
Potential Tumor Suppressor Role of Polo-like Kinase 5 in Cancer
by
 , , and
, , and
Shengqin Su
1,†,‡ ,
,
Mary Ann Ndiaye
1,†,
Glorimar Guzmán-Pérez
1,
Rebecca Michael Baus
2,
Wei Huang
2,
Manish Suresh Patankar
3,4 and
Nihal Ahmad
1,4,* 1
Department of Dermatology, University of Wisconsin, Madison, WI 53705, USA
2
Department of Pathology and Laboratory Medicine, University of Wisconsin, Madison, WI 53705, USA
3
Department of Obstetrics and Gynecology, University of Wisconsin, Madison, WI 53792, USA
4
William S. Middleton VA Medical Center, Madison, WI 53705, USA
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
‡
Current address: Department of Radiation Oncology, Stanford Cancer Institute and Stanford University School of Medicine, Stanford, CA 94304, USA.
Cancers 2023, 15(22), 5457; https://doi.org/10.3390/cancers15225457
Submission received: 24 October 2023
/
Revised: 11 November 2023
/
Accepted: 15 November 2023
/
Published: 17 November 2023
(This article belongs to the Special Issue The Signal-Processing in Tumor)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Cancer is a complex disease and the underlying molecular mechanisms driving cancer initiation and progression is crucial for the development of effective approaches for cancer management. Exploring the molecular landscape of cancer can shed light on novel biomarkers, therapeutic targets, and strategies for personalized medicine. In this study, we determined the potential involvement of PLK5 in multiple cancers employing cancer tissue microarrays (TMAs). We found a downregulation of PLK5 in these cancers. Further, using publicly available GTEx and TCGA databases, we validated our findings and extended our investigations to additional cancer types. Overall, our data suggested a potential tumor suppressor role for PLK5.
Abstract
The polo-like kinase (PLK) family of serine/threonine kinases contains five members (PLK1–5). Most PLKs are involved in cell cycle regulation and DNA damage response. However, PLK5 is different as it lacks a functional kinase domain and is not involved in cell cycle control. PLK5 remains the least-studied family member, and its role in oncogenesis remains enigmatic. Here, we identified tissues with high PLK5 expression by leveraging the Protein Atlas and GTEx databases with relevant literature and selected ovarian, lung, testis, endometrium, cervix, and fallopian tube tissues as candidates for further investigation. Subsequently, we performed immunohistochemical staining for PLK5 on multiple tissue microarrays followed by Vectra scanning and quantitative inForm analysis. This revealed consistently downregulated PLK5 expression in these cancers compared to normal tissues. To validate and extend our findings, we performed pan-cancer analysis of PLK5 expression using public RNAseq databases (TCGA and GTEx). We found PLK5 is downregulated in 18 cancer types, including our selected candidates. Interestingly, we also observed PLK5 expression remains consistently low in later stages of cancer, suggesting PLK5 may have a greater role in tumor initiation than cancer progression. Overall, our study demonstrates PLK5 downregulation in multiple cancers, highlighting its role as a tumor suppressor.
1. Introduction
Cancer, a complex, heterogeneous, and potentially fatal disease characterized by uncontrolled cell growth and proliferation continues to pose significant challenges in global health. Understanding the underlying molecular mechanisms driving cancer initiation, progression, and metastasis is crucial for the development of effective approaches for cancer management. Exploring the molecular landscape of cancer can shed light on novel biomarkers, therapeutic targets, and strategies for personalized medicine. In this context, the polo-like kinase (PLK) family of serine/threonine kinases, primarily shown to be key regulators of cell cycle, has emerged as a compelling area of investigation in cancer research [1,2,3,4,5,6]. PLKs are evolutionarily conserved from yeast to humans and are distinguished by two essential components: the polo-box domain (PBD), which facilitates protein interactions, and the kinase domain (KD), which regulates the catalytic activity of the protein [1,2,3,4,5,6,7]. In humans, five PLK family members have been identified (PLK1-5). Although primarily involved in cell cycle regulation, several of the family members have also been implicated in other key cellular pathways and processes, including apoptosis [8], DNA damage response [9], and cancers [9,10]. The most recently identified member of the PLK family, PLK5, was initially classified as a pseudogene [11,12]. However, recent evidence indicates that PLK5 encodes a functional protein [13,14]. Interestingly, although PLK5 is conserved in most mammals, human PLK5 has a marked difference as compared to others such as mouse. Full-length mouse PLK5 protein consists of 599 amino acids, including a complete KD and two PBDs [5,7,14]. In contrast, human PLK5 contains a premature stop codon in the KD, followed by an in-frame ATG codon [13]. Consequently, the human PLK5 protein is truncated, resulting in the translation of a smaller protein containing 336 amino acids [5,13]. Despite this incomplete KD in human PLK5, the protein appears to be functional, as evidenced by an impaired G2/M checkpoint after etoposide treatment in cells depleted of PLK5 by shRNA [14]. Curiously, while in mice the PLK5 protein has a full length KD, it has been shown to lack kinase activity, suggesting that PLK5 function may be independent of its kinase domain [7,13,15].
Interestingly, although PLK5 is generally a low expressing protein, its levels have been found to be higher in more differentiated and specialized tissues such as ovary, brain, and eye [13]. Among these tissues, PLK5 expression is particularly high in the cerebellum, suggesting its potential involvement in neuronal development and function. Nevertheless, the role PLK5 plays in different cells and tissues remains vastly unknown, despite its identification over two decades ago. To date, very limited research has been performed regarding the role of PLK5 in cancer, with only a few studies showing decreased PLK5 expression in non-small-cell lung cancer [16,17], breast cancer [18], medulloblastoma [19] and glioblastomas [13]. Additionally, a 2016 study found a novel metastasis-associated three-nucleotide deletion mutation of PLK5 in clear cell renal cell carcinoma [20] and a 2020 study identified a rare p.G223V variant of PLK5 in a small subset of ovarian cancer patients [21]. Therefore, given the limited available knowledge and potential impact of changes to PLK5 expression and/or sequence, it is crucial to expand our understanding to further elucidate the involvement of PLK5 in cancer. In this study, we employed quantitative measurement of immunohistochemical staining to conduct analysis of PLK5 protein expression on a range of disease-specific tissue microarrays (TMAs), including lung, cervix, ovarian, endometrial, fallopian, and testicular cancers. Additionally, using GTEx and TCGA databases, we validated our findings in these tumor types and extended our investigations to an additional 12 cancer types. Further, using the TCGA database, we looked at overall survival and progression-free survival of cancer patients with high and low PLK5 expression levels to determine if PLK5 expression may have an influence on survival. Overall, our data suggested a potential tumor suppressor role for PLK5.
2. Materials and Methods
2.1. Immunohistochemistry
Tissue microarrays (TMAs) were obtained from US Biomax (Rockville, MD, USA) (Cervix: #CR1001b; Endometrium: #OD-CT-RpUTR03-02; Fallopian Tube: #UTE601; Lung: #BC04002a; Normal Ovary: #OV806; Ovarian Disease: #OV1005b; Testis: #TE803) and University of Wisconsin (UW) Carbone Cancer Center Translational Science BioBank (Ovarian Cancer TMA). Immunostaining was performed by the UW Translational Research Initiatives in Pathology (TRIP) Lab on a Ventana Ultra BioMarker Platform (Roche, Indianapolis, IN, USA). Deparaffinization was carried out on the instrument, as was heat-induced epitope retrieval with CC2 buffer, a citrate-based buffer, for approximately 60 min at 100 °C. Primary antibody (Anti-PLK5 (#HPA035024; Sigma-Aldrich, St. Louis, MO, USA) diluted 1:50 in DaVinci Green (#PD900H; BioCare Medical, Pacheco, CA, USA)) was added and the slides were incubated for 60 min at 37 °C. After rinsing, discovery OmniMap anti-Rabbit HRP was added (Ventana #760-4311; Roche) and incubated for 16 min at 37 °C before rinsing again and adding Discovery ChromoMap DAB detection (Ventana #760-159; Roche). The slides were then rinsed again and Cytokeratin primary antibody (Dako #M3515; Agilent, Santa Clara, CA, USA) diluted 1:1000 in DaVinci Green (BioCare Medical #PD904H) was applied and incubated for 8 min at 37 °C. This was followed by incubation with Discovery UltraMap anti-Mouse AP (Ventana #760-4312; Roche) for 8 min at 37 °C and subsequent detection with Discovery RED (Ventana #760-228; Roche). The slides were counterstained with hematoxylin (1:5) for 30 sec, rinsed, dehydrated and coverslips were mounted.
