
Invasion-Associated Reorganization of Laminin 332 in Oral Squamous Cell Carcinomas: The Role of the Laminin γ2 Chain in Tumor Biology, Diagnosis, and Therapy

Abstract
:Simple Summary
Abstract
1. Introduction
2. ECM Reorganization in the OSCC Invasive Front
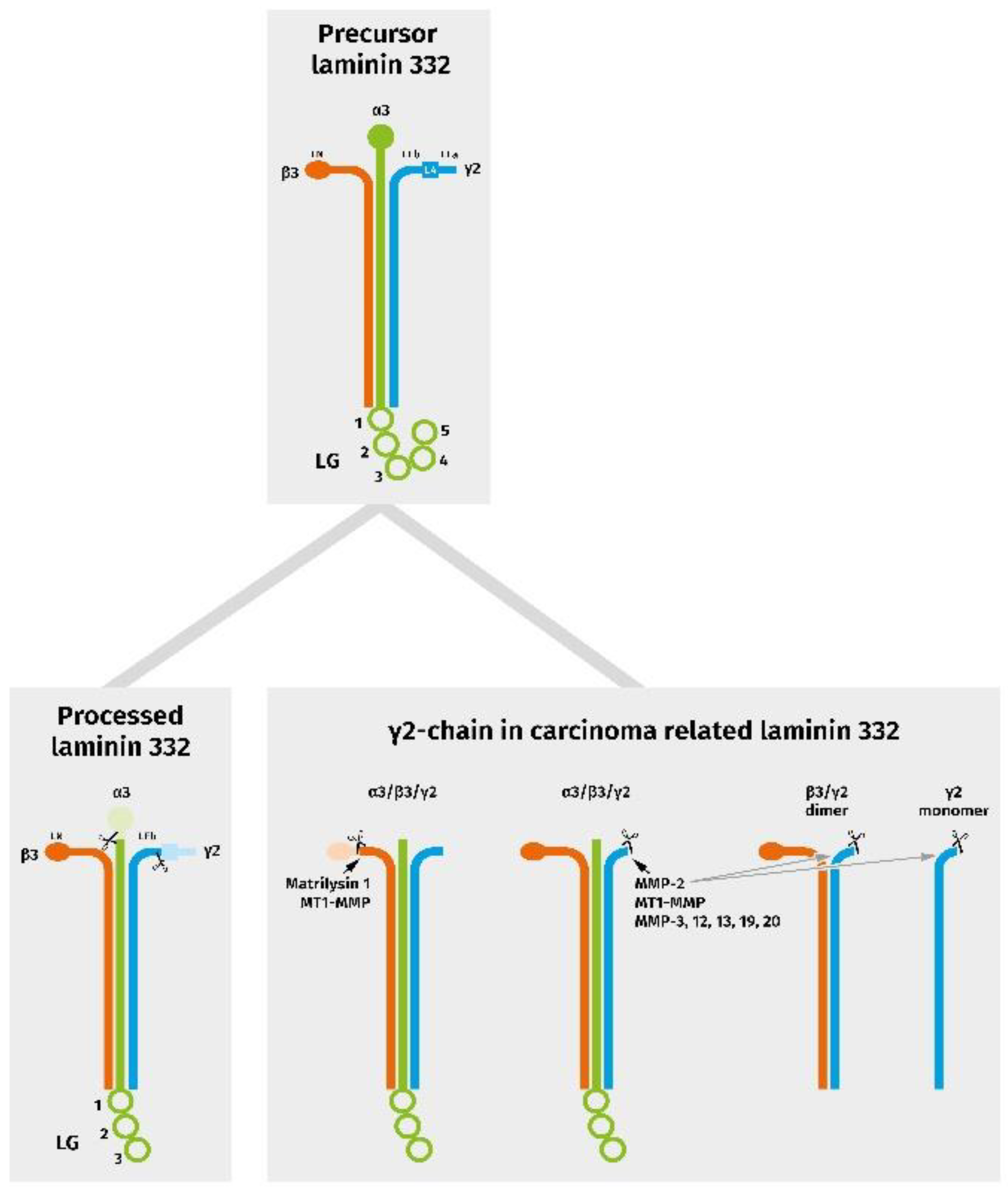
3. Laminin and Laminin Isoforms
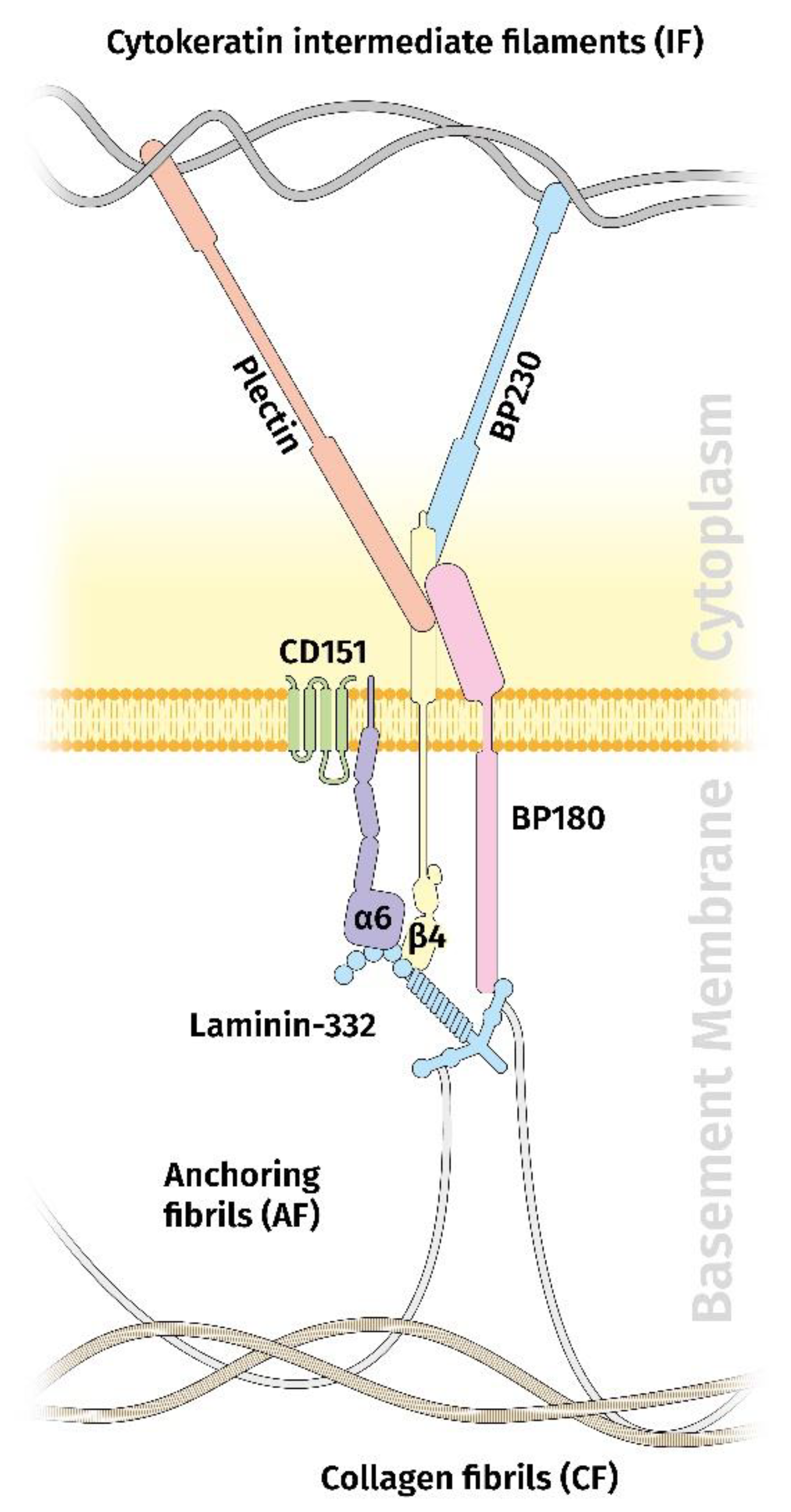
4. Ln332 and Hemidesmosomes
5. Laminin Reorganization in OSCCs
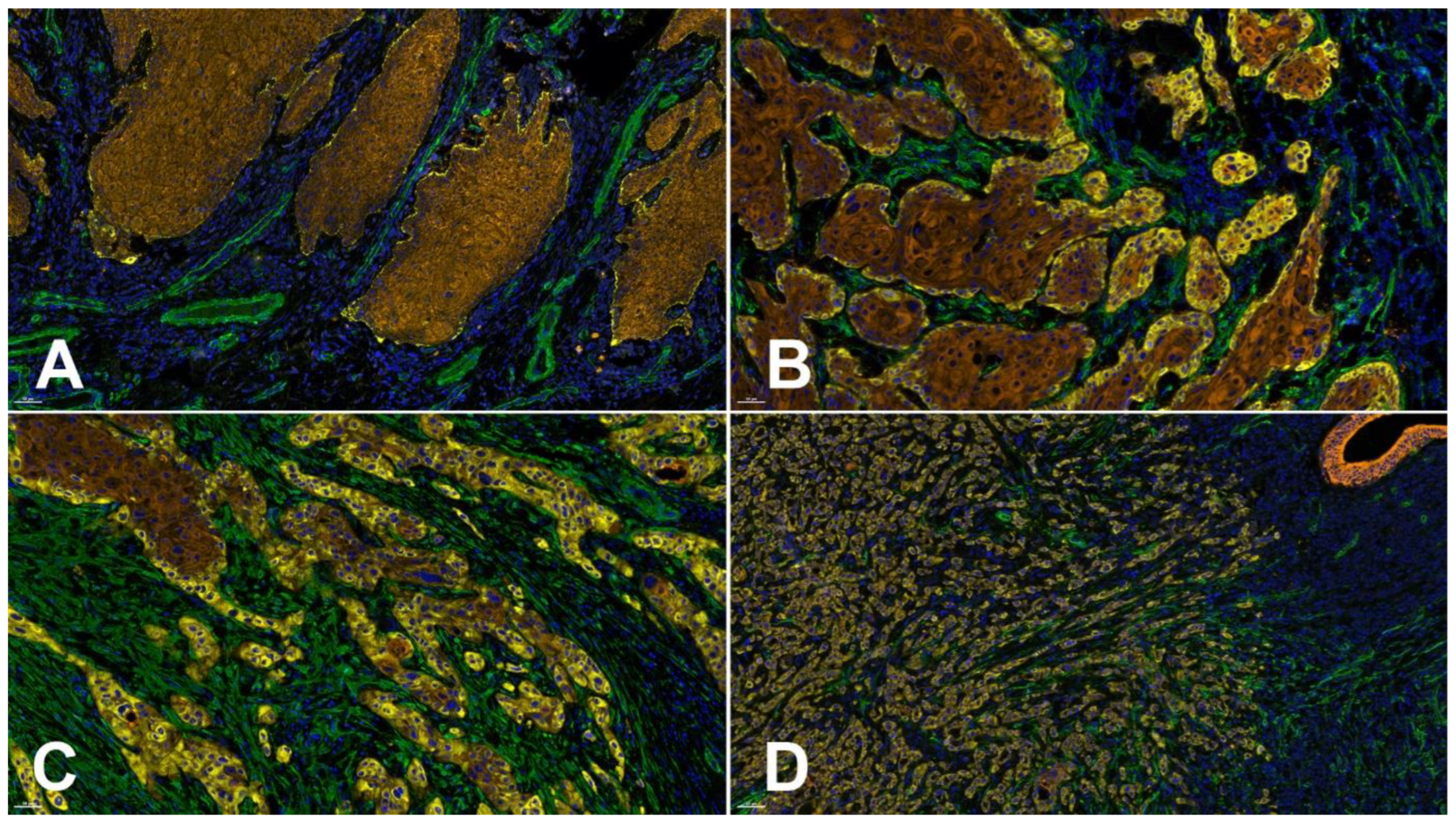
6. Laminin γ2 Chain Reorganization in the OSCC Invasive Front: Tumor Biological Implications
6.1. Laminin γ2 Chain Expression Is Related to the Migration of Normal and Neoplastic Keratinocytes
6.2. Proteolytic Processing of Laminin γ2 Generates Migration Promoting Matrikines
6.3. Modulation of Laminin γ2 Expression Pattern Is a Result of Tumor–Stroma Cross-Talk and EMT
6.4. Laminin γ2 Interacts with Oncofetal Fibronectin and Tenascin-C
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 3D | 3-dimensional |
| aSMA | alpha smooth muscle actin |
| BM | basement membrane |
| CAF | cancer-associated fibroblast |
| ECM | extracellular matrix |
