
Combined BRCA2 and MAGEC3 Expression Predict Outcome in Advanced Ovarian Cancers
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ovarian Cancer Patients
2.2. Whole Tissue Sections
2.3. Tissue Microarray (TMA) Construction
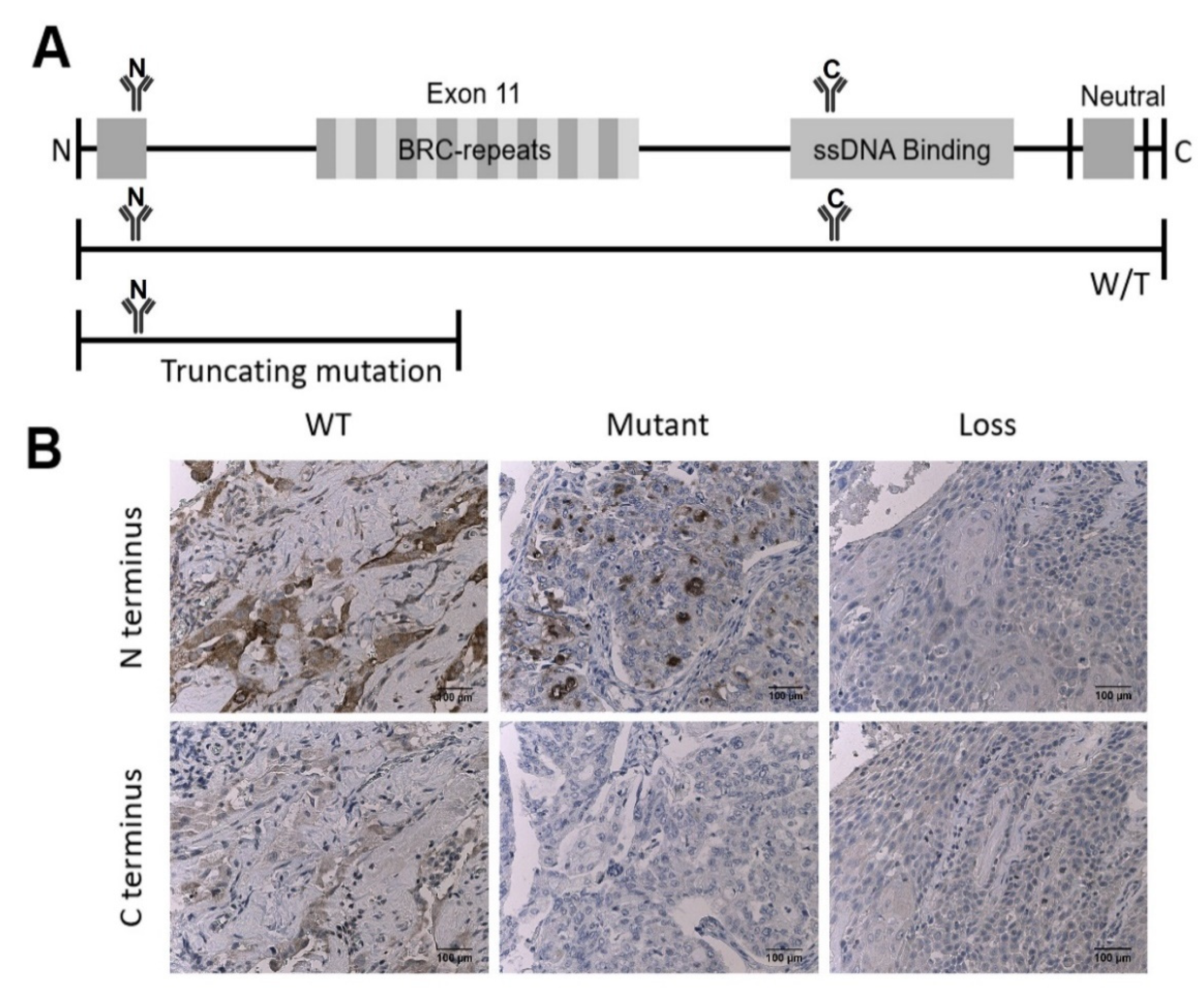
2.4. Antibody Validation
2.5. Immunohistochemistry
2.6. Digital Image Analysis
2.7. Statistical Methods
3. Results
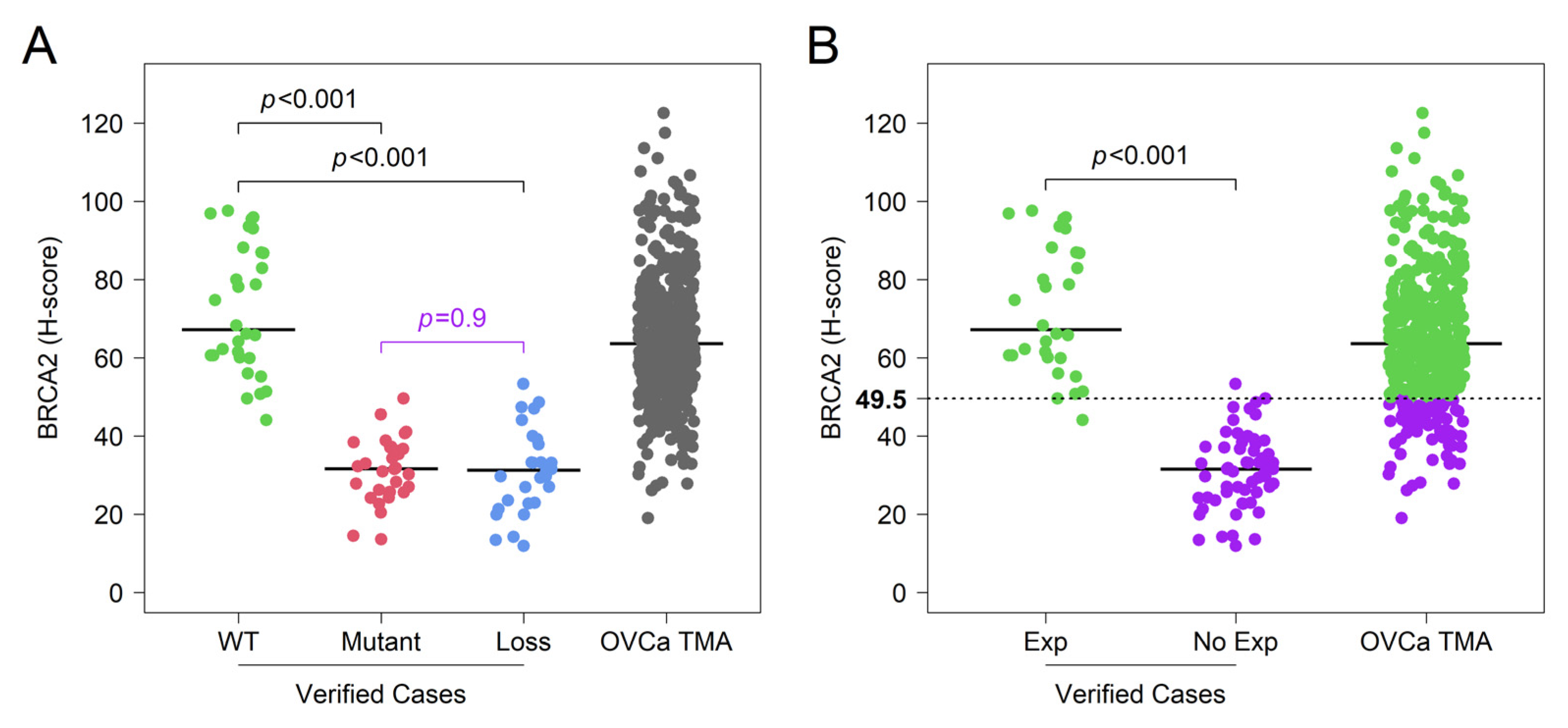
3.1. Validation of IHC Measurements of BRCA2 Protein Expression in Cancer Tissue
3.2. Clinical Characteristics of Ovarian Cancer Microarray Patient Population
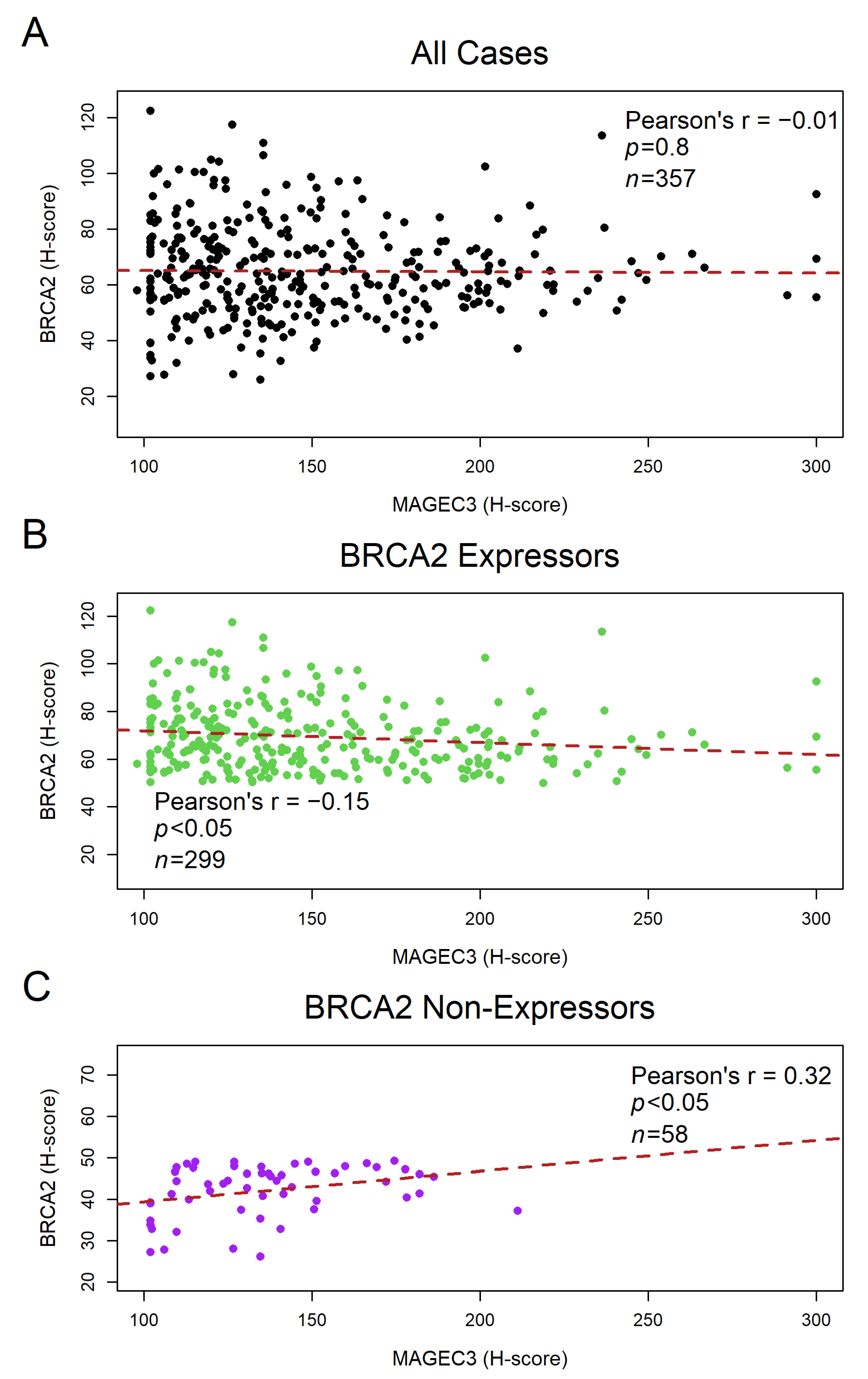
