
The Role of microRNAs in Multidrug Resistance of Glioblastoma
by
 , , and
, , and
Parvaneh Mahinfar
1,†, 
Behnaz Mansoori
1,2,† ,
,
Davoud Rostamzadeh
3,4, 
Behzad Baradaran
1 ,
,
William C. Cho
5,* and
and
Behzad Mansoori
6,* 1
Immunology Research Center, Tabriz University of Medical Sciences, Tabriz 5166/15731, Iran
2
Department of Molecular Genetics, Faculty of Biological Sciences, Tarbiat Modares University, Tehran 175-14115, Iran
3
Department of Clinical Biochemistry, Yasuj University of Medical Sciences, Yasuj 7591994799, Iran
4
Medicinal Plants Research Center, Yasuj University of Medical Sciences, Yasuj 7591994799, Iran
5
Department of Clinical Oncology, Queen Elizabeth Hospital, Kowloon, Hong Kong SAR, China
6
The Wistar Institute, Molecular & Cellular Oncogenesis Program, Philadelphia, PA 19104, USA
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Cancers 2022, 14(13), 3217; https://doi.org/10.3390/cancers14133217
Submission received: 23 May 2022
/
Revised: 20 June 2022
/
Accepted: 25 June 2022
/
Published: 30 June 2022
(This article belongs to the Special Issue Oncogenes and Tumor Suppressor Genes in Brain Tumor)
Abstract
:Simple Summary
Glioblastoma (GBM) is one of the most malignant types of central nervous system tumor which accounts for more than 60% of all brain tumors in adults. Owing to poor prognosis and drug resistance of most GBM, it is urged to further develop the diagnosis and treatment strategies. The aim of this article is to highlight the roles of some functional microRNAs in the diagnosis and treatment of drug-resistant GBM. Besides, we suggest effective treatment strategies based on the expression profiles of these effective miRNAs to provide an alternative solution to deal with this cancer.
Abstract
Glioblastoma (GBM) is an aggressive brain tumor that develops from neuroglial stem cells and represents a highly heterogeneous group of neoplasms. These tumors are predominantly correlated with a dismal prognosis and poor quality of life. In spite of major advances in developing novel and effective therapeutic strategies for patients with glioblastoma, multidrug resistance (MDR) is considered to be the major reason for treatment failure. Several mechanisms contribute to MDR in GBM, including upregulation of MDR transporters, alterations in the metabolism of drugs, dysregulation of apoptosis, defects in DNA repair, cancer stem cells, and epithelial–mesenchymal transition. MicroRNAs (miRNAs) are a large class of endogenous RNAs that participate in various cell events, including the mechanisms causing MDR in glioblastoma. In this review, we discuss the role of miRNAs in the regulation of the underlying mechanisms in MDR glioblastoma which will open up new avenues of inquiry for the treatment of glioblastoma.
1. Introduction
Glioblastoma is a malignant brain tumor which develops from neuroglial stem cells [1,2]. According to the fifth edition of the WHO classification of Central Nervous System (WHO CNS5) tumor in 2021, 14 distinct tumors have been identified. This alteration incorporates numerous molecular changes with clinicopathologic utility for the most accurate classification of CNS neoplasms. On the basis of a recent version of WHO CNS5, fundamental changes in molecular and practical approaches including histology and immunohistochemistry to CNS tumor taxonomy were added and introduced the role of molecular diagnostics in CNS tumor classification. The momentous changes in WHO CNS5 involve the classification of gliomas, differentiating gliomas that occur primarily in adults. In fact, in adults, diffuse gliomas have been categorized into three types including astrocytoma, IDH-mutant; oligodendroglioma, IDH-mutant, 1p/19q-co-deleted; and glioblastoma, IDH-wildtype. Additionally, in WHO CNS5, glioblastomas will comprise only IDH-wildtype tumors. In the new classification, all IDH-mutant diffuse astrocytic tumors are considered a single type (astrocytoma, IDH-mutant) and are graded as 2, 3, or 4 [3,4]. It is estimated that the 5-year survival rate for glioblastomas is less than 5% [3]. Although, there have been significant improvements in both the research and development for combating this type of cancer, the prognosis and long-term outlook for patients with glioblastoma remain poor [5]. Glioblastoma treatment failure is thought to be due to its anatomical location, and the presence of the blood–brain barrier which hinders the transport of chemotherapeutic agents. Commute inhibition of immune cells results in aggressive tumor cell behavior [6]. One of the main reasons why glioblastoma treatments have failed is the occurrence of multi-drug resistance (MDR) to common chemotherapy drugs. According to recent information that release through FDA-approved chemotropic drugs, there are four drugs for the treatment of glioblastoma: temozolomide (TMZ), lomustine, intravenous carmustine, and bevacizumab (BVZ). As mentioned above, only TMZ, tumor treatment fields, is approved for new diagnoses in high-grade gliomas (HGG). There is no standard of care (SOC) for these reversions. Moreover, only MDR to TMZ has been reported in glioblastoma malignancy [7,8]. According to recent studies, over 90% of cancer patients who die due to drug resistance are receiving traditional chemotherapeutics or novel targeted drugs [9]. In response to chemotherapy, a large number of tumor cells become resistant to the amount of drug administered [9,10]. It is widely acknowledged that drug resistance is a significant health issue that adversely affects cancer treatment effectiveness [11]. It has been found that a variety of molecular mechanisms are involved in the development of MDR, such as dysregulation of drug efflux proteins expression, modification of drug targets, disruption of cell cycle checkpoints, altered apoptosis, enhanced DNA damage repair in cancer stem cells, and mediators of posttranscriptional regulation, such as microRNAs (miRNAs) [12]. There are numerous biomarkers, such as biological markers, found in body fluids which assume an essential role in many aspects of oncology such as diagnosis. These biomarkers are signals of the cancer’s presence and consist of molecular alteration, proliferation, or process in the patient’s body. For instance, an alteration in the expression of several proteins may indicate the presence of cancer. The prognosis of GBM patients is poor in most cases. Therefore, utilizing some specific and sensitive biomarkers is a great approach to the diagnosis of GBM. According to the recent studies, there are some putative biomarkers of glioblastoma such as CD133, CD44, CD15, CD70, S100A4, ALDH1A3, NANOG, OCT-4, and SOX-2. Furthermore, all of these biomarkers consider MDR GBM because of fluctuations in expression, especially in glioblastoma stem cells (GSCs). GSCs make a great impact on chemotherapy resistance and cause tumor recurrence. These markers are associated with cascading pathways and interactions with some important and functional miRNAs such as miR-20a which are upregulated in MDR GBM. In addition, CCL22, ADCY2, PDK1, ZFP36, CP, CD2, PLAUR, ACAP1, COL5A1, FAM83D, PBK, FANCA, ANXA7, and TACC3 were identified as genetic biomarkers that were all associated with pathways in GBM progression and MDR. Investigation of the expression these hallmarks is one of the appropriate methods to detect cancer cells in the early phase [13,14]. Among these, in general, miRNAs are small (usually 19–24 nucleotides) non-coding RNAs that play an important role in various critical cellular processes by targeting and modulating the expression of key genes involved [15]. MiRNAs play a key regulatory role in MDR through modulating various drug resistant mechanisms that are more significant in current treatment approaches. In particular, miRNAs that are deregulated play a momentous role in glioblastoma, in which they participate in multiple cellular processes, including proliferation, apoptosis, autophagy, invasion, metastasis, and angiogenesis [16,17]. This review aimed to highlight the importance of miRNAs in the regulation of MDR-related mechanisms.
2. MDR in GBM
A major cause of chemotherapy treatments failure is MDR, the mechanism by which cancers become resistant to chemotherapy drugs. A major cause of many chemotherapy treatments failing is MDR, the mechanism by which cancers become resistant to chemotherapy drugs. Different kinds of chemotherapy agents are used to treat cancer—either alone or in combination with other agents. These agents are various in their chemical composition. The mainly chemotropic agents include antimetabolites (5-fluorouracil (5-FU)), alkylating agents (temozolomide, cisplatin), topoisomerase inhibitors (doxorubicin), and mitotic spindle inhibitors (paclitaxel) [9]. The development of MDR in GBM has been linked to numerous molecular mechanisms including upregulation of MDR transporters, changes in the metabolism of drugs, dysregulation of apoptosis, defects in DNA repair, cancer stem cells, and epithelial-mesenchymal transition (EMT).
2.1. Upregulation of MDR Transporters
Increasing the efflux of drugs in cancer cells, especially in glioblastoma cells is one of the most significant and well-known mechanisms for developing MDR [18,19]. ATP binding cassette (ABC) transporter family members are considered to be essential transmembrane proteins which play a significant role in MDR with respect to pumping chemotherapeutic agents from tumor cells into the extracellular space as a result of ATP cleavage [20,21]. Therefore, these transporters diminish the cytotoxicity of anticancer agents by reducing the intracellular concentration of drugs. Among 48 members of ABC transporter members P-glycoprotein (P-gp/ABCB1) a transmembrane glycoprotein, and multidrug resistance-associated protein-1 (MRP1/ABCC1) are two extensively investigated members in GBM MDR [22]. Various studies have shown that these proteins are involved in the development of resistance against chemotherapeutic agents in this type of cancer. Moreover, co-localization of both P-gp and ABCG2 proteins is observed in glioblastoma cells, which is correlated to their joint functioning as drug transporters [19]. Consequently, blocking or inactivating ABC transporters increases the concentration of anti-neoplastic drugs in a cell [23].