| EDA-Fn | EDA domain containing fibronectin |
| EDB-Fn | EDB domain containing fibronectin |
| EGF | epidermal growth factor |
| EGFR | epidermal growth factor receptor |
| EMT | Epithelial–mesenchymal transition |
| Fn | fibronectin |
| FNIII | fibronectin type III-like repeats |
| HD | hemidesmosome |
| HNSCC | head and neck squamous cell carcinoma |
| Ln | laminin |
| Ln332 | laminin 332 |
| Ln5 | laminin 5 |
| Lng2 | laminin gamma 2 chain |
| MMP | matrix metalloproteinase |
| mRNA | messenger RNA |
| myCAF | cancer-associated fibroblast with a myofibroblast phenotype |
| oncf | oncofetal |
| oncfECM | oncofetal extracellular matrix |
| oncfFn | oncofetal fibronectin |
| oncfTn-C | oncofetal tenascin-C |
| OSCC | oral squamous cell carcinoma |
| RNA | ribonucleic acid |
| TGFβ1 | transforming growth factor beta 1 |
| TIMP | tissue inhibitor of metalloproteinases |
| TME | tumor microenvironment |
| Tn-C | tenascin-C |
References
- Observatory, T.G.C. GLOBOCAN 2020/Lip, Oral Cavity. 2022. Available online: https://gco.iarc.fr/today/data/factsheets/cancers/1-Lip-oral-cavity-fact-sheet.pdf (accessed on 1 September 2022).
