2021, 25, 155–167. [Google Scholar] [CrossRef]
- Wang, W.; Zhong, G.-Z.; Long, K.-B.; Liu, Y.; Liu, Y.-Q.; Xu, A.-L. Silencing MiR-181b-5p Upregulates PIAS1 to Repress Oxidative Stress and Inflammatory Response in Rats with Alcoholic Fatty Liver Disease through Inhibiting PRMT1. Int. Immunopharmacol. 2021, 101, 108151. [Google Scholar] [CrossRef] [PubMed]
- Saha, B.; Bruneau, J.C.; Kodys, K.; Szabo, G. Alcohol-Induced MiR-27a Regulates Differentiation and M2 Macrophage Polarization of Normal Human Monocytes. J. Immunol. Baltim. Md 1950 2015, 194, 3079–3087. [Google Scholar] [CrossRef]
- Saha, B.; Momen-Heravi, F.; Kodys, K.; Szabo, G. MicroRNA Cargo of Extracellular Vesicles from Alcohol-Exposed Monocytes Signals Naive Monocytes to Differentiate into M2 Macrophages. J. Biol. Chem. 2016, 291, 149–159. [Google Scholar] [CrossRef]
- Eguchi, A.; Yan, R.; Pan, S.Q.; Wu, R.; Kim, J.; Chen, Y.; Ansong, C.; Smith, R.D.; Tempaku, M.; Ohno-Machado, L.; et al. Comprehensive Characterization of Hepatocyte-Derived Extracellular Vesicles Identifies Direct MiRNA-Based Regulation of Hepatic Stellate Cells and DAMP-Based Hepatic Macrophage IL-1β and IL-17 Upregulation in Alcoholic Hepatitis Mice. J. Mol. Med. Berl. Ger. 2020, 98, 1021–1034. [Google Scholar] [CrossRef]
- Wan, Y.; McDaniel, K.; Wu, N.; Ramos-Lorenzo, S.; Glaser, T.; Venter, J.; Francis, H.; Kennedy, L.; Sato, K.; Zhou, T.; et al. Regulation of Cellular Senescence by MiR-34a in Alcoholic Liver Injury. Am. J. Pathol. 2017, 187, 2788–2798. [Google Scholar] [CrossRef]
- Lee, J.; Padhye, A.; Sharma, A.; Song, G.; Miao, J.; Mo, Y.-Y.; Wang, L.; Kemper, J.K. A Pathway Involving Farnesoid X Receptor and Small Heterodimer Partner Positively Regulates Hepatic Sirtuin 1 Levels via MicroRNA-34a Inhibition. J. Biol. Chem. 2010, 285, 12604–12611. [Google Scholar] [CrossRef]
- Wan, Y.; Slevin, E.; Koyama, S.; Huang, C.-K.; Shetty, A.K.; Li, X.; Harrison, K.; Li, T.; Zhou, B.; Lorenzo, S.R.; et al. MiR-34a Regulates Macrophage-Associated Inflammation and Angiogenesis in Alcohol-Induced Liver Injury. Hepatol. Commun. 2023, 7, e0089. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; French, B.A.; Li, J.; Tillman, B.; French, S.W. Altered Regulation of MiR-34a and MiR-483-3p in Alcoholic Hepatitis and DDC Fed Mice. Exp. Mol. Pathol. 2015, 99, 552–557. [Google Scholar] [CrossRef] [PubMed]
- Iwagami, Y.; Zou, J.; Zhang, H.; Cao, K.; Ji, C.; Kim, M.; Huang, C. Alcohol-mediated MiR-34a Modulates Hepatocyte Growth and Apoptosis. J. Cell. Mol. Med. 2018, 22, 3987–3995. [Google Scholar] [CrossRef] [PubMed]
- Francis, H.; McDaniel, K.; Han, Y.; Liu, X.; Kennedy, L.; Yang, F.; McCarra, J.; Zhou, T.; Glaser, S.; Venter, J.; et al. Regulation of the Extrinsic Apoptotic Pathway by MicroRNA-21 in Alcoholic Liver Injury. J. Biol. Chem. 2014, 289, 27526–27539. [Google Scholar] [CrossRef] [PubMed]
- Dippold, R.P.; Vadigepalli, R.; Gonye, G.E.; Hoek, J.B. Chronic Ethanol Feeding Enhances MiR-21 Induction during Liver Regeneration While Inhibiting Proliferation in Rats. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G733–G743. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; McDaniel, K.; Zhou, T.; Ramos-Lorenzo, S.; Wu, C.; Huang, L.; Chen, D.; Annable, T.; Francis, H.; Glaser, S.; et al. Knockout of MicroRNA-21 Attenuates Alcoholic Hepatitis through the VHL/NF-ΚB Signaling Pathway in Hepatic Stellate Cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G385–G398. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, A.; Lazaro, R.G.; Wang, J.; Kim, J.; Povero, D.; Willliams, B.; Ho, S.B.; Stärkel, P.; Schnabl, B.; Ohno-Machado, L.; et al. Extracellular Vesicles Released by Hepatocytes from Gastric Infusion Model of Alcoholic Liver Disease Contain a MicroRNA Barcode That Can Be Detected in Blood. Hepatol. Baltim. Md 2017, 65, 475–490. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Wang, H.-F.; Chen, Y.-M.; Bai, R.-X.; Du, S.-Y. Baicalin Confers Hepatoprotective Effect against Alcohol-Associated Liver Disease by Upregulating MicroRNA-205. Int. Immunopharmacol. 2022, 107, 108553. [Google Scholar] [CrossRef]
- Zhou, K.; Yin, F.; Li, Y.; Ma, C.; Liu, P.; Xin, Z.; Ren, R.; Wei, S.; Khan, M.; Wang, H.; et al. MicroRNA-29b Ameliorates Hepatic Inflammation via Suppression of STAT3 in Alcohol-Associated Liver Disease. Alcohol Fayettev. N 2022, 99, 9–22. [Google Scholar] [CrossRef]
- Sabater, L.; Locatelli, L.; Oakley, F.; Hardy, T.; French, J.; Robinson, S.M.; Sen, G.; Mann, D.A.; Mann, J. RNA Sequencing Reveals Changes in the MicroRNAome of Transdifferentiating Hepatic Stellate Cells That Are Conserved between Human and Rat. Sci. Rep. 2020, 10, 21708. [Google Scholar] [CrossRef]
- Xiong, J.; Ni, J.; Chen, C.; Wang, K. MiR-148a-3p Regulates Alcoholic Liver Fibrosis through Targeting ERBB3. Int. J. Mol. Med. 2020, 46, 1003–1012. [Google Scholar] [CrossRef] [PubMed]
- McDaniel, K.; Huang, L.; Sato, K.; Wu, N.; Annable, T.; Zhou, T.; Ramos-Lorenzo, S.; Wan, Y.; Huang, Q.; Francis, H.; et al. The Let-7/Lin28 Axis Regulates Activation of Hepatic Stellate Cells in Alcoholic Liver Injury. J. Biol. Chem. 2017, 292, 11336–11347. [Google Scholar] [CrossRef] [PubMed]
- Massey, V.L.; Qin, L.; Cabezas, J.; Caballeria, J.; Sancho-Bru, P.; Bataller, R.; Crews, F.T. TLR7-Let-7 Signaling Contributes to Ethanol-Induced Hepatic Inflammatory Response in Mice and in Alcoholic Hepatitis. Alcohol. Clin. Exp. Res. 2018, 42, 2107–2122. [Google Scholar] [CrossRef] [PubMed]
- Brandon-Warner, E.; Feilen, N.A.; Culberson, C.R.; Field, C.O.; deLemos, A.S.; Russo, M.W.; Schrum, L.W. Processing of MiR17-92 Cluster in Hepatic Stellate Cells Promotes Hepatic Fibrogenesis During Alcohol-Induced Injury. Alcohol. Clin. Exp. Res. 2016, 40, 1430–1442. [Google Scholar] [CrossRef]
- Krauskopf, J.; de Kok, T.M.; Schomaker, S.J.; Gosink, M.; Burt, D.A.; Chandler, P.; Warner, R.L.; Johnson, K.J.; Caiment, F.; Kleinjans, J.C.; et al. Serum MicroRNA Signatures as “Liquid Biopsies” for Interrogating Hepatotoxic Mechanisms and Liver Pathogenesis in Human. PLoS ONE 2017, 12, e0177928. [Google Scholar] [CrossRef] [PubMed]
- Roderburg, C.; Mollnow, T.; Bongaerts, B.; Elfimova, N.; Vargas Cardenas, D.; Berger, K.; Zimmermann, H.; Koch, A.; Vucur, M.; Luedde, M.; et al. Micro-RNA Profiling in Human Serum Reveals Compartment-Specific Roles of MiR-571 and MiR-652 in Liver Cirrhosis. PLoS ONE 2012, 7, e32999. [Google Scholar] [CrossRef] [PubMed]
- Fründt, T.; Krause, L.; Hussey, E.; Steinbach, B.; Köhler, D.; von Felden, J.; Schulze, K.; Lohse, A.W.; Wege, H.; Schwarzenbach, H. Diagnostic and Prognostic Value of MiR-16, MiR-146a, MiR-192 and MiR-221 in Exosomes of Hepatocellular Carcinoma and Liver Cirrhosis Patients. Cancers 2021, 13, 2484. [Google Scholar] [CrossRef]
- Fuller-Pace, F.V. DExD/H Box RNA Helicases: Multifunctional Proteins with Important Roles in Transcriptional Regulation. Nucleic Acids Res. 2006, 34, 4206–4215. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, T.; Kusumanchi, P.; Tang, Q.; Sun, Z.; Radaeva, S.; Peiffer, B.; Shah, V.H.; Kamath, P.; Gores, G.J.; et al. Transcriptomic Analysis Reveals the MicroRNAs Responsible for Liver Regeneration Associated With Mortality in Alcohol-Associated Hepatitis. Hepatol. Baltim. Md 2021, 74, 2436–2451. [Google Scholar] [CrossRef]
- Heo, M.J.; Kim, T.H.; You, J.S.; Blaya, D.; Sancho-Bru, P.; Kim, S.G. Alcohol Dysregulates MiR-148a in Hepatocytes through FoxO1, Facilitating Pyroptosis via TXNIP Overexpression. Gut 2019, 68, 708–720. [Google Scholar] [CrossRef]
- Jin, X.; Yu, M.-S.; Huang, Y.; Xiang, Z.; Chen, Y.-P. MiR-30e-UCP2 Pathway Regulates Alcoholic Hepatitis Progress by Influencing ATP and Hydrogen Peroxide Expression. Oncotarget 2017, 8, 64294–64302. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yu, Y.; Li, S.; Liu, Y.; Zhou, S.; Cao, S.; Yin, J.; Li, G. MicroRNA-30a Ameliorates Hepatic Fibrosis by Inhibiting Beclin1-Mediated Autophagy. J. Cell. Mol. Med. 2017, 21, 3679–3692. [Google Scholar] [CrossRef] [PubMed]