3.3. BRCA2 Protein Shows a Weak Negative Correlation with MAGEC3 Expression in Ovarian Cancer Tumor Samples
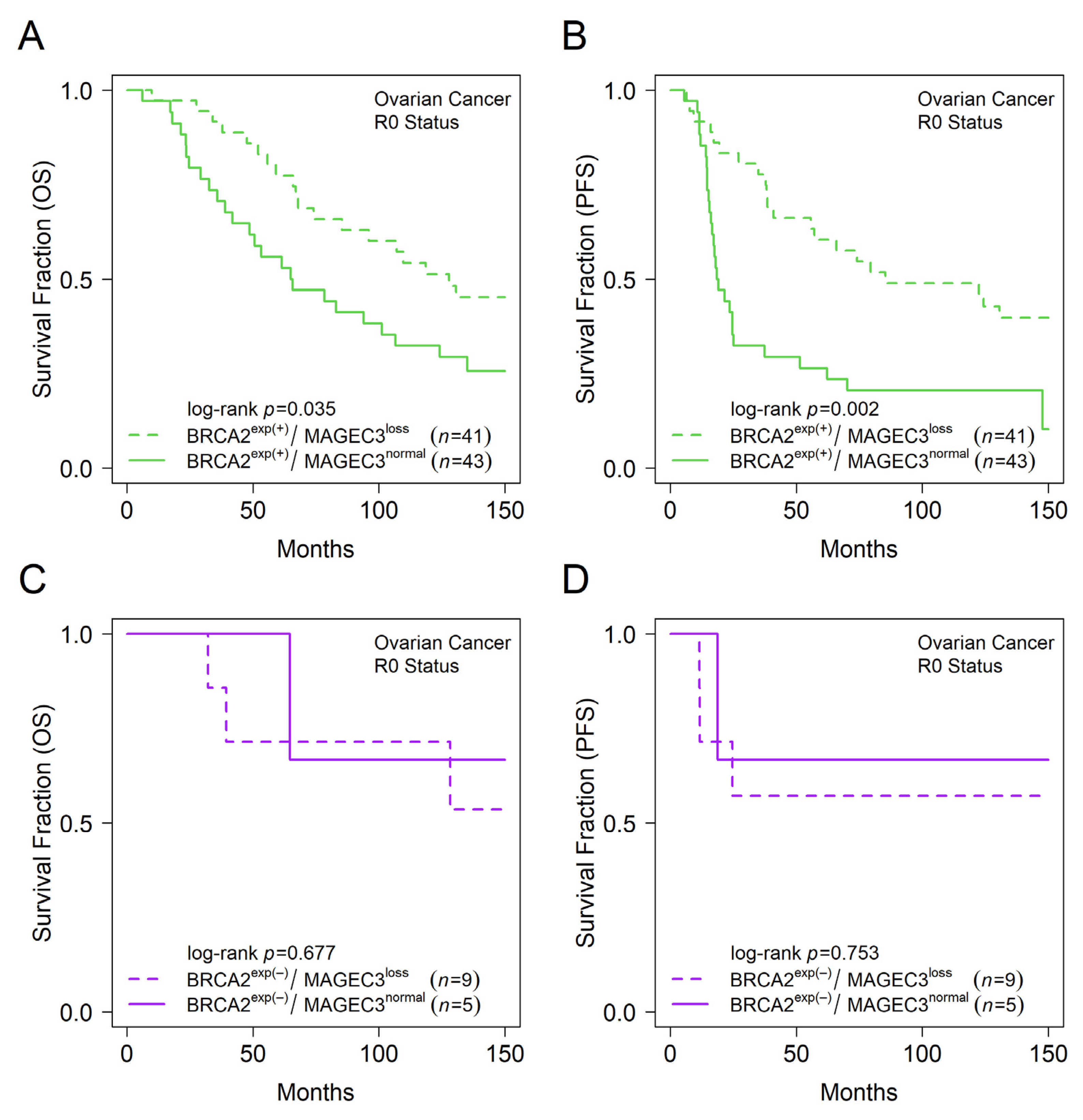
3.4. MAGEC3 and BRCA2 Association with Prognosis in Epithelial Ovarian Cancer
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Matulonis, U.A.; Sood, A.K.; Fallowfield, L.; Howitt, B.E.; Sehouli, J.; Karlan, B.Y. Ovarian cancer. Nat. Rev. Dis. Primers 2016, 2, 16061. [Google Scholar] [CrossRef] [PubMed]
- Jayson, G.C.; Kohn, E.C.; Kitchener, H.C.; Ledermann, J.A. Ovarian cancer. Lancet 2014, 384, 1376–1388. [Google Scholar] [CrossRef]
- Morgan, R.D.; Clamp, A.R.; Evans, D.G.R.; Edmondson, R.J.; Jayson, G.C. PARP inhibitors in platinum-sensitive high-grade serous ovarian cancer. Cancer Chemother. Pharmacol. 2018, 81, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Bowtell, D.D.; Bohm, S.; Ahmed, A.A.; Aspuria, P.J.; Bast, R.C., Jr.; Beral, V.; Berek, J.S.; Birrer, M.J.; Blagden, S.; Bookman, M.A.; et al. Rethinking ovarian cancer II: Reducing mortality from high-grade serous ovarian cancer. Nat. Rev. Cancer 2015, 15, 668–679. [Google Scholar] [CrossRef]
- Rendi, M.H.; Garcia, R.L.; Dizon, D.S. Epithelial Carcinoma of the Ovary, Fallopian Tube, and Peritoneum: Histopathology. Last Updated February. 2022. Available online: www.uptodate.com (accessed on 12 April 2022).