- Coletta, R.D.; Yeudall, W.A.; Salo, T. Grand Challenges in Oral Cancers. Front. Oral Health 2020, 1, 3. [Google Scholar] [CrossRef]
- Ling, Z.; Cheng, B.; Tao, X. Epithelial-to-mesenchymal transition in oral squamous cell carcinoma: Challenges and opportunities. Int. J. Cancer 2021, 148, 1548–1561. [Google Scholar] [CrossRef]
- Savagner, P. Epithelial-mesenchymal transitions: From cell plasticity to concept elasticity. Curr. Top. Dev. Biol. 2015, 112, 273–300. [Google Scholar] [CrossRef]
- Van den Bossche, V.; Zaryouh, H.; Vara-Messler, M.; Vignau, J.; Machiels, J.P.; Wouters, A.; Schmitz, S.; Corbet, C. Microenvironment-driven intratumoral heterogeneity in head and neck cancers: Clinical challenges and opportunities for precision medicine. Drug Resist. Updates 2022, 60, 100806. [Google Scholar] [CrossRef]
- Qin, Y.; Zheng, X.; Gao, W.; Wang, B.; Wu, Y. Tumor microenvironment and immune-related therapies of head and neck squamous cell carcinoma. Mol. Ther. Oncolytics 2021, 20, 342–351. [Google Scholar] [CrossRef]
- Kaspar, M.; Zardi, L.; Neri, D. Fibronectin as target for tumor therapy. Int. J. Cancer 2006, 118, 1331–1339. [Google Scholar] [CrossRef] [PubMed]
- Friedl, P.; Alexander, S. Cancer invasion and the microenvironment: Plasticity and reciprocity. Cell 2011, 147, 992–1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Te Boekhorst, V.; Friedl, P. Plasticity of Cancer Cell Invasion-Mechanisms and Implications for Therapy. Adv. Cancer Res. 2016, 132, 209–264. [Google Scholar] [CrossRef]
- Mughees, M.; Sengupta, A.; Khowal, S.; Wajid, S. Mechanism of tumour microenvironment in the progression and development of oral cancer. Mol. Biol. Rep. 2021, 48, 1773–1786. [Google Scholar] [CrossRef]
- Nisar, S.; Yousuf, P.; Masoodi, T.; Wani, N.A.; Hashem, S.; Singh, M.; Sageena, G.; Mishra, D.; Kumar, R.; Haris, M.; et al. Chemokine-Cytokine Networks in the Head and Neck Tumor Microenvironment. Int. J. Mol. Sci. 2021, 22, 4584. [Google Scholar] [CrossRef] [PubMed]
- Koontongkaew, S. The tumor microenvironment contribution to development, growth, invasion and metastasis of head and neck squamous cell carcinomas. J. Cancer 2013, 4, 66–83. [Google Scholar] [CrossRef] [PubMed]
- Pakshir, P.; Noskovicova, N.; Lodyga, M.; Son, D.O.; Schuster, R.; Goodwin, A.; Karvonen, H.; Hinz, B. The myofibroblast at a glance. J. Cell Sci. 2020, 133, jcs227900. [Google Scholar] [CrossRef]
- Rasanen, K.; Vaheri, A. Activation of fibroblasts in cancer stroma. Exp. Cell Res. 2010, 316, 2713–2722. [Google Scholar] [CrossRef] [PubMed]
- Shiga, K.; Hara, M.; Nagasaki, T.; Sato, T.; Takahashi, H.; Takeyama, H. Cancer-Associated Fibroblasts: Their Characteristics and Their Roles in Tumor Growth. Cancers 2015, 7, 2443–2458. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, M.; Gibbons, D.L.; Zong, C.; Fradette, J.J.; Bota-Rabassedas, N.; Kurie, J.M. Fibroblast heterogeneity and its impact on extracellular matrix and immune landscape remodeling in cancer. Matrix Biol. 2020, 91–92, 8–18. [Google Scholar] [CrossRef]
- Augsten, M. Cancer-associated fibroblasts as another polarized cell type of the tumor microenvironment. Front. Oncol. 2014, 4, 62. [Google Scholar] [CrossRef]
- Bienkowska, K.J.; Hanley, C.J.; Thomas, G.J. Cancer-Associated Fibroblasts in Oral Cancer: A Current Perspective on Function and Potential for Therapeutic Targeting. Front. Oral Health 2021, 2, 686337. [Google Scholar] [CrossRef]
- Mezawa, Y.; Orimo, A. Phenotypic heterogeneity, stability and plasticity in tumor-promoting carcinoma-associated fibroblasts. FEBS J. 2022, 289, 2429–2447. [Google Scholar] [CrossRef]
- Tanis, T.; Cincin, Z.B.; Gokcen-Rohlig, B.; Bireller, E.S.; Ulusan, M.; Tanyel, C.R.; Cakmakoglu, B. The role of components of the extracellular matrix and inflammation on oral squamous cell carcinoma metastasis. Arch. Oral Biol. 2014, 59, 1155–1163. [Google Scholar] [CrossRef] [PubMed]
- Patankar, S.R.; Wankhedkar, D.P.; Tripathi, N.S.; Bhatia, S.N.; Sridharan, G. Extracellular matrix in oral squamous cell carcinoma: Friend or foe? Indian J. Dent. Res. 2016, 27, 184–189. [Google Scholar] [CrossRef]
- Eble, J.A.; Niland, S. The extracellular matrix in tumor progression and metastasis. Clin. Exp. Metastasis 2019, 36, 171–198. [Google Scholar] [CrossRef]
- Kosmehl, H.; Berndt, A.; Katenkamp, D. Molecular variants of fibronectin and laminin: Structure, physiological occurrence and histopathological aspects. Virchows Arch. 1996, 429, 311–322. [Google Scholar] [CrossRef]
- Kosmehl, H.; Berndt, A.; Strassburger, S.; Borsi, L.; Rousselle, P.; Mandel, U.; Hyckel, P.; Zardi, L.; Katenkamp, D. Distribution of laminin and fibronectin isoforms in oral mucosa and oral squamous cell carcinoma. Br. J. Cancer 1999, 81, 1071–1079. [Google Scholar] [CrossRef] [Green Version]
- Kosmehl, H.; Berndt, A.; Katenkamp, D.; Mandel, U.; Bohle, R.; Gabler, U.; Celeda, D. Differential expression of fibronectin splice variants, oncofetal glycosylated fibronectin and laminin isoforms in nodular palmar fibromatosis. Pathol. Res. Pract. 1995, 191, 1105–1113. [Google Scholar] [CrossRef]
- Franz, M.; Grun, K.; Richter, P.; Brehm, B.R.; Fritzenwanger, M.; Hekmat, K.; Neri, D.; Gummert, J.; Figulla, H.R.; Kosmehl, H.; et al. Extra cellular matrix remodelling after heterotopic rat heart transplantation: Gene expression profiling and involvement of ED-A+ fibronectin, alpha-smooth muscle actin and B+ tenascin-C in chronic cardiac allograft rejection. Histochem. Cell Biol. 2010, 134, 503–517. [Google Scholar] [CrossRef]
- Franz, M.; Jung, C.; Lauten, A.; Figulla, H.R.; Berndt, A. Tenascin-C in cardiovascular remodeling: Potential impact for diagnosis, prognosis estimation and targeted therapy. Cell Adhes. Migr. 2015, 9, 90–95. [Google Scholar] [CrossRef] [Green Version]
- Imanaka-Yoshida, K.; Tawara, I.; Yoshida, T. Tenascin-C in cardiac disease: A sophisticated controller of inflammation, repair, and fibrosis. Am. J. Physiol. Cell Physiol. 2020, 319, C781–C796. [Google Scholar] [CrossRef] [PubMed]
- Barker, T.H.; Engler, A.J. The provisional matrix: Setting the stage for tissue repair outcomes. Matrix Biol. 2017, 60–61, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyons, A.J.; Bateman, A.C.; Spedding, A.; Primrose, J.N.; Mandel, U. Oncofetal fibronectin and oral squamous cell carcinoma. Br. J. Oral Maxillofac. Surg. 2001, 39, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, H.; Hakomori, S. The oncofetal domain of fibronectin defined by monoclonal antibody FDC-6: Its presence in fibronectins from fetal and tumor tissues and its absence in those from normal adult tissues and plasma. Proc. Natl. Acad. Sci. USA 1985, 82, 6517–6521. [Google Scholar] [CrossRef]
- Matsuura, H.; Takio, K.; Titani, K.; Greene, T.; Levery, S.B.; Salyan, M.E.; Hakomori, S. The oncofetal structure of human fibronectin defined by monoclonal antibody FDC-6. Unique structural requirement for the antigenic specificity provided by a glycosylhexapeptide. J. Biol. Chem. 1988, 263, 3314–3322. [Google Scholar] [CrossRef]