2.2. Vectra Scanning/inForm Analysis
TMA slide scanning and image acquisition was carried out using the Vectra automated multispectral imaging system (Akoya Biosciences, Marlborough, MA, USA). Employing the Nuance software (version 3.02; PerkinElmer, Waltham, MA, USA) we created a spectral library in order to unmix the signals on the acquired images stained with DAB, Discovery RED, and hematoxylin. This aids in the recognition of the different chromogens based on their unique spectral curves and to analyze protein levels in each TMA core. Using the inForm software (version 2.4.8; Akoya Biosciences, Marlborough, MA, USA), we trained the program on distinguishing between different tissue segments (epithelium vs. stroma) and cell segments (nucleus, cytoplasm, cell membrane) within the TMA tissue cores. Following the software training, quantitative image analysis was carried out in each tissue core using inForm.
2.3. Statistical Analysis for Tissue Microarrays (TMAs)
In each of the tissue cores, values for epithelium only were used. The normalized optical density (OD) per unit area [OD/peak weighting value within each cell was divided by the cell area (square microns)] for the DAB chromogen was considered as the level of PLK5 expression. Statistical analysis was conducted using R software (v4.3.1) and its developmental environment RStudio (v 2023.06.1+524). PLK5 expression was compared across the disease stages in different cancer types. The boxplot between two or multiple groups and simple linear regression were conducted using ggplot2 (v3.3.3) and ggpmisc (v0.5.2) packages.
2.4. TCGA and GTEx Database Analyses of PLK5 Expression
The results shown here are in part based upon data generated by The Cancer Genome Atlas (TCGA) research network: https://www.cancer.gov/tcga (accessed on 10 May 2023). The Genotype-Tissue Expression (GTEx) project was supported by the common fund of the Office of the Director of the National Institutes of Health and by NCI NHGRI NHLBI NIDA NIMH and NINDS. The data used for the analyses described in this manuscript were obtained from GTEx Analysis Release V8 (dbGaP Accession phs000424.v8.p2).” PLK5 expression profiles in normal tissues was queried in the GTEx portal (https://www.gtexportal.org/home/gene/PLK5 (accessed on 10 May 2023)). PLK5 expression between normal vs. tumor in selected tissues were downloaded from OncoDB [22]. The pan-cancer and stage-wise analysis of PLK5 were queried in Gepia2 [23]. T-test was performed with q < 0.05 as significance cutoff for pan-cancer PLK5 analysis. ANOVA was conducted to compare PLK5 stage-wise expression analysis in selected cancers. The correlation of PLK5 and age/sex were queried in cBioPortal [24]. For TCGA/GTEx analysis, PLK5 expression is plotted in log2(TPM+1) scale.
2.5. Survival Analysis of PLK5 Expression in TCGA Database
The available data on PLK5 expression and the overall and progression-free survival of patients were downloaded from cBioPortal for selected cancer types (accessed on 6 November 2023) [25]. Subsequently, we conducted Cox proportional-hazards (PH) regression and log-rank test using the survival (v3.5-7) and survminer (v0.4.9) packages in R. For Cox proportional-hazards (PH) regression, we computed the hazard ratio and the p-value from the Wald test to assess the association between PLK5 expression and the survival of cancer patients.
For the log-rank test, the cancer patients were divided into two groups based on their PLK5 expression: (1) If more than half of the patients had zero PLK5 expression, we categorized patients with non-zero PLK5 expression as “High PLK5,” while those with zero PLK5 expression were labeled as “Low PLK5.” (2) If more than half of the patients had non-zero PLK5 expression, we assigned patients with PLK5 expression greater than or equal to the median as “High PLK5,” and those with PLK5 expression less than the median as “Low PLK5.” Next, Kaplan–Meier survival curves were generated for the two groups using survminer (v0.4.9) within R.
3. Results and Discussion
3.1. PLK5 Expression Is High in the Normal Tissues of Brain, Respiratory System, and Reproductive System
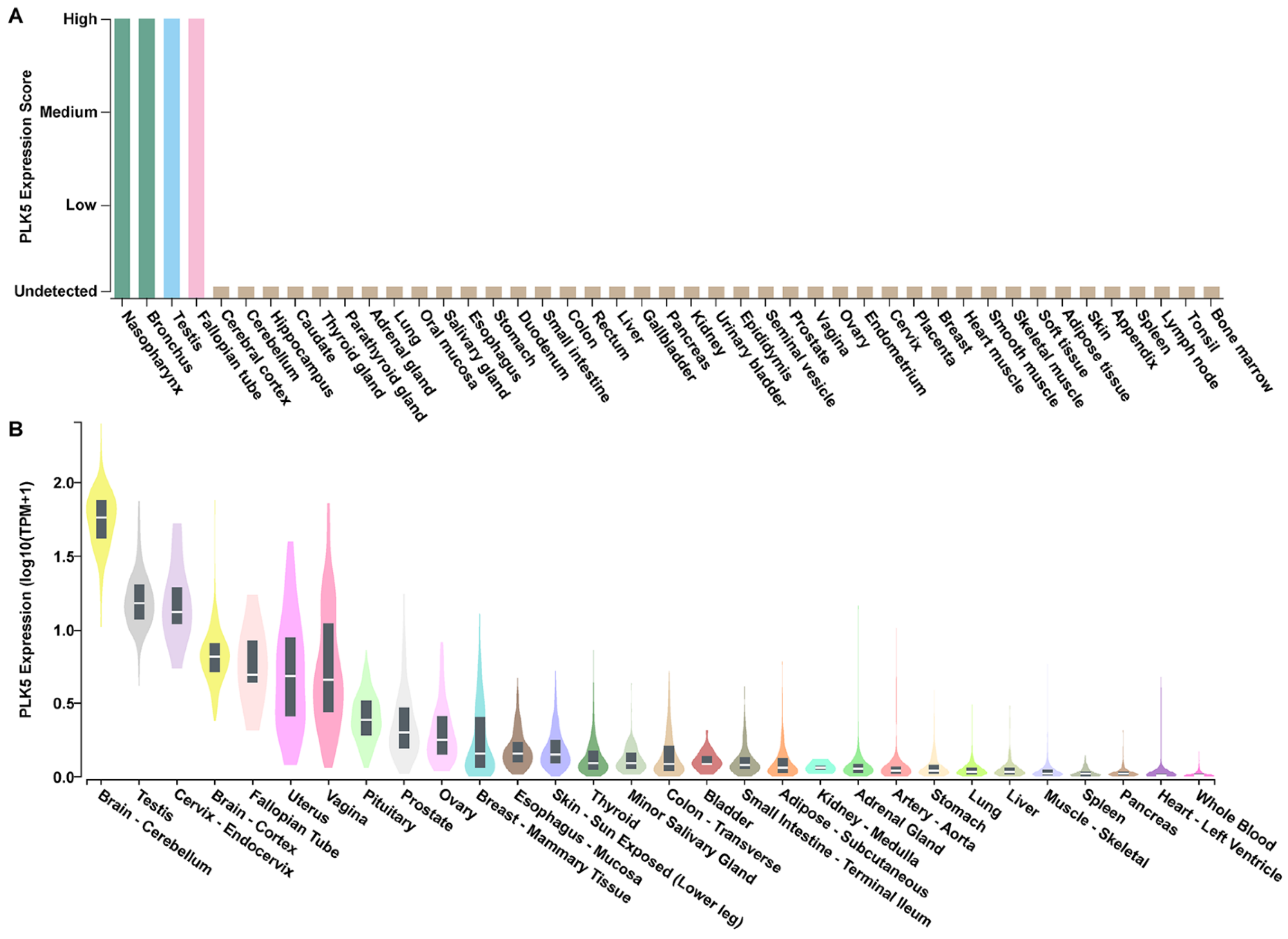
To determine the tissue-specific expression pattern of PLK5 and its potential association with cancer, we conducted a comprehensive analysis using publicly available databases. First, we utilized data compiled by the Human Protein Atlas [26,27,28,29] on PLK5 expression across 44 different normal tissue types as determined by immunohistochemical staining and interpretation by trained pathologists. PLK5 expression was shown to be highest in nasopharynx, bronchus, testis, and fallopian tube tissues, while the remaining tissues exhibited very low PLK5 (Figure 1A).