- Menon, U.; Gentry-Maharaj, A.; Burnell, M.; Singh, N.; Ryan, A.; Karpinskyj, C.; Carlino, G.; Taylor, J.; Massingham, S.K.; Raikou, M.; et al. Ovarian cancer population screening and mortality after long-term follow-up in the UK Collaborative Trial of Ovarian Cancer Screening (UKCTOCS): A randomised controlled trial. Lancet 2021, 397, 2182–2193. [Google Scholar] [CrossRef]
- Guo, T.; Dong, X.; Xie, S.; Zhang, L.; Zeng, P.; Zhang, L. Cellular Mechanism of Gene Mutations and Potential Therapeutic Targets in Ovarian Cancer. Cancer Manag. Res. 2021, 13, 3081–3100. [Google Scholar] [CrossRef]
- Badgwell, D.; Bast, R.C., Jr. Early detection of ovarian cancer. Dis. Markers 2007, 23, 397–410. [Google Scholar] [CrossRef]
- Bast, R.C., Jr.; Lu, Z.; Han, C.Y.; Lu, K.H.; Anderson, K.S.; Drescher, C.W.; Skates, S.J. Biomarkers and Strategies for Early Detection of Ovarian Cancer. Cancer Epidemiol. Biomark. Prev. 2020, 29, 2504–2512. [Google Scholar] [CrossRef]
- Ashworth, A. A synthetic lethal therapeutic approach: Poly(ADP) ribose polymerase inhibitors for the treatment of cancers deficient in DNA double-strand break repair. J. Clin. Oncol. 2008, 26, 3785–3790. [Google Scholar] [CrossRef]
- Koczkowska, M.; Zuk, M.; Gorczynski, A.; Ratajska, M.; Lewandowska, M.; Biernat, W.; Limon, J.; Wasag, B. Detection of somatic BRCA1/2 mutations in ovarian cancer-next-generation sequencing analysis of 100 cases. Cancer Med. 2016, 5, 1640–1646. [Google Scholar] [CrossRef] [PubMed]
- Girolimetti, G.; Perrone, A.M.; Santini, D.; Barbieri, E.; Guerra, F.; Ferrari, S.; Zamagni, C.; De Iaco, P.; Gasparre, G.; Turchetti, D. BRCA-associated ovarian cancer: From molecular genetics to risk management. Biomed. Res. Int. 2014, 2014, 787143. [Google Scholar] [CrossRef] [PubMed]
- Bolton, K.L.; Chenevix-Trench, G.; Goh, C.; Sadetzki, S.; Ramus, S.J.; Karlan, B.Y.; Lambrechts, D.; Despierre, E.; Barrowdale, D.; McGuffog, L.; et al. Association between BRCA1 and BRCA2 mutations and survival in women with invasive epithelial ovarian cancer. JAMA 2012, 307, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Fu, Y.; Xue, H.; Guo, K.; Song, Z.; Yu, Z.; Jia, T.; Yan, Y.; Zhao, L.; Mi, X.; et al. BRCA1 promoter hypermethylation in sporadic epithelial ovarian carcinoma: Association with low expression of BRCA1, improved survival and co-expression of DNA methyltransferases. Oncol. Lett. 2014, 7, 1088–1096. [Google Scholar] [CrossRef] [PubMed]
- Garg, K.; Levine, D.A.; Olvera, N.; Dao, F.; Bisogna, M.; Secord, A.A.; Berchuck, A.; Cerami, E.; Schultz, N.; Soslow, R.A. BRCA1 immunohistochemistry in a molecularly characterized cohort of ovarian high-grade serous carcinomas. Am. J. Surg. Pathol. 2013, 37, 138–146. [Google Scholar] [CrossRef]
- Huang, M.; Kamath, P.; Schlumbrecht, M.; Miao, F.; Driscoll, D.; Oldak, S.; Slomovitz, B.; Koru-Sengul, T.; George, S. Identifying disparities in germline and somatic testing for ovarian cancer. Gynecol. Oncol. 2019, 153, 297–303. [Google Scholar] [CrossRef]
- Wooster, R.; Neuhausen, S.L.; Mangion, J.; Quirk, Y.; Ford, D.; Collins, N.; Nguyen, K.; Seal, S.; Tran, T.; Averill, D.; et al. Localization of a breast cancer susceptibility gene, BRCA2, to chromosome 13q12–13. Science 1994, 265, 2088–2090. [Google Scholar] [CrossRef]
- Prat, J.; Ribe, A.; Gallardo, A. Hereditary ovarian cancer. Hum. Pathol. 2005, 36, 861–870. [Google Scholar] [CrossRef]
- Spain, B.H.; Larson, C.J.; Shihabuddin, L.S.; Gage, F.H.; Verma, I.M. Truncated BRCA2 is cytoplasmic: Implications for cancer-linked mutations. Proc. Natl. Acad. Sci. USA 1999, 96, 13920–13925. [Google Scholar] [CrossRef]