- Ziober, A.F.; Falls, E.M.; Ziober, B.L. The extracellular matrix in oral squamous cell carcinoma: Friend or foe? Head Neck 2006, 28, 740–749. [Google Scholar] [CrossRef]
- Fromme, J.E.; Zigrino, P. The Role of Extracellular Matrix Remodeling in Skin Tumor Progression and Therapeutic Resistance. Front. Mol. Biosci. 2022, 9, 864302. [Google Scholar] [CrossRef] [PubMed]
- Lyons, A.J.; Jones, J. Cell adhesion molecules, the extracellular matrix and oral squamous carcinoma. Int. J. Oral Maxillofac. Surg. 2007, 36, 671–679. [Google Scholar] [CrossRef]
- Berndt, A.; Hyckel, P.; Konneker, A.; Katenkamp, D.; Kosmehl, H. Oral squamous cell carcinoma invasion is associated with a laminin-5 matrix re-organization but independent of basement membrane and hemidesmosome formation. clues from an in vitro invasion model. Invasion Metastasis 1997, 17, 251–258. [Google Scholar]
- Berndt, A.; Borsi, L.; Hyckel, P.; Kosmehl, H. Fibrillary co-deposition of laminin-5 and large unspliced tenascin-C in the invasive front of oral squamous cell carcinoma in vivo and in vitro. J. Cancer Res. Clin. Oncol. 2001, 127, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Meireles Da Costa, N.; Mendes, F.A.; Pontes, B.; Nasciutti, L.E.; Ribeiro Pinto, L.F.; Palumbo Junior, A. Potential Therapeutic Significance of Laminin in Head and Neck Squamous Carcinomas. Cancers 2021, 13, 1890. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wei, J.R.; Dong, J.; Lin, Q.G.; Tang, H.; Jia, Y.X.; Tan, W.; Chen, Q.Y.; Zeng, T.T.; Xing, S.; et al. Laminin gamma2-mediating T cell exclusion attenuates response to anti-PD-1 therapy. Sci. Adv. 2021, 7, eabc8346. [Google Scholar] [CrossRef]
- Kariya, Y.; Mori, T.; Yasuda, C.; Watanabe, N.; Kaneko, Y.; Nakashima, Y.; Ogawa, T.; Miyazaki, K. Localization of laminin alpha3B chain in vascular and epithelial basement membranes of normal human tissues and its down-regulation in skin cancers. J. Mol. Histol. 2008, 39, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Aumailley, M. Laminins and interaction partners in the architecture of the basement membrane at the dermal-epidermal junction. Exp. Dermatol. 2021, 30, 17–24. [Google Scholar] [CrossRef]
- Burgeson, R.E.; Chiquet, M.; Deutzmann, R.; Ekblom, P.; Engel, J.; Kleinman, H.; Martin, G.R.; Meneguzzi, G.; Paulsson, M.; Sanes, J.; et al. A new nomenclature for the laminins. Matrix Biol. 1994, 14, 209–211. [Google Scholar] [CrossRef]
- Aumailley, M.; Bruckner-Tuderman, L.; Carter, W.G.; Deutzmann, R.; Edgar, D.; Ekblom, P.; Engel, J.; Engvall, E.; Hohenester, E.; Jones, J.C.; et al. A simplified laminin nomenclature. Matrix Biol. 2005, 24, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Timpl, R.; Rohde, H.; Robey, P.G.; Rennard, S.I.; Foidart, J.M.; Martin, G.R. Laminin—A glycoprotein from basement membranes. J. Biol. Chem. 1979, 254, 9933–9937. [Google Scholar] [CrossRef]
- Aumailley, M. The laminin family. Cell Adhes. Migr. 2013, 7, 48–55. [Google Scholar] [CrossRef] [Green Version]
- Colognato, H.; Yurchenco, P.D. Form and function: The laminin family of heterotrimers. Dev. Dyn. 2000, 218, 213–234. [Google Scholar] [CrossRef]
- Walko, G.; Castanon, M.J.; Wiche, G. Molecular architecture and function of the hemidesmosome. Cell Tissue Res. 2015, 360, 529–544. [Google Scholar] [CrossRef] [Green Version]
- Te Molder, L.; de Pereda, J.M.; Sonnenberg, A. Regulation of hemidesmosome dynamics and cell signaling by integrin alpha6beta4. J. Cell Sci. 2021, 134, jcs259004. [Google Scholar] [CrossRef]
- Litjens, S.H.; de Pereda, J.M.; Sonnenberg, A. Current insights into the formation and breakdown of hemidesmosomes. Trends Cell Biol. 2006, 16, 376–383. [Google Scholar] [CrossRef] [PubMed]
- Guess, C.M.; Quaranta, V. Defining the role of laminin-332 in carcinoma. Matrix Biol. 2009, 28, 445–455. [Google Scholar] [CrossRef] [Green Version]
- Rousselle, P.; Scoazec, J.Y. Laminin 332 in cancer: When the extracellular matrix turns signals from cell anchorage to cell movement. Semin. Cancer Biol. 2020, 62, 149–165. [Google Scholar] [CrossRef] [PubMed]
- Marinkovich, M.P. Tumour microenvironment: Laminin 332 in squamous-cell carcinoma. Nat. Rev. Cancer 2007, 7, 370–380. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.R.; Silverman, S., Jr.; Daniels, T.E.; Kramer, R.H.; Greenspan, J.S. Distribution of fibronectin and laminin in oral leukoplakia and carcinoma. J. Oral Pathol. 1985, 14, 247–255. [Google Scholar] [CrossRef]
- Noguchi, M.; Kohama, G.; Hiratsuka, H.; Miyakawa, A.; Yamaguchi, A.; Nagai, I.; Kyogoku, J.; Odajima, T. Immunohistochemical localization of laminin and its relation to the grade of histological malignancy in oral cancer. Gan No Rinsho 1989, 35, 880–885. [Google Scholar] [PubMed]
- Tosios, K.I.; Kapranos, N.; Papanicolaou, S.I. Loss of basement membrane components laminin and type IV collagen parallels the progression of oral epithelial neoplasia. Histopathology 1998, 33, 261–268. [Google Scholar] [CrossRef]
- Kumagai, S.; Kojima, S.; Imai, K.; Nakagawa, K.; Yamamoto, E.; Kawahara, E.; Nakanishi, I. Immunohistologic distribution of basement membrane in oral squamous cell carcinoma. Head Neck 1994, 16, 51–57. [Google Scholar] [CrossRef]
- Shinohara, M.; Nakamura, S.; Harada, T.; Shimada, M.; Oka, M. Mode of tumor invasion in oral squamous cell carcinoma: Improved grading based on immunohistochemical examination of extracellular matrices. Head Neck 1996, 18, 153–159. [Google Scholar] [CrossRef]
- Yellapurkar, S.; Natarajan, S.; Boaz, K.; Manaktala, N.; Baliga, M.; Shetty, P.; Prasad, M.; Ravi, M. Expression of Laminin in Oral Squamous Cell Carcinomas. Asian Pac. J. Cancer Prev. 2018, 19, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Harada, T.; Shinohara, M.; Nakamura, S.; Oka, M. An immunohistochemical study of the extracellular matrix in oral squamous cell carcinoma and its association with invasive and metastatic potential. Virchows Arch. 1994, 424, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Domloge-Hultsch, N.; Gammon, W.R.; Briggaman, R.A.; Gil, S.G.; Carter, W.G.; Yancey, K.B. Epiligrin, the major human keratinocyte integrin ligand, is a target in both an acquired autoimmune and an inherited subepidermal blistering skin disease. J. Clin. Investig. 1992, 90, 1628–1633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kainulainen, T.; Autio-Harmainen, H.; Oikarinen, A.; Salo, S.; Tryggvason, K.; Salo, T. Altered distribution and synthesis of laminin-5 (kalinin) in oral lichen planus, epithelial dysplasias and squamous cell carcinomas. Br. J. Dermatol. 1997, 136, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Haas, K.M.; Berndt, A.; Stiller, K.J.; Hyckel, P.; Kosmehl, H. A comparative quantitative analysis of laminin-5 in the basement membrane of normal, hyperplastic, and malignant oral mucosa by confocal immunofluorescence imaging. J. Histochem. Cytochem. 2001, 49, 1261–1268. [Google Scholar] [CrossRef]