To validate these findings, we analyzed the mRNA expression profile of PLK5 using the GTEx database. We found that while PLK5 expression is generally low across most tissues, it is significantly elevated in brain tissues (cerebellum, cortex, and pituitary), female reproductive tissues (cervix, fallopian tube, uterus, vagina, ovary, and breast), and male reproductive tissues (testis and prostate) (Figure 1B). To reinforce the robustness of our findings, we performed additional analysis using two RNAseq databases, HPA (Human Protein Atlas [28]) and FANTOM5 [30,31]. The results from these independent cohorts are consistent with the findings that PLK5 is highly expressed in brain as well as female and male reproductive tissues (Supplementary Figure S1A,B).
3.2. PLK5 Expression Is Downregulated in Cancer Tissues, as Shown by TMA Analyses
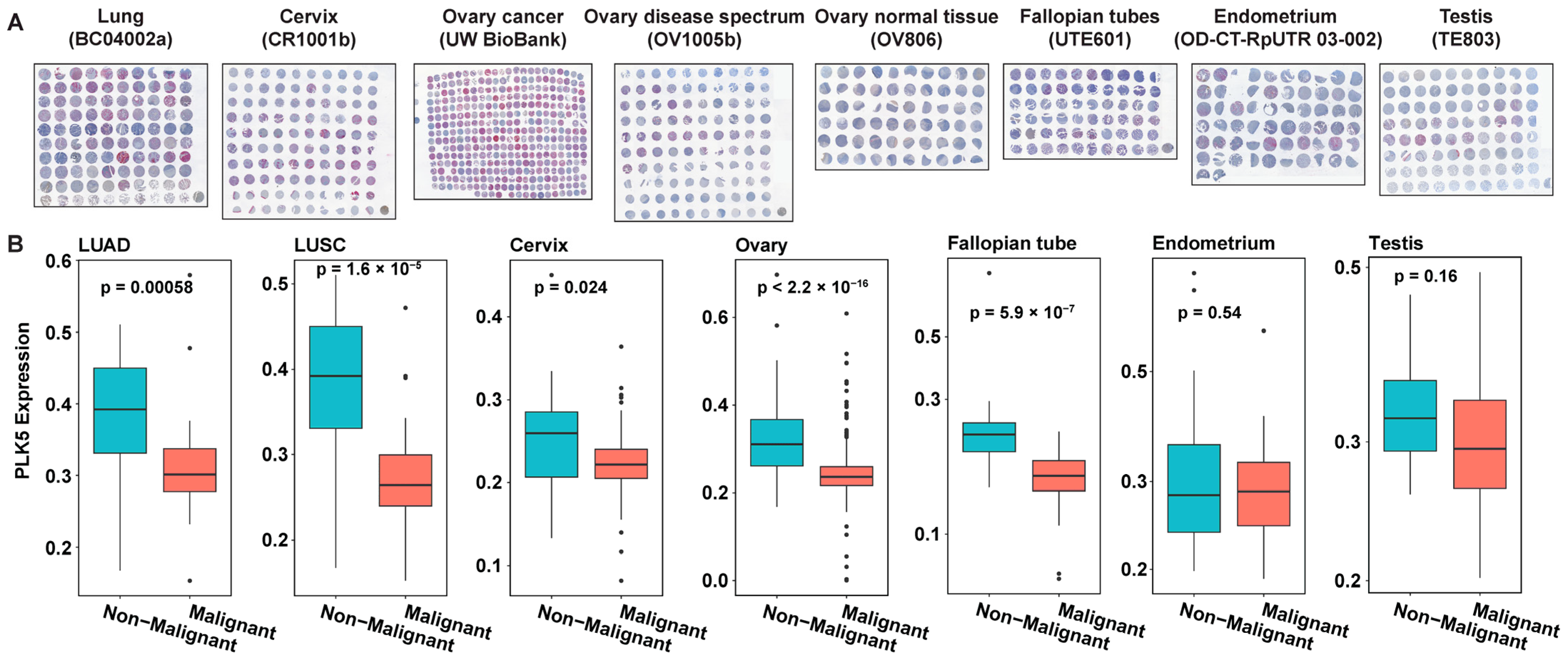
In view of earlier published studies supporting a tumor suppressor role of PLK5 expression in non-small-cell lung cancer [16,17], breast cancer [18], medulloblastoma [19] and glioblastomas [13], we were interested in expanding the available knowledge on the role of this kinase in additional cancer types. Our observation of high PLK5 expression in certain specific normal tissues led us to further investigate the expression profile of PLK5 in cancers of these same tissue types to determine its potential tumor suppressor role in these tissues. For this purpose, we employed multiple tissue microarrays (TMAs, for more information please see Supplementary Table S1) containing samples from lung, cervix, ovary, fallopian tube, endometrium, and testis (Figure 2A). The TMAs were immunostained for PLK5, followed by Vectra scanning and inForm analysis for quantitative analyses of protein expression, where we compared the PLK5 protein levels in cancer tissues versus their normal counterparts (Supplementary Table S2). We excluded brain tissue from our investigation, due to an earlier published paper on certain brain cancers [13]. As shown by our data, we found significantly reduced levels of PLK5 protein in lung, cervical, ovarian, and fallopian tube cancers, and a decreasing trend towards significance in endometrial and testicular cancer tissues (Figure 2B). These findings are consistent with a potential tumor suppressor role of PLK5 as reported earlier in certain cancers as mentioned earlier. However, additional detailed studies are required to determine the exact role of PLK5 in cancer.