2.2. Changes in the Metabolism of Drugs
The alteration of antitumor drug metabolism is another important mechanism used by glioblastoma cancer cells to reduce the cytotoxic effects of chemotherapy. Cytochrome P450 (CYP) enzymes which are expressed at higher levels in tumors of the digestive tract, liver, kidney, and brain, play a significant role in the MDR against vincristine, taxanes, etoposide, vinblastine, ifosfamide, doxorubicin, irinotecan, and cyclophosphamide [24]. Upregulating the expression of drug-metabolizing CYP (17A1) which catalyzes the metabolism of cholesterol to neurosteroids in GBM, causes MDR during treatment [25,26]. Many factors that affect CYP activities, including genetic polymorphisms, diseases, medications, certain foods, physiological conditions, and smoking, can alter pharmacokinetic profiles which are affecting chemotherapeutic efficacy in glioblastoma tumor cells [27].
2.3. Dysregulation of Apoptosis
As a consequence of chemotherapeutic-induced DNA damage, cancer cells can undergo two biological events, including cell cycle arrest and DNA repair, or apoptosis and cell death. During this process, TP53 plays a fundamental role, particularly during GBM MDR [28,29]. TP53 mutations in GBM mainly point to mutations that promote the development of MDR [30]. Normally, the most effective way of killing cancer cells is by inducing apoptosis with chemotherapeutic agents such as DNA cross-linking agents, antimetabolites, topoisomerase I/II inhibitors, and intercalating agents, and tyrosine kinase inhibitors (TKIs).
The disruption of apoptotic pathways, both intrinsic and extrinsic, has been implicated in the development of glioma MDR [31]. Tumor cells can evade apoptosis via downregulating pro-apoptotic proteins and upregulating anti-apoptotic proteins. The escape of apoptosis has been widely observed in MDR against a variety of chemotherapeutics, including paclitaxel, doxorubicin, mitoxantrone, etoposide, cisplatin, and camptothecin [32,33,34].
2.4. Defects in DNA Repair
As a consequence of chemotherapy-induced DNA damage, cancer cells have increased DNA repair capability which is implicated in MDR development. At the beginning of gliomagenesis, the DNA damage repair (DDR) system is fundamentally activated via oncogene-evoked replication and oxidative stress. By reason of, DNA repair machinery diminishes the efficacy of genotoxic treatments, understanding and characterizing the DDR is essential to developing new therapeutic strategies in GBM [35]. A recent study reported that combining chemotherapeutics with DNA repair inhibitors helps improve treatment efficacy by decreasing the likelihood of chemo-resistant cancer arising [36].
2.5. Cancer Stem Cells
Cancer stem cells (CSCs) are subpopulations of tumor cells that are characterized by the ability to differentiate and self-renew, aberrant proliferation, long lifespan, active DNA repair capacity, and resistance to apoptosis [37]. As a result of their specific intrinsic mechanisms, CSCs do not disappear with chemotherapy because they overexpress drug transporters which prevent them from undergoing anti-cancer agent-induced apoptosis [38]. GSC are multipotent cells that share many of the characteristics of CSCs, such as their capacity for self-renewal. Moreover, they are evidence of a critical role in tumor maintenance, recurrence, and the development of glioblastoma MDR [39,40]. Additionally, CSC-like populations of glioblastoma cells are highly resistant to glioblastoma therapeutic agents [39].
2.6. Epithelial to Mesenchymal Transition
Another possible mechanism for the development of MDR in cancer cells is the converting EMT is a reversible biological process to mesenchymal–epithelial transition (MET) characteristics [41]. In this complex process, some significant morphologic alterations occur in epithelial cells which are transformed into elongated fibroblastic mesenchymal cells phenotype, that are significantly more invasive and motile, as well as developing an MDR [42]. Such diversity in mesenchymal features may be brought about by various microenvironmental factors, as well as intrinsic genetic alterations in glioma tissues. Simultaneous with genetic and especially epigenetic alterations that cancer cells endure making them sensitive to EMT-inducing signals. Mesenchyme-like cancer cells are commonly observed at the invasive foreside, considering that signals that related to dedifferentiation usually originate from the tumor microenvironment. In these processes, Snail, as a member of the Snail family of transcriptional activators, modulates various other EMT phenotypes, such as the decreased expression of diverse epithelial markers including claudins, cytokeratin, and occludins. Furthermore, in the next step increase the expression of mesenchymal markers including vitronectin and fibronectin. Slug is another member of the Snail family of transcriptional activators and performs as a suppressing the epithelial phenotype in numerous cancer cells. Moreover, this transcriptional factor increases both migration and invasion of malignant gliomas [43,44]. These alterations cause MDR in GBM which is more considerable during chemotherapy (Figure 1).
3. MiRNAs
MiRNA plays a significant role in biological processes including cell proliferation and differentiation by targeting protein-coding mRNA at the posttranscriptional level [45]. MiRNA is one of the key regulators of the above-mentioned mechanisms of MDR by modulating target gene expression [46]. It is estimated that miRNAs control the translation status of more than 50% of the human genome. It means that a particular target gene can be controlled by multiple miRNAs, as well as a miRNA also can be involved in the regulation of various target mRNAs [47]. Multiple studies have highlighted the role of miRNAs in the process of carcinogenesis. However, intriguingly, it has appeared that the identical miRNA molecules might act as either suppressors and/or oncogenes, contingent on the organ or tissue [48]. The dysregulated miRNAs have been revealed to alter hallmarks of cancer including, evading growth suppressors, activating invasion, metastasis, angiogenesis, and resisting cell death.
4. Roles of MiRNAs in Glioblastoma
A broad range of microarray analyses has demonstrated that the expression profile of multiple miRNAs shows significant alteration in glioblastomas [49]. There is a long list of miRNAs, which facilitate cell growth, proliferation, invasion, metastasis, angiogenesis, evade immune destruction, and reprogram cellular energy in glioblastoma (Table 1). Exosomes (microvesicles) are extracellular vesicles (EVs) that contain various molecules such as DNA, mRNA, growth factors, oncogenic receptors, enzymes, and microRNA molecules. Tumor cells in aggressive glioblastomas can release these molecules, thereby inducing the oncogenic transformation of neighboring cells [50]. It is reported that the co-culture of U87MG astrocytoma cells and human neural stem cell-derived astrocytes led to the induction of malignant-like phenotypes in astrocytes acquired from tumor cells by inducing the expression of GFAP, MMP-2, TGF-B1, SPARC, and CX43 [51]. Moreover, co-culturing of MSCs with U87MG simultaneously leads to a decrease in MMP inhibitor (TIMP-2) expression, indicating that U87MG could elevate a modification of the phenotype of neighboring astrocytes which may provide a significant change to the extracellular matrix of the tumor microenvironment and allow tumor invasion [52,53]. In addition, it was suggested that a certain miRNA was shared between normal and glioblastoma cells. Therefore, these studies proved that miRNAs play a direct role in glioblastoma malignancy.
5. The Most Frequent miRNAs Involved in the Signaling Pathway in Glioblastoma
It has been some miRNA being dysregulated in glioblastoma, including upregulation and downregulation which can bridge the widening gap between cancer treatment and failure. Some miRNAs play a crucial role in certain signaling pathways, with a particular mechanism that might be used as a sensitive and effective therapeutic approach for glioblastoma. Several important pathways have been identified as frequently genetically modified in cancer, including the EGFR/Ras/Raf/MEK/ERK pathways. Furthermore, this pathway plays a pivotal role in regulating cell proliferation and differentiation within the signaling network. The association of this pathway with some of the important microarrays in glioblastoma is considerable. For, instance, the EGFR gene expression has been altered in glioblastoma. EGFR amplification triggers downstream signaling pathways with moderate carcinogenicity. There are some new and important functional miRNAs in glioblastoma cells development:
5.1. MiR-218
miR-218 as a tumor-suppressive microRNA, which is decreased significantly in highly necrotic mesenchymal GBM. Studies revealed that reduced miR-218 levels confer GBM resistance to chemotherapy. Therefore, this miRNA was identified as a tumor suppressor gene in glioblastoma and regulated by RTK signaling in glioma cells [45,130]. RTK signaling pathway promotes tumor growth and plays an important role in tumor progression. Mathew et al. reported that in glioblastoma, miR-218 inhibition increased multiple RTK activities through the regulatory feedback loop; this mechanism may suppress RTK signaling and ultimately result in the proliferation of glioblastoma. Briefly, the activation of the RTK signaling promotes the expression of STAT3 which binds to the miR-218 locus with B-cell lymphoma 2-associated transcription factor 1 (BCLAF1) and consequently inhibits the expression of miR-218 and resulting in the suppression of glioblastoma proliferation [60]. In another study, glioblastoma samples with deep necrosis are substantially intensified in the mesenchymal transcriptional gene signature. In particular, hypoxic glioblastoma cells surrounding necrotic zones express high levels of C/EBP-β and C/EBP-δ, the mesenchymal transcription factors, indicating a link between hypoxia, necrosis, and specific mesenchymal transcription factors in glioblastoma cellular identity. The miR-218 levels were found to be lower in highly necrotic and hypoxic glioblastomas than in less necrotic tissues. Furthermore, miR-218 downregulation results in resistance to chemotherapy in glioblastoma. GSCs express more hypoxia-induced factors (HIFs), particularly, HIF2, which is required for GSC growth and survival. MiR-218 is downregulated in mesenchymal glioblastomas that have high levels of necrosis and hypoxia and promotes either RTK or HIF activation. Therefore, the miR-218–RTK–HIF2α as an efficient signaling pathway influences mesenchymal glioblastoma that exhibits a high degree of aggressiveness. Furthermore, HIF2α knockdown diminishes VEGF expression, precludes GSC-induced angiogenesis, and therefore is considered a promising target for anti-GBM therapeutics. The upregulation of miR-218 as a newly discovered tumor suppressor miRNA has proven to be a viable approach to dealing with chemoresistance [62].