- Matsumoto, K.; Horikoshi, M.; Rikimaru, K.; Enomoto, S. A study of an in vitro model for invasion of oral squamous cell carcinoma. J. Oral Pathol. Med. 1989, 18, 498–501. [Google Scholar] [CrossRef] [PubMed]
- Kulasekara, K.K.; Lukandu, O.M.; Neppelberg, E.; Vintermyr, O.K.; Johannessen, A.C.; Costea, D.E. Cancer progression is associated with increased expression of basement membrane proteins in three-dimensional in vitro models of human oral cancer. Arch. Oral Biol. 2009, 54, 924–931. [Google Scholar] [CrossRef] [PubMed]
- Franz, M.; Wolheim, A.; Richter, P.; Umbreit, C.; Dahse, R.; Driemel, O.; Hyckel, P.; Virtanen, I.; Kosmehl, H.; Berndt, A. Stromal laminin chain distribution in normal, hyperplastic and malignant oral mucosa: Relation to myofibroblast occurrence and vessel formation. J. Oral Pathol. Med. 2010, 39, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Ono, Y.; Nakanishi, Y.; Ino, Y.; Niki, T.; Yamada, T.; Yoshimura, K.; Saikawa, M.; Nakajima, T.; Hirohashi, S. Clinocopathologic significance of laminin-5 gamma2 chain expression in squamous cell carcinoma of the tongue: Immunohistochemical analysis of 67 lesions. Cancer 1999, 85, 2315–2321. [Google Scholar] [CrossRef]
- Kuratomi, Y.; Kumamoto, M.; Kidera, K.; Toh, S.; Masuda, M.; Nakashima, T.; Inokuchi, A. Diffuse expression of laminin gamma2 chain in disseminating and infiltrating cancer cells indicates a highly malignant state in advanced tongue cancer. Oral Oncol. 2006, 42, 73–76. [Google Scholar] [CrossRef]
- Thangaraj, S.V.; Shyamsundar, V.; Krishnamurthy, A.; Ramshankar, V. Deregulation of extracellular matrix modeling with molecular prognostic markers revealed by transcriptome sequencing and validations in Oral Tongue squamous cell carcinoma. Sci. Rep. 2021, 11, 250. [Google Scholar] [CrossRef]
- Xin, Z.; Yamaguchi, A.; Sakamoto, K. Aberrant expression and altered cellular localization of desmosomal and hemidesmosomal proteins are associated with aggressive clinicopathological features of oral squamous cell carcinoma. Virchows Arch. 2014, 465, 35–47. [Google Scholar] [CrossRef]
- Gasparoni, A.; Della Casa, M.; Milillo, L.; Lorenzini, G.; Rubini, C.; Urso, R.; Lo Muzio, L. Prognostic value of differential expression of Laminin-5 gamma2 in oral squamous cell carcinomas: Correlation with survival. Oncol. Rep. 2007, 18, 793–800. [Google Scholar]
- Nordemar, S.; Hogmo, A.; Lindholm, J.; Auer, G.; Munck-Wikland, E. Laminin-5 gamma 2: A marker to identify oral mucosal lesions at risk for tumor development? Anticancer Res. 2003, 23, 4985–4989. [Google Scholar]
- Silva, E.M.R.; Freitas, V.M.; Bautz, W.G.; de Barros, L.A.P.; da Gama de Souza, L.N. Immunohistochemical Study of Laminin-332 gamma2 Chain and MMP-9 in High Risk of Malignant Transformation Oral Lesions and OSCC. J. Oral Maxillofac. Res. 2018, 9, e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanaruddin, S.N.; Saleh, A.; Yang, Y.H.; Hamid, S.; Mustafa, W.M.; Khairul Bariah, A.A.; Zain, R.B.; Lau, S.H.; Cheong, S.C. Four-protein signature accurately predicts lymph node metastasis and survival in oral squamous cell carcinoma. Hum. Pathol. 2013, 44, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Kawano, K.; Yanagisawa, S. Predictive value of laminin-5 and membrane type 1-matrix metalloproteinase expression for cervical lymph node metastasis in T1 and T2 squamous cell carcinomas of the tongue and floor of the mouth. Head Neck 2006, 28, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Pyke, C.; Romer, J.; Kallunki, P.; Lund, L.R.; Ralfkiaer, E.; Dano, K.; Tryggvason, K. The gamma 2 chain of kalinin/laminin 5 is preferentially expressed in invading malignant cells in human cancers. Am. J. Pathol. 1994, 145, 782–791. [Google Scholar]
- Pyke, C.; Salo, S.; Ralfkiaer, E.; Romer, J.; Dano, K.; Tryggvason, K. Laminin-5 is a marker of invading cancer cells in some human carcinomas and is coexpressed with the receptor for urokinase plasminogen activator in budding cancer cells in colon adenocarcinomas. Cancer Res. 1995, 55, 4132–4139. [Google Scholar]
- Driemel, O.; Dahse, R.; Hakim, S.G.; Tsioutsias, T.; Pistner, H.; Reichert, T.E.; Kosmehl, H. Laminin-5 immunocytochemistry: A new tool for identifying dysplastic cells in oral brush biopsies. Cytopathology 2007, 18, 348–355. [Google Scholar] [CrossRef]
- Zargaran, M.; Eshghyar, N.; Vaziri, P.B.; Mortazavi, H. Immunohistochemical evaluation of type IV collagen and laminin-332 gamma2 chain expression in well-differentiated oral squamous cell carcinoma and oral verrucous carcinoma: A new recommended cut-off. J. Oral Pathol. Med. 2011, 40, 167–173. [Google Scholar] [CrossRef]
- Marangon Junior, H.; Rocha, V.N.; Leite, C.F.; de Aguiar, M.C.; Souza, P.E.; Horta, M.C. Laminin-5 gamma 2 chain expression is associated with intensity of tumor budding and density of stromal myofibroblasts in oral squamous cell carcinoma. J. Oral Pathol. Med. 2014, 43, 199–204. [Google Scholar] [CrossRef]
- Kuratomi, Y.; Sato, S.; Monji, M.; Shimazu, R.; Tanaka, G.; Yokogawa, K.; Inoue, A.; Inokuchi, A.; Katayama, M. Serum concentrations of laminin gamma2 fragments in patients with head and neck squamous cell carcinoma. Head Neck 2008, 30, 1058–1063. [Google Scholar] [CrossRef]
- Masaoka, T.; Hashimoto, S.; Kinumatsu, T.; Muramatsu, T.; Jung, H.S.; Yamada, S.; Shimono, M. Immunolocalization of laminin and integrin in regenerating junctional epithelium of mice after gingivectomy. J. Periodontal Res. 2009, 44, 489–495. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.C.; Gordon, M.K.; Gerecke, D.R. Expression of Laminin 332 in Vesicant Skin Injury and Wound Repair. Clin. Dermatol. 2018, 2, 115. [Google Scholar]
- Rousselle, P.; Braye, F.; Dayan, G. Re-epithelialization of adult skin wounds: Cellular mechanisms and therapeutic strategies. Adv. Drug Deliv. Rev. 2019, 146, 344–365. [Google Scholar] [CrossRef] [PubMed]
- Rousselle, P.; Montmasson, M.; Garnier, C. Extracellular matrix contribution to skin wound re-epithelialization. Matrix Biol. 2019, 75–76, 12–26. [Google Scholar] [CrossRef] [PubMed]
- Sugisawa, M.; Masaoka, T.; Enokiya, Y.; Muramatsu, T.; Hashimoto, S.; Yamada, S.; Shimono, M. Expression and function of laminin and integrins on adhesion/migration of primary culture cells derived from rat oral epithelium. J. Periodontal Res. 2010, 45, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Choma, D.P.; Milano, V.; Pumiglia, K.M.; DiPersio, C.M. Integrin alpha3beta1-dependent activation of FAK/Src regulates Rac1-mediated keratinocyte polarization on laminin-5. J. Investig. Dermatol. 2007, 127, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Choma, D.P.; Pumiglia, K.; DiPersio, C.M. Integrin alpha3beta1 directs the stabilization of a polarized lamellipodium in epithelial cells through activation of Rac1. J. Cell Sci. 2004, 117, 3947–3959. [Google Scholar] [CrossRef]