- Rahman, N.; Stratton, M.R. The genetics of breast cancer susceptibility. Annu. Rev. Genet. 1998, 32, 95–121. [Google Scholar] [CrossRef]
- Hakansson, S.; Johannsson, O.; Johansson, U.; Sellberg, G.; Loman, N.; Gerdes, A.M.; Holmberg, E.; Dahl, N.; Pandis, N.; Kristoffersson, U.; et al. Moderate frequency of BRCA1 and BRCA2 germ-line mutations in Scandinavian familial breast cancer. Am. J. Hum. Genet. 1997, 60, 1068–1078. [Google Scholar] [PubMed]
- Chatterjee, G.; Jimenez-Sainz, J.; Presti, T.; Nguyen, T.; Jensen, R.B. Distinct binding of BRCA2 BRC repeats to RAD51 generates differential DNA damage sensitivity. Nucleic Acids Res. 2016, 44, 5256–5270. [Google Scholar] [CrossRef] [PubMed]
- Krejci, L.; Altmannova, V.; Spirek, M.; Zhao, X. Homologous recombination and its regulation. Nucleic Acids Res. 2012, 40, 5795–5818. [Google Scholar] [CrossRef]
- Xia, B.; Sheng, Q.; Nakanishi, K.; Ohashi, A.; Wu, J.; Christ, N.; Liu, X.; Jasin, M.; Couch, F.J.; Livingston, D.M. Control of BRCA2 cellular and clinical functions by a nuclear partner, PALB2. Mol. Cell 2006, 22, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Baughan, S.; Tainsky, M.A. K3326X and Other C-Terminal BRCA2 Variants Implicated in Hereditary Cancer Syndromes: A Review. Cancers 2021, 13, 447. [Google Scholar] [CrossRef]
- Rodriguez-Freixinos, V.; Farinas-Madrid, L.; Gil-Martin, M.; Barretina-Ginesta, P.; Romeo, M.; Villacampa, G.; Pardo, B.; Ahmed, H.; Recalde, S.; Piulats, J.M.; et al. Chemotherapy and PARP inhibitors in heavily pretreated BRCA1/2 mutation ovarian cancer (BMOC) patients. Gynecol. Oncol. 2019, 152, 270–277. [Google Scholar] [CrossRef]
- Redelico, T. Rucaparib and Niraparib in Advanced Ovarian Cancer. J. Adv. Pract. Oncol. 2019, 10, 402–408. [Google Scholar] [CrossRef]
- Banerjee, S.; Kaye, S. PARP inhibitors in BRCA gene-mutated ovarian cancer and beyond. Curr. Oncol. Rep. 2011, 13, 442–449. [Google Scholar] [CrossRef]
- Rose, M.; Burgess, J.T.; O’Byrne, K.; Richard, D.J.; Bolderson, E. PARP Inhibitors: Clinical Relevance, Mechanisms of Action and Tumor Resistance. Front. Cell Dev. Biol. 2020, 8, 564601. [Google Scholar] [CrossRef]
- Moore, K.; Colombo, N.; Scambia, G.; Kim, B.G.; Oaknin, A.; Friedlander, M.; Lisyanskaya, A.; Floquet, A.; Leary, A.; Sonke, G.S.; et al. Maintenance Olaparib in Patients with Newly Diagnosed Advanced Ovarian Cancer. N. Engl. J. Med. 2018, 379, 2495–2505. [Google Scholar] [CrossRef]
- Ganguly, B.; Dolfi, S.C.; Rodriguez-Rodriguez, L.; Ganesan, S.; Hirshfield, K.M. Role of Biomarkers in the Development of PARP Inhibitors. Biomark. Cancer 2016, 8, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Swisher, E.M.; Lin, K.K.; Oza, A.M.; Scott, C.L.; Giordano, H.; Sun, J.; Konecny, G.E.; Coleman, R.L.; Tinker, A.V.; O’Malley, D.M.; et al. Rucaparib in relapsed, platinum-sensitive high-grade ovarian carcinoma (ARIEL2 Part 1): An international, multicentre, open-label, phase 2 trial. Lancet Oncol. 2017, 18, 75–87. [Google Scholar] [CrossRef]
- Mateo, J.; Carreira, S.; Sandhu, S.; Miranda, S.; Mossop, H.; Perez-Lopez, R.; Nava Rodrigues, D.; Robinson, D.; Omlin, A.; Tunariu, N.; et al. DNA-Repair Defects and Olaparib in Metastatic Prostate Cancer. N. Engl. J. Med. 2015, 373, 1697–1708. [Google Scholar] [CrossRef] [PubMed]
- Criscuolo, D.; Morra, F.; Giannella, R.; Cerrato, A.; Celetti, A. Identification of Novel Biomarkers of Homologous Recombination Defect in DNA Repair to Predict Sensitivity of Prostate Cancer Cells to PARP-Inhibitors. Int. J. Mol. Sci. 2019, 20, 3100. [Google Scholar] [CrossRef]