5.2. MiR-7
MiR-7 is one of the most potent tumor suppressors in GBM and has been shown to regulate proliferation, migration, and invasion. MiR-7 is expressed mostly in normal brain and pancreatic tissue, which illustrates a high degree of tissue specificity may be an ideal target for cancer therapy, particularly in GBM. Liu et al. reported that miR-7 can target multiple oncogenes including PI3K and Raf-1 by the EGFR pathway, bringing up a strong perception of the role of this miRNA in tumor cell proliferation. This miR is a common regulator of the important pathways including PI3K, ATK, Raf, MEK, and ERK. Bioinformatic studies revealed that there are four potential binding sites of miR-7 in the 3′-UTR of EGFR, PI3K, and Raf-1. This study confirmed that PI3K and Raf-1 mRNAs are direct targets of miR-7 through luciferase assay. However, there was no identified clear targeting relationship between EGFR and miR-7 in this experiment. As a result, miR-7 inhibits synchronously the PI3K/ATK and Raf/MEK/ ERK pathways via PI3K and Raf-1, which are placed downstream of EGFR. All of these findings imply that miR-7 can be a key factor and a potential therapeutic target in GBM [17,131,132].
5.3. MiR-21
MiR-21 has been consistently upregulated in glioblastomas and involved in a wide variety of biological pathways, promoting tumor cell survival and invasiveness [48]. This miR is the first one to be found in human glioblastoma. STAT3, as a part of the STAT family of transcription factors, has been illustrated to play a very essential role in glioma tumorigenesis via promoting angiogenesis, and invasion. Moreover, miR-21 can be activated through a variety of other growth factor receptors and cytokines, including EGFR, IL-6R, JAK, and other kinases [133]. It has been suggested that reversion-inducing cysteine-rich protein with kazal motifs (RECK), a glycosylphosphatidylinositol-anchored membrane-bound regulator of matrix metalloproteinases (MMPs) as well as tissue inhibitor of metalloproteinase-3 (TIMP3), are both unlikely candidates to act as tumor suppressors during carcinogenic processes. In addition, these genes act as inhibitors of MMPs. It was discovered that miR-21 upregulated in glioma samples with significantly higher stages, followed by a lower mRNA expression of RECK and TIMP3 and subsequently, this process caused either reduction in MMP-2 activity or cellular motility. MMPs disrupt the extracellular matrix and help glioma cells motility and metastasis [133]. MiR-21 enhances glioma cells resistance to carmustine (BCNU) and promotes cell cycle arrest in the G2/M phase, phosphatase and PTEN gene expression. Therefore, miR-21 inhibition increases the chemosensitivity of glioma cells [134]. Accordingly, miR-21 inhibition enhanced the effectiveness of treatment. In recent studies, it has been demonstrated that inhibiting miR-21 and inducing miR-7 could be a promising strategy to inhibit parallel survival pathways and produce a synergistic effect in malignant glioma cells by inhibiting BCL2, PI3K/AKT, and Raf/MEK/ERK [135]. These findings provide evidence that miRNAs might be promising targets for glioblastoma treatment by regulating signaling pathways and opening the possibility for novel therapeutic approaches such as combined therapy to achieve synergistic inhibitions (Figure 2).
6. The Role of miRNAs in Glioblastoma MDR
6.1. MiRNAs That Target MDR Transporters
Tumor cells that overexpress drug transporters may be able to maintain low intracellular levels of cytotoxic chemotherapeutic agents by pumping them into extracellular space. MiRNAs play a significant role in regulating glioblastoma MDR by influencing the level of MDR transporter expression. ABCG2 is a main member of ABC transporters with high expression levels in glioblastoma [136]. It has been demonstrated that miR-328 targets and inhibits ABCG2 in glioblastoma cells, thereby sensitizing the cells to chemotherapeutics [17]. It was also reported that upregulation of miR-9 levels led to the inhibition of ABC transporters, including MDR1, ABCC3, and ABCC6, which reversed MDR in glioblastoma cells [137]. MiR-381 is a common tumor suppressor miRNA that is downregulated in glioblastoma [138]. It has been demonstrated that miR-381 overexpression effectively sensitized glioblastoma U251 cells to temozolomide by targeting various ABC transporters including ABCG2, ABCC3, and ABCC5 [139]. MiR-1268a is another tumor suppressor miRNA with downregulated expression in glioblastoma. Li et al. [140]. reported downregulation of miR-1268a following temozolomide treatment in glioblastoma cells. The authors found that overexpression of miR-1268a suppressed protein translation of ABCC1 and reversed upregulation of ABCC1 due to temozolomide. Inversely, knockdown of miR-1268a increased ABCC1 at the protein level and enhanced upregulation of ABCC1 with TMZ treatment [140].
6.2. MiRNAs Targeting Apoptosis
One of the most important mechanisms used by tumor cells in MDR is evading anti-cancer drug-induced apoptosis. Various microRNAs are ectopically expressed which disrupts apoptotic pathways and is implicated in the development of MDR in glioblastoma. For example, the inhibition of miR-497 which is upregulated in glioblastoma cells, resulted in a significant increase in apoptosis and enhancement in the sensitivity of glioblastoma cells to temozolomide [141]. Treatment of glioblastoma cells with miR-21 inhibitors leads to a significantly higher apoptotic rate than treatment with temozolomide alone, thus overcoming drug resistance [17]. As reported by Yang et al., upregulation of miR-29a in CD133+ glioblastoma cells increased cisplatin-induced apoptosis and decreased survival of CD133+ tumor-bearing mice after treatment with cisplatin [142] (Figure 3). Increasing the expression level of miR-181b led to a synergistic effect on temozolomide-induced apoptosis [143]. Further, miRNAs have been demonstrated to target the intrinsic and extrinsic pathways of apoptosis to reverse drug resistance in glioblastoma cells. Silencing an oncogenic microRNA, miR-21, in sunitinib-resistant glioblastoma cells led to an increase in the apoptotic rate of cancer cells by overexpressing PTEN and PDCD4, as well as increased activity of caspase 3/7, reversing MDR in these cells [144]. Shi et al. reported that combination treatment of glioblastoma cells with miR-125b-2 and temozolomide potently increased cancer cells apoptosis through activation in the mitochondrial pathway by targeting APAF-1, CASPASE-3, BAX, BCL-2, and poly-ADP-ribose polymerase (PARP) [145]. BCL-2 is a target of various microRNAs, such as miR-181b-5p and miR-18, that acts to overcome temozolomide-induced resistance in glioblastoma cells [146,147]. MiR-155-5p, miR-221-3p, miR-21, and miR-125b were reported to play critical roles in developing MDR in glioblastoma cells through targeting caspase-3 [148,149,150].
Mouse double minute 2 (MDM2) is an important negative regulator of the TP53 tumor suppressor which is also a direct target of miR-181b [151,152]. Sun et al. found that miR-181b overexpression sensitized U87 glioblastoma cell lines to temozolomide-mediated apoptosis by downregulating MDM2 [146]. Pro-apoptotic BCL-2 antagonist killer 1 (Bak1) is targeted by miR-125b, which increases the chemosensitivity of glioblastoma stem cells to temozolomide [153].
6.3. MiRNAs Targeting DNA Repair
Another well-established mechanism for miRNA-mediated MDR in glioblastoma involves targeting specific components of the DNA repair machinery.
The temozolomide is activated in the CNS by a chemical reaction which results in DNA methylation at various sites [154]. A DNA repair enzyme, O6-methyl-guanine-methyltransferase (MGMT), removes any methyl adducts from DNA [155]. It is important to note that MGMT has also been shown to contribute significantly to glioblastoma MDR [156]. MiR-10a, miR-195, and miR-455-3p are among the upregulated miRNAs in temozolomide-resistant glioblastoma cells [157]. miR-181b and miR-181c were also downregulated in patients with a glioblastoma that was resistant to temozolomide [158]. In both studies, it was established that the methylation status of MGMT was an independent predictor of response to temozolomide. Nie et al. [159] reported that miR-198 was downregulated in glioblastoma patients. Patients with downregulation of this miRNA were more likely to have a poor prognosis. Moreover, in vitro and in vivo studies demonstrated that overexpression of miR-198 was associated with enhanced chemosensitivity to temozolomide. This was accomplished by miR-198 directly targeting MGMT and suppressing its protein translation. Therefore, miR-198 induced chemosensitivity to temozolomide in glioblastoma by targeting MGMT [159]. In another study by Gao et al. [160], it was found that transfection of temozolomide-resistant glioblastoma cells with the miR-370-3p, which is downregulated in glioblastoma, enhanced the sensitivity of the cells to the anticancer drug by inhibiting self-repair capacity of tumor cells’ DNA. Based on their results MGMT is a direct target of miR-370-3p and that plays a critical role in the miRNA-mediated reversal of MDR in glioblastoma [160]. Non-homologous end-joining (NHEJ) is another DNA repair mechanism playing crucial functions in temozolomide sensitivity in glioblastoma.
An important component of this pathway is the XRCC4 protein, which has recently been discovered to be a direct target of miR-151a in resistant glioblastoma cells [161]. Zeng et al. [162] showed that low miR-151a levels in glioblastoma patients correlated with poor response to temozolomide therapy. Restoring miR-151a expression sensitized temozolomide-resistant glioblastoma cells through inhibition of XRCC4-mediated DNA repair [162].
6.4. MiRNA Regulating Cancer Stem Cells
MicroRNAs have been demonstrated to play a role in the CSCs-linked MDR in glioblastoma. MiR-125b-2 is an oncogenic miRNA that is highly expressed in glioblastoma cells and GSCs. Shi et al. reported that treatment of GSCs with miR-125b-2 inhibitors significantly increased stem cell sensitivity to temozolomide [8]. Furthermore, downregulation of miR-21 inhibition in GSCs resulted in the suppression of cell proliferation and the induction of apoptosis, resulting in enhanced sensitivity to chemotherapeutic agents [163,164]. Cheng et al. found that miR-132 plays a vital role in the development of resistance against temozolomide and induces the formation of CSC-like phenotypes in glioblastoma U87MG cells. Their finding indicated that miR-132 inhibited the expression levels of tumor suppressor candidate 3 (TUSC3), which is downregulated in temozolomide-resistant U87MG cells (U87MG-res cells) and its renewal sensitized U87MG-res cells to temozolomide. This protein is able to inhibit the formation of GIC phenotypes in the U87MG-res cells. Hence, high expression levels of TUSC3 were associated with the high sensitivity of cancer cells to temozolomide [165]. As discussed above, all of the studies have highlighted the important role played by miRNAs in the MDR induced by CSCs in glioblastoma.