- Decline, F.; Rousselle, P. Keratinocyte migration requires alpha2beta1 integrin-mediated interaction with the laminin 5 gamma2 chain. J. Cell Sci. 2001, 114, 811–823. [Google Scholar] [CrossRef]
- Lu, W.; Miyazaki, K.; Mizushima, H.; Nemoto, N. Immunohistochemical distribution of laminin-5 gamma2 chain and its developmental change in human embryonic and foetal tissues. Histochem. J. 2001, 33, 629–637. [Google Scholar] [CrossRef]
- Pei, Y.F.; Liu, J.; Cheng, J.; Wu, W.D.; Liu, X.Q. Silencing of LAMC2 Reverses Epithelial-Mesenchymal Transition and Inhibits Angiogenesis in Cholangiocarcinoma via Inactivation of the Epidermal Growth Factor Receptor Signaling Pathway. Am. J. Pathol. 2019, 189, 1637–1653. [Google Scholar] [CrossRef]
- Huang, C.; Chen, J. Laminin332 mediates proliferation, apoptosis, invasion, migration and epithelialtomesenchymal transition in pancreatic ductal adenocarcinoma. Mol. Med. Rep. 2021, 23, 11. [Google Scholar] [CrossRef]
- Kirtonia, A.; Pandey, A.K.; Ramachandran, B.; Mishra, D.P.; Dawson, D.W.; Sethi, G.; Ganesan, T.S.; Koeffler, H.P.; Garg, M. Overexpression of laminin-5 gamma-2 promotes tumorigenesis of pancreatic ductal adenocarcinoma through EGFR/ERK1/2/AKT/mTOR cascade. Cell Mol. Life. Sci. 2022, 79, 362. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.M.; Yao, Y.L.; Liu, W.; Shen, X.M.; Shi, L.J.; Wu, L. MicroRNA-134 inhibits tumor stem cell migration and invasion in oral squamous cell carcinomas via downregulation of PI3K-Akt signaling pathway by inhibiting LAMC2 expression. Cancer Biomark. 2020, 29, 51–67. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Shi, W.; Chen, M. Long non-coding RNA BBOX1-antisense RNA 1 enhances cell proliferation and migration and suppresses apoptosis in oral squamous cell carcinoma via the miR-3940-3p/laminin subunit gamma 2 axis. Bioengineered 2022, 13, 11138–11153. [Google Scholar] [CrossRef]
- Richter, P.; Bohmer, F.D.; Hindermann, W.; Borsi, L.; Hyckel, P.; Schleier, P.; Katenkamp, D.; Kosmehl, H.; Berndt, A. Analysis of activated EGFR signalling pathways and their relation to laminin-5 gamma2 chain expression in oral squamous cell carcinoma (OSCC). Histochem. Cell Biol. 2005, 124, 151–160. [Google Scholar] [CrossRef]
- Fukai, Y.; Masuda, N.; Kato, H.; Fukuchi, M.; Miyazaki, T.; Nakajima, M.; Sohda, M.; Kuwano, H.; Nakajima, T. Correlation between laminin-5 gamma2 chain and epidermal growth factor receptor expression in esophageal squamous cell carcinomas. Oncology 2005, 69, 71–80. [Google Scholar] [CrossRef]
- Richter, P.; Umbreit, C.; Franz, M.; Berndt, A.; Grimm, S.; Uecker, A.; Bohmer, F.D.; Kosmehl, H.; Berndt, A. EGF/TGFbeta1 co-stimulation of oral squamous cell carcinoma cells causes an epithelial-mesenchymal transition cell phenotype expressing laminin 332. J. Oral Pathol. Med. 2011, 40, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Ono, Y.; Nakanishi, Y.; Gotoh, M.; Sakamoto, M.; Hirohashi, S. Epidermal growth factor receptor gene amplification is correlated with laminin-5 gamma2 chain expression in oral squamous cell carcinoma cell lines. Cancer Lett. 2002, 175, 197–204. [Google Scholar] [CrossRef]
- Franz, M.; Richter, P.; Geyer, C.; Hansen, T.; Acuna, L.D.; Hyckel, P.; Bohmer, F.D.; Kosmehl, H.; Berndt, A. Mesenchymal cells contribute to the synthesis and deposition of the laminin-5 gamma2 chain in the invasive front of oral squamous cell carcinoma. J. Mol. Histol. 2007, 38, 183–190. [Google Scholar] [CrossRef]
- Takkunen, M.; Grenman, R.; Hukkanen, M.; Korhonen, M.; Garcia de Herreros, A.; Virtanen, I. Snail-dependent and -independent epithelial-mesenchymal transition in oral squamous carcinoma cells. J. Histochem. Cytochem. 2006, 54, 1263–1275. [Google Scholar] [CrossRef]
- Niland, S.; Riscanevo, A.X.; Eble, J.A. Matrix Metalloproteinases Shape the Tumor Microenvironment in Cancer Progression. Int. J. Mol. Sci. 2021, 23, 146. [Google Scholar] [CrossRef]
- Giannelli, G.; Falk-Marzillier, J.; Schiraldi, O.; Stetler-Stevenson, W.G.; Quaranta, V. Induction of cell migration by matrix metalloprotease-2 cleavage of laminin-5. Science 1997, 277, 225–228. [Google Scholar] [CrossRef] [PubMed]
- Koshikawa, N.; Giannelli, G.; Cirulli, V.; Miyazaki, K.; Quaranta, V. Role of cell surface metalloprotease MT1-MMP in epithelial cell migration over laminin-5. J. Cell Biol. 2000, 148, 615–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koshikawa, N.; Minegishi, T.; Sharabi, A.; Quaranta, V.; Seiki, M. Membrane-type matrix metalloproteinase-1 (MT1-MMP) is a processing enzyme for human laminin gamma 2 chain. J. Biol. Chem. 2005, 280, 88–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.C.; Wang, J.D.; Chang, H.Y.; Zhou, P.; Hahn, R.A.; Gordon, M.K.; Laskin, J.D.; Gerecke, D.R. Expression of Laminin gamma2 Proteolytic Fragments in Murine Skin Following Exposure to Sulfur Mustard. Anat. Rec. 2020, 303, 1642–1652. [Google Scholar] [CrossRef]
- Udayakumar, T.S.; Chen, M.L.; Bair, E.L.; Von Bredow, D.C.; Cress, A.E.; Nagle, R.B.; Bowden, G.T. Membrane type-1-matrix metalloproteinase expressed by prostate carcinoma cells cleaves human laminin-5 beta3 chain and induces cell migration. Cancer Res. 2003, 63, 2292–2299. [Google Scholar]
- Remy, L.; Trespeuch, C.; Bachy, S.; Scoazec, J.Y.; Rousselle, P. Matrilysin 1 influences colon carcinoma cell migration by cleavage of the laminin-5 beta3 chain. Cancer Res. 2006, 66, 11228–11237. [Google Scholar] [CrossRef] [Green Version]
- Oku, N.; Sasabe, E.; Ueta, E.; Yamamoto, T.; Osaki, T. Tight junction protein claudin-1 enhances the invasive activity of oral squamous cell carcinoma cells by promoting cleavage of laminin-5 gamma2 chain via matrix metalloproteinase (MMP)-2 and membrane-type MMP-1. Cancer Res. 2006, 66, 5251–5257. [Google Scholar] [CrossRef]
- Munshi, H.G.; Wu, Y.I.; Ariztia, E.V.; Stack, M.S. Calcium regulation of matrix metalloproteinase-mediated migration in oral squamous cell carcinoma cells. J. Biol. Chem. 2002, 277, 41480–41488. [Google Scholar] [CrossRef] [Green Version]
- Kurahara, S.; Shinohara, M.; Ikebe, T.; Nakamura, S.; Beppu, M.; Hiraki, A.; Takeuchi, H.; Shirasuna, K. Expression of MMPS, MT-MMP, and TIMPs in squamous cell carcinoma of the oral cavity: Correlations with tumor invasion and metastasis. Head Neck 1999, 21, 627–638. [Google Scholar] [CrossRef]
- Myoung, H.; Kim, M.J.; Hong, S.D.; Lee, J.I.; Lim, C.Y.; Hong, S.P. Expression of membrane type I-matrix metalloproteinase in oral squamous cell carcinoma. Cancer Lett. 2002, 185, 201–209. [Google Scholar] [CrossRef]