3.3. PLK5 Expression Is Reduced in Multiple Malignant Tissues Compared to Normal Tissues, as Shown by Analysis of the GTEx and TCGA Databases
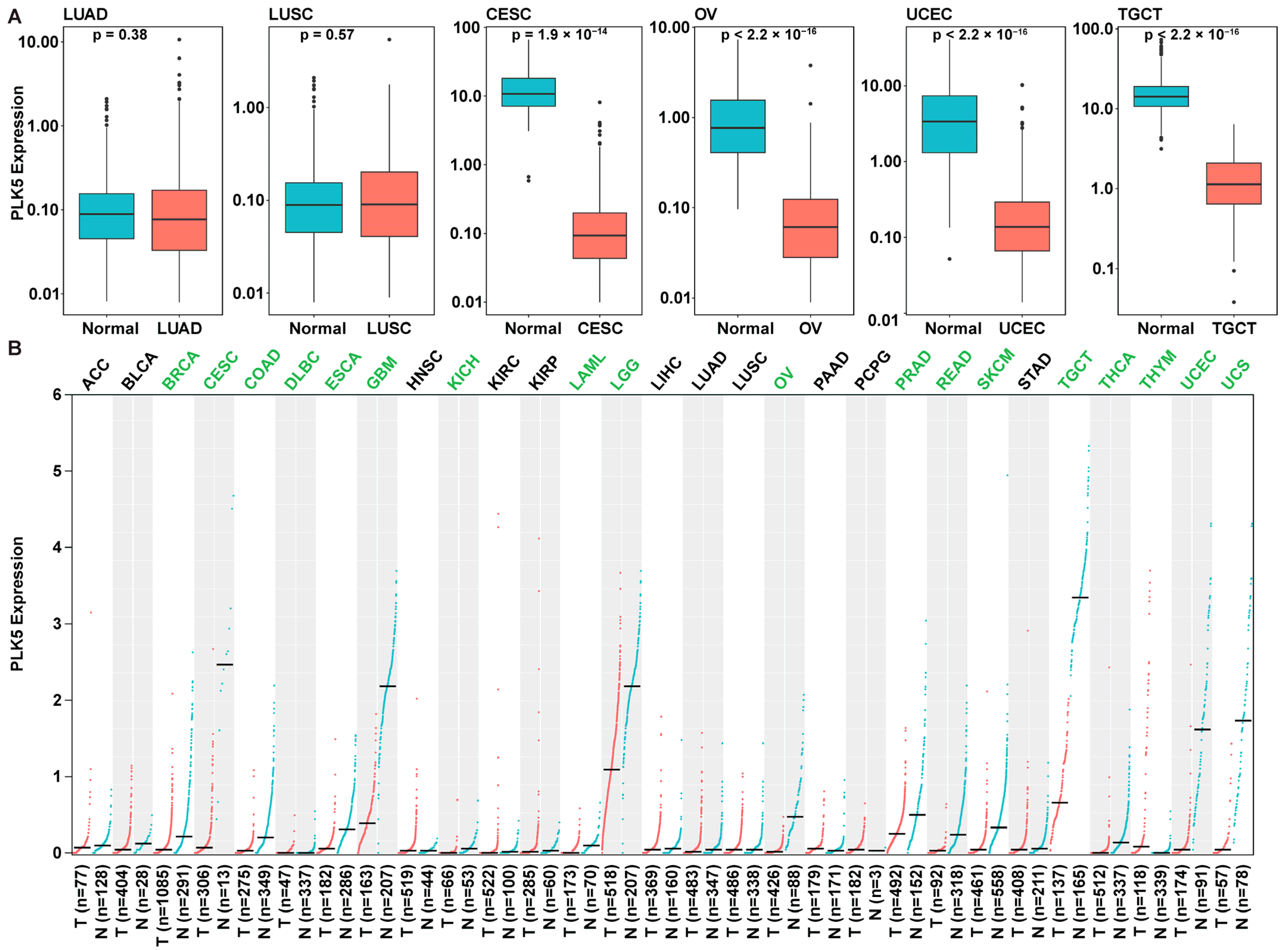
To validate our findings from the TMA analysis, we corroborated our results using independent cohorts from the GTEx and TCGA databases. We compared the PLK5 expressions in normal tissue from both TCGA and GTEx alongside cancer tissues from TCGA. Encouragingly, our results were shown to be aligned with our earlier TMA analysis findings, and we found a consistent downregulation of PLK5 expression in ovary (OV; ovarian serous cystadenocarcinoma), cervical (CESC; cervical squamous cell carcinoma and endocervical adenocarcinoma), endometrial (UCEC; uterine corpus endometrial carcinoma), and testicular cancers (TGCT; uterine corpus endometrial carcinoma) (Figure 3A). This congruence between TMA analysis and the independent GTEx/TCGA datasets lends robustness to our findings and strengthens the reliability of our conclusions regarding a potential tumor suppressor function of PLK5.
To broaden the scope of our investigation, we conducted a pan-cancer analysis of PLK5 expression using the extensive TCGA and GTEx databases (Figure 3B). As shown in the figure, we observed a broader downregulation of PLK5 in an additional 13 malignancies arising from tissues with both high and low PLK5 expression. Notably, we confirmed a reduction of PLK5 in cancers originating from tissues with elevated PLK5 expression, including brain cancers (e.g., glioblastoma, low-grade gliomas), breast cancers, prostate adenocarcinoma, and tenosynovial giant cell tumors. Moreover, reduced PLK5 levels were also observed in cancers arising from tissues with relatively low PLK5 expression, including colon adenocarcinoma, diffuse large B-cell lymphoma, esophageal carcinoma, kidney chromophobe, acute myeloid leukemia, rectum adenocarcinoma, cutaneous melanoma, thyroid carcinoma, thymoma, and uterine carcinosarcoma. Our findings suggest that PLK5 may play an important role in cancer and may have potential as a diagnostic or therapeutic biomarker.
3.4. PLK5 Levels Remain Consistent during Cancer Progression and Are Independent of Age or Sex
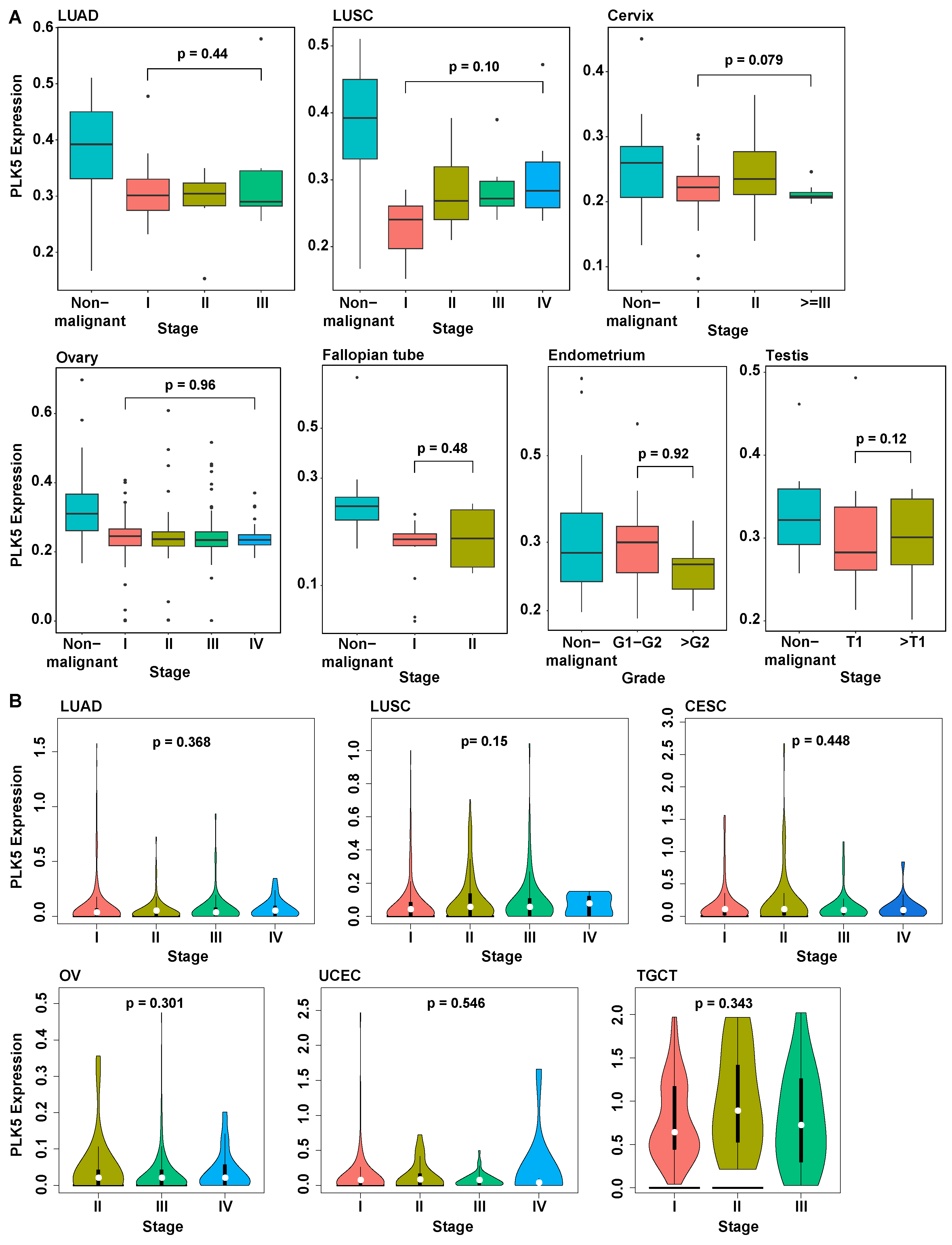
To further investigate the involvement of PLK5 in cancer progression and metastasis, we analyzed its expression levels in various cancer stages. In our TMA dataset, we observed that, although PLK5 levels were reduced in cancer tissues compared to normal, the PLK5 protein levels remained consistently low as cancer progressed (Figure 4A). Analysis of PLK5 levels across different cancer stages using TCGA analysis corroborated our results, demonstrating that during the progression of cancer, PLK5 expression remained stable in cervix, endometrium, lung, ovary, and testicular cancers (Figure 4B). Additionally, our analysis of an additional nine tumor types showed the same trends of stable PLK5 expression after the initial reduction in expression (Supplementary Figure S2). Collectively, these findings suggest that although PLK5 may be involved in tumor initiation and/or early-stage development, its significance may be limited in cancer progression and metastasis once a malignancy is established. Further experiments are warranted to fully elucidate the precise role of PLK5 in carcinogenesis and cancer progression.