7. MiRNA Targeting EMT
Glioblastoma MDR may be influenced by miRNAs targeting the key components of EMT [166]. MiR-203 is a tumor suppressor miRNA, which is significantly downregulated in the resistant glioblastoma cells. This miRNA can bind to 3′-UTR SNAI2, as an embryonic protein with the ability to suppress E-cadherin transcription and induce EMT directly [117]. SNAI2 was expressed at a higher level in glioblastoma-resistant cells. Transfection of miR-203 in resistant cells inhibited SNAI2 expression, reversing EMT and MDR in response to imatinib [117]. MiR-26b is also downregulated in temozolomide-resistant glioblastoma cells. It has been demonstrated that miR-26b can sensitize resistant cells to temozolomide by targeting Wee-1 [167]. The expression of Wee-1 plays an important role in regulating EMT and drug resistance by modulating the expression of sensible drug resistance genes and the activity of the MEK/ERK pathway [167]
8. Crosstalk between Signaling Pathways and miRNAs in Glioblastoma MDR
An accumulating number of recent studies have reported the mutual interaction between miRNAs and key components of various signaling pathways including epidermal growth factor, Wnt/β-catenin, nuclear factor kappa B (NF-κB), and PI3K signaling pathways in developing MDR in glioblastoma [168,169,170,171,172,173]. The EGFR is an important signaling pathway that ectopic activation of this receptor has been extensively characterized in glioblastoma cells [174]. Through this receptor, glioblastoma cells are induced to proliferate, differentiate, and survive [174]. Chen et al. [175] showed that miR-181b upregulation resulted in the significant enhancement in the chemo-sensitivity of glioblastoma cells to temozolomide through potentiating temozolomide-induced apoptosis. MiR-181b directly targets the EGFR, restoring EGFR decreased the suppressive effects of miR-181b and temozolomide treatment [175]. A separate study by Zhang et al. found that miR-566 overexpression was associated with nimotuzumab resistance in glioblastoma cell lines. Moreover, treating cells with miR-566 inhibitor decrease the EGFR pathway activity, reversing nimotuzumab resistance in glioblastoma cells [60]. Another study has demonstrated that exosomal miR-1238 contributed to the development of temozolomide-resistance in glioblastoma cells in vitro and in vivo through significant activation of the EGFR-PI3K-Akt-mTOR pathways [176]. Zhang et al. [177] found that an increase in the expression levels of miR-625 in glioblastoma cells inhibited cellular proliferation, induced apoptosis, and arrested cell cycle as well as suppressed tumor growth in the animal model of glioblastoma. Moreover, miR-625 targeted and inhibited AKT2, thereby sensitizing cells to temozolomide [177]. The upregulation of miR-423-5p in glioblastoma cells was shown to lead to overexpression of signaling molecules such as p-AKT and p-ERK1/2. Therefore, this miRNA was responsible for glioblastoma MDR through activation of the AKT/ERK pathway [178]. NF-κB signaling is another signaling cross-talking with miRNAs and plays a key role in the glioblastoma MDR. Wang et al. showed that miR-133a induced TRAIL resistance in glioblastoma by inhibiting death receptor (DR)-5 expression and activating NF-κB signaling [84]. It was demonstrated that miR-126-3p and miR-101 sensitized glioblastoma cells to temozolomide via targeting and inhibiting Wnt/β-catenin signaling [179,180].
9. Conclusions and Perspective
In cancer treatment, a major obstacle to patient treatment is the occurrence of MDR [9]. Various mechanisms contribute to the development of MDR during the treatment process [181]. It is noteworthy that miRNAs are important modulators of cellular pathways by regulating the expression of target genes during MDR; thus contributing significantly to the complexity of treatment failure in cancer progression, including glioblastoma [182]. The importance of some miRNAs for tumorigenesis and MDR mechanisms cannot be overstated. MiRNAs-mediated MDR in glioblastoma comes with a number of mechanisms including targeting MDR transporters, modulating apoptosis, targeting DNA repair machinery, controlling cancer stem cells, regulating EMT, and cross-talking with major oncogenic pathways. By targeting signaling pathways such as EGFR, RTK, and HIF2α, miRNAs have a direct effect on causing MDR and therefore increasing the effectiveness of glioblastoma treatment. There is no doubt that miRNAs are targeting the critical components of EMT, suppressing E-cadherin transcription and promoting EMT. Furthermore, miRNAs regulate signaling pathways such as epidermal growth factor, Wnt/β-catenin, NF-κB, and PI3K in the progression of MDR in glioblastoma. These studies demonstrate the importance of miRNAs in glioblastoma and glioma, but there are significant gaps in the knowledge in this area which need to be filled or improved. It will be necessary to clarify the role of potential miRNAs and their networking in modulating MDR mechanisms involved in glioblastoma. Furthermore, in the terms of therapeutics, there are not sufficient studies that address how miRNA applications can be translated for MDR glioblastoma patients to help move the bench-top research into clinical trials. There is a significant challenge in miRNA systemic delivery, especially which of natural and/or synthetic miRNA carriers work as the best systemic carrier for miRNA, their pharmacokinetics, as well as conducting related trials studies to understand their safety and effectiveness. In addition, in the terms of diagnosis and prognosis, profiling of miRNA needs more investigation to identify a miRNA signature for diagnosis and prognosis of resistance GBM via collecting the samples from body fluids such as serum, and CNS. This could be achieved by novel RNA-Based detection platforms such as Nonostring®.
Lastly but more importantly, we can suggest a potential application of miRNA studies in glioblastoma based on the analysis of the different studies discussed in this review. MiRNA can be used primarily for diagnostic and prognostic purposes in patients with glioblastoma. To recapitulate, by contemplating the subjects, miRNAs play an important role in the development of drug resistance in glioblastoma by modulating different types of important mechanisms and signaling pathways. A greater understanding of the roles of miRNAs in MDR glioblastoma will increase the number of miRNAs which may be nominated for replacement or blocking as potential therapeutic procedures. These candidates will be validated through extensive in vivo, in vitro, and ultimately clinical trials. In particular, miR-21 and miR-7 are momentous and functional miRNAs in MDR glioblastoma cells, which should be excellent candidates for further investigation both in vivo and in vitro. MiR-21 is one of the most prominent oncogenic miRNAs in cancer, especially glioblastoma. Blocking this miRNA with anti-miR or miRspong is a potential strategy to decrease its oncogenic function. Moreover, inhibiting miR-21a is an effective therapeutic approach, especially for GCS (CD133+) cells, which are the main population for GBM relapse. In contrast, MiR-7 is a tissue-specific miRNA that is significantly reduced in GBM cells, and its replacement may prove to be an ideal treatment strategy for the disease. Furthermore, combining commonly used GBM chemotherapeutic agents or small inhibitor molecules with miRNA-based therapeutics may have the potential to overcome patient resistance. The combination approach requires extensive knowledge of the functional properties of therapeutic agents and miRNAs; selecting miRNAs that complement agent function is crucial in this approach.