- Rosenthal, E.L.; Matrisian, L.M. Matrix metalloproteases in head and neck cancer. Head Neck 2006, 28, 639–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogura, Y.; Matsunaga, Y.; Nishiyama, T.; Amano, S. Plasmin induces degradation and dysfunction of laminin 332 (laminin 5) and impaired assembly of basement membrane at the dermal-epidermal junction. Br. J. Dermatol. 2008, 159, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Karamanos, N.K.; Theocharis, A.D.; Piperigkou, Z.; Manou, D.; Passi, A.; Skandalis, S.S.; Vynios, D.H.; Orian-Rousseau, V.; Ricard-Blum, S.; Schmelzer, C.E.H.; et al. A guide to the composition and functions of the extracellular matrix. FEBS J. 2021, 288, 6850–6912. [Google Scholar] [CrossRef]
- Walker, C.; Mojares, E.; Del Rio Hernandez, A. Role of Extracellular Matrix in Development and Cancer Progression. Int. J. Mol. Sci. 2018, 19, 3028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, K.T.; Lamb, P.; Deng, J.S. Matrikines and matricryptins: Implications for cutaneous cancers and skin repair. J. Dermatol. Sci. 2005, 40, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Swindle, C.S.; Tran, K.T.; Johnson, T.D.; Banerjee, P.; Mayes, A.M.; Griffith, L.; Wells, A. Epidermal growth factor (EGF)-like repeats of human tenascin-C as ligands for EGF receptor. J. Cell Biol. 2001, 154, 459–468. [Google Scholar] [CrossRef] [Green Version]
- Schenk, S.; Hintermann, E.; Bilban, M.; Koshikawa, N.; Hojilla, C.; Khokha, R.; Quaranta, V. Binding to EGF receptor of a laminin-5 EGF-like fragment liberated during MMP-dependent mammary gland involution. J. Cell Biol. 2003, 161, 197–209. [Google Scholar] [CrossRef]
- Fullar, A.; Kovalszky, I.; Bitsche, M.; Romani, A.; Schartinger, V.H.; Sprinzl, G.M.; Riechelmann, H.; Dudas, J. Tumor cell and carcinoma-associated fibroblast interaction regulates matrix metalloproteinases and their inhibitors in oral squamous cell carcinoma. Exp. Cell Res. 2012, 318, 1517–1527. [Google Scholar] [CrossRef] [Green Version]
- Buttner, R.; Berndt, A.; Valkova, C.; Richter, P.; Korn, A.; Kosan, C.; Liebmann, C. Myofibroblasts have an impact on expression, dimerization and signaling of different ErbB receptors in OSCC cells. J. Recept. Signal Transduct. Res. 2017, 37, 25–37. [Google Scholar] [CrossRef]
- Wang, S.H.; Liou, G.G.; Liu, S.H.; Chang, J.S.; Hsiao, J.R.; Yen, Y.C.; Chen, Y.L.; Wu, W.L.; Chang, J.Y.; Chen, Y.W. Laminin gamma2-enriched extracellular vesicles of oral squamous cell carcinoma cells enhance in vitro lymphangiogenesis via integrin alpha3-dependent uptake by lymphatic endothelial cells. Int. J. Cancer 2019, 144, 2795–2810. [Google Scholar] [CrossRef]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Luksic, I.; Suton, P.; Manojlovic, S.; Virag, M.; Petrovecki, M.; Macan, D. Significance of myofibroblast appearance in squamous cell carcinoma of the oral cavity on the occurrence of occult regional metastases, distant metastases, and survival. Int. J. Oral Maxillofac. Surg. 2015, 44, 1075–1080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dourado, M.R.; Guerra, E.N.S.; Salo, T.; Lambert, D.W.; Coletta, R.D. Prognostic value of the immunohistochemical detection of cancer-associated fibroblasts in oral cancer: A systematic review and meta-analysis. J. Oral Pathol. Med. 2018, 47, 443–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharjee, K.; Girish, H.C.; Murgod, S.; Alshame, A.M.J.; Shyamala, K.; Nayak, V.N. A Comparative Immunohistochemical Study of Presence and Distribution Pattern of Stromal Myofibroblast in Oral Dysplasia and in Different Grades of Oral Squamous Cell Carcinoma. J. Int. Soc. Prev. Community Dent. 2018, 8, 451–456. [Google Scholar] [CrossRef]
- Zhou, B.; Chen, W.L.; Wang, Y.Y.; Lin, Z.Y.; Zhang, D.M.; Fan, S.; Li, J.S. A role for cancer-associated fibroblasts in inducing the epithelial-to-mesenchymal transition in human tongue squamous cell carcinoma. J. Oral Pathol. Med. 2014, 43, 585–592. [Google Scholar] [CrossRef]
- Berndt, A.; Buttner, R.; Guhne, S.; Gleinig, A.; Richter, P.; Chen, Y.; Franz, M.; Liebmann, C. Effects of activated fibroblasts on phenotype modulation, EGFR signalling and cell cycle regulation in OSCC cells. Exp. Cell Res. 2014, 322, 402–414. [Google Scholar] [CrossRef]
- Ding, L.; Zhang, Z.; Shang, D.; Cheng, J.; Yuan, H.; Wu, Y.; Song, X.; Jiang, H. alpha-Smooth muscle actin-positive myofibroblasts, in association with epithelial-mesenchymal transition and lymphogenesis, is a critical prognostic parameter in patients with oral tongue squamous cell carcinoma. J. Oral Pathol. Med. 2014, 43, 335–343. [Google Scholar] [CrossRef]
- Zlobec, I.; Bachli, M.; Galuppini, F.; Berger, M.D.; Dawson, H.E.; Nagtegaal, I.D.; Lugli, A. Refining the ITBCC tumor budding scoring system with a “zero-budding” category in colorectal cancer. Virchows Arch. 2021, 479, 1085–1090. [Google Scholar] [CrossRef]
- Wahab, A.; Onkamo, O.; Pirinen, M.; Almangush, A.; Salo, T. The budding and depth of invasion model in oral cancer: A systematic review and meta-analysis. Oral Dis. 2020, 28, 275–283. [Google Scholar] [CrossRef]
- Almangush, A.; Salo, T.; Hagstrom, J.; Leivo, I. Tumour budding in head and neck squamous cell carcinoma–a systematic review. Histopathology 2014, 65, 587–594. [Google Scholar] [CrossRef]
- Almangush, A.; Pirinen, M.; Heikkinen, I.; Makitie, A.A.; Salo, T.; Leivo, I. Tumour budding in oral squamous cell carcinoma: A meta-analysis. Br. J. Cancer 2018, 118, 577–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okado, Y.; Aoki, M.; Hamasaki, M.; Koga, K.; Sueta, T.; Shiratsuchi, H.; Oda, Y.; Nakagawa, T.; Nabeshima, K. Tumor budding and laminin5-gamma2 in squamous cell carcinoma of the external auditory canal are associated with shorter survival. Springerplus 2015, 4, 814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patten, J.; Wang, K. Fibronectin in development and wound healing. Adv. Drug Deliv. Rev. 2021, 170, 353–368. [Google Scholar] [CrossRef] [PubMed]
- Mandel, U.; Hamilton Therkildsen, M.; Reibel, J.; Sweeney, B.; Matsuura, H.; Hakomori, S.; Dabelsteen, E.; Clausen, H. Cancer-associated changes in glycosylation of fibronectin. Immunohistological localization of oncofetal fibronectin defined by monoclonal antibodies. APMIS 1992, 100, 817–826. [Google Scholar] [CrossRef]
- Berndt, A.; Borsi, L.; Luo, X.; Zardi, L.; Katenkamp, D.; Kosmehl, H. Evidence of ED-B+ fibronectin synthesis in human tissues by non-radioactive RNA in situ hybridization. Investigations on carcinoma (oral squamous cell and breast carcinoma), chronic inflammation (rheumatoid synovitis) and fibromatosis (Morbus Dupuytren). Histochem. Cell Biol. 1998, 109, 249–255. [Google Scholar] [CrossRef]
- Ramos Gde, O.; Bernardi, L.; Lauxen, I.; Sant’Ana Filho, M.; Horwitz, A.R.; Lamers, M.L. Fibronectin Modulates Cell Adhesion and Signaling to Promote Single Cell Migration of Highly Invasive Oral Squamous Cell Carcinoma. PLoS ONE 2016, 11, e0151338. [Google Scholar] [CrossRef] [Green Version]
- Midwood, K.S.; Chiquet, M.; Tucker, R.P.; Orend, G. Tenascin-C at a glance. J. Cell Sci. 2016, 129, 4321–4327. [Google Scholar] [CrossRef]