- Saikia, P.; Roychowdhury, S.; Bellos, D.; Pollard, K.A.; McMullen, M.R.; McCullough, R.L.; McCullough, A.J.; Gholam, P.; de la Motte, C.; Nagy, L.E. Hyaluronic Acid 35 Normalizes TLR4 Signaling in Kupffer Cells from Ethanol-Fed Rats via Regulation of MicroRNA291b and Its Target Tollip. Sci. Rep. 2017, 7, 15671. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Wu, J.; Poulsen, K.L.; Kim, A.; Wu, X.; Huang, E.; Miyata, T.; Sanz-Garcia, C.; Nagy, L.E. Identification of a MicroRNA-E3 Ubiquitin Ligase Regulatory Network for Hepatocyte Death in Alcohol-Associated Hepatitis. Hepatol. Commun. 2021, 5, 830–845. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, Z.; Wang, L.; Sun, L.; Liu, Z.; Li, Q.; Yao, B.; Chen, T.; Wang, C.; Yang, W.; et al. MiR-532-3p Promotes Hepatocellular Carcinoma Progression by Targeting PTPRT. Biomed. Pharmacother. Biomed. Pharmacother. 2019, 109, 991–999. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Yu, W.; Wang, Q.; Huang, T.; Ding, Y. CircANTXR1 Contributes to the Malignant Progression of Hepatocellular Carcinoma by Promoting Proliferation and Metastasis. J. Hepatocell. Carcinoma 2021, 8, 1339–1353. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Wang, Z.; Jin, Y.; Wang, Y.; Duan, W. Loss of MiR-532-5p in Vitro Promotes Cell Proliferation and Metastasis by Influencing CXCL2 Expression in HCC. Am. J. Transl. Res. 2015, 7, 2254–2261. [Google Scholar]
- Chen, D.; Yan, Y.; Wang, X.; Li, S.; Liu, Y.; Yu, D.; He, Y.; Deng, R.; Liu, Y.; Xu, M.; et al. Chronic Alcohol Exposure Promotes HCC Stemness and Metastasis through β-Catenin/MiR-22-3p/TET2 Axis. Aging 2021, 13, 14433–14455. [Google Scholar] [CrossRef]
- Zhao, Y.; Ye, L.; Yu, Y. MicroRNA-126-5p Suppresses Cell Proliferation, Invasion and Migration by Targeting EGFR in Liver Cancer. Clin. Res. Hepatol. Gastroenterol. 2020, 44, 865–873. [Google Scholar] [CrossRef]
- Jones, K.R.; Nabinger, S.C.; Lee, S.; Sahu, S.S.; Althouse, S.; Saxena, R.; Johnson, M.S.; Chalasani, N.; Gawrieh, S.; Kota, J. Lower Expression of Tumor MicroRNA-26a Is Associated with Higher Recurrence in Patients with Hepatocellular Carcinoma Undergoing Surgical Treatment. J. Surg. Oncol. 2018, 118, 431–439. [Google Scholar] [CrossRef]
- Di Ciaula, A.; Bonfrate, L.; Krawczyk, M.; Frühbeck, G.; Portincasa, P. Synergistic and Detrimental Effects of Alcohol Intake on Progression of Liver Steatosis. Int. J. Mol. Sci. 2022, 23, 2636. [Google Scholar] [CrossRef] [PubMed]
- Tsutsumi, M.; Lasker, J.M.; Takahashi, T.; Lieber, C.S. In Vivo Induction of Hepatic P4502E1 by Ethanol: Role of Increased Enzyme Synthesis. Arch. Biochem. Biophys. 1993, 304, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Rasineni, K.; Casey, C.A. Molecular Mechanism of Alcoholic Fatty Liver. Indian J. Pharmacol. 2012, 44, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yu, D.; Tolleson, W.H.; Yu, L.-R.; Green, B.; Zeng, L.; Chen, Y.; Chen, S.; Ren, Z.; Guo, L.; et al. A Systematic Evaluation of MicroRNAs in Regulating Human Hepatic CYP2E1. Biochem. Pharmacol. 2017, 138, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.; Yao, H.; Li, C.; Pu, M.; Yao, X.; Yang, H.; Qi, X.; Ren, J.; Wang, Y. A Dual Inhibition: MicroRNA-552 Suppresses Both Transcription and Translation of Cytochrome P450 2E1. Biochim. Biophys. Acta BBA—Gene Regul. Mech. 2016, 1859, 650–662. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Sun, Q.; Zhong, W.; Sun, X.; Zhou, Z. Hepatic Peroxisome Proliferator-Activated Receptor Gamma Signaling Contributes to Alcohol-Induced Hepatic Steatosis and Inflammation in Mice. Alcohol. Clin. Exp. Res. 2016, 40, 988–999. [Google Scholar] [CrossRef]
- Zaiou, M. Peroxisome Proliferator-Activated Receptor-γ as a Target and Regulator of Epigenetic Mechanisms in Nonalcoholic Fatty Liver Disease. Cells 2023, 12, 1205. [Google Scholar] [CrossRef]
- Chen, Y.; Patel, V.; Bang, S.; Cohen, N.; Millar, J.; Kim, S.F. Maturation and Activity of Sterol Regulatory Element Binding Protein 1 Is Inhibited by Acyl-CoA Binding Domain Containing 3. PLoS ONE 2012, 7, e49906. [Google Scholar] [CrossRef]
- Xiao, X.; Song, B.-L. SREBP: A Novel Therapeutic Target. Acta Biochim. Biophys. Sin. 2013, 45, 2–10. [Google Scholar] [CrossRef]
- Zhu, L.; Liao, R.; Huang, J.; Yan, H.; Xiao, C.; Yang, Y.; Wang, H.; Yang, C. The MiR-216/MiR-217 Cluster Regulates Lipid Metabolism in Laying Hens With Fatty Liver Syndrome via PPAR/SREBP Signaling Pathway. Front. Vet. Sci. 2022, 9, 913841. [Google Scholar] [CrossRef]
- Hu, M.; Wang, F.; Li, X.; Rogers, C.Q.; Liang, X.; Finck, B.N.; Mitra, M.S.; Zhang, R.; Mitchell, D.A.; You, M. Regulation of Hepatic Lipin-1 by Ethanol: Role of AMP-Activated Protein Kinase/Sterol Regulatory Element-Binding Protein 1 Signaling in Mice. Hepatol. Baltim. Md 2012, 55, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Harris, T.E.; Finck, B.N. Dual Function Lipin Proteins and Glycerolipid Metabolism. Trends Endocrinol. Metab. TEM 2011, 22, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Ponugoti, B.; Kim, D.-H.; Xiao, Z.; Smith, Z.; Miao, J.; Zang, M.; Wu, S.-Y.; Chiang, C.-M.; Veenstra, T.D.; Kemper, J.K. SIRT1 Deacetylates and Inhibits SREBP-1C Activity in Regulation of Hepatic Lipid Metabolism. J. Biol. Chem. 2010, 285, 33959–33970. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Zhu, X.; Jiang, X.; Li, S.; Lv, Y. Transcriptional Control by HNF-1: Emerging Evidence Showing Its Role in Lipid Metabolism and Lipid Metabolism Disorders. Genes Dis. 2022, 9, 1248–1257. [Google Scholar] [CrossRef] [PubMed]
- Lawler, J.F.; Yin, M.; Diehl, A.M.; Roberts, E.; Chatterjee, S. Tumor Necrosis Factor-α Stimulates the Maturation of Sterol Regulatory Element Binding Protein-1 in Human Hepatocytes through the Action of Neutral Sphingomyelinase. J. Biol. Chem. 1998, 273, 5053–5059. [Google Scholar] [CrossRef] [PubMed]
- Donohue, T.M.; Osna, N.A.; Trambly, C.S.; Whitaker, N.P.; Thomes, P.G.; Todero, S.L.; Davis, J.S. Early Growth Response-1 Contributes to Steatosis Development after Acute Ethanol Administration. Alcohol. Clin. Exp. Res. 2012, 36, 759–767. [Google Scholar] [CrossRef] [PubMed]
- McMullen, M.R.; Pritchard, M.T.; Wang, Q.; Millward, C.A.; Croniger, C.M.; Nagy, L.E. Early Growth Response-1 Transcription Factor Is Essential for Ethanol-Induced Fatty Liver Injury in Mice. Gastroenterology 2005, 128, 2066–2076. [Google Scholar] [CrossRef]
- Li, W.; Li, K.; Wang, Z.; Fa, Z. MicroRNA-377-3p Promotes Cell Proliferation and Inhibits Cell Cycle Arrest and Cell Apoptosis in Hepatocellular Carcinoma by Affecting EGR1-Mediated P53 Activation. Pathol. Res. Pract. 2022, 234, 153855. [Google Scholar] [CrossRef]
- Osna, N.A.; Donohue, T.M.; Kharbanda, K.K. Alcoholic Liver Disease: Pathogenesis and Current Management. Alcohol Res. Curr. Rev. 2017, 38, 147–161. [Google Scholar]
- Krammer, J.; Digel, M.; Ehehalt, F.; Stremmel, W.; Füllekrug, J.; Ehehalt, R. Overexpression of CD36 and Acyl-CoA Synthetases FATP2, FATP4 and ACSL1 Increases Fatty Acid Uptake in Human Hepatoma Cells. Int. J. Med. Sci. 2011, 8, 599–614. [Google Scholar] [CrossRef]
- Lin, H.-Y.; Wang, F.-S.; Yang, Y.-L.; Huang, Y.-H. MicroRNA-29a Suppresses CD36 to Ameliorate High Fat Diet-Induced Steatohepatitis and Liver Fibrosis in Mice. Cells 2019, 8, 1298. [Google Scholar] [CrossRef]
- Wang, X.; Ma, Y.; Yang, L.-Y.; Zhao, D. MicroRNA-20a-5p Ameliorates Non-Alcoholic Fatty Liver Disease via Inhibiting the Expression of CD36. Front. Cell Dev. Biol. 2020, 8, 596329. [Google Scholar] [CrossRef] [PubMed]
- Ding, D.; Ye, G.; Lin, Y.; Lu, Y.; Zhang, H.; Zhang, X.; Hong, Z.; Huang, Q.; Chi, Y.; Chen, J.; et al. MicroRNA-26a-CD36 Signaling Pathway: Pivotal Role in Lipid Accumulation in Hepatocytes Induced by PM2.5 Liposoluble Extracts. Environ. Pollut. Barking Essex 1987 2019, 248, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Schmoldt, A.; Benthe, H.F.; Haberland, G. Digitoxin Metabolism by Rat Liver Microsomes. Biochem. Pharmacol. 1975, 24, 1639–1641. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Wang, Z.; Fillmore, R.; Xi, Y. MiR-200, a New Star MiRNA in Human Cancer. Cancer Lett. 2014, 344, 166–173. [Google Scholar] [CrossRef]
- Huang, Q.; Li, J.; Zheng, J.; Wei, A. The Carcinogenic Role of the Notch Signaling Pathway in the Development of Hepatocellular Carcinoma. J. Cancer 2019, 10, 1570–1579. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.-B.; Chen, X.-B.; Xu, L.-L.; Zhang, M.; Feng, L.; Yi, P.-S.; Tang, J.-W.; Xu, M.-Q. MiR-203 Inhibits Augmented Proliferation and Metastasis of Hepatocellular Carcinoma Residual in the Promoted Regenerating Liver. Cancer Sci. 2017, 108, 338–346. [Google Scholar] [CrossRef]