Author Contributions
Conceptualization, P.M. and B.M. (Behnaz Mansoori).; methodology, B.M. (Behnaz Mansoori); software, D.R.; validation, D.R., B.B. and B.M. (Behzad Mansoori).; investiga-tion, P.M. and B.M.(Behnaz Mansoori).; resources, B.M. (Behzad Mansoori). and W.C.C.; data cu-ration, B.M. (Behzad Mansoori) and W.C.C.; writing—original draft preparation, P.M. and B.M.; writing—review and editing, D.R., W.C.C. and B.M. (Behzad Mansoori); visualization, B.M. (Behzad Mansoori); supervision, B.M. (Behzad Mansoori) and W.C.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Desjardins, A.; Gromeier, M.; Herndon, J.E.; Beaubier, N.; Bolognesi, D.P.; Friedman, A.H.; Friedman, H.S.; McSherry, F.; Muscat, A.M.; Nair, S.; et al. Recurrent Glioblastoma Treated with Recombinant Poliovirus. N. Engl. J. Med. 2018, 379, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Hanif, F.; Muzaffar, K.; Perveen, K.; Malhi, S.M.; Simjee, S.U. Glioblastoma Multiforme: A Review of Its Epidemiology and Pathogenesis through Clinical Presentation and Treatment. Asian Pacific J. Cancer Prev. 2017, 18, 3–9. [Google Scholar] [CrossRef]
- Shergalis, A.; Bankhead, A.; Luesakul, U.; Muangsin, N.; Neamati, N. Current Challenges and Opportunities in Treating Glioblastomas. Pharmacol. Rev. 2018, 70, 412–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 WHO Classification of Tumors of the Central Nervous System: A Summary. Neuro. Oncol. 2021, 23, 1231–1251. [Google Scholar] [CrossRef]
- Shahneh, F.Z.; Valiyari, S.; Baradaran, B.; Abdolalizadeh, J.; Bandehagh, A.; Azadmehr, A.; Hajiaghaee, R. Inhibitory and Cytotoxic Activities of Salvia Officinalis l. Extract on Human Lymphoma and Leukemia Cells by Induction of Apoptosis. Adv. Pharm. Bull. 2013, 3, 51–55. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, H.; Xu, S.; Liu, Z.; Cheng, Q. The Adaptive Transition of Glioblastoma Stem Cells and Its Implications on Treatments. Signal Transduct. Target. Ther. 2021, 6, 124. [Google Scholar] [CrossRef]
- Fisher, J.P.; Adamson, D.C. Current FDA-Approved Therapies for High-Grade Malignant Gliomas. Biomedicines 2021, 9, 324. [Google Scholar] [CrossRef]
- Olivier, C.; Oliver, L.; Lalier, L.; Vallette, F.M. Drug Resistance in Glioblastoma: The Two Faces of Oxidative Stress. Front. Mol. Biosci. 2021, 7, 620677. [Google Scholar] [CrossRef]
- Bukowski, K.; Kciuk, M.; Kontek, R. Mechanisms of Multidrug Resistance in Cancer Chemotherapy. Int. J. Mol. Sci. 2020, 21, 3233. [Google Scholar] [CrossRef]
- Mansoori, B.; Mohammadi, A.; Davudian, S.; Shirjang, S.; Baradaran, B. The Different Mechanisms of Cancer Drug Resistance: A Brief Review. Adv. Pharm. Bull. 2017, 7, 339–348. [Google Scholar] [CrossRef]
- Hamed, A.R.; Abdel-Azim, N.S.; Shams, K.A.; Hammouda, F.M. Targeting Multidrug Resistance in Cancer by Natural Chemosensitizers. Bull. Natl. Res. Cent. 2019, 43, 8. [Google Scholar] [CrossRef] [Green Version]
- Zheng, T.; Wang, J.; Chen, X.; Liu, L. Role of MicroRNA in Anticancer Drug Resistance. Int. J. Cancer 2010, 126, 2–10. [Google Scholar] [CrossRef]
- Hassn Mesrati, M.; Behrooz, A.B.; Abuhamad, A.Y.; Syahir, A. Understanding Glioblastoma Biomarkers: Knocking a Mountain with a Hammer. Cells 2020, 9, 1236. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, H.; Chen, X. Drug Resistance and Combating Drug Resistance in Cancer. Cancer Drug Resist. 2019, 2, 141–160. [Google Scholar] [CrossRef] [Green Version]
- Vaghari-Tabari, M.; Majidinia, M.; Moein, S.; Qujeq, D.; Asemi, Z.; Alemi, F.; Mohamadzadeh, R.; Targhazeh, N.; Safa, A.; Yousefi, B. MicroRNAs and Colorectal Cancer Chemoresistance: New Solution for Old Problem. Life Sci. 2020, 259, 118255. [Google Scholar] [CrossRef]
- Ward, R.A.; Fawell, S.; Floc’h, N.; Flemington, V.; McKerrecher, D.; Smith, P.D. Challenges and Opportunities in Cancer Drug Resistance. Chem. Rev. 2021, 121, 3297–3351. [Google Scholar] [CrossRef]
- Chen, M.; Medarova, Z.; Moore, A. Role of Micrornas in Glioblastoma. Oncotarget 2021, 12, 1707–1723. [Google Scholar] [CrossRef]
- Nikolaou, M.; Pavlopoulou, A.; Georgakilas, A.G.; Kyrodimos, E. The Challenge of Drug Resistance in Cancer Treatment: A Current Overview. Clin. Exp. Metastasis 2018, 35, 309–318. [Google Scholar] [CrossRef]
- Chernov, A.N.; Alaverdian, D.A.; Galimova, E.S.; Renieri, A.; Frullanti, E.; Meloni, I.; Shamova, O.V. The Phenomenon of Multidrug Resistance in Glioblastomas. Hematol. Oncol. Stem Cell Ther. 2021; in press. [Google Scholar] [CrossRef]
- Li, Y.J.; Lei, Y.H.; Yao, N.; Wang, C.R.; Hu, N.; Ye, W.C.; Zhang, D.M.; Chen, Z.S. Autophagy and Multidrug Resistance in Cancer. Chin. J. Cancer 2017, 36, 52. [Google Scholar] [CrossRef]
- Feng, Z.; Liu, D.; Wang, L.; Wang, Y.; Zang, Z.; Liu, Z.; Song, B.; Gu, L.; Fan, Z.; Yang, S.; et al. A Putative Efflux Transporter of the ABC Family, YbhFSR, in Escherichia Coli Functions in Tetracycline Efflux and Na+(Li+)/H+ Transport. Front. Microbiol. 2020, 11, 556. [Google Scholar] [CrossRef]
- Vivo, I.; Characterization, P.; Dual, P.; Molinari, A.; Bugli, F.; Sanguinetti, M.; Torelli, R.; Martini, M.; Maccari, L.; Valoti, M.; et al. A New Strategy for Glioblastoma Treatment: In Vitro. Cancers 2019, 11, 848. [Google Scholar]
- Arya, V.; Kiser, J.J. Role of Transporters in Drug Development. J. Clin. Pharmacol. 2016, 56, S7–S10. [Google Scholar] [CrossRef]
- Bart, A.G.; Morais, G.; Vangala, V.R.; Loadman, P.M.; Pors, K.; Scott, E.E. Cytochrome P450 Binding and Bioactivation of Tumor-Targeted Duocarmycin Agents. Drug Metab. Dispos. 2022, 50, 49–57. [Google Scholar] [CrossRef]
- Tsai, Y.T.; Lo, W.L.; Chen, P.Y.; Ko, C.Y.; Chuang, J.Y.; Kao, T.J.; Yang, W.B.; Chang, K.Y.; Hung, C.Y.; Kikkawa, U.; et al. Reprogramming of Arachidonate Metabolism Confers Temozolomide Resistance to Glioblastoma through Enhancing Mitochondrial Activity in Fatty Acid Oxidation. J. Biomed. Sci. 2022, 29, 21. [Google Scholar] [CrossRef]
- Kuban, W.; Daniel, W.A. Cytochrome P450 Expression and Regulation in the Brain. Drug Metab. Rev. 2021, 53, 1–29. [Google Scholar] [CrossRef]
- Sanchez-Spitman, A.B.; Swen, J.J.; Dezentjé, V.O.; Moes, D.J.A.R.; Gelderblom, H.; Guchelaar, H.J. Effect of CYP2C19 Genotypes on Tamoxifen Metabolism and Early-Breast Cancer Relapse. Sci. Rep. 2021, 11, 415. [Google Scholar] [CrossRef]
- Aubrey, B.J.; Kelly, G.L.; Janic, A.; Herold, M.J.; Strasser, A. How Does P53 Induce Apoptosis and How Does This Relate to P53-Mediated Tumour Suppression? Cell Death Differ. 2018, 25, 104–113. [Google Scholar] [CrossRef] [Green Version]
- Mantovani, F.; Collavin, L.; Del Sal, G. Mutant P53 as a Guardian of the Cancer Cell. Cell Death Differ. 2019, 26, 199–212. [Google Scholar] [CrossRef]
- Zhang, Y.; Dube, C.; Gibert, M.; Cruickshanks, N.; Wang, B.; Coughlan, M.; Yang, Y.; Setiady, I.; Deveau, C.; Saoud, K.; et al. The P53 Pathway in Glioblastoma. Cancers 2018, 10, 297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neophytou, C.M.; Trougakos, I.P.; Erin, N.; Papageorgis, P. Apoptosis Deregulation and the Development of Cancer Multi-Drug Resistance. Cancers 2021, 13, 4363. [Google Scholar] [CrossRef] [PubMed]
- Morris, J.L.; Gillet, G.; Prudent, J.; Popgeorgiev, N. Bcl-2 Family of Proteins in the Control of Mitochondrial Calcium Signalling: An Old Chap with New Roles. Int. J. Mol. Sci. 2021, 22, 3730. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.H.; Hsiao, M.Y.; Kung, Y.; Huang, A.P.H.; Chen, W.S. Investigation of the Therapeutic Effect of Doxorubicin Combined With Focused Shockwave on Glioblastoma. Front. Oncol. 2021, 11, 711088. [Google Scholar] [CrossRef] [PubMed]