- Ellegate, J., Jr.; Mastri, M.; Isenhart, E.; Krolewski, J.J.; Chatta, G.; Kauffman, E.; Moffitt, M.; Eng, K.H. Loss of MAGEC3 Expression Is Associated with Prognosis in Advanced Ovarian Cancers. Cancers 2022, 14, 731. [Google Scholar] [CrossRef]
- Weon, J.L.; Potts, P.R. The MAGE protein family and cancer. Curr. Opin. Cell Biol. 2015, 37, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Pineda, C.T.; Ramanathan, S.; Fon Tacer, K.; Weon, J.L.; Potts, M.B.; Ou, Y.H.; White, M.A.; Potts, P.R. Degradation of AMPK by a cancer-specific ubiquitin ligase. Cell 2015, 160, 715–728. [Google Scholar] [CrossRef]
- Gordeeva, O.; Gordeev, A.; Khaydukov, S. Expression dynamics of Mage family genes during self-renewal and differentiation of mouse pluripotent stem and teratocarcinoma cells. Oncotarget 2019, 10, 3248–3266. [Google Scholar] [CrossRef]
- Doyle, J.M.; Gao, J.; Wang, J.; Yang, M.; Potts, P.R. MAGE-RING protein complexes comprise a family of E3 ubiquitin ligases. Mol. Cell 2010, 39, 963–974. [Google Scholar] [CrossRef]
- Hao, J.; Shen, R.; Li, Y.; Zhang, Y.; Yin, Y. Cancer-testis antigen HCA587/MAGE-C2 interacts with BS69 and promotes its degradation in the ubiquitin-proteasome pathway. Biochem. Biophys. Res. Commun. 2014, 449, 386–391. [Google Scholar] [CrossRef]
- Eng, K.H.; Szender, J.B.; Etter, J.L.; Kaur, J.; Poblete, S.; Huang, R.Y.; Zhu, Q.; Grzesik, K.A.; Battaglia, S.; Cannioto, R.; et al. Paternal lineage early onset hereditary ovarian cancers: A Familial Ovarian Cancer Registry study. PLoS Genet. 2018, 14, e1007194. [Google Scholar] [CrossRef] [PubMed]
- George, S.H.; Shaw, P. BRCA and Early Events in the Development of Serous Ovarian Cancer. Front. Oncol. 2014, 4, 5. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Jia, N.; Ma, H.; Liu, G.; Xiao, Z.; Wu, Y.; Zhou, L.; Lei, J.; Wang, L.; Liu, Y.; et al. 0D/3D coupling of g-C3N4 QDs/hierarchical macro-mesoporous CuO-SiO2 for high-efficiency norfloxacin removal in photo-Fenton-like processes. J. Hazard. Mater. 2021, 419, 126359. [Google Scholar] [CrossRef]
- Liu, Q.; Tong, D.; Liu, G.; Yi, Y.; Xu, J.; Yang, X.; Wang, L.; Zhang, J.; Ye, J.; Zhang, Y.; et al. A novel BRCA2 mutation in prostate cancer sensitive to combined radiotherapy and androgen deprivation therapy. Cancer Biol. Ther. 2018, 19, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Zhang, W.; Wang, Y.; Min, Q.; Zhang, H.; Dong, D.; Zhan, Q. MAGE-C3 promotes cancer metastasis by inducing epithelial-mesenchymal transition and immunosuppression in esophageal squamous cell carcinoma. Cancer Commun. 2021, 41, 1354–1372. [Google Scholar] [CrossRef] [PubMed]
- Daudi, S.; Eng, K.H.; Mhawech-Fauceglia, P.; Morrison, C.; Miliotto, A.; Beck, A.; Matsuzaki, J.; Tsuji, T.; Groman, A.; Gnjatic, S.; et al. Expression and immune responses to MAGE antigens predict survival in epithelial ovarian cancer. PLoS ONE 2014, 9, e104099. [Google Scholar] [CrossRef]
- Uhlen, M.; Bjorling, E.; Agaton, C.; Szigyarto, C.A.; Amini, B.; Andersen, E.; Andersson, A.C.; Angelidou, P.; Asplund, A.; Asplund, C.; et al. A human protein atlas for normal and cancer tissues based on antibody proteomics. Mol. Cell Proteom. 2005, 4, 1920–1932. [Google Scholar] [CrossRef]
- Uhlen, M.; Bandrowski, A.; Carr, S.; Edwards, A.; Ellenberg, J.; Lundberg, E.; Rimm, D.L.; Rodriguez, H.; Hiltke, T.; Snyder, M. A proposal for validation of antibodies. Nat. Methods 2016, 13, 823–827. [Google Scholar] [CrossRef]