We next sought to explore the correlation of PLK5 expression with age and/or sex in the context of our TMAs as well as TCGA database (Supplementary Figure S3). In our TMA analysis, we found decreased PLK5 expression with increasing age in fallopian and cervical cancer, as well as increasing PLK5 with increasing age in ovarian cancer (Figure S3A). However, no significant correlation was observed between PLK5 expression and age in endometrium, lung, or cervical cancer. On the other hand, other than lung adenocarcinoma (LUAD), no significant correlation was found between age and PLK5 expression in any of the other cancers in the TCGA database, although a trend was seen in the testicular cancer dataset. Similarly, no significant correlation between PLK5 and sex was found in lung cancer in either the TMA or TCGA datasets (Supplementary Figure S4). These findings highlight the potentially complex nature of PLK5 regulation in different cancer types. Further studies are warranted to unravel the intricate interplay between PLK5 expression, age, sex, and specific cancer types to enhance our understanding of its clinical implications.
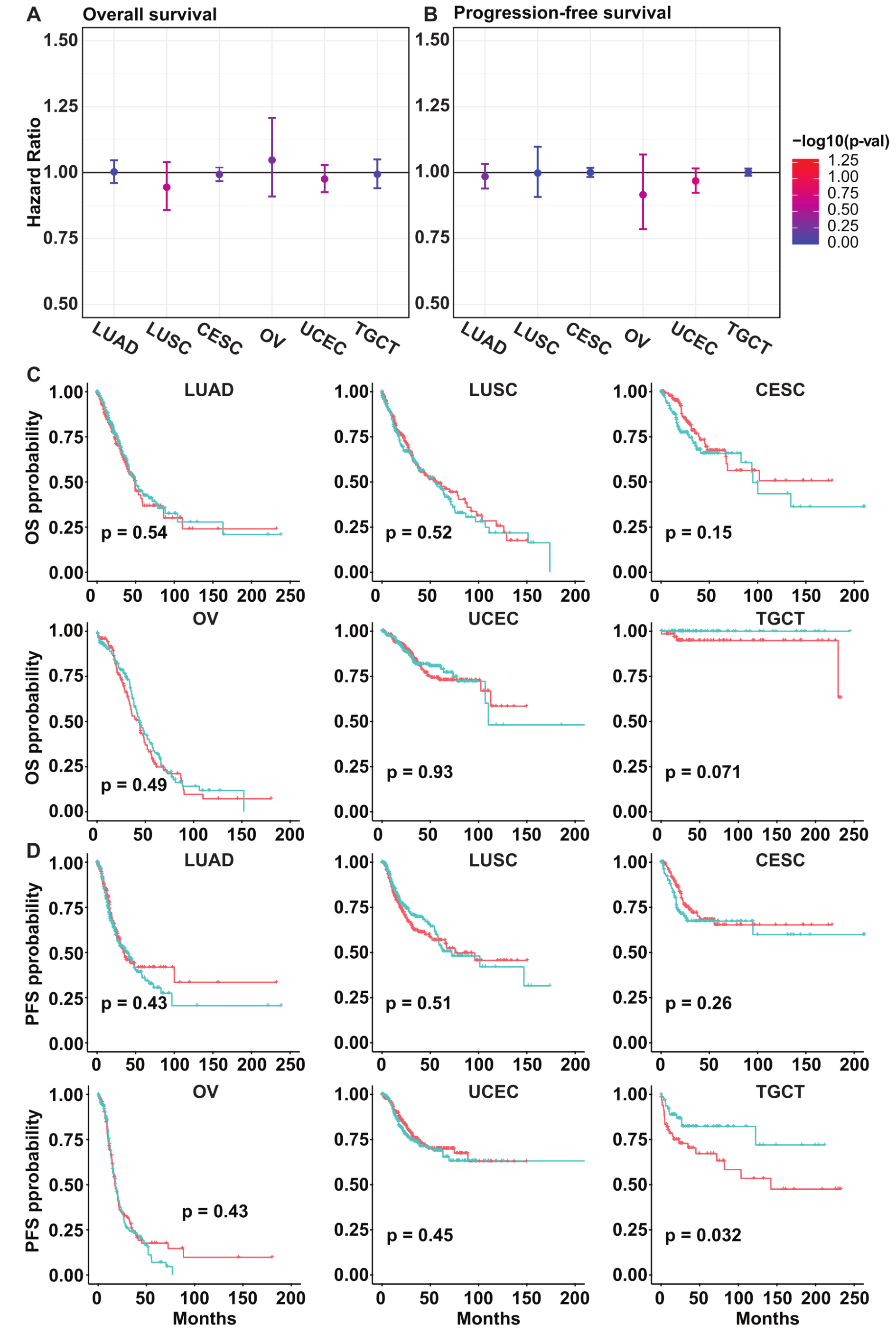
3.5. PLK5 Levels Have No Association with the Overall or Progression-Free Survival in Various Cancer Types
Given that PLK5 expression remains consistently low during cancer progression, we anticipate that PLK5 plays a limited role in cancer prognosis. To further investigate this, we conducted Cox proportional-hazards (PH) regression analysis on cervix, endometrium, lung, ovary, and testis cancers using the TCGA database. Our analysis revealed that the hazard ratio for PLK5 is not significantly different from 1 in both overall survival (Figure 5A) and progression-free survival (Figure 5B). This indicates that PLK5 expression does not appear to impact cancer prognosis in these cancers. In addition to these findings, Kaplan–Meier curves were plotted to compare survival between PLK5-high and PLK5-low patients. Similar to the hazard ratio, the data show no discernible difference in overall survival (Figure 5C) or progression-free survival (Figure 5D) among most cancer types. Furthermore, we extended our hazard ratio and Kaplan–Meier analyses to an additional nine tumor types we identified earlier. Similar to the other cancers tested, these results showed no consistent association between PLK5 expression and patient survival (Supplementary Figure S5). In summary, our findings suggest that PLK5 expression remains low during cancer progression and has negligible influence on patient survival. This is not an unexpected finding since PLK5 levels, in general, are extremely low in cancer throughout the different stages of cancer progression. However, although there appears to be no effect on survival, further research is needed to determine what role PLK5 may play in cancer due to its reduced expression in cancer tissues and the tumor suppressive qualities found by us and others.