- Maliyakkal, N.; Appadath Beeran, A.; Udupa, N. Nanoparticles of Cisplatin Augment Drug Accumulations and Inhibit Multidrug Resistance Transporters in Human Glioblastoma Cells. Saudi Pharm. J. 2021, 29, 857–873. [Google Scholar] [CrossRef]
- Annovazzi, L.; Mellai, M.; Schiffer, D. Chemotherapeutic Drugs: DNA Damage and Repair in Glioblastoma. Cancers 2017, 9, 57. [Google Scholar] [CrossRef] [Green Version]
- Majidinia, M.; Yousefi, B. Long non-coding RNAs in cancer drug resistance development. DNA Repair 2016, 45, 25–33. [Google Scholar] [CrossRef]
- Phi, L.T.H.; Sari, I.N.; Yang, Y.G.; Lee, S.H.; Jun, N.; Kim, K.S.; Lee, Y.K.; Kwon, H.Y. Cancer Stem Cells (CSCs) in Drug Resistance and Their Therapeutic Implications in Cancer Treatment. Stem Cells Int. 2018, 2018, 5416923. [Google Scholar] [CrossRef] [Green Version]
- Suresh, R.; Ali, S.; Ahmad, A.; Philip, P.A.; Sarkar, F.H. The Role of Cancer Stem Cells in Recurrent and Drug-Resistant Lung Cancer. In Lung Cancer and Personalized Medicine: Novel Therapies and Clinical Management; Springer: Berlin/Heidelberg, Germany, 2016; pp. 57–74. [Google Scholar] [CrossRef]
- Ong, D.S.T.; Hu, B.; Ho, Y.W.; Sauvé, C.E.G.; Bristow, C.A.; Wang, Q.; Multani, A.S.; Chen, P.; Nezi, L.; Jiang, S.; et al. PAF Promotes Stemness and Radioresistance of Glioma Stem Cells. Proc. Natl. Acad. Sci. USA 2017, 114, E9085–E9095. [Google Scholar] [CrossRef] [Green Version]
- Gravina, G.L.; Mancini, A.; Marampon, F.; Colapietro, A.; Delle Monache, S.; Sferra, R.; Vitale, F.; Richardson, P.J.; Patient, L.; Burbidge, S.; et al. The Brain-Penetrating CXCR4 Antagonist, PRX177561, Increases the Antitumor Effects of Bevacizumab and Sunitinib in Preclinical Models of Human Glioblastoma. J. Hematol. Oncol. 2017, 10, 5. [Google Scholar] [CrossRef] [Green Version]
- Hill, C.; Wang, Y. The Importance of Epithelial-Mesenchymal Transition and Autophagy in Cancer Drug Resistance. Cancer Drug Resist. 2020, 3, 38–47. [Google Scholar] [CrossRef] [Green Version]
- Shibue, T.; Weinberg, R.A. EMT, CSCs, and Drug Resistance: The Mechanistic Link and Clinical Implications. Nat. Publ. Gr. 2017, 14, 611–629. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.K.; Huo, S.F.; Zhang, G.; Zhang, F.; Lian, Z.P.; Tang, X.L.; Jin, C. CDA-2 Induces Cell Differentiation through Suppressing Twist/SLUG Signaling via MiR-124 in Glioma. J. Neurooncol. 2012, 110, 179–186. [Google Scholar] [CrossRef]
- Iwadate, Y. Epithelial-Mesenchymal Transition in Glioblastoma Progression. Oncol. Lett. 2016, 11, 1615–1620. [Google Scholar] [CrossRef] [Green Version]
- Setlai, B.P.; Hull, R.; Reis, R.M.; Agbor, C.; Ambele, M.A.; Mulaudzi, T.V.; Dlamini, Z. MicroRNA Interrelated Epithelial Mesenchymal Transition (EMT) in Glioblastoma. Genes 2022, 13, 244. [Google Scholar] [CrossRef]
- Brook, O.R. 34 Fibrotic Lung Disease. In Radiology Structured Reporting Handbook; Thieme: New York, NY, USA, 2021; pp. 1–26. [Google Scholar] [CrossRef]
- Kucherenko, M.M.; Shcherbata, H.R. miRNA Targeting and Alternative Splicing in the Stress Response—Events Hosted by Membrane-Less Compartments. J. Cell Sci. 2018, 131, jcs202002. [Google Scholar] [CrossRef] [Green Version]
- Gajda, E.; Godlewska, M.; Mariak, Z.; Nazaruk, E. Combinatory Treatment with MiR-7-5p and Drug-Loaded Cubosomes E Ff Ectively Impairs Cancer Cells. Int. J. Mol. Sci. 2020, 21, 5039. [Google Scholar] [CrossRef]
- Keeler, J.R.; Roth, E.A.; Neuser, B.L.; Spitsbergen, J.M.; Waters, D.J.M.; Vianney, J.M. The Neurochemistry and Social Flow of Singing: Bonding and Oxytocin. Front. Hum. Neurosci. 2015, 9, 518. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Ali, N.; Zhong, L.; Shi, J. MicroRNAs as Biomarkers for Human Glioblastoma: Progress and Potential. Acta Pharmacol. Sin. 2018, 39, 1405–1413. [Google Scholar] [CrossRef] [Green Version]
- Hallal, S.; Mallawaaratchy, D.M.; Wei, H.; Ebrahimkhani, S.; Stringer, B.W.; Day, B.W.; Boyd, A.W. Extracellular Vesicles Released by Glioblastoma Cells Stimulate Normal Astrocytes to Acquire a Tumor-Supportive Phenotype Via P53 and MYC Signaling Pathways. Mol. Neurobiol. 2019, 56, 4566–4581. [Google Scholar] [CrossRef] [Green Version]
- Yi, G.; Feng, W.; Zhou, Q.; Liu, Y.; Qi, S. The Impact of MMP-2 and Its Specific Inhibitor TIMP-2 Expression on the WHO Grade and Prognosis of Gliomas in Chinese Population: A Meta-Analysis. Mol. Neurobiol. 2016, 54, 22–30. [Google Scholar] [CrossRef] [Green Version]
- Breznik, B.; Motaln, H.; Vittori, M.; Rotter, A.; Lah, T. Mesenchymal Stem Cells Differentially Affect the Invasion of Distinct Glioblastoma Cell Lines. Oncotarget 2017, 8, 25482–25499. [Google Scholar] [CrossRef] [Green Version]
- Korać, P.; Antica, M.; Matulić, M. Mir-7 in Cancer Development. Biomedicines 2021, 9, 325. [Google Scholar] [CrossRef]
- Xu, F.; Li, F.; Zhang, W.; Jia, P. Growth of Glioblastoma Is Inhibited by MiR-133-Mediated EGFR Suppression. Tumor Biol. 2015, 36, 9553–9558. [Google Scholar] [CrossRef]
- Florian, A.I.; Timis, T.; Buruian, A.; Florian, I.; Hraps, I.; Mihu, C.M.; Micl, M.; Sergiu, S. The Roles of MiRNA in Glioblastoma Tumor Cell Communication: Diplomatic and Aggressive Negotiations. Int. J. Mol. Sci. 2020, 21, 1950. [Google Scholar]
- Li, X.; Liu, Y.; Granberg, K.J.; Wang, Q.; Moore, L.M.; Ji, P.; Gumin, J.; Sulman, E.P.; Calin, G.A.; Haapasalo, H.; et al. Two Mature Products of MIR-491 Coordinate to Suppress Key Cancer Hallmarks in Glioblastoma. Oncogene 2015, 34, 1619–1628. [Google Scholar] [CrossRef] [Green Version]
- Zirak, R.G.; Tajik, H.; Asadi, J. The Role of Micro RNAs in Regulating PI3K/AKT Signaling Pathways in Glioblastoma. Iran. J. Pathol. 2022, 17, 122–136. [Google Scholar] [CrossRef]
- Ma, Z.; Liu, T.; Huang, W.; Liu, H.; Zhang, H.M.; Li, Q.; Chen, Z.; Guo, A.Y. MicroRNA Regulatory Pathway Analysis Identifies MiR-142-5p as a Negative Regulator of TGF-ß Pathway via Targeting SMAD3. Oncotarget 2016, 7, 71504–71513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, B.; Mei, J.; Ji, W.; Huo, Z.; Bian, Z.; Jiao, J.; Li, X.; Sun, J.; Shao, J. MicroRNAs Involved in the EGFR Pathway in Glioblastoma. Biomed. Pharmacother. 2021, 134, 111115. [Google Scholar] [CrossRef]
- Neurochemistry, J.O.F. miR-144-3p exerts anti-tumor effects in glioblastoma by targeting c-Met. J. Neurochem. 2015, 135, 274–286. [Google Scholar] [CrossRef]
- Bouzari, B.; Mohammadi, S.; Bokov, D.O.; Krasnyuk, I.I.; Hosseini-Fard, S.R.; Hajibaba, M.; Mirzaei, R.; Karampoor, S. Angioregulatory Role of MiRNAs and Exosomal MiRNAs in Glioblastoma Pathogenesis. Biomed. Pharmacother. 2022, 148, 112760. [Google Scholar] [CrossRef]
- Bhaskaran, V.; Nowicki, M.O.; Idriss, M.; Jimenez, M.A.; Lugli, G.; Hayes, J.L.; Mahmoud, A.B.; Zane, R.E.; Passaro, C.; Ligon, K.L.; et al. The Functional Synergism of MicroRNA Clustering Provides Therapeutically Relevant Epigenetic Interference in Glioblastoma. Nat. Commun. 2019, 10, 442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, W.; Zheng, J.; Liu, X.; Ma, J.; Liu, Y.; Xue, Y. Knockdown of NEAT1 Restrained the Malignant Progression of Glioma Stem Cells by Activating MicroRNA Let-7e. Oncotarget 2016, 7, 62208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Zhao, M.; Guo, Q.; Lou, J.; Wang, L. Non-Small Cell Lung Cancer Cell–Derived Exosomal MiR-17-5p Promotes Osteoclast Differentiation by Targeting PTEN. Exp. Cell Res. 2021, 408, 112834. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Hao, Y.; Zhang, A.; Yang, W.; Wei, W.; Wang, G.; Jia, Z. MiR-19a/b Promote EMT and Proliferation in Glioma Cells via SEPT7-AKT-NF-ΚB Pathway. Mol. Ther.-Oncolytics 2021, 20, 290–305. [Google Scholar] [CrossRef]
- Cai, J.; Zhao, J.; Zhang, N.; Xu, X.; Li, R.; Yi, Y.; Fang, L.; Zhang, L.; Li, M.; Wu, J.; et al. MicroRNA-542-3p Suppresses Tumor Cell Invasion via Targeting AKT Pathway in Human Astrocytoma. J. Biol. Chem. 2015, 290, 24678–24688. [Google Scholar] [CrossRef] [Green Version]