- Varghese, F.; Bukhari, A.B.; Malhotra, R.; De, A. IHC Profiler: An open source plugin for the quantitative evaluation and automated scoring of immunohistochemistry images of human tissue samples. PLoS ONE 2014, 9, e96801. [Google Scholar] [CrossRef]
- Thiele, C.; Hirschfeld, G. Cutpointr: Improved Estimation and Validation of Optimal Cutpoints in R. J. Stat. Softw. 2021, 98, 1–27. [Google Scholar] [CrossRef]
- Huang, J.; Hu, W.; Sood, A.K. Prognostic biomarkers in ovarian cancer. Cancer Biomark. 2010, 8, 231–251. [Google Scholar] [CrossRef] [PubMed]
- Kashima, K.; Oite, T.; Aoki, Y.; Takakuwa, K.; Aida, H.; Nagata, H.; Sekine, M.; Wu, H.J.; Hirai, Y.; Wada, Y.; et al. Screening of BRCA1 mutation using immunohistochemical staining with C-terminal and N-terminal antibodies in familial ovarian cancers. Jpn. J. Cancer Res. 2000, 91, 399–409. [Google Scholar] [CrossRef]
- Vorrius, T.R.; Snyder, K.; Pica-Mendez, A.; Tan, C.; Laterza, O.; Toniatti, C.; Carpenter, C.; Lee, H.; Tanaka, W.; Zhang, Z.-Q. Immunohistochemical detection of BRCA-1 and BRCA-2 expression in human breast and ovarian tumors. J. Histotechnol. 2009, 32, 202–203. [Google Scholar] [CrossRef]
- Thorgeirsson, T.; Jordahl, K.M.; Flavin, R.; Epstein, M.M.; Fiorentino, M.; Andersson, S.-O.; Andren, O.; Rider, J.R.; Mosquera, J.M.; Ingoldsby, H. Intracellular location of BRCA2 protein expression and prostate cancer progression in the Swedish Watchful Waiting Cohort. Carcinogenesis 2016, 37, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Aijaz, I. Roswell Park Comprehensive Cancer Center: Buffalo, NY, USA, Unpublished work. 2022.
- Stordal, B.; Timms, K.; Farrelly, A.; Gallagher, D.; Busschots, S.; Renaud, M.; Thery, J.; Williams, D.; Potter, J.; Tran, T. BRCA1/2 mutation analysis in 41 ovarian cell lines reveals only one functionally deleterious BRCA1 mutation. Mol. Oncol. 2013, 7, 567–579. [Google Scholar] [CrossRef] [PubMed]
- Guirado, R.; Carceller, H.; Castillo-Gomez, E.; Castren, E.; Nacher, J. Automated analysis of images for molecular quantification in immunohistochemistry. Heliyon 2018, 4, e00669. [Google Scholar] [CrossRef]
- Vrekoussis, T.; Chaniotis, V.; Navrozoglou, I.; Dousias, V.; Pavlakis, K.; Stathopoulos, E.; Zoras, O. Image analysis of breast cancer immunohistochemistry-stained sections using ImageJ: An RGB-based model. Anticancer. Res. 2009, 29, 4995–4998. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | MAGEloss | MAGEnormal | ||||
|---|---|---|---|---|---|---|
| All Ovary Patients | BRCA2exp(−) | BRCA2exp(+) | BRCA2exp(−) | BRCA2exp(+) | p-Value | |
| n = 357 | n = 34 | n = 139 | n = 24 | n = 160 | ||
| Age of Diagnosis (years) | 0.1 † | |||||
| Mean (Range) | 63 (21–93) | 61 (21–85) | 63 (31–93) | 69 (47–89) | 63 (21–89) | |
| Missing | 0 | 0 | 0 | 0 | 0 | |
| Year of Diagnosis (n (%)) | <0.001 ‡ | |||||
| Before 2006 | 177 (52.1) | 14 (43.8) | 104 (81.9) | 5 (21.7) | 54 (34.2) | |
| After 2006 | 163 (47.9) | 18 (56.3) | 23 (18.1) | 18 (78.3) | 104 (65.8) | |
| Missing | 17 | 2 | 12 | 1 | 2 | |
| Race (n (%)) | ||||||
| White | 334 (94.9) | 31 (94) | 130 (96.3) | 24 (100) | 149 (93.1) | 0.2 § |
| Black or African American | 8 (2.3) | 1 (3) | 0 (0) | 0 (0) | 7 (4.4) | |
| Other | 10 (2.8) | 1 (3) | 5 (3.7) | 0 (0) | 4 (2.5) | |
| Missing | 5 | 1 | 4 | 0 | 0 | |
| Hispanic (n (%)) | ||||||
| No | 350 (99.4) | 34 (100) | 133 (99.3) | 24 (100) | 159 (99.4) | 0.9 § |
| Yes | 2 (0.6) | 0 (0) | 1 (0.7) | 0 (0) | 1 (0.6) | |
| Missing | 5 | 0 | 5 | 0 | 0 | |
| Primary (n (%)) | ||||||
| Ovary | 310 (88.1) | 27 (81.8) | 131 (94.9) | 16 (66.7) | 136 (86.6) | <0.001 ‡ |