4. Conclusions
The role of PLK5 in cancer remains vastly understudied, warranting further research to clarify its role in cancer development and progression. In this study, we have found that PLK5 is downregulated in multiple types of cancers, suggesting a potential tumor suppressor function of this protein. Interestingly, we observed that PLK5 expression remains consistent as cancer progresses and that PLK5 expression does not affect either overall survival or progression-free survival in the cancer types we tested, suggesting a limited role for PLK5 in driving cancer progression and metastasis. However, in-depth research is necessary to confirm these results in independent datasets and experimental models.
While the downregulation of PLK5 in many cancers holds promise for further research, the mechanisms underlying its regulation and potential therapeutic implications are still elusive. Future studies could explore the molecular pathways involved in PLK5 regulation and its interactions with other PLK family members and cancer-related genes, which may provide valuable insights into its contribution to tumorigenesis. A detailed understanding of the upstream and downstream effectors of PLK5 could be very useful towards the potential clinical exploitation of this target in cancer. Additionally, further investigation is needed to determine if PLK5 could be used as a target for cancer management. Continued research in these areas will advance our understanding of the complex role of PLK5 in cancer and may unveil novel avenues for cancer management. For example, novel small molecule and natural product agonists could be developed for cancer chemoprevention and cancer interception, since PLK5 appears to be involved in the early stages of the carcinogenesis process.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/cancers15225457/s1, Figure S1: Expression profile of PLK5 across multiple tissue types. Figure S2: PLK5 stage-wise expression in TCGA. Figure S3: Correlation of PLK5 expression with age in selcted tumor types. Figure S4: Correlation of PLK5 expression with biological sex in selected non-reproductive tissues. Figure S5: Analysis of PLK5 expression and patient survival in additional cancer types. Table S1: Tissue microarray (TMA) Details. Table S2: Clinical information of TMA samples and quantification of PLK5.
Author Contributions
Conceptualization, S.S., M.A.N., G.G.-P., M.S.P. and N.A.; data curation, S.S., G.G.-P., M.A.N. and N.A.; formal analysis, S.S., G.G.-P. and M.A.N.; funding acquisition, N.A.; investigation, S.S., M.A.N., G.G.-P., R.M.B. and W.H.; resources, N.A.; supervision, N.A.; validation, S.S., G.G.-P., M.A.N., R.M.B., W.H. and N.A.; writing-original draft, S.S., M.A.N. and G.G.-P.; writing-review and editing, S.S., M.A.N., G.G.-P., R.M.B., W.H., M.S.P. and N.A. All authors have read and agreed to the published version of the manuscript.
Funding
This work was partially supported by funding from the NIH (R01CA261937), and the Department of Veterans Affairs (Merit Review Awards; BLR&D I01BX005917; CSR&D I01CX002210; and a Senior Research Career Scientist Award BLR&D IK6BX006041).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Data used in this study were from TCGA and GTEx databases. These are publicly available databases and data submitted to the databases were collected using ethically approved methods and IRB boards. The tissue microarrays were obtained from a reputable supplier that collects human tissues under HIPPA-approve protocols with informed consent. Our study was based on open source and publicly available data, so no ethical issues or conflicts of interest exist.
Data Availability Statement
Data used are from publicly available databases and are accessible to everyone. Our tissue microarray data has been provided in the Supplementary Materials section as well.
Acknowledgments
The authors would like to acknowledge the University of Wisconsin Translational Research Initiatives in Pathology laboratory (TRIP), supported by the UW Department of Pathology and Laboratory Medicine, UWCCC (P30CA014520) and the Office of The Director-NIH (S10OD023526), for use of its facilities and services for TMA staining and analyses. The authors are also thankful to the University of Wisconsin Carbone Cancer Center pilot funds and BioBank, supported by P30CA014520. We would also like to acknowledge support from the Dr. Frederic E. Mohs’s Skin Cancer Research Chair endowment to N.A.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cholewa, B.D.; Liu, X.; Ahmad, N. The role of polo-like kinase 1 in carcinogenesis: Cause or consequence? Cancer Res. 2013, 73, 6848–6855. [Google Scholar] [CrossRef] [PubMed]
- Garvey, D.R.; Chhabra, G.; Ndiaye, M.A.; Ahmad, N. Role of Polo-Like Kinase 4 (PLK4) in Epithelial Cancers and Recent Progress in its Small Molecule Targeting for Cancer Management. Mol. Cancer Ther. 2021, 20, 632–640. [Google Scholar] [CrossRef]
- Gutteridge, R.E.; Ndiaye, M.A.; Liu, X.; Ahmad, N. Plk1 Inhibitors in Cancer Therapy: From Laboratory to Clinics. Mol. Cancer Ther. 2016, 15, 1427–1435. [Google Scholar] [CrossRef]
- Su, S.; Chhabra, G.; Singh, C.K.; Ndiaye, M.A.; Ahmad, N. PLK1 inhibition-based combination therapies for cancer management. Transl. Oncol. 2022, 16, 101332. [Google Scholar] [CrossRef] [PubMed]
- Strebhardt, K. Multifaceted polo-like kinases: Drug targets and antitargets for cancer therapy. Nat. Rev. Drug Discov. 2010, 9, 643–660. [Google Scholar] [CrossRef] [PubMed]
- Takai, N.; Hamanaka, R.; Yoshimatsu, J.; Miyakawa, I. Polo-like kinases (Plks) and cancer. Oncogene 2005, 24, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Archambault, V.; Lepine, G.; Kachaner, D. Understanding the Polo Kinase machine. Oncogene 2015, 34, 4799–4807. [Google Scholar] [CrossRef]
- Lee, S.Y.; Jang, C.; Lee, K.A. Polo-like kinases (plks), a key regulator of cell cycle and new potential target for cancer therapy. Dev. Reprod. 2014, 18, 65–71. [Google Scholar] [CrossRef]
- Winkles, J.A.; Alberts, G.F. Differential regulation of polo-like kinase 1, 2, 3, and 4 gene expression in mammalian cells and tissues. Oncogene 2005, 24, 260–266. [Google Scholar] [CrossRef]
- Liu, X. Targeting Polo-Like Kinases: A Promising Therapeutic Approach for Cancer Treatment. Transl. Oncol. 2015, 8, 185–195. [Google Scholar] [CrossRef]
- Caenepeel, S.; Charydczak, G.; Sudarsanam, S.; Hunter, T.; Manning, G. The mouse kinome: Discovery and comparative genomics of all mouse protein kinases. Proc. Natl. Acad. Sci. USA 2004, 101, 11707–11712. [Google Scholar] [CrossRef] [PubMed]