- Shen, L.; Sun, C.; Li, Y.; Li, X.; Sun, T.; Liu, C.; Zhou, Y.; Du, Z. MicroRNA-199a-3p Suppresses Glioma Cell Proliferation by Regulating the AKT/MTOR Signaling Pathway. Tumor Biol. 2015, 36, 6929–6938. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Wang, Y.; Liu, R.; Kasinski, A.L.; Shen, H.; Slack, F.J.; Tang, D.G. MicroRNA-34a: Potent Tumor Suppressor, Cancer Stem Cell Inhibitor, and Potential Anticancer Therapeutic. Front. Cell Dev. Biol. 2021, 9, 640587. [Google Scholar] [CrossRef]
- Rathod, S.S.; Rani, S.B.; Khan, M.; Muzumdar, D.; Shiras, A. Tumor Suppressive MiRNA-34a Suppresses Cell Proliferation and Tumor Growth of Glioma Stem Cells by Targeting Akt and Wnt Signaling Pathways. FEBS Open Bio 2014, 4, 485–495. [Google Scholar] [CrossRef] [Green Version]
- Sasayama, T.; Tanaka, K.; Kohmura, E. The Roles of MicroRNAs in Glioblastoma Biology and Biomarker. In Neurooncology—Newer Developments; IntechOpen: London, UK, 2016. [Google Scholar] [CrossRef] [Green Version]
- Mazurek, M.; Litak, J.; Kamieniak, P.; Osuchowska, I.; Maciejewski, R.; Roliński, J.; Grajkowska, W.; Grochowski, C. Micro RNA Molecules as Modulators of Treatment Resistance, Immune Checkpoints Controllers and Sensitive Biomarkers in Glioblastoma Multiforme. Int. J. Mol. Sci. 2020, 21, 1507. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zhi, H.; Ma, D.; Li, T. MiR-217 Promoted the Proliferation and Invasion of Glioblastoma by Repressing YWHAG. Cytokine 2017, 92, 93–102. [Google Scholar] [CrossRef]
- Liang, H.Q.; Wang, R.J.; Diao, C.F.; Li, J.W.; Su, J.L.; Zhang, S. The PTTG1-Targeting MiRNAs MiR-329, MiR-300, MiR-381, and MiR-655 Inhibit Pituitary Tumor Cell Tumorigenesis and Are Involved in a P53/PTTG1 Regulation Feedback Loop. Oncotarget 2015, 6, 29413–29427. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Zhang, Y.; Cavazos, D.; Ma, X.; Zhao, Z.; Du, L.; Pertsemlidis, A. MIR-195 Targets Cyclin D3 and Survivin to Modulate the Tumorigenesis of Non-Small Cell Lung Cancer. Cell Death Dis. 2018, 9, 193. [Google Scholar] [CrossRef]
- Xu, Q.; Liu, L.Z.; Yin, Y.; He, J.; Li, Q.; Qian, X.; You, Y.; Lu, Z.; Peiper, S.C.; Shu, Y.; et al. Regulatory Circuit of PKM2/NF-ΚB/MIR-148a/152-Modulated Tumor Angiogenesis and Cancer Progression. Oncogene 2015, 34, 5482–5493. [Google Scholar] [CrossRef]
- Ye, J.; Li, L.; Feng, P.; Wan, J.; Li, J. Downregulation of MiR-34a Contributes to the Proliferation and Migration of Laryngeal Carcinoma Cells by Targeting Cyclin D1. Oncol. Rep. 2016, 36, 390–398. [Google Scholar] [CrossRef] [Green Version]
- Cao, P.; Feng, Y.; Deng, M.; Li, J.; Cai, H.; Meng, Q.; Fang, W.; Li, Y.; Ke, J.; Long, X. MiR-15b Is a Key Regulator of Proliferation and Apoptosis of Chondrocytes from Patients with Condylar Hyperplasia by Targeting IGF1, IGF1R and BCL2. Osteoarthr. Cartil. 2019, 27, 336–346. [Google Scholar] [CrossRef] [Green Version]
- Kouhkan, F.; Mobarra, N.; Soufi-Zomorrod, M.; Keramati, F.; Ali Hosseini Rad, S.M.; Fathi-Roudsari, M.; Tavakoli, R.; Hajarizadeh, A.; Ziaei, S.; Lahmi, R.; et al. MicroRNA-129-1 Acts as Tumour Suppressor and Induces Cell Cycle Arrest of GBM Cancer Cells through Targeting IGF2BP3 and MAPK1. J. Med. Genet. 2015, 53, 24–33. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, Q.; Chen, G.; Zhou, J.; Li, L.; Dong, Z.; Yang, R.; Xu, L.; Cui, H.; Xu, M.; Yi, L. Neurotensin Signaling Stimulates Glioblastoma Cell Proliferation by Upregulating C-Myc and Inhibiting MiR-29b-1 and MiR-129-3p. Neuro. Oncol. 2016, 18, 216–226. [Google Scholar] [CrossRef] [Green Version]
- Ofek, P.; Calderón, M.; Mehrabadi, F.S.; Krivitsky, A.; Ferber, S.; Tiram, G.; Yerushalmi, N.; Kredo-Russo, S.; Grossman, R.; Ram, Z.; et al. Restoring the Oncosuppressor Activity of MicroRNA-34a in Glioblastoma Using a Polyglycerol-Based Polyplex. Nanomed. Nanotechnol. Biol. Med. 2016, 12, 2201–2214. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Y.; Ye, Q.; Chen, C.; Wang, M.; Wang, H. Ezh2 Promotes Clock Function and Hematopoiesis Independent of Histone Methyltransferase Activity in Zebrafish. Nucleic Acids Res. 2018, 46, 3382–3399. [Google Scholar] [CrossRef]
- Shang, C.; Guo, Y.; Hong, Y.; Liu, Y.H.; Xue, Y. MiR-21 up-Regulation Mediates Glioblastoma Cancer Stem Cells Apoptosis and Proliferation by Targeting FASLG. Mol. Biol. Rep. 2015, 42, 721–727. [Google Scholar] [CrossRef]
- Wang, S.; Feng, L.; Hu, B.G.; Lu, Y.F.; Wang, W.M.; Guo, W.; Suen, C.W.; Jiao, B.H.; Pang, J.X.; Fu, W.M.; et al. MiR-133a Promotes TRAIL Resistance in Glioblastoma via Suppressing Death Receptor 5 and Activating NF-ΚB Signaling. Mol. Ther.-Nucleic Acids 2017, 8, 482–492. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhang, Y.; Yang, J.; Li, S.; Chen, J. Upregulation of MiR-582-5p Inhibits Cell Proliferation, Cell Cycle Progression and Invasion by Targeting Rab27a in Human Colorectal Carcinoma. Cancer Gene Ther. 2015, 22, 475–480. [Google Scholar] [CrossRef] [Green Version]
- Fu, Z.J.; Chen, Y.; Xu, Y.Q.; Lin, M.A.; Wen, H.; Chen, Y.T.; Pan, P.L. Regulation of MiR-30b in Cancer Development, Apoptosis, and Drug Resistance. Open Life Sci. 2022, 17, 102–106. [Google Scholar] [CrossRef]
- Duan, K.; Ge, Y.C.; Zhang, X.P.; Wu, S.Y.; Feng, J.S.; Chen, S.L.; Zhang, L.; Yuan, Z.H.; Fu, C.H. MiR-34a Inhibits Cell Proliferation in Prostate Cancer by Downregulation of SIRT1 Expression. Oncol. Lett. 2015, 10, 3223–3227. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhou, Q.; Miao, Y.; Tian, H.; Li, Y.; Feng, X.; Song, X. MiR-429 Induces Apoptosis of Glioblastoma Cell through Bcl-2. Tumor Biol. 2016, 37, 15607–15613. [Google Scholar] [CrossRef]
- Luo, B.; Zhang, J. MicroRNA-16 Inhibits the Migration and Invasion of Glioma Cell by Targeting Bcl-2 Gene. Trop. J. Pharm. Res. 2020, 19, 2499–2504. [Google Scholar] [CrossRef]
- Chung, H.J.; Choi, Y.E.; Kim, E.S.; Han, Y.H.; Park, M.J.; Bae, I.H. MiR-29b Attenuates Tumorigenicity and Stemness Maintenance in Human Glioblastoma Multiforme by Directly Targeting BCL2L2. Oncotarget 2015, 6, 18429–18444. [Google Scholar] [CrossRef]
- Wang, L.; Liu, Y.; Yu, Z.; Gong, J.; Deng, Z.; Ren, N.; Zhong, Z.; Cai, H.; Tang, Z.; Cheng, H.; et al. Mir-139-5p Inhibits Glioma Cell Proliferation and Progression by Targeting GABRA1. J. Transl. Med. 2021, 19, 213. [Google Scholar] [CrossRef]
- Cui, J.; Placzek, W.J. Post-Transcriptional Regulation of Anti-Apoptotic BCL2 Family Members. Int. J. Mol. Sci. 2018, 19, 308. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Zhu, X.; Sun, Y.; Liu, J. MicroRNA-141 Inhibits the Self-Renewal of Glioblastoma Stem Cells via Jaggedl. Mol. Med. Rep. 2017, 16, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.X.; Zhao, Z.Y.; Weng, G.H.; He, X.Y.; Wu, C.J.; Fu, C.Y.; Sui, Z.Y.; Ma, Y.S.; Liu, T. Upregulation of MiR-181a Suppresses the Formation of Glioblastoma Stem Cells by Targeting the Notch2 Oncogene and Correlates with Good Prognosis in Patients with Glioblastoma Multiforme. Biochem. Biophys. Res. Commun. 2017, 486, 1129–1136. [Google Scholar] [CrossRef] [PubMed]
- Fm, K.; La, H.; Wl, D.; Es, D.; Hua, Y.; Hao, L.; Cy, P.; Tj, M.; Ma, Q.; Ritner, C. Differentiation Programs in Glioblastoma. Cold Spring Harb. Lab. Press 2015, 2, 25838542. [Google Scholar] [CrossRef]