| Primary Peritoneal | 42 (11.9) | 6 (18.2) | 7 (5.1) | 8 (33.3) | 21 (13.4) | |
| Missing | 5 | 1 | 1 | 0 | 3 | |
| FIGO Stage (n (%)) | 0.6 ‡ | |||||
| I/II/IIIA/B | 65 (18.5) | 8 (24.2) | 28 (20.6) | 4 (17.4) | 25 (15.7) | |
| IIIC/IV | 286 (81.5) | 25 (75.8) | 108 (79.4) | 19 (82.6) | 134 (84.3) | |
| Missing | 6 | 1 | 3 | 1 | 1 | |
| Grade (n (%)) | 0.03 ‡ | |||||
| Well/Moderately differentiated | 93 (26.5) | 5 (14.7) | 44 (32.6) | 2 (8.3) | 42 (26.6) | |
| Poorly/Undifferentiated | 258 (73.5) | 29 (85.3) | 91 (67.4) | 22 (91.7) | 116 (73.4) | |
| Missing | 6 | 0 | 4 | 0 | 2 | |
| Histology (n (%)) | ||||||
| Serous | 284 (79.6) | 17 (50) | 114 (82) | 20 (83.3) | 133 (83.1) | <0.001 ‡ |
| Other Epithelial | 73 (20.4) | 17 (50) | 25 (18) | 4 (16.7) | 27 (16.9) | |
| Missing | 0 | 0 | 0 | 0 | 0 | |
| Cytoreduction (n (%)) | 0.09 ‡ | |||||
| R0 | 98 (27.8) | 9 (28.1) | 41 (29.9) | 5 (20.8) | 43 (26.9) | |
| Not R0 | 255 (72.2) | 23 (71.9) | 96 (70.1) | 19 (79.2) | 117 (73.1) | |
| Missing | 4 | 2 | 2 | 0 | 0 | |
| Platinum sensitivity (n (%)) | ||||||
| Sensitive | 156 (55.5) | 9 (37.5) | 65 (55.1) | 11 (68.8) | 71 (57.7) | 0.8 ‡ |
| Resistant | 125 (44.5) | 15 (62.5) | 53 (44.9) | 5 (31.3) | 52 (42.3) | |
| Missing | 76 | 10 | 21 | 8 | 37 | |
| Treatment Outcome (n (%)) | ||||||
| Complete Response | 169 (57.5) | 9 (37.5) | 65 (55.1) | 9 (52.9) | 86 (63.7) | 0.2 ‡ |
| Not Complete Response | 125 (42.5) | 15 (62.5) | 53 (44.9) | 8 (47.1) | 49 (36.3) | |
| Missing | 63 | 10 | 21 | 7 | 25 | |
| Ovarian Cancer | Univariate Analysis (n = 357) † | Multivariate Analysis (n = 342) | |||||
|---|---|---|---|---|---|---|---|
| Covariate | Risk Level | Hazard Ratio | 95% CI | p-Value ‡ | Hazard Ratio | 95% CI | p-Value ‡ |
| Age | +10 years | 1.3 | (1.18–1.42) | <0.001 | 1.32 | (1.19–1.46) | <0.001 |
| Stage | I/II/IIIA/B | Reference | Reference | ||||
| IIIC/IV | 3.45 | (2.4–4.96) | <0.001 | 3.28 | (2.05–5.26) | <0.001 | |
| Grade | Well/Moderately differentiated | Reference | Reference | ||||
| Poorly/Undifferentiated | 1.15 | (0.89–1.49) | 0.3 | 1.34 | (1.02–1.76) | 0.04 | |
| Histology | Other Epithelial | Reference | Stratifier § | ||||
| Serous | 1.17 | (0.87–1.57) | 0.3 | ||||
| Cytoreduction | R0 | Reference | Stratifier § | ||||
| Not R0 | 2.43 | (1.84–3.15) | <0.001 | ||||
| MAGEC3 and | Loss~Exp(−) | Reference | Reference | ||||
| BRCA2 | Normal~Exp(−) | 1.29 | (0.85–1.95) | 0.2 | 1.14 | (0.73–1.79) | 0.6 |
| Loss~Exp(+) | 1.17 | (0.72–1.89) | 0.5 | 1 | (0.61–1.63) | 1 | |
| Normal~Exp(+) | 1.16 | (0.91–1.49) | 0.2 | 1.14 | (0.88–1.48) | 0.3 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Omole, E.B.; Aijaz, I.; Ellegate, J., Jr.; Isenhart, E.; Desouki, M.M.; Mastri, M.; Humphrey, K.; Dougherty, E.M.; Rosario, S.R.; Nastiuk, K.L.; et al. Combined BRCA2 and MAGEC3 Expression Predict Outcome in Advanced Ovarian Cancers. Cancers 2022, 14, 4724. https://doi.org/10.3390/cancers14194724
Omole EB, Aijaz I, Ellegate J Jr., Isenhart E, Desouki MM, Mastri M, Humphrey K, Dougherty EM, Rosario SR, Nastiuk KL, et al. Combined BRCA2 and MAGEC3 Expression Predict Outcome in Advanced Ovarian Cancers. Cancers. 2022; 14(19):4724. https://doi.org/10.3390/cancers14194724
Chicago/Turabian StyleOmole, Emmanuel B., Iqbal Aijaz, James Ellegate, Jr., Emily Isenhart, Mohamed M. Desouki, Michalis Mastri, Kristen Humphrey, Emily M. Dougherty, Spencer R. Rosario, Kent L. Nastiuk, and et al. 2022. "Combined BRCA2 and MAGEC3 Expression Predict Outcome in Advanced Ovarian Cancers" Cancers 14, no. 19: 4724. https://doi.org/10.3390/cancers14194724