- Giblin, S.P.; Midwood, K.S. Tenascin-C: Form versus function. Cell Adhes. Migr. 2015, 9, 48–82. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, A.; Loustau, T.; Salome, N.; Poilil Surendran, S.; Li, C.; Tucker, R.P.; Izzi, V.; Lamba, R.; Koch, M.; Orend, G. Advances on the roles of tenascin-C in cancer. J. Cell Sci. 2022, 135, jcs260244. [Google Scholar] [CrossRef]
- Berndt, A.; Richter, P.; Kosmehl, H.; Franz, M. Tenascin-C and carcinoma cell invasion in oral and urinary bladder cancer. Cell Adhes. Migr. 2015, 9, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Spenle, C.; Loustau, T.; Burckel, H.; Riegel, G.; Abou Faycal, C.; Li, C.; Yilmaz, A.; Petti, L.; Steinbach, F.; Ahowesso, C.; et al. Impact of Tenascin-C on Radiotherapy in a Novel Syngeneic Oral Squamous Cell Carcinoma Model With Spontaneous Dissemination to the Lymph Nodes. Front. Immunol. 2021, 12, 636108. [Google Scholar] [CrossRef] [PubMed]
- Spenle, C.; Loustau, T.; Murdamoothoo, D.; Erne, W.; Beghelli-de la Forest Divonne, S.; Veber, R.; Petti, L.; Bourdely, P.; Morgelin, M.; Brauchle, E.M.; et al. Tenascin-C Orchestrates an Immune-Suppressive Tumor Microenvironment in Oral Squamous Cell Carcinoma. Cancer Immunol. Res. 2020, 8, 1122–1138. [Google Scholar] [CrossRef] [PubMed]
- Hindermann, W.; Berndt, A.; Borsi, L.; Luo, X.; Hyckel, P.; Katenkamp, D.; Kosmehl, H. Synthesis and protein distribution of the unspliced large tenascin-C isoform in oral squamous cell carcinoma. J. Pathol. 1999, 189, 475–480. [Google Scholar] [CrossRef]
- Ramos, D.M.; Chen, B.; Regezi, J.; Zardi, L.; Pytela, R. Tenascin-C matrix assembly in oral squamous cell carcinoma. Int. J. Cancer 1998, 75, 680–687. [Google Scholar] [CrossRef]
- Franz, M.; Hansen, T.; Richter, P.; Borsi, L.; Bohmer, F.D.; Hyckel, P.; Schleier, P.; Katenkamp, D.; Zardi, L.; Kosmehl, H.; et al. Complex formation of the laminin-5 gamma2 chain and large unspliced tenascin-C in oral squamous cell carcinoma in vitro and in situ: Implications for sequential modulation of extracellular matrix in the invasive tumor front. Histochem. Cell Biol. 2006, 126, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Franz, M.; Hansen, T.; Borsi, L.; Geier, C.; Hyckel, P.; Schleier, P.; Richter, P.; Altendorf-Hofmann, A.; Kosmehl, H.; Berndt, A. A quantitative co-localization analysis of large unspliced tenascin-C(L) and laminin-5/gamma2-chain in basement membranes of oral squamous cell carcinoma by confocal laser scanning microscopy. J. Oral Pathol. Med. 2007, 36, 6–11. [Google Scholar] [CrossRef]
- Spenle, C.; Gasser, I.; Saupe, F.; Janssen, K.P.; Arnold, C.; Klein, A.; van der Heyden, M.; Mutterer, J.; Neuville-Mechine, A.; Chenard, M.P.; et al. Spatial organization of the tenascin-C microenvironment in experimental and human cancer. Cell Adhes. Migr. 2015, 9, 4–13. [Google Scholar] [CrossRef]
- Sundquist, E.; Kauppila, J.H.; Veijola, J.; Mroueh, R.; Lehenkari, P.; Laitinen, S.; Risteli, J.; Soini, Y.; Kosma, V.M.; Sawazaki-Calone, I.; et al. Tenascin-C and fibronectin expression divide early stage tongue cancer into low- and high-risk groups. Br. J. Cancer 2017, 116, 640–648. [Google Scholar] [CrossRef] [Green Version]
- Berndt, A.; Kollner, R.; Richter, P.; Franz, M.; Voigt, A.; Berndt, A.; Borsi, L.; Giavazzi, R.; Neri, D.; Kosmehl, H. A comparative analysis of oncofetal fibronectin and tenascin-C incorporation in tumour vessels using human recombinant SIP format antibodies. Histochem. Cell Biol. 2010, 133, 467–475. [Google Scholar] [CrossRef]
- Katayama, M.; Sanzen, N.; Funakoshi, A.; Sekiguchi, K. Laminin gamma2-chain fragment in the circulation: A prognostic indicator of epithelial tumor invasion. Cancer Res. 2003, 63, 222–229. [Google Scholar]
- Noguchi, M.; Kohama, G.; Hiratsuka, H.; Sekiguchi, T. Clinical significance of laminin deposition and T-cell infiltration in oral cancer. Head Neck 1993, 15, 125–132. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Tissue Type | Method | Laminin γ2 Chain Pattern Analyzed | Relevance for Diagnosis/Prognosis | Reference |
|---|---|---|---|---|
| Dysplastic oral cells (brush biopsies) | Immunohistochemistry | Cytology, presence of positive tumor cells | Method-enhanced brush cytology with enhanced sensitivity | [77] |
| Preneoplastic oral lesions | Immunohistochemistry | Positive versus negative staining | Higher risk for tumor progression | [71] |
| Oral verrucous carcinoma versus well differentiated OSCC | Immunohistochemistry | Number of cytoplasmic positive tumor cells | >5% positive cells favors diagnosis of well-differentiated OSCCs | [78] |
| OSCC | Immunohistochemistry | Number of cytoplasmic positive tumor cells | Shorter survival period | [70] |
| OSCC | Immunohistochemistry | Immunohistochemical expression/4-point intensity scoring | Lng2: disease specific survival/4 gene signature including Lng2 predicts metastasis | [73] |
| OSCC | Genome expression profiling | Expression changes | Separation of invasive and metastatic OSCC | [20] |
| OSCC | Immunohistochemistry | Increased number of cytoplasmic positive tumor cells | Associated with tumor budding | [79] |
| OSCC | Immunohistochemistry | Extension of cytoplasmic tumor cell staining | Associated with aggressive growth patterns | [69] |
| OSCC versus high-risk oral lesions | Immunohistochemistry | Continuity of BM staining and basal versus suprabasal tumor cell staining | Associated with smoking and OSCC diagnosis | [72] |
| SCC from the tongue and floor of the mouth | Immunohistochemistry | Focal type versus diffuse type expression pattern | Independent factor for nodal metastasis | [74] |
| Tongue SCC | Immunohistochemistry | Number of cytoplasmic positive tumor cells | Decreased survival time | [66] |
| Tongue SCC | Immunohistochemistry | Peripheral versus diffuse cytoplasmic positive tumor cells | Decreased 3-year survival rate, increased cervical metastases | [67] |
| Tongue SCC | Transcriptome sequencing/Immunohistochemistry | Increased expression | Treatment failure in T1, T2/decreased disease free and overall survival | [68] |
| Head and neck SCC | Lng2 fragment ELISA | Changes in serum concentration | Monitoring clinical course and treatment results | [80] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berndt, A.; Gaßler, N.; Franz, M. Invasion-Associated Reorganization of Laminin 332 in Oral Squamous Cell Carcinomas: The Role of the Laminin γ2 Chain in Tumor Biology, Diagnosis, and Therapy. Cancers 2022, 14, 4903. https://doi.org/10.3390/cancers14194903
Berndt A, Gaßler N, Franz M. Invasion-Associated Reorganization of Laminin 332 in Oral Squamous Cell Carcinomas: The Role of the Laminin γ2 Chain in Tumor Biology, Diagnosis, and Therapy. Cancers. 2022; 14(19):4903. https://doi.org/10.3390/cancers14194903
Chicago/Turabian StyleBerndt, Alexander, Nikolaus Gaßler, and Marcus Franz. 2022. "Invasion-Associated Reorganization of Laminin 332 in Oral Squamous Cell Carcinomas: The Role of the Laminin γ2 Chain in Tumor Biology, Diagnosis, and Therapy" Cancers 14, no. 19: 4903. https://doi.org/10.3390/cancers14194903