- Wong, H.K.A.; El Fatimy, R.; Onodera, C.; Wei, Z.; Yi, M.; Mohan, A.; Gowrisankaran, S.; Karmali, P.; Marcusson, E.; Wakimoto, H.; et al. The Cancer Genome Atlas Analysis Predicts MicroRNA for Targeting Cancer Growth and Vascularization in Glioblastoma. Mol. Ther. 2015, 23, 1234–1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, W.; Liu, B.; Qu, S.; Liang, G.; Luo, W.; Gong, C. MicroRNAs and Cancer: Key Paradigms in Molecular Therapy (Review). Oncol. Lett. 2018, 15, 2735–2742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Xue, Y.; Wang, P.; Zhu, J.; Ma, J. MIR-608 Inhibits the Migration and Invasion of Glioma Stem Cells by Targeting Macrophage Migration Inhibitory Factor. Oncol. Rep. 2016, 35, 2733–2742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, B.; Wang, Y.; Fang, X.; Gu, L.; Zhang, C. Targeting KLF6. Animals 2021, 11, 2–9. [Google Scholar]
- Yao, Y.L.; Ma, J.; Wang, P.; Xue, Y.X.; Li, Z.; Zhao, L.N.; Li, Z.Q.; Feng, T.D.; Liu, Y.H. MiR-101 Acts as a Tumor Suppressor by Targeting Kruppel-like Factor 6 in Glioblastoma Stem Cells. CNS Neurosci. Ther. 2015, 21, 40–51. [Google Scholar] [CrossRef]
- Yao, Y.; Ma, J.; Xue, Y.; Wang, P.; Li, Z.; Li, Z.; Hu, Y.; Shang, X.; Liu, Y. MiR-449a Exerts Tumor-Suppressive Functions in Human Glioblastoma by Targeting Myc-Associated Zinc-Finger Protein. Mol. Oncol. 2015, 9, 640–656. [Google Scholar] [CrossRef] [Green Version]
- Xi, Z.; Wang, P.; Xue, Y.; Shang, C.; Liu, X.; Ma, J.; Li, Z.; Li, Z.; Bao, M.; Liu, Y. Overexpression of MiR-29a Reduces the Oncogenic Properties of Glioblastoma Stem Cells by Downregulating Quaking Gene Isoform 6. Oncotarget 2017, 8, 24949–24963. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Chen, C.; Yu, S.Z.; Liu, Q.; Rao, J.; Zhang, H.R.; Xiao, H.L.; Fu, T.W.; Long, H.; He, Z.C.; et al. MiR-663 Suppresses Oncogenic Function of CXCR4 in Glioblastoma. Clin. Cancer Res. 2015, 21, 4004–4013. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Chen, R.; Zhou, X.; Guo, R.; Yin, J.; Li, Y.; Ma, G. MiR-137: A Novel Therapeutic Target for Human Glioma. Mol. Ther.-Nucleic Acids 2020, 21, 614–622. [Google Scholar] [CrossRef]
- Li, Q.; Cheng, Q.; Chen, Z.; Peng, R.; Chen, R.; Ma, Z.; Wan, X.; Liu, J.; Meng, M.; Peng, Z.; et al. MicroRNA-663 Inhibits the Proliferation, Migration and Invasion of Glioblastoma Cells via Targeting TGF-1. Oncol. Rep. 2016, 35, 1125–1134. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Liu, Z.; Peng, Y.; Yu, C. MicroRNA-181c Inhibits Glioblastoma Cell Invasion, Migration and Mesenchymal Transition by Targeting TGF-β Pathway. Biochem. Biophys. Res. Commun. 2016, 469, 1041–1048. [Google Scholar] [CrossRef]
- Jiang, C.; Shen, F.; Du, J.; Hu, Z.; Li, X.; Su, J.; Wang, X.; Huang, X. MicroRNA-564 Is Downregulated in Glioblastoma and Inhibited Proliferation and Invasion of Glioblastoma Cells by Targeting TGF-Β1. Oncotarget 2016, 7, 56200–56208. [Google Scholar] [CrossRef] [Green Version]
- Wei, F.; Wang, Q.; Su, Q.; Huang, H.; Luan, J.; Xu, X.; Wang, J. MiR-373 Inhibits Glioma Cell U251 Migration and Invasion by Down-Regulating CD44 and TGFBR2. Cell. Mol. Neurobiol. 2016, 36, 1389–1397. [Google Scholar] [CrossRef]
- Hu, S.; Chen, H.; Zhang, Y.; Wang, C.; Liu, K.; Wang, H.; Luo, J. MicroRNA-520c Inhibits Glioma Cell Migration and Invasion by the Suppression of Transforming Growth Factor-β Receptor Type 2. Oncol. Rep. 2017, 37, 1691–1697. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Li, J.; Qin, F.; Dai, S. MiR-152 as a Tumor Suppressor MicroRNA: Target Recognition and Regulation in Cancer. Oncol. Lett. 2016, 11, 3911–3916. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Sun, H.; Wang, X.; Hou, N.; Zhao, L.; Tong, D.; He, K.; Yang, Y.; Song, T.; Yang, J.; et al. EGR1 Mediates MiR-203a Suppress the Hepatocellular Carcinoma Cells Progression by Targeting HOXD3 through EGFR Signaling Pathway. Oncotarget 2016, 7, 45302–45316. [Google Scholar] [CrossRef] [Green Version]
- Shea, A.; Harish, V.; Afzal, Z.; Chijioke, J.; Kedir, H.; Dusmatova, S.; Roy, A.; Ramalinga, M.; Harris, B.; Blancato, J.; et al. MicroRNAs in Glioblastoma Multiforme Pathogenesis and Therapeutics. Cancer Med. 2016, 5, 1917–1946. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Kacimi, S.E.O.; Nguyen, T.L.; Suman, K.H.; Lemus-Martin, R.; Saleem, H.; Do, D.N. MiR-21 in the Cancers of the Digestive System and Its Potential Role as a Diagnostic, Predictive, and Therapeutic Biomarker. Biology 2021, 10, 417. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, B.; Shi, Y.; Xu, C.; Xiao, H.L.; Ma, L.N.; Xu, S.L.; Yang, L.; Wang, Q.L.; Dang, W.Q.; et al. Oncogenic MiR-20a and MiR-106a Enhance the Invasiveness of Human Glioma Stem Cells by Directly Targeting TIMP-2. Oncogene 2015, 34, 1407–1419. [Google Scholar] [CrossRef]
- Cai, J.J.; Qi, Z.X.; Chen, L.C.; Yao, Y.; Gong, Y.; Mao, Y. MiR-124 Suppresses the Migration and Invasion of Glioma Cells in Vitro via Capn4. Oncol. Rep. 2016, 35, 284–290. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.F.; Zhu, T.; Chen, J.; Liu, L.; Ouyang, R. Knockdown of Collagen α-1(Iii) Inhibits Glioma Cell Proliferation and Migration and Is Regulated by Mir128-3p. Oncol. Lett. 2018, 16, 1917–1923. [Google Scholar] [CrossRef] [Green Version]
- Liao, H.; Bai, Y.; Qiu, S.; Zheng, L.; Huang, L.; Liu, T.; Wang, X.; Liu, Y.; Xu, N.; Yan, X.; et al. MiR-203 Downregulation Is Responsible for Chemoresistance in Human Glioblastoma by Promoting Epithelial-Mesenchymal Transition via SNAI2. Oncotarget 2015, 6, 8914–8928. [Google Scholar] [CrossRef] [Green Version]
- Pang, H.; Zheng, Y.; Zhao, Y.; Xiu, X.; Wang, J. MiR-590-3p Suppresses Cancer Cell Migration, Invasion and Epithelial-Mesenchymal Transition in Glioblastoma Multiforme by Targeting ZEB1 and ZEB2. Biochem. Biophys. Res. Commun. 2015, 468, 739–745. [Google Scholar] [CrossRef]
- Xiao, B.; Zhou, X.; Ye, M.; Lv, S.; Wu, M.; Liao, C.; Han, L.; Kang, C.; Zhu, X. MicroRNA-566 Modulates Vascular Endothelial Growth Factor by Targeting von Hippel-Landau in Human Glioblastoma in Vitro and in Vivo. Mol. Med. Rep. 2016, 13, 379–385. [Google Scholar] [CrossRef] [Green Version]
- Salinas-Vera, Y.M.; Marchat, L.A.; Gallardo-Rincón, D.; Ruiz-García, E.; Astudillo-De la Vega, H.; Echavarría-Zepeda, R.; López-Camarillo, C. AngiomiRs: MicroRNAs Driving Angiogenesis in Cancer (Review). Int. J. Mol. Med. 2019, 43, 657–670. [Google Scholar] [CrossRef] [Green Version]
- Mazurek, M.; Grochowski, C.; Litak, J.; Osuchowska, I.; Maciejewski, R.; Kamieniak, P. Recent Trends of MicroRNA Significance in Pediatric Population Glioblastoma and Current Knowledge of Micro RNA Function in Glioblastoma Multiforme. Int. J. Mol. Sci. 2020, 21, 3046. [Google Scholar] [CrossRef]
- Marisetty, A.; Wei, J.; Kong, L.Y.; Ott, M.; Fang, D.; Sabbagh, A.; Heimberger, A.B. Mir-181 Family Modulates Osteopontin in Glioblastoma Multiforme. Cancers 2020, 12, 3813. [Google Scholar] [CrossRef]
- Liu, J.; Xu, J.; Li, H.; Sun, C.; Yu, L.; Li, Y.; Shi, C.; Zhou, X.; Bian, X.; Ping, Y.; et al. MiR-146b-5p Functions as a Tumor Suppressor by Targeting TRAF6 and Predicts the Prognosis of Human Gliomas. Oncotarget 2015, 6, 29129. [Google Scholar] [CrossRef] [Green Version]
- Kosaka, A.; Ohkuri, T.; Ikeura, M.; Kohanbash, G.; Okada, H. Transgene-Derived Overexpression of MiR-17-92 in CD8+ T-Cells Confers Enhanced Cytotoxic Activity. Biochem. Biophys. Res. Commun. 2015, 458, 549–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, M.; Wang, Y.Y.; Yoo, J.Y.; Oh, C.; Otani, Y.; Kang, J.M.; Park, E.S.; Kim, E.; Chung, S.; Jeon, Y.J.; et al. MicroRNA-138 Suppresses Glioblastoma Proliferation through Downregulation of CD44. Sci. Rep. 2021, 11, 9219. [Google Scholar] [CrossRef] [PubMed]
- Zingoni, A.; Molfetta, R.; Fionda, C.; Soriani, A.; Paolini, R.; Cippitelli, M.; Cerboni, C.; Santoni, A. NKG2D and Its Ligands: “One for All, All for One”. Front. Immunol. 2018, 9, 476. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Shi, Z.; Wei, W.; Lu, C.; Wei, Y.; Yan, W.; Li, R.; Zhang, J.; You, Y.; Wang, X. MiR-181b Suppress Glioblastoma Multiforme Growth through Inhibition of