- Manning, G.; Whyte, D.B.; Martinez, R.; Hunter, T.; Sudarsanam, S. The protein kinase complement of the human genome. Science 2002, 298, 1912–1934. [Google Scholar] [CrossRef] [PubMed]
- de Carcer, G.; Escobar, B.; Higuero, A.M.; Garcia, L.; Anson, A.; Perez, G.; Mollejo, M.; Manning, G.; Melendez, B.; Abad-Rodriguez, J.; et al. Plk5, a polo box domain-only protein with specific roles in neuron differentiation and glioblastoma suppression. Mol. Cell Biol. 2011, 31, 1225–1239. [Google Scholar] [CrossRef] [PubMed]
- Andrysik, Z.; Bernstein, W.Z.; Deng, L.; Myer, D.L.; Li, Y.Q.; Tischfield, J.A.; Stambrook, P.J.; Bahassi, E.M. The novel mouse Polo-like kinase 5 responds to DNA damage and localizes in the nucleolus. Nucleic Acids Res. 2010, 38, 2931–2943. [Google Scholar] [CrossRef] [PubMed]
- de Carcer, G.; Manning, G.; Malumbres, M. From Plk1 to Plk5: Functional evolution of polo-like kinases. Cell Cycle 2011, 10, 2255–2262. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Shen, S.; Dong, L.; Fang, Q.; Hou, X.; Shi, X. Polo-like kinase (PLK) 5, a new member of the PLK family, serves as a biomarker to indicate anabatic tumor burden and poor prognosis for resectable non-small cell lung cancer. Front. Surg. 2022, 9, 964044. [Google Scholar] [CrossRef]
- Zeng, Y.; Li, N.; Liu, W.; Zeng, M.; Cheng, J.; Huang, J. Analyses of expressions and prognostic values of Polo-like kinases in non-small cell lung cancer. J. Cancer Res. Clin. Oncol. 2020, 146, 2447–2460. [Google Scholar] [CrossRef]
- Jiawei, W.; Xiajun, B.; Tian, S.; Xuzheng, G.; Zhenwang, Z. Comprehensive analysis of PLKs expression and prognosis in breast cancer. Cancer Genet. 2022, 268–269, 83–92. [Google Scholar] [CrossRef]
- Pezuk, J.A.; Brassesco, M.S.; de Oliveira, R.S.; Machado, H.R.; Neder, L.; Scrideli, C.A.; Tone, L.G. PLK1-associated microRNAs are correlated with pediatric medulloblastoma prognosis. Childs Nerv. Syst. 2017, 33, 609–615. [Google Scholar] [CrossRef]
- Li, Z.; Hao, P.; Wu, Q.; Li, F.; Zhao, J.; Wu, K.; Qu, C.; Chen, Y.; Li, M.; Chen, X.; et al. Genetic mutations associated with metastatic clear cell renal cell carcinoma. Oncotarget 2016, 7, 16172–16179. [Google Scholar] [CrossRef]
- Auguste, A.; Blanc-Durand, F.; Deloger, M.; Le Formal, A.; Bareja, R.; Wilkes, D.C.; Richon, C.; Brunn, B.; Caron, O.; Devouassoux-Shisheboran, M.; et al. Small Cell Carcinoma of the Ovary, Hypercalcemic Type (SCCOHT) beyond SMARCA4 Mutations: A Comprehensive Genomic Analysis. Cells 2020, 9, 1496. [Google Scholar] [CrossRef] [PubMed]
- Tang, G.; Cho, M.; Wang, X. OncoDB: An interactive online database for analysis of gene expression and viral infection in cancer. Nucleic Acids Res. 2022, 50, D1334–D1339. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Kang, B.; Li, C.; Chen, T.; Zhang, Z. GEPIA2: An enhanced web server for large-scale expression profiling and interactive analysis. Nucleic Acids Res. 2019, 47, W556–W560. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal 2013, 6, pl1. [Google Scholar] [CrossRef]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef]
- Uhlen, M.; Zhang, C.; Lee, S.; Sjostedt, E.; Fagerberg, L.; Bidkhori, G.; Benfeitas, R.; Arif, M.; Liu, Z.; Edfors, F.; et al. A pathology atlas of the human cancer transcriptome. Science 2017, 357, 6352. [Google Scholar] [CrossRef]
- Thul, P.J.; Akesson, L.; Wiking, M.; Mahdessian, D.; Geladaki, A.; Ait Blal, H.; Alm, T.; Asplund, A.; Bjork, L.; Breckels, L.M.; et al. A subcellular map of the human proteome. Science 2017, 356, 6340. [Google Scholar] [CrossRef]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Uhlen, M.; Bjorling, E.; Agaton, C.; Szigyarto, C.A.; Amini, B.; Andersen, E.; Andersson, A.C.; Angelidou, P.; Asplund, A.; Asplund, C.; et al. A human protein atlas for normal and cancer tissues based on antibody proteomics. Mol. Cell Proteomics 2005, 4, 1920–1932. [Google Scholar] [CrossRef]
- Lizio, M.; Abugessaisa, I.; Noguchi, S.; Kondo, A.; Hasegawa, A.; Hon, C.C.; de Hoon, M.; Severin, J.; Oki, S.; Hayashizaki, Y.; et al. Update of the FANTOM web resource: Expansion to provide additional transcriptome atlases. Nucleic Acids Res. 2019, 47, D752–D758. [Google Scholar] [CrossRef]
- Lizio, M.; Harshbarger, J.; Shimoji, H.; Severin, J.; Kasukawa, T.; Sahin, S.; Abugessaisa, I.; Fukuda, S.; Hori, F.; Ishikawa-Kato, S.; et al. Gateways to the FANTOM5 promoter level mammalian expression atlas. Genome Biol. 2015, 16, 22. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
PLK5 expression using publicly available databases. (A) PLK5 protein expression profile using immunohistochemical staining of pan-tissue TMAs containing 44 different normal tissue types, as available from proteinatlas.org. Image credit: Human Protein Atlas, www.proteinatlas.org (accessed on 10 May 2023) [28]. Image has been edited for style only and original image is available at the following URL: v23.proteinatlas.org/ENSG00000185988-PLK5/tissue. (B) PLK5 mRNA expression as determined using the GTEx RNA-Seq database consisting of 54 non-diseased tissue sites from ~1000 individuals to create a public dataset. Tissues with the same organ of origin are labeled with the same color.
Figure 1.
PLK5 expression using publicly available databases. (A) PLK5 protein expression profile using immunohistochemical staining of pan-tissue TMAs containing 44 different normal tissue types, as available from proteinatlas.org. Image credit: Human Protein Atlas, www.proteinatlas.org (accessed on 10 May 2023) [28]. Image has been edited for style only and original image is available at the following URL: v23.proteinatlas.org/ENSG00000185988-PLK5/tissue. (B) PLK5 mRNA expression as determined using the GTEx RNA-Seq database consisting of 54 non-diseased tissue sites from ~1000 individuals to create a public dataset. Tissues with the same organ of origin are labeled with the same color.

Figure 2.
PLK5 protein levels in multiple cancer tissue microarrays (TMAs). (A) Composite staining images of TMAs used in the analysis. PLK5 is stained with DAB (brown), cytokeratin (pink) is used as a tumor marker, and blue is a nuclear stain. (B) PLK5 levels in each cancer type with differential expression shown between non-malignant and malignant tissues from each TMA. LUAD: Lung adenocarcinoma; LUSC: Lung squamous cell carcinoma.
Figure 2.
PLK5 protein levels in multiple cancer tissue microarrays (TMAs). (A) Composite staining images of TMAs used in the analysis. PLK5 is stained with DAB (brown), cytokeratin (pink) is used as a tumor marker, and blue is a nuclear stain. (B) PLK5 levels in each cancer type with differential expression shown between non-malignant and malignant tissues from each TMA. LUAD: Lung adenocarcinoma; LUSC: Lung squamous cell carcinoma.

Figure 3.