- Yang, F.; Hu, Y.; Liu, H.-X.; Wan, Y.-J.Y. MiR-22-Silenced Cyclin A Expression in Colon and Liver Cancer Cells Is Regulated by Bile Acid Receptor. J. Biol. Chem. 2015, 290, 6507–6515. [Google Scholar] [CrossRef]
- Csak, T.; Ganz, M.; Pespisa, J.; Kodys, K.; Dolganiuc, A.; Szabo, G. Fatty Acid and Endotoxin Activate Inflammasomes in Mouse Hepatocytes That Release Danger Signals to Stimulate Immune Cells. Hepatol. Baltim. Md 2011, 54, 133–144. [Google Scholar] [CrossRef]
- Glaser, T.; Baiocchi, L.; Zhou, T.; Francis, H.; Lenci, I.; Grassi, G.; Kennedy, L.; Liangpunsakul, S.; Glaser, S.; Alpini, G.; et al. Pro-Inflammatory Signalling and Gut-Liver Axis in Non-Alcoholic and Alcoholic Steatohepatitis: Differences and Similarities along the Path. J. Cell. Mol. Med. 2020, 24, 5955–5965. [Google Scholar] [CrossRef]
- Li, S.; Tan, H.-Y.; Wang, N.; Zhang, Z.-J.; Lao, L.; Wong, C.-W.; Feng, Y. The Role of Oxidative Stress and Antioxidants in Liver Diseases. Int. J. Mol. Sci. 2015, 16, 26087–26124. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Svegliati-Baroni, G.; Poniachik, J.; Baraona, E.; Lieber, C.S. Collagen Synthesis by Liver Stellate Cells Is Released from Its Normal Feedback Regulation by Acetaldehyde-Induced Modification of the Carboxyl-Terminal Propeptide of Procollagen. Alcohol. Clin. Exp. Res. 1997, 21, 1204–1211. [Google Scholar] [CrossRef] [PubMed]
- Niemelä, O.; Parkkila, S.; Ylä-Herttuala, S.; Halsted, C.; Witztum, J.L.; Lanca, A.; Israel, Y. Covalent Protein Adducts in the Liver as a Result of Ethanol Metabolism and Lipid Peroxidation. Lab. Investig. J. Tech. Methods Pathol. 1994, 70, 537–546. [Google Scholar]
- Thiele, G.M.; Duryee, M.J.; Willis, M.S.; Sorrell, M.F.; Freeman, T.L.; Tuma, D.J.; Klassen, L.W. Malondialdehyde-Acetaldehyde (MAA) Modified Proteins Induce pro-Inflammatory and pro-Fibrotic Responses by Liver Endothelial Cells. Comp. Hepatol. 2004, 3 (Suppl. 1), S25. [Google Scholar] [CrossRef] [PubMed]
- Yan, A.W.; Schnabl, B. Bacterial Translocation and Changes in the Intestinal Microbiome Associated with Alcoholic Liver Disease. World J. Hepatol. 2012, 4, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Rao, R. Endotoxemia and Gut Barrier Dysfunction in Alcoholic Liver Disease. Hepatol. Baltim. Md 2009, 50, 638–644. [Google Scholar] [CrossRef] [PubMed]
- Petrasek, J.; Csak, T.; Szabo, G. Toll-like Receptors in Liver Disease. Adv. Clin. Chem. 2013, 59, 155–201. [Google Scholar] [CrossRef]
- Takano, T.; Abe, S.; Hata, S. A Selected Ion Monitoring Method for Quantifying Simvastatin and Its Acid Form in Human Plasma, Using the Ferroceneboronate Derivative. Biomed. Environ. Mass Spectrom. 1990, 19, 577–581. [Google Scholar] [CrossRef]
- Gao, B.; Bataller, R. Alcoholic Liver Disease: Pathogenesis and New Therapeutic Targets. Gastroenterology 2011, 141, 1572–1585. [Google Scholar] [CrossRef]
- Nagy, L.E. The Role of Innate Immunity in Alcoholic Liver Disease. Alcohol Res. Curr. Rev. 2015, 37, 237–250. [Google Scholar]
- Gao, B.; Ahmad, M.F.; Nagy, L.E.; Tsukamoto, H. Inflammatory Pathways in Alcoholic Steatohepatitis. J. Hepatol. 2019, 70, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Bala, S.; Csak, T.; Kodys, K.; Catalano, D.; Ambade, A.; Furi, I.; Lowe, P.; Cho, Y.; Iracheta-Vellve, A.; Szabo, G. Alcohol-Induced MiR-155 and HDAC11 Inhibit Negative Regulators of the TLR4 Pathway and Lead to Increased LPS Responsiveness of Kupffer Cells in Alcoholic Liver Disease. J. Leukoc. Biol. 2017, 102, 487–498. [Google Scholar] [CrossRef] [PubMed]
- Bala, S.; Petrasek, J.; Mundkur, S.; Catalano, D.; Levin, I.; Ward, J.; Alao, H.; Kodys, K.; Szabo, G. Circulating MicroRNAs in Exosomes Indicate Hepatocyte Injury and Inflammation in Alcoholic, Drug-Induced, and Inflammatory Liver Diseases. Hepatol. Baltim. Md 2012, 56, 1946–1957. [Google Scholar] [CrossRef]
- Nan, Y.-M.; Wang, R.-Q.; Fu, N. Peroxisome Proliferator-Activated Receptor α, a Potential Therapeutic Target for Alcoholic Liver Disease. World J. Gastroenterol. 2014, 20, 8055–8060. [Google Scholar] [CrossRef]
- Wu, L.; Guo, C.; Wu, J. Therapeutic Potential of PPARγ Natural Agonists in Liver Diseases. J. Cell. Mol. Med. 2020, 24, 2736–2748. [Google Scholar] [CrossRef]
- Li, J.; Guo, C.; Wu, J. The Agonists of Peroxisome Proliferator-Activated Receptor-γ for Liver Fibrosis. Drug Des. Dev. Ther. 2021, 15, 2619–2628. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Friedman, J.R. MiR-122 Regulates Hepatic Lipid Metabolism and Tumor Suppression. J. Clin. Investig. 2012, 122, 2773–2776. [Google Scholar] [CrossRef] [PubMed]
- Szabo, G.; Satishchandran, A. MicroRNAs in Alcoholic Liver Disease. Semin. Liver Dis. 2015, 35, 36–42. [Google Scholar] [CrossRef]
- Li, H.-D.; Chen, X.; Yang, Y.; Huang, H.-M.; Zhang, L.; Zhang, X.; Zhang, L.; Huang, C.; Meng, X.-M.; Li, J. Wogonin Attenuates Inflammation by Activating PPAR-γ in Alcoholic Liver Disease. Int. Immunopharmacol. 2017, 50, 95–106. [Google Scholar] [CrossRef]
- Nowak, A.J.; Relja, B. The Impact of Acute or Chronic Alcohol Intake on the NF-ΚB Signaling Pathway in Alcohol-Related Liver Disease. Int. J. Mol. Sci. 2020, 21, 9407. [Google Scholar] [CrossRef]
- Luedde, T.; Schwabe, R.F. NF-ΚB in the Liver--Linking Injury, Fibrosis and Hepatocellular Carcinoma. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; He, S.; Wara, A.K.M.; Icli, B.; Shvartz, E.; Tesmenitsky, Y.; Belkin, N.; Li, D.; Blackwell, T.S.; Sukhova, G.K.; et al. Systemic Delivery of MicroRNA-181b Inhibits Nuclear Factor-ΚB Activation, Vascular Inflammation, and Atherosclerosis in Apolipoprotein E-Deficient Mice. Circ. Res. 2014, 114, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, L.; Yuan, H.; Guo, S.; Wu, G. LncRNA DANCR Promotes Sorafenib Resistance via Activation of IL-6/STAT3 Signaling in Hepatocellular Carcinoma Cells. OncoTargets Ther. 2020, 13, 1145–1157. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Lafdil, F.; Kong, X.; Gao, B. Signal Transducer and Activator of Transcription 3 in Liver Diseases: A Novel Therapeutic Target. Int. J. Biol. Sci. 2011, 7, 536–550. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.M.; Horiguchi, N.; Jeong, W.-I.; Radaeva, S.; Gao, B. Molecular Mechanisms of Alcoholic Liver Disease: Innate Immunity and Cytokines. Alcohol. Clin. Exp. Res. 2011, 35, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Servais, F.A.; Kirchmeyer, M.; Hamdorf, M.; Minoungou, N.W.E.; Rose-John, S.; Kreis, S.; Haan, C.; Behrmann, I. Modulation of the IL-6-Signaling Pathway in Liver Cells by MiRNAs Targeting Gp130, JAK1, and/or STAT3. Mol. Ther. Nucleic Acids 2019, 16, 419–433. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.M.; Cho, Y.E.; Hwang, S. Crosstalk between Oxidative Stress and Inflammatory Liver Injury in the Pathogenesis of Alcoholic Liver Disease. Int. J. Mol. Sci. 2022, 23, 774. [Google Scholar] [CrossRef]
- Shen, Z.; Liang, X.; Rogers, C.Q.; Rideout, D.; You, M. Involvement of Adiponectin-SIRT1-AMPK Signaling in the Protective Action of Rosiglitazone against Alcoholic Fatty Liver in Mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G364–G374. [Google Scholar] [CrossRef]
- Jung, Y.; Brown, K.D.; Witek, R.P.; Omenetti, A.; Yang, L.; Vandongen, M.; Milton, R.J.; Hines, I.N.; Rippe, R.A.; Spahr, L.; et al. Accumulation of Hedgehog-Responsive Progenitors Parallels Alcoholic Liver Disease Severity in Mice and Humans. Gastroenterology 2008, 134, 1532–1543. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, Y.; Wang, L.; Zhao, R.; Qiao, Y.; Han, D.; Sun, Q.; Dong, N.; Liu, Y.; Wu, D.; et al. MiR-200a Targets Gelsolin: A Novel Mechanism Regulating Secretion of Microvesicles in Hepatocellular Carcinoma Cells. Oncol. Rep. 2017, 37, 2711–2719. [Google Scholar] [CrossRef]
- Ramasamy, S.; Duraisamy, S.; Barbashov, S.; Kawano, T.; Kharbanda, S.; Kufe, D. The MUC1 and Galectin-3 Oncoproteins Function in a MicroRNA-Dependent Regulatory Loop. Mol. Cell 2007, 27, 992–1004. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yue, Y.; Wang, X.; Jin, H. Function and Clinical Potential of MicroRNAs in Hepatocellular Carcinoma. Oncol. Lett. 2015, 10, 3345–3353. [Google Scholar] [CrossRef] [PubMed]
- Chrysavgis, L.; Giannakodimos, I.; Diamantopoulou, P.; Cholongitas, E. Non-Alcoholic Fatty Liver Disease and Hepatocellular Carcinoma: Clinical Challenges of an Intriguing Link. World J. Gastroenterol. 2022, 28, 310–331. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.-C.; Zhang, Q.-B.; Qiao, L. Pathogenesis of Liver Cirrhosis. World J. Gastroenterol. 2014, 20, 7312–7324. [Google Scholar] [CrossRef] [PubMed]
- Seitz, H.K.; Bataller, R.; Cortez-Pinto, H.; Gao, B.; Gual, A.; Lackner, C.; Mathurin, P.; Mueller, S.; Szabo, G.; Tsukamoto, H. Alcoholic Liver Disease. Nat. Rev. Dis. Primer 2018, 4, 16. [Google Scholar] [CrossRef] [PubMed]
- Koyama, Y.; Brenner, D.A. Liver Inflammation and Fibrosis. J. Clin. Investig. 2017, 127, 55–64. [Google Scholar] [CrossRef] [PubMed]
- De Bleser, P.J.; Xu, G.; Rombouts, K.; Rogiers, V.; Geerts, A. Glutathione Levels Discriminate between Oxidative Stress and Transforming Growth Factor-Beta Signaling in Activated Rat Hepatic Stellate Cells. J. Biol. Chem. 1999, 274, 33881–33887. [Google Scholar] [CrossRef]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The Extracellular Matrix at a Glance. J. Cell Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef]
- Pradere, J.-P.; Kluwe, J.; De Minicis, S.; Jiao, J.-J.; Gwak, G.-Y.; Dapito, D.H.; Jang, M.-K.; Guenther, N.D.; Mederacke, I.; Friedman, R.; et al. Hepatic Macrophages but Not Dendritic Cells Contribute to Liver Fibrosis by Promoting the Survival of Activated Hepatic Stellate Cells in Mice. Hepatol. Baltim. Md 2013, 58, 1461–1473. [Google Scholar] [CrossRef]
- Szabo, G.; Bala, S. Alcoholic Liver Disease and the Gut-Liver Axis. World J. Gastroenterol. 2010, 16, 1321–1329. [Google Scholar] [CrossRef]
- Seki, E.; De Minicis, S.; Österreicher, C.H.; Kluwe, J.; Osawa, Y.; Brenner, D.A.; Schwabe, R.F. TLR4 Enhances TGF-β Signaling and Hepatic Fibrosis. Nat. Med. 2007, 13, 1324–1332. [Google Scholar] [CrossRef] [PubMed]
- Luangmonkong, T.; Suriguga, S.; Mutsaers, H.A.M.; Groothuis, G.M.M.; Olinga, P.; Boersema, M. Targeting Oxidative Stress for the Treatment of Liver Fibrosis. In Reviews of Physiology, Biochemistry and Pharmacology, Vol. 175; Nilius, B., de Tombe, P., Gudermann, T., Jahn, R., Lill, R., Eds.; Reviews of Physiology, Biochemistry and Pharmacology; Springer International Publishing: Cham, Switzerland, 2018; Volume 175, pp. 71–102. ISBN 978-3-319-95287-1. [Google Scholar]
- Paik, Y.-H.; Iwaisako, K.; Seki, E.; Inokuchi, S.; Schnabl, B.; Osterreicher, C.H.; Kisseleva, T.; Brenner, D.A. The Nicotinamide Adenine Dinucleotide Phosphate Oxidase (NOX) Homologues NOX1 and NOX2/Gp91(Phox) Mediate Hepatic Fibrosis in Mice. Hepatol. Baltim. Md 2011, 53, 1730–1741. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, H.; Yao, L.; Zhao, P.; Wu, X. MiR-34a Promotes Fibrosis of Hepatic Stellate Cells via the TGF-β Pathway. Ann. Transl. Med. 2021, 9, 1520. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Li, W.; Guo, K.; Xiao, Y.; Wang, Y.; Fan, J. MiR-181b Promotes Hepatic Stellate Cells Proliferation by Targeting P27 and Is Elevated in the Serum of Cirrhosis Patients. Biochem. Biophys. Res. Commun. 2012, 421, 4–8. [Google Scholar] [CrossRef] [PubMed]
- Pivonello, C.; De Martino, M.C.; Negri, M.; Cuomo, G.; Cariati, F.; Izzo, F.; Colao, A.; Pivonello, R. The GH-IGF-SST System in Hepatocellular Carcinoma: Biological and Molecular Pathogenetic Mechanisms and Therapeutic Targets. Infect. Agent. Cancer 2014, 9, 27. [Google Scholar] [CrossRef] [PubMed]
- Gilgenkrantz, H.; Collin de l’Hortet, A. New Insights into Liver Regeneration. Clin. Res. Hepatol. Gastroenterol. 2011, 35, 623–629. [Google Scholar] [CrossRef]
- Brandão, D.F.; Ramalho, L.N.Z.; Ramalho, F.S.; Zucoloto, S.; Martinelli, A.d.L.C.; Silva, O.d.C.e. Liver Cirrhosis and Hepatic Stellate Cells. Acta Cir. Bras. 2006, 21 (Suppl. 1), 54–57. [Google Scholar] [CrossRef]
- Seol, H.S.; Akiyama, Y.; Lee, S.-E.; Shimada, S.; Jang, S.J. Loss of MiR-100 and MiR-125b Results in Cancer Stem Cell Properties through IGF2 Upregulation in Hepatocellular Carcinoma. Sci. Rep. 2020, 10, 21412. [Google Scholar] [CrossRef]
- Ge, Y.-Y.; Shi, Q.; Zheng, Z.-Y.; Gong, J.; Zeng, C.; Yang, J.; Zhuang, S.-M. MicroRNA-100 Promotes the Autophagy of Hepatocellular Carcinoma Cells by Inhibiting the Expression of MTOR and IGF-1R. Oncotarget 2014, 5, 6218–6228. [Google Scholar] [CrossRef]
- Su, H.; Yang, J.-R.; Xu, T.; Huang, J.; Xu, L.; Yuan, Y.; Zhuang, S.-M. MicroRNA-101, down-Regulated in Hepatocellular Carcinoma, Promotes Apoptosis and Suppresses Tumorigenicity. Cancer Res. 2009, 69, 1135–1142. [Google Scholar] [CrossRef]
- Xu, Y.; An, Y.; Wang, Y.; Zhang, C.; Zhang, H.; Huang, C.; Jiang, H.; Wang, X.; Li, X. MiR-101 Inhibits Autophagy and Enhances Cisplatin-Induced Apoptosis in Hepatocellular Carcinoma Cells. Oncol. Rep. 2013, 29, 2019–2024. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Wang, Q.-L.; Shen, L.; Tao, Y.-Y.; Liu, C.-H. MicroRNA-101 Suppresses Liver Fibrosis by Downregulating PI3K/Akt/MTOR Signaling Pathway. Clin. Res. Hepatol. Gastroenterol. 2019, 43, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Xiang, T.; Ren, G.; Tan, C.; Liu, R.; Xu, X.; Wu, Z. MiR-101 Is down-Regulated by the Hepatitis B Virus x Protein and Induces Aberrant DNA Methylation by Targeting DNA Methyltransferase 3A. Cell. Signal. 2013, 25, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Yip-Schneider, M.T.; Doyle, C.J.; McKillop, I.H.; Wentz, S.C.; Brandon-Warner, E.; Matos, J.M.; Sandrasegaran, K.; Saxena, R.; Hennig, M.E.; Wu, H.; et al. Alcohol Induces Liver Neoplasia in a Novel Alcohol-Preferring Rat Model. Alcohol. Clin. Exp. Res. 2011, 35, 2216–2225. [Google Scholar] [CrossRef] [PubMed]
- Kew, M.C. The Role of Cirrhosis in the Etiology of Hepatocellular Carcinoma. J. Gastrointest. Cancer 2014, 45, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Mogilyansky, E.; Rigoutsos, I. The MiR-17/92 Cluster: A Comprehensive Update on Its Genomics, Genetics, Functions and Increasingly Important and Numerous Roles in Health and Disease. Cell Death Differ. 2013, 20, 1603–1614. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Han, C.; Wu, T. MiR-17-92 Cluster Promotes Hepatocarcinogenesis. Carcinogenesis 2015, 36, 1213–1222. [Google Scholar] [CrossRef]
- Jin, B.; Wang, W.; Meng, X.-X.; Du, G.; Li, J.; Zhang, S.-Z.; Zhou, B.-H.; Fu, Z.-H. Let-7 Inhibits Self-Renewal of Hepatocellular Cancer Stem-like Cells through Regulating the Epithelial-Mesenchymal Transition and the Wnt Signaling Pathway. BMC Cancer 2016, 16, 863. [Google Scholar] [CrossRef]
- Zhang, K.; Wong, P.; Salvaggio, C.; Salhi, A.; Osman, I.; Bedogni, B. Synchronized Targeting of Notch and ERBB Signaling Suppresses Melanoma Tumor Growth through Inhibition of Notch1 and ERBB3. J. Investig. Dermatol. 2016, 136, 464–472. [Google Scholar] [CrossRef]
- Huang, X.-P.; Hou, J.; Shen, X.-Y.; Huang, C.-Y.; Zhang, X.-H.; Xie, Y.-A.; Luo, X.-L. MicroRNA-486-5p, Which Is Downregulated in Hepatocellular Carcinoma, Suppresses Tumor Growth by Targeting PIK3R1. FEBS J. 2015, 282, 579–594. [Google Scholar] [CrossRef]
- Felgendreff, P.; Raschzok, N.; Kunze, K.; Leder, A.; Lippert, S.; Klunk, S.; Tautenhahn, H.-M.; Hau, H.-M.; Schmuck, R.B.; Reutzel-Selke, A.; et al. Tissue-Based MiRNA Mapping in Alcoholic Liver Cirrhosis: Different Profiles in Cirrhosis with or without Hepatocellular Carcinoma. Biomark. Biochem. Indic. Expo. Response Susceptibility Chem. 2020, 25, 62–68. [Google Scholar] [CrossRef]
- Szabo, G. Gut-Liver Axis in Alcoholic Liver Disease. Gastroenterology 2015, 148, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Kanel, G.C.; Korula, J. Part II Liver Biopsy Evaluation: Morphology with Differential Diagnoses. In Atlas of Liver Pathology; Elsevier: Amsterdam, The Netherlands, 2011; pp. 379–488. ISBN 978-1-4377-0765-6. [Google Scholar]
- Avila, M.A.; Dufour, J.-F.; Gerbes, A.L.; Zoulim, F.; Bataller, R.; Burra, P.; Cortez-Pinto, H.; Gao, B.; Gilmore, I.; Mathurin, P.; et al. Recent Advances in Alcohol-Related Liver Disease (ALD): Summary of a Gut Round Table Meeting. Gut 2020, 69, 764–780. [Google Scholar] [CrossRef] [PubMed]
- Shah, N.J.; Royer, A.; John, S. Alcoholic Hepatitis. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Thursz, M.; Morgan, T.R. Treatment of Severe Alcoholic Hepatitis. Gastroenterology 2016, 150, 1823–1834. [Google Scholar] [CrossRef] [PubMed]
- Bou Saleh, M.; Louvet, A.; Ntandja-Wandji, L.C.; Boleslawski, E.; Gnemmi, V.; Lassailly, G.; Truant, S.; Maggiotto, F.; Ningarhari, M.; Artru, F.; et al. Loss of Hepatocyte Identity Following Aberrant YAP Activation: A Key Mechanism in Alcoholic Hepatitis. J. Hepatol. 2021, 75, 912–923. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.-H.; Kim, S.J.; Hyun, J. MicroRNAs Regulating Hippo-YAP Signaling in Liver Cancer. Biomedicines 2021, 9, 347. [Google Scholar] [CrossRef] [PubMed]
- Momen-Heravi, F.; Bala, S.; Kodys, K.; Szabo, G. Exosomes Derived from Alcohol-Treated Hepatocytes Horizontally Transfer Liver Specific MiRNA-122 and Sensitize Monocytes to LPS. Sci. Rep. 2015, 5, 9991. [Google Scholar] [CrossRef] [PubMed]
- Raposo, G.; Stoorvogel, W. Extracellular Vesicles: Exosomes, Microvesicles, and Friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Minakawa, T.; Yamashita, J.K. Extracellular Vesicles and MicroRNAs in the Regulation of Cardiomyocyte Differentiation and Proliferation. Arch. Biochem. Biophys. 2023, 749, 109791. [Google Scholar] [CrossRef]
- Liao, M.; Qin, M.; Liu, L.; Huang, H.; Chen, N.; Du, H.; Huang, D.; Wang, P.; Zhou, H.; Tong, G. Exosomal MicroRNA Profiling Revealed Enhanced Autophagy Suppression and Anti-Tumor Effects of a Combination of Compound Phyllanthus Urinaria and Lenvatinib in Hepatocellular Carcinoma. Phytomed. Int. J. Phytother. Phytopharm. 2023, 122, 155091. [Google Scholar] [CrossRef]
- Beylerli, O.; Encarnacion Ramirez, M.d.J.; Shumadalova, A.; Ilyasova, T.; Zemlyanskiy, M.; Beilerli, A.; Montemurro, N. Cell-Free MiRNAs as Non-Invasive Biomarkers in Brain Tumors. Diagn. Basel Switz. 2023, 13, 2888. [Google Scholar] [CrossRef]
- Schulze, K.; Imbeaud, S.; Letouzé, E.; Alexandrov, L.B.; Calderaro, J.; Rebouissou, S.; Couchy, G.; Meiller, C.; Shinde, J.; Soysouvanh, F.; et al. Exome Sequencing of Hepatocellular Carcinomas Identifies New Mutational Signatures and Potential Therapeutic Targets. Nat. Genet. 2015, 47, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Barrena, M.G.; Arechederra, M.; Colyn, L.; Berasain, C.; Avila, M.A. Epigenetics in Hepatocellular Carcinoma Development and Therapy: The Tip of the Iceberg. JHEP Rep. Innov. Hepatol. 2020, 2, 100167. [Google Scholar] [CrossRef] [PubMed]
- Oura, K.; Morishita, A.; Masaki, T. Molecular and Functional Roles of MicroRNAs in the Progression of Hepatocellular Carcinoma-A Review. Int. J. Mol. Sci. 2020, 21, 8362. [Google Scholar] [CrossRef]
- Xu, X.; Tao, Y.; Shan, L.; Chen, R.; Jiang, H.; Qian, Z.; Cai, F.; Ma, L.; Yu, Y. The Role of MicroRNAs in Hepatocellular Carcinoma. J. Cancer 2018, 9, 3557–3569. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Ayub, H.; Khan, T.; Wahid, F. MicroRNA Biogenesis, Gene Silencing Mechanisms and Role in Breast, Ovarian and Prostate Cancer. Biochimie 2019, 167, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Annese, T.; Tamma, R.; De Giorgis, M.; Ribatti, D. MicroRNAs Biogenesis, Functions and Role in Tumor Angiogenesis. Front. Oncol. 2020, 10, 581007. [Google Scholar] [CrossRef]
- Liu, K.; Chen, J.; McCaughan, G.W. Animal Models for Hepatocellular Carcinoma Arising from Alcoholic and Metabolic Liver Diseases. Hepatoma Res. 2020, 6, 7. [Google Scholar] [CrossRef]
- Yasmin, A.; Regan, D.P.; Schook, L.B.; Gaba, R.C.; Schachtschneider, K.M. Transcriptional Regulation of Alcohol Induced Liver Fibrosis in a Translational Porcine Hepatocellular Carcinoma Model. Biochimie 2021, 182, 73–84. [Google Scholar] [CrossRef]
- Shen, J.; Siegel, A.B.; Remotti, H.; Wang, Q.; Santella, R.M. Identifying MicroRNA Panels Specifically Associated with Hepatocellular Carcinoma and Its Different Etiologies. Hepatoma Res. 2016, 2, 151–162. [Google Scholar] [CrossRef]
- Yang, H.; Zheng, W.; Shuai, X.; Chang, R.-M.; Yu, L.; Fang, F.; Yang, L.-Y. MicroRNA-424 Inhibits Akt3/E2F3 Axis and Tumor Growth in Hepatocellular Carcinoma. Oncotarget 2015, 6, 27736–27750. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.-Y.; Wei, H.-Y.; Liu, L.-Y.; Li, K.-M.; Wang, R.-B.; Xu, X.-Q.; Feng, R. MiR-3607, a Biomarker of Hepatocellular Carcinoma Invasion and Aggressiveness: Its Relationship with Epithelial-Mesenchymal Transition Process. IUBMB Life 2020, 72, 1686–1697. [Google Scholar] [CrossRef] [PubMed]
- Gu, W.; Li, X.; Wang, J. MiR-139 Regulates the Proliferation and Invasion of Hepatocellular Carcinoma through the WNT/TCF-4 Pathway. Oncol. Rep. 2014, 31, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Lv, T.; Jiang, L.; Kong, L.; Yang, J. MicroRNA-29c-3p Acts as a Tumor Suppressor Gene and Inhibits Tumor Progression in Hepatocellular Carcinoma by Targeting TRIM31. Oncol. Rep. 2020, 43, 953–964. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, G.; Xu, H.; Wang, H.; Bai, D. Identification of a Novel Metastasis-Related MiRNAs-Based Signature for Predicting the Prognosis of Hepatocellular Carcinoma. J. Oncol. 2021, 2021, 6629633. [Google Scholar] [CrossRef] [PubMed]
- Che, J.; Su, Z.; Yang, W.; Xu, L.; Li, Y.; Wang, H.; Zhou, W. Tumor-Suppressor P53 Specifically Binds to MiR-29c-3p and Reduces ADAM12 Expression in Hepatocellular Carcinoma. Dig. Liver Dis. Off. J. Ital. Soc. Gastroenterol. Ital. Assoc. Study Liver 2023, 55, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z.; Liu, X.; Li, Z.; Wang, G.; Feng, Z.; Liu, Y.; Yang, H.; Tan, C.; Zhang, Z.; Li, K. LncRNA LINC00667 Aggravates the Progression of Hepatocellular Carcinoma by Regulating Androgen Receptor Expression as a MiRNA-130a-3p Sponge. Cell Death Discov. 2021, 7, 387. [Google Scholar] [CrossRef]
- Li, L.; He, K.; Chen, S.; Wei, W.; Tian, Z.; Tang, Y.; Xiao, C.; Xiang, G. Circ_0001175 Promotes Hepatocellular Carcinoma Cell Proliferation and Metastasis by Regulating MiR-130a-5p. OncoTargets Ther. 2020, 13, 13315–13327. [Google Scholar] [CrossRef]
- Wang, J.; Chu, Y.; Xu, M.; Zhang, X.; Zhou, Y.; Xu, M. MiR-21 Promotes Cell Migration and Invasion of Hepatocellular Carcinoma by Targeting KLF5. Oncol. Lett. 2019, 17, 2221–2227. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, H.; Ren, L. MiR-21 Promoted Proliferation and Migration in Hepatocellular Carcinoma through Negative Regulation of Navigator-3. Biochem. Biophys. Res. Commun. 2015, 464, 1228–1234. [Google Scholar] [CrossRef]
- Franck, M.; Thon, C.; Schütte, K.; Malfertheiner, P.; Link, A. Circulating MiR-21-5p Level Has Limited Prognostic Value in Patients with Hepatocellular Carcinoma and Is Influenced by Renal Function. World J. Hepatol. 2020, 12, 1031–1045. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-Y.; Zhang, H.-F.; Wang, L.; Ma, Y.-P.; Gao, F.; Zhang, S.-J.; Wang, L.-C. MiR-21 Expression Predicts Prognosis in Hepatocellular Carcinoma. Clin. Res. Hepatol. Gastroenterol. 2014, 38, 715–719. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Yang, J.; Chen, M.; Cui, L.; Wang, T.; Gao, W.; Tian, J.; Wei, R. MicroRNA-21 as a Diagnostic Marker for Hepatocellular Carcinoma: A Systematic Review and Meta-Analysis. Pak. J. Med. Sci. 2019, 35, 1466–1471. [Google Scholar] [CrossRef] [PubMed]
- Correia de Sousa, M.; Calo, N.; Sobolewski, C.; Gjorgjieva, M.; Clément, S.; Maeder, C.; Dolicka, D.; Fournier, M.; Vinet, L.; Montet, X.; et al. Mir-21 Suppression Promotes Mouse Hepatocarcinogenesis. Cancers 2021, 13, 4983. [Google Scholar] [CrossRef] [PubMed]
- Ladeiro, Y.; Couchy, G.; Balabaud, C.; Bioulac-Sage, P.; Pelletier, L.; Rebouissou, S.; Zucman-Rossi, J. MicroRNA Profiling in Hepatocellular Tumors Is Associated with Clinical Features and Oncogene/Tumor Suppressor Gene Mutations. Hepatol. Baltim. Md 2008, 47, 1955–1963. [Google Scholar] [CrossRef] [PubMed]
- Gulyaeva, L.F.; Kushlinskiy, N.E. Regulatory Mechanisms of MicroRNA Expression. J. Transl. Med. 2016, 14, 143. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Xu, Z.; Zhang, Y.; Evert, M.; Calvisi, D.F.; Chen, X. β-Catenin Signaling in Hepatocellular Carcinoma. J. Clin. Investig. 2022, 132, e154515. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.; Lin, J.; Li, J.; Wang, X.-D.; Xu, L.-M.; Chen, D.-Z.; Chen, Y.-P. MiRNA-432 and SLC38A1 as Predictors of Hepatocellular Carcinoma Complicated with Alcoholic Steatohepatitis. Oxid. Med. Cell. Longev. 2022, 2022, 4832611. [Google Scholar] [CrossRef]
- Zheng, H.; Zou, A.E.; Saad, M.A.; Wang, X.Q.; Kwok, J.G.; Korrapati, A.; Li, P.; Kisseleva, T.; Wang-Rodriguez, J.; Ongkeko, W.M. Alcohol-Dysregulated MicroRNAs in Hepatitis B Virus-Related Hepatocellular Carcinoma. PLoS ONE 2017, 12, e0178547. [Google Scholar] [CrossRef]
- Mercer, K.E.; Hennings, L.; Sharma, N.; Lai, K.; Cleves, M.A.; Wynne, R.A.; Badger, T.M.; Ronis, M.J.J. Alcohol Consumption Promotes Diethylnitrosamine-Induced Hepatocarcinogenesis in Male Mice through Activation of the Wnt/β-Catenin Signaling Pathway. Cancer Prev. Res. Phila. Pa 2014, 7, 675–685. [Google Scholar] [CrossRef]
- Xu, Y.; Liu, L.; Liu, J.; Zhang, Y.; Zhu, J.; Chen, J.; Liu, S.; Liu, Z.; Shi, H.; Shen, H.; et al. A Potentially Functional Polymorphism in the Promoter Region of MiR-34b/c Is Associated with an Increased Risk for Primary Hepatocellular Carcinoma. Int. J. Cancer 2011, 128, 412–417. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Beckebaum, S.; Iacob, S.; Wu, G.; Kaiser, G.M.; Radtke, A.; Liu, C.; Kabar, I.; Schmidt, H.H.; Zhang, X.; et al. MicroRNA-101 Inhibits Human Hepatocellular Carcinoma Progression through EZH2 Downregulation and Increased Cytostatic Drug Sensitivity. J. Hepatol. 2014, 60, 590–598. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, X.; Jia, L.-T.; Hu, S.-J.; Zhao, J.; Yang, J.-D.; Wen, W.-H.; Wang, Z.; Wang, T.; Zhao, J.; et al. C-Myc-Mediated Epigenetic Silencing of MicroRNA-101 Contributes to Dysregulation of Multiple Pathways in Hepatocellular Carcinoma. Hepatol. Baltim. Md 2014, 59, 1850–1863. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Tian, W.; Chen, H.; Deng, Y. MicroRNA-101 Sensitizes Hepatocellular Carcinoma Cells to Doxorubicin-Induced Apoptosis via Targeting Mcl-1. Mol. Med. Rep. 2016, 13, 1923–1929. [Google Scholar] [CrossRef] [PubMed]
- Chiang, C.-W.; Huang, Y.; Leong, K.-W.; Chen, L.-C.; Chen, H.-C.; Chen, S.-J.; Chou, C.-K. PKCalpha Mediated Induction of MiR-101 in Human Hepatoma HepG2 Cells. J. Biomed. Sci. 2010, 17, 35. [Google Scholar] [CrossRef]
- Yang, J.; Lu, Y.; Lin, Y.-Y.; Zheng, Z.-Y.; Fang, J.-H.; He, S.; Zhuang, S.-M. Vascular Mimicry Formation Is Promoted by Paracrine TGF-β and SDF1 of Cancer-Associated Fibroblasts and Inhibited by MiR-101 in Hepatocellular Carcinoma. Cancer Lett. 2016, 383, 18–27. [Google Scholar] [CrossRef]
- Yuan, R.; Zhi, Q.; Zhao, H.; Han, Y.; Gao, L.; Wang, B.; Kou, Z.; Guo, Z.; He, S.; Xue, X.; et al. Upregulated Expression of MiR-106a by DNA Hypomethylation Plays an Oncogenic Role in Hepatocellular Carcinoma. Tumour Biol. J. Int. Soc. Oncodevelopmental Biol. Med. 2015, 36, 3093–3100. [Google Scholar] [CrossRef]
- Gao, J.; Dai, C.; Yu, X.; Yin, X.-B.; Zhou, F. Long Noncoding RNA LEF1-AS1 Acts as a MicroRNA-10a-5p Regulator to Enhance MSI1 Expression and Promote Chemoresistance in Hepatocellular Carcinoma Cells through Activating AKT Signaling Pathway. J. Cell. Biochem. 2021, 122, 86–99. [Google Scholar] [CrossRef]
- Gong, J.; Zhang, J.-P.; Li, B.; Zeng, C.; You, K.; Chen, M.-X.; Yuan, Y.; Zhuang, S.-M. MicroRNA-125b Promotes Apoptosis by Regulating the Expression of Mcl-1, Bcl-w and IL-6R. Oncogene 2013, 32, 3071–3079. [Google Scholar] [CrossRef]
- Song, S.; Yang, Y.; Liu, M.; Liu, B.; Yang, X.; Yu, M.; Qi, H.; Ren, M.; Wang, Z.; Zou, J.; et al. MiR-125b Attenuates Human Hepatocellular Carcinoma Malignancy through Targeting SIRT6. Am. J. Cancer Res. 2018, 8, 993–1007. [Google Scholar]
- Zhao, L.; Wang, W. MiR-125b Suppresses the Proliferation of Hepatocellular Carcinoma Cells by Targeting Sirtuin7. Int. J. Clin. Exp. Med. 2015, 8, 18469–18475. [Google Scholar] [PubMed]
- Jiang, F.; Mu, J.; Wang, X.; Ye, X.; Si, L.; Ning, S.; Li, Z.; Li, Y. The Repressive Effect of MiR-148a on TGF Beta-SMADs Signal Pathway Is Involved in the Glabridin-Induced Inhibition of the Cancer Stem Cells-like Properties in Hepatocellular Carcinoma Cells. PLoS ONE 2014, 9, e96698. [Google Scholar] [CrossRef] [PubMed]
- You, Z.; Peng, D.; Cao, Y.; Zhu, Y.; Yin, J.; Zhang, G.; Peng, X. P53 Suppresses the Progression of Hepatocellular Carcinoma via MiR-15a by Decreasing OGT Expression and EZH2 Stabilization. J. Cell. Mol. Med. 2021, 25, 9168–9182. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y. The Inhibition of MicroRNA-15a Suppresses Hepatitis B Virus-Associated Liver Cancer Cell Growth through the Smad/TGF-β Pathway. Oncol. Rep. 2017, 37, 3520–3526. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Jiao, T.; Huang, Y.; Liu, W.; Li, Z.; Ye, X. Hepatitis B Virus Regulates Apoptosis and Tumorigenesis through the MicroRNA-15a-Smad7-Transforming Growth Factor Beta Pathway. J. Virol. 2015, 89, 2739–2749. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Wu, Y.; Yang, Y.; Wang, J.; Niu, M.; Gao, S.; Qin, T.; Bao, D. Long Noncoding RNA LINC00662 Promotes M2 Macrophage Polarization and Hepatocellular Carcinoma Progression via Activating Wnt/β-Catenin Signaling. Mol. Oncol. 2020, 14, 462–483. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, S.; He, Y.; Huang, X.; Hui, Y.; Tang, Y. HOXA11-AS Regulates JAK-STAT Pathway by MiR-15a-3p/STAT3 Axis to Promote the Growth and Metastasis in Liver Cancer. J. Cell. Biochem. 2019, 120, 15941–15951. [Google Scholar] [CrossRef]
- Lin, Z.; Liu, J. LncRNA DQ786243 Promotes Hepatocellular Carcinoma Cell Invasion and Proliferation by Regulating the MiR-15b-5p/Wnt3A Axis. Mol. Med. Rep. 2021, 23, 318. [Google Scholar] [CrossRef]
- Pan, W.; Wang, L.; Zhang, X.-F.; Zhang, H.; Zhang, J.; Wang, G.; Xu, P.; Zhang, Y.; Hu, P.; Zhang, X.-D.; et al. Hypoxia-Induced MicroRNA-191 Contributes to Hepatic Ischemia/Reperfusion Injury through the ZONAB/Cyclin D1 Axis. Cell Death Differ. 2019, 26, 291–305. [Google Scholar] [CrossRef]
- Fornari, F.; Milazzo, M.; Chieco, P.; Negrini, M.; Calin, G.A.; Grazi, G.L.; Pollutri, D.; Croce, C.M.; Bolondi, L.; Gramantieri, L. MiR-199a-3p Regulates MTOR and c-Met to Influence the Doxorubicin Sensitivity of Human Hepatocarcinoma Cells. Cancer Res. 2010, 70, 5184–5193. [Google Scholar] [CrossRef]
- Henry, J.C.; Park, J.-K.; Jiang, J.; Kim, J.H.; Nagorney, D.M.; Roberts, L.R.; Banerjee, S.; Schmittgen, T.D. MiR-199a-3p Targets CD44 and Reduces Proliferation of CD44 Positive Hepatocellular Carcinoma Cell Lines. Biochem. Biophys. Res. Commun. 2010, 403, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Guan, J.; Liu, Z.; Xiao, M.; Hao, F.; Wang, C.; Chen, Y.; Lu, Y.; Liang, J. MicroRNA-199a-3p Inhibits Tumorigenesis of Hepatocellular Carcinoma Cells by Targeting ZHX1/PUMA Signal. Am. J. Transl. Res. 2017, 9, 2457–2465. [Google Scholar] [PubMed]
- Zhang, J.; Yang, Y.; Yang, T.; Liu, Y.; Li, A.; Fu, S.; Wu, M.; Pan, Z.; Zhou, W. MicroRNA-22, Downregulated in Hepatocellular Carcinoma and Correlated with Prognosis, Suppresses Cell Proliferation and Tumourigenicity. Br. J. Cancer 2010, 103, 1215–1220. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Setayesh, T.; Vaziri, F.; Wu, X.; Hwang, S.T.; Chen, X.; Yvonne Wan, Y.-J. MiR-22 Gene Therapy Treats HCC by Promoting Anti-Tumor Immunity and Enhancing Metabolism. Mol. Ther. J. Am. Soc. Gene Ther. 2023, 31, 1829–1845. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Xu, X. MicroRNA-22 Is down-Regulated in Hepatitis B Virus-Related Hepatocellular Carcinoma. Biomed. Pharmacother. Biomedecine Pharmacother. 2013, 67, 375–380. [Google Scholar] [CrossRef]
- Yang, Y.-F.; Wang, F.; Xiao, J.-J.; Song, Y.; Zhao, Y.-Y.; Cao, Y.; Bei, Y.-H.; Yang, C.-Q. MiR-222 Overexpression Promotes Proliferation of Human Hepatocellular Carcinoma HepG2 Cells by Downregulating P27. Int. J. Clin. Exp. Med. 2014, 7, 893–902. [Google Scholar]
- Wong, Q.W.-L.; Ching, A.K.-K.; Chan, A.W.-H.; Choy, K.-W.; To, K.-F.; Lai, P.B.-S.; Wong, N. MiR-222 Overexpression Confers Cell Migratory Advantages in Hepatocellular Carcinoma through Enhancing AKT Signaling. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2010, 16, 867–875. [Google Scholar] [CrossRef]
- Ma, D.; Tao, X.; Gao, F.; Fan, C.; Wu, D. MiR-224 Functions as an Onco-MiRNA in Hepatocellular Carcinoma Cells by Activating AKT Signaling. Oncol. Lett. 2012, 4, 483–488. [Google Scholar] [CrossRef]
- An, F.; Wu, X.; Zhang, Y.; Chen, D.; Lin, Y.; Wu, F.; Ding, J.; Xia, M.; Zhan, Q. MiR-224 Regulates the Aggressiveness of Hepatoma Cells Through the IL-6/STAT3/SMAD4 Pathway. Turk. J. Gastroenterol. Off. J. Turk. Soc. Gastroenterol. 2021, 32, 532–542. [Google Scholar] [CrossRef]
- Wang, Y.; Ren, J.; Gao, Y.; Ma, J.Z.I.; Toh, H.C.; Chow, P.; Chung, A.Y.F.; Ooi, L.L.P.J.; Lee, C.G.L. MicroRNA-224 Targets SMAD Family Member 4 to Promote Cell Proliferation and Negatively Influence Patient Survival. PLoS ONE 2013, 8, e68744. [Google Scholar] [CrossRef]
- Cao, Y.; Lv, W.; Ding, W.; Li, J. Sevoflurane Inhibits the Proliferation and Invasion of Hepatocellular Carcinoma Cells through Regulating the PTEN/Akt/GSK-3β/Β-catenin Signaling Pathway by Downregulating MiR-25-3p. Int. J. Mol. Med. 2020, 46, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Xue, J.; Yang, S.; Chen, Y.; Wang, Y.; Zhu, Y.; Wang, X.; Kuang, D.; Ruan, Q.; Duan, Y.; et al. Co-Targeting of IGF1R/MTOR Pathway by MiR-497 and MiR-99a Impairs Hepatocellular Carcinoma Development. Oncotarget 2017, 8, 47984–47997. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Liu, X.; Lin, L.; Hou, J.; Li, N.; Wang, C.; Wang, P.; Zhang, Q.; Zhang, P.; Zhou, W.; et al. MicroRNA-99a Inhibits Hepatocellular Carcinoma Growth and Correlates with Prognosis of Patients with Hepatocellular Carcinoma. J. Biol. Chem. 2011, 286, 36677–36685. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Jin, H.; Liu, H.; Lv, S.; Wang, B.; Wang, R.; Liu, H.; Ding, M.; Yang, Y.; Li, L.; et al. MiRNA-99a Directly Regulates AGO2 through Translational Repression in Hepatocellular Carcinoma. Oncogenesis 2014, 3, e97. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Chen, J.; Huang, Z. Recent Progress in MicroRNA-Based Delivery Systems for the Treatment of Human Disease. ExRNA 2019, 1, 24. [Google Scholar] [CrossRef]
- To, K.K.W.; Fong, W.; Tong, C.W.S.; Wu, M.; Yan, W.; Cho, W.C.S. Advances in the Discovery of MicroRNA-Based Anticancer Therapeutics: Latest Tools and Developments. Expert Opin. Drug Discov. 2020, 15, 63–83. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Wan, X.; Shan, X.; Fan, R.; Zha, W. Drugging the “Undruggable” MicroRNAs. Cell. Mol. Life Sci. CMLS 2021, 78, 1861–1871. [Google Scholar] [CrossRef]
- Krichevsky, A.; Nguyen, L.; Wei, Z.; Silva, M.; Barberán-Soler, S.; Rabinovsky, R.; Muratore, C.; Stricker, J.; Hortman, C.; Young-Pearse, T.; et al. Small Molecule Regulators of MicroRNAs Identified by High-Throughput Screen Coupled with High-Throughput Sequencing. Res. Sq. 2023. preprint. [Google Scholar] [CrossRef]
- Dasgupta, I.; Chatterjee, A. Recent Advances in MiRNA Delivery Systems. Methods Protoc. 2021, 4, 10. [Google Scholar] [CrossRef]
- Pan, Y.; Zhang, Y.; Jia, T.; Zhang, K.; Li, J.; Wang, L. Development of a MicroRNA Delivery System Based on Bacteriophage MS2 Virus-like Particles. FEBS J. 2012, 279, 1198–1208. [Google Scholar] [CrossRef]
- Sun, Y.; Sun, Y.; Zhao, R. Establishment of MicroRNA Delivery System by PP7 Bacteriophage-like Particles Carrying Cell-Penetrating Peptide. J. Biosci. Bioeng. 2017, 124, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, D.; Zhang, R.; Peng, R.; Li, J. Delivery of MicroRNA-21-Sponge and Pre-MicroRNA-122 by MS2 Virus-like Particles to Therapeutically Target Hepatocellular Carcinoma Cells. Exp. Biol. Med. Maywood NJ 2021, 246, 2463–2472. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.K.; Steinmetz, N.F. MicroRNA-181a Silencing by Antisense Oligonucleotides Delivered by Virus-like Particles. J. Mater. Chem. B 2023, 11, 816–825. [Google Scholar] [CrossRef] [PubMed]
- Vandenberghe, L.H.; Wilson, J.M. AAV as an Immunogen. Curr. Gene Ther. 2007, 7, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Verdera, H.C.; Kuranda, K.; Mingozzi, F. AAV Vector Immunogenicity in Humans: A Long Journey to Successful Gene Transfer. Mol. Ther. J. Am. Soc. Gene Ther. 2020, 28, 723–746. [Google Scholar] [CrossRef] [PubMed]
- Munir, J.; Yoon, J.K.; Ryu, S. Therapeutic MiRNA-Enriched Extracellular Vesicles: Current Approaches and Future Prospects. Cells 2020, 9, 2271. [Google Scholar] [CrossRef] [PubMed]
- Shinde, S.S.; Ahmed, S.; Malik, J.A.; Hani, U.; Khanam, A.; Ashraf Bhat, F.; Ahmad Mir, S.; Ghazwani, M.; Wahab, S.; Haider, N.; et al. Therapeutic Delivery of Tumor Suppressor MiRNAs for Breast Cancer Treatment. Biology 2023, 12, 467. [Google Scholar] [CrossRef]
- Böttger, R.; Pauli, G.; Chao, P.-H.; Al Fayez, N.; Hohenwarter, L.; Li, S.-D. Lipid-Based Nanoparticle Technologies for Liver Targeting. Adv. Drug Deliv. Rev. 2020, 154–155, 79–101. [Google Scholar] [CrossRef]
- Luo, F.; Yu, Y.; Li, M.; Chen, Y.; Zhang, P.; Xiao, C.; Lv, G. Polymeric Nanomedicines for the Treatment of Hepatic Diseases. J. Nanobiotechnol. 2022, 20, 488. [Google Scholar] [CrossRef]
- Scott, L.J. Givosiran: First Approval. Drugs 2020, 80, 335–339. [Google Scholar] [CrossRef]
- Yamaguchi, K.; Yang, L.; McCall, S.; Huang, J.; Yu, X.X.; Pandey, S.K.; Bhanot, S.; Monia, B.P.; Li, Y.-X.; Diehl, A.M. Inhibiting Triglyceride Synthesis Improves Hepatic Steatosis but Exacerbates Liver Damage and Fibrosis in Obese Mice with Nonalcoholic Steatohepatitis. Hepatol. Baltim. Md 2007, 45, 1366–1374. [Google Scholar] [CrossRef] [PubMed]
- Listenberger, L.L.; Han, X.; Lewis, S.E.; Cases, S.; Farese, R.V.; Ory, D.S.; Schaffer, J.E. Triglyceride Accumulation Protects against Fatty Acid-Induced Lipotoxicity. Proc. Natl. Acad. Sci. USA 2003, 100, 3077–3082. [Google Scholar] [CrossRef] [PubMed]
- Parlati, L.; Régnier, M.; Guillou, H.; Postic, C. New Targets for NAFLD. JHEP Rep. Innov. Hepatol. 2021, 3, 100346. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yu, B.; Ren, W.; Mo, X.; Zhou, C.; He, H.; Jia, H.; Wang, L.; Jacob, S.T.; Lee, R.J.; et al. Enhanced Hepatic Delivery of SiRNA and MicroRNA Using Oleic Acid Based Lipid Nanoparticle Formulations. J. Control. Release Off. J. Control. Release Soc. 2013, 172, 690–698. [Google Scholar] [CrossRef] [PubMed]
- Ellipilli, S.; Wang, H.; Binzel, D.W.; Shu, D.; Guo, P. Ligand-Displaying-Exosomes Using RNA Nanotechnology for Targeted Delivery of Multi-Specific Drugs for Liver Cancer Regression. Nanomed. Nanotechnol. Biol. Med. 2023, 50, 102667. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Zhang, F.; Wu, J.; Zhuge, Y. Nanotechnology in Drug Delivery for Liver Fibrosis. Front. Mol. Biosci. 2021, 8, 804396. [Google Scholar] [CrossRef]
- Kim, A.; Saikia, P.; Nagy, L.E. MiRNAs Involved in M1/M2 Hyperpolarization Are Clustered and Coordinately Expressed in Alcoholic Hepatitis. Front. Immunol. 2019, 10, 1295. [Google Scholar] [CrossRef]
- Zeng, T.; Zhang, C.-L.; Xiao, M.; Yang, R.; Xie, K.-Q. Critical Roles of Kupffer Cells in the Pathogenesis of Alcoholic Liver Disease: From Basic Science to Clinical Trials. Front. Immunol. 2016, 7, 538. [Google Scholar] [CrossRef]
- Hong, D.S.; Kang, Y.-K.; Borad, M.; Sachdev, J.; Ejadi, S.; Lim, H.Y.; Brenner, A.J.; Park, K.; Lee, J.-L.; Kim, T.-Y.; et al. Phase 1 Study of MRX34, a Liposomal MiR-34a Mimic, in Patients with Advanced Solid Tumours. Br. J. Cancer 2020, 122, 1630–1637. [Google Scholar] [CrossRef]
- Cheng, S.-H.; Li, F.-C.; Souris, J.S.; Yang, C.-S.; Tseng, F.-G.; Lee, H.-S.; Chen, C.-T.; Dong, C.-Y.; Lo, L.-W. Visualizing Dynamics of Sub-Hepatic Distribution of Nanoparticles Using Intravital Multiphoton Fluorescence Microscopy. ACS Nano 2012, 6, 4122–4131. [Google Scholar] [CrossRef]
- Shilpi, S. Drug Targeting Strategies for Liver Cancer and Other Liver Diseases. MOJ Drug Des. Dev. Ther. 2018, 2, 171–177. [Google Scholar] [CrossRef]
- Hanna, J.; Hossain, G.S.; Kocerha, J. The Potential for MicroRNA Therapeutics and Clinical Research. Front. Genet. 2019, 10, 478. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, C.; Sharma, A.R.; Sharma, G.; Lee, S.-S. Therapeutic Advances of MiRNAs: A Preclinical and Clinical Update. J. Adv. Res. 2021, 28, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Iacomino, G. MiRNAs: The Road from Bench to Bedside. Genes 2023, 14, 314. [Google Scholar] [CrossRef]
- Roberts, T.C.; Langer, R.; Wood, M.J.A. Advances in Oligonucleotide Drug Delivery. Nat. Rev. Drug Discov. 2020, 19, 673–694. [Google Scholar] [CrossRef]
- Kim, T.; Croce, C.M. MicroRNA: Trends in Clinical Trials of Cancer Diagnosis and Therapy Strategies. Exp. Mol. Med. 2023, 55, 1314–1321. [Google Scholar] [CrossRef]
- Chakraborty, C.; Sharma, A.R.; Sharma, G.; Doss, C.G.P.; Lee, S.-S. Therapeutic MiRNA and SiRNA: Moving from Bench to Clinic as Next Generation Medicine. Mol. Ther. Nucleic Acids 2017, 8, 132–143. [Google Scholar] [CrossRef]
- Hsu, S.-H.; Yu, B.; Wang, X.; Lu, Y.; Schmidt, C.R.; Lee, R.J.; Lee, L.J.; Jacob, S.T.; Ghoshal, K. Cationic Lipid Nanoparticles for Therapeutic Delivery of SiRNA and MiRNA to Murine Liver Tumor. Nanomed. Nanotechnol. Biol. Med. 2013, 9, 1169–1180. [Google Scholar] [CrossRef]
- Yang, C.; Yin, M.; Xu, G.; Lin, W.-J.; Chen, J.; Zhang, Y.; Feng, T.; Huang, P.; Chen, C.-K.; Yong, K.-T. Biodegradable Polymers as a Noncoding MiRNA Nanocarrier for Multiple Targeting Therapy of Human Hepatocellular Carcinoma. Adv. Healthc. Mater. 2019, 8, e1801318. [Google Scholar] [CrossRef]
- Ning, Q.; Liu, Y.-F.; Ye, P.-J.; Gao, P.; Li, Z.-P.; Tang, S.-Y.; He, D.-X.; Tang, S.-S.; Wei, H.; Yu, C.-Y. Delivery of Liver-Specific MiRNA-122 Using a Targeted Macromolecular Prodrug toward Synergistic Therapy for Hepatocellular Carcinoma. ACS Appl. Mater. Interfaces 2019, 11, 10578–10588. [Google Scholar] [CrossRef]
- Li, Y.; Luan, Y.; Li, J.; Song, H.; Li, Y.; Qi, H.; Sun, B.; Zhang, P.; Wu, X.; Liu, X.; et al. Exosomal MiR-199a-5p Promotes Hepatic Lipid Accumulation by Modulating MST1 Expression and Fatty Acid Metabolism. Hepatol. Int. 2020, 14, 1057–1074. [Google Scholar] [CrossRef]
- Niu, Q.; Wang, T.; Wang, Z.; Wang, F.; Huang, D.; Sun, H.; Liu, H. Adipose-Derived Mesenchymal Stem Cell-Secreted Extracellular Vesicles Alleviate Non-Alcoholic Fatty Liver Disease via Delivering MiR-223-3p. Adipocyte 2022, 11, 572–587. [Google Scholar] [CrossRef]
- He, S.; Guo, W.; Deng, F.; Chen, K.; Jiang, Y.; Dong, M.; Peng, L.; Chen, X. Targeted Delivery of MicroRNA 146b Mimic to Hepatocytes by Lactosylated PDMAEMA Nanoparticles for the Treatment of NAFLD. Artif. Cells Nanomed. Biotechnol. 2018, 46, 217–228. [Google Scholar] [CrossRef]
- Wang, Z.; Zhao, K.; Zhang, Y.; Duan, X.; Zhao, Y. Anti-GPC3 Antibody Tagged Cationic Switchable Lipid-Based Nanoparticles for the Co-Delivery of Anti-MiRNA27a And Sorafenib in Liver Cancers. Pharm. Res. 2019, 36, 145. [Google Scholar] [CrossRef]
- Wu, J.; Huang, J.; Kuang, S.; Chen, J.; Li, X.; Chen, B.; Wang, J.; Cheng, D.; Shuai, X. Synergistic MicroRNA Therapy in Liver Fibrotic Rat Using MRI-Visible Nanocarrier Targeting Hepatic Stellate Cells. Adv. Sci. Weinh. Baden-Wurtt. Ger. 2019, 6, 1801809. [Google Scholar] [CrossRef]
- El-Mezayen, N.S.; El-Hadidy, W.F.; El-Refaie, W.M.; Shalaby, T.I.; Khattab, M.M.; El-Khatib, A.S. Hepatic Stellate Cell-Targeted Imatinib Nanomedicine versus Conventional Imatinib: A Novel Strategy with Potent Efficacy in Experimental Liver Fibrosis. J. Control. Release Off. J. Control. Release Soc. 2017, 266, 226–237. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Niu, D.; Zhang, J.; Zhang, W.; Yao, Y.; Li, P.; Gong, J. Amphiphilic Core-Shell Nanoparticles Containing Dense Polyethyleneimine Shells for Efficient Delivery of MicroRNA to Kupffer Cells. Int. J. Nanomed. 2016, 11, 2785–2797. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, L.O.; Skerlj, R.T.; Bridger, G.J.; Schwartz, T.W. Molecular Interactions of Cyclam and Bicyclam Non-Peptide Antagonists with the CXCR4 Chemokine Receptor. J. Biol. Chem. 2001, 276, 14153–14160. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Hang, Y.; Tang, W.; Sil, D.; Jensen-Smith, H.C.; Bennett, R.G.; McVicker, B.L.; Oupický, D. Dually Active Polycation/MiRNA Nanoparticles for the Treatment of Fibrosis in Alcohol-Associated Liver Disease. Pharmaceutics 2022, 14, 669. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Ding, L.; Yu, A.; Tang, W.; Tang, S.; Zhang, C.; Oupický, D. A Boronate-Based Modular Assembly Nanosystem to Block the Undesirable Crosstalk between Hepatic Stellate Cells and Kupffer Cells. Bioact. Mater. 2023, 25, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.T.; Zhang, D.; Zhou, K.; Lee, S.M.; Liu, S.; Dilliard, S.A.; Farbiak, L.; Chatterjee, S.; Lin, Y.-H.; Siegwart, D.J. Lipid Nanoparticle (LNP) Chemistry Can Endow Unique In Vivo RNA Delivery Fates within the Liver That Alter Therapeutic Outcomes in a Cancer Model. Mol. Pharm. 2022, 19, 3973–3986. [Google Scholar] [CrossRef] [PubMed]
- Vaschetto, L.M. MiRNA Activation Is an Endogenous Gene Expression Pathway. RNA Biol. 2018, 15, 826–828. [Google Scholar] [CrossRef] [PubMed]
- Xiang, H.; Tu, B.; Luo, M.; Hou, P.; Wang, J.; Zhang, R.; Wu, L. Knockdown of UCA1 Attenuated the Progression of Alcoholic Fatty Disease by Sponging MiR-214. Mamm. Genome Off. J. Int. Mamm. Genome Soc. 2022, 33, 534–542. [Google Scholar] [CrossRef] [PubMed]
- Cho, C.J.; Myung, S.-J.; Chang, S. ADAR1 and MicroRNA; A Hidden Crosstalk in Cancer. Int. J. Mol. Sci. 2017, 18, 799. [Google Scholar] [CrossRef]
- Tomaselli, S.; Bonamassa, B.; Alisi, A.; Nobili, V.; Locatelli, F.; Gallo, A. ADAR Enzyme and MiRNA Story: A Nucleotide That Can Make the Difference. Int. J. Mol. Sci. 2013, 14, 22796–22816. [Google Scholar] [CrossRef]






| Identification | Title | Phase | miRNA Target | Disease |
|---|---|---|---|---|
| NCT01727934 | Miravirsen Study in Null Responder to Pegylated Interferon Alpha Plus Ribavirin Subjects with Chronic Hepatitis C | II | miR-122 | Hepatitis C virus infection |
| NCT02862145 | Pharmacodynamics Study of MRX34, MicroRNA Liposomal Injection in Melanoma Patients with Biopsy Accessible Lesions (MRX34-102) | I | miR-34 | Advanced melanoma |
| NCT03373786 | A Study of RG-012 in Subjects with Alport Syndrome | I | miR-21 | Alport syndrome |
| NCT02369198 | MesomiR 1: A Phase I Study of TargomiRs as 2nd or 3rd Line Treatment for Patients with Recurrent MPM and NSCLC | I | miR-16 | Malignant Pleural Mesothelioma (MPM) and Advanced Non-Small Cell Lung Cancer (NSCLC) |
| NCT03601052 | Efficacy, Safety, and Tolerability of Remlarsen (MRG-201) Following Intradermal Injection in Subjects with a History of Keloids | II | miR-29 | Keloid formation |
| NCT03837457 | PRISM: Efficacy and Safety of Cobomarsen (MRG-106) in Subjects with Mycosis Fungoides Who Have Completed the SOLAR Study (PRISM) | II | miR-155 | Cutaneous T-Cell Lymphoma (CTCL) and Mycosis Fungoides (MF) |
| NCT0280552 | Safety, Tolerability and Pharmacokinetics of MRG-106 in Patients with Mycosis Fungoides (MF), CLL, DLBCL or ATLL | I | miR-155 | Cutaneous T-Cell Lymphoma (CTCL), Mycosis Fungoides (MF), Chronic Lymphocytic Leukemia (CLL), Diffuse Large B-Cell Lymphoma (DLBCL) and Adult T-Cell Leukemia/Lymphoma (ATLL) |
| NCT03713320 | SOLAR: Efficacy and Safety of Cobomarsen (MRG-106) vs. Active Comparator in Subjects with Mycosis Fungoides (SOLAR) | II | miR-155 | Cutaneous T-Cell Lymphoma (CTCL) and Mycosis Fungoides (MF) |
| NCT03603431 | Safety, Tolerability, Pharmacokinetics, and Pharmacodynamics of MRG-110 Following Intradermal Injection in Healthy Volunteers | I | miR-92a | Ischemia |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jouve, M.; Carpentier, R.; Kraiem, S.; Legrand, N.; Sobolewski, C. MiRNAs in Alcohol-Related Liver Diseases and Hepatocellular Carcinoma: A Step toward New Therapeutic Approaches? Cancers 2023, 15, 5557. https://doi.org/10.3390/cancers15235557
Jouve M, Carpentier R, Kraiem S, Legrand N, Sobolewski C. MiRNAs in Alcohol-Related Liver Diseases and Hepatocellular Carcinoma: A Step toward New Therapeutic Approaches? Cancers. 2023; 15(23):5557. https://doi.org/10.3390/cancers15235557
Chicago/Turabian StyleJouve, Mickaël, Rodolphe Carpentier, Sarra Kraiem, Noémie Legrand, and Cyril Sobolewski. 2023. "MiRNAs in Alcohol-Related Liver Diseases and Hepatocellular Carcinoma: A Step toward New Therapeutic Approaches?" Cancers 15, no. 23: 5557. https://doi.org/10.3390/cancers15235557