PLK5 expression in normal tissues and cancers using GTEx and TCGA databases. (A) Expression of PLK5 in same tissues as were analyzed in TMAs. (B) Pan-cancer analysis of PLK5 expression. Green text means PLK5 is down regulated in tumors (q < 0.05), the black means PLK5 is not down regulated in tumors. Blue lines and boxes are normal tissues and red are cancer tissues. ACC: Adrenocortical carcinoma; BLCA: Bladder urothelial carcinoma; BRCA: Breast invasive carcinoma; CESC: Cervical squamous cell carcinoma and endocervical adenocarcinoma; COAD: Colon adenocarcinoma; DLBC: Lymphoid neoplasm diffuse large B-cell lymphoma; ESCA: Esophageal carcinoma; GBM: Glioblastoma multiforme; HNSC: Head and neck squamous cell carcinoma; KICH: Kidney chromophobe; KIRC: Kidney renal clear cell carcinoma; KIRP: Kidney renal papillary cell carcinoma; LAML: Acute myeloid leukemia; LGG: Brain lower grade glioma; LIHC: Liver hepatocellular carcinoma; LUAD: Lung adenocarcinoma; LUSC: Lung squamous cell carcinoma; OV: Ovarian serous cystadenocarcinoma; PAAD: Pancreatic adenocarcinoma; PCPG: Pheochromocytoma and paraganglioma; PRAD: Prostate adenocarcinoma; READ: Rectum adenocarcinoma; SKCM: Skin cutaneous melanoma; STAD: Stomach adenocarcinoma; TGCT: Testicular germ cell tumors; THCA: Thyroid carcinoma; THYM: Thymoma; UCEC: Uterine corpus endometrial carcinoma; UCS: Uterine carcinosarcoma.
Figure 3.
PLK5 expression in normal tissues and cancers using GTEx and TCGA databases. (A) Expression of PLK5 in same tissues as were analyzed in TMAs. (B) Pan-cancer analysis of PLK5 expression. Green text means PLK5 is down regulated in tumors (q < 0.05), the black means PLK5 is not down regulated in tumors. Blue lines and boxes are normal tissues and red are cancer tissues. ACC: Adrenocortical carcinoma; BLCA: Bladder urothelial carcinoma; BRCA: Breast invasive carcinoma; CESC: Cervical squamous cell carcinoma and endocervical adenocarcinoma; COAD: Colon adenocarcinoma; DLBC: Lymphoid neoplasm diffuse large B-cell lymphoma; ESCA: Esophageal carcinoma; GBM: Glioblastoma multiforme; HNSC: Head and neck squamous cell carcinoma; KICH: Kidney chromophobe; KIRC: Kidney renal clear cell carcinoma; KIRP: Kidney renal papillary cell carcinoma; LAML: Acute myeloid leukemia; LGG: Brain lower grade glioma; LIHC: Liver hepatocellular carcinoma; LUAD: Lung adenocarcinoma; LUSC: Lung squamous cell carcinoma; OV: Ovarian serous cystadenocarcinoma; PAAD: Pancreatic adenocarcinoma; PCPG: Pheochromocytoma and paraganglioma; PRAD: Prostate adenocarcinoma; READ: Rectum adenocarcinoma; SKCM: Skin cutaneous melanoma; STAD: Stomach adenocarcinoma; TGCT: Testicular germ cell tumors; THCA: Thyroid carcinoma; THYM: Thymoma; UCEC: Uterine corpus endometrial carcinoma; UCS: Uterine carcinosarcoma.

Figure 4.
Analysis of stage-wise PLK5 expression. (A) Analysis of PLK5 protein levels in TMAs by tumor stage. (B) Analysis of TCGA data for PLK5 expression by tumor stages of the same tumor types as in TMA. Note that TCGA does not have data for fallopian tube tumors, and therefore, it was excluded. LUAD: Lung adenocarcinoma; LUSC: Lung squamous cell carcinoma; CESC: Cervical squamous cell carcinoma and endocervical adenocarcinoma; OV: Ovarian serous cystadenocarcinoma; TGCT: Testicular germ cell tumors; UCEC: Uterine corpus endometrial carcinoma.
Figure 4.
Analysis of stage-wise PLK5 expression. (A) Analysis of PLK5 protein levels in TMAs by tumor stage. (B) Analysis of TCGA data for PLK5 expression by tumor stages of the same tumor types as in TMA. Note that TCGA does not have data for fallopian tube tumors, and therefore, it was excluded. LUAD: Lung adenocarcinoma; LUSC: Lung squamous cell carcinoma; CESC: Cervical squamous cell carcinoma and endocervical adenocarcinoma; OV: Ovarian serous cystadenocarcinoma; TGCT: Testicular germ cell tumors; UCEC: Uterine corpus endometrial carcinoma.

Figure 5.
Survival analysis of cancers according to PLK5 expression using the TCGA database. (A) PLK5 hazard ratio plot for overall survival in the same tissues as were analyzed in TMAs. The error bar denotes 95% confidence interval. (B) PLK5 hazard ratio plot for progression-free survival in same tissues as were analyzed in TMAs. The error bar denotes 95% confidence interval. (C) Kaplan–Meier plots comparing the overall survival between PLK5-High and -Low patients. (D) Kaplan–Meier plots comparing progression-free survival curves between PLK5-High and -Low patients. For (C,D), red color is PLK-High patients, while blue color is PLK-Low patients. CESC: Cervical squamous cell carcinoma and endocervical adenocarcinoma; LUAD: Lung adenocarcinoma; LUSC: Lung squamous cell carcinoma; OV: Ovarian serous cystadenocarcinoma; TGCT: Testicular germ cell tumors; UCEC: Uterine corpus endometrial carcinoma.
Figure 5.
Survival analysis of cancers according to PLK5 expression using the TCGA database. (A) PLK5 hazard ratio plot for overall survival in the same tissues as were analyzed in TMAs. The error bar denotes 95% confidence interval. (B) PLK5 hazard ratio plot for progression-free survival in same tissues as were analyzed in TMAs. The error bar denotes 95% confidence interval. (C) Kaplan–Meier plots comparing the overall survival between PLK5-High and -Low patients. (D) Kaplan–Meier plots comparing progression-free survival curves between PLK5-High and -Low patients. For (C,D), red color is PLK-High patients, while blue color is PLK-Low patients. CESC: Cervical squamous cell carcinoma and endocervical adenocarcinoma; LUAD: Lung adenocarcinoma; LUSC: Lung squamous cell carcinoma; OV: Ovarian serous cystadenocarcinoma; TGCT: Testicular germ cell tumors; UCEC: Uterine corpus endometrial carcinoma.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Su, S.; Ndiaye, M.A.; Guzmán-Pérez, G.; Baus, R.M.; Huang, W.; Patankar, M.S.; Ahmad, N. Potential Tumor Suppressor Role of Polo-like Kinase 5 in Cancer. Cancers 2023, 15, 5457. https://doi.org/10.3390/cancers15225457
AMA Style
Su S, Ndiaye MA, Guzmán-Pérez G, Baus RM, Huang W, Patankar MS, Ahmad N. Potential Tumor Suppressor Role of Polo-like Kinase 5 in Cancer. Cancers. 2023; 15(22):5457. https://doi.org/10.3390/cancers15225457
Chicago/Turabian StyleSu, Shengqin, Mary Ann Ndiaye, Glorimar Guzmán-Pérez, Rebecca Michael Baus, Wei Huang, Manish Suresh Patankar, and Nihal Ahmad. 2023. "Potential Tumor Suppressor Role of Polo-like Kinase 5 in Cancer" Cancers 15, no. 22: 5457. https://doi.org/10.3390/cancers15225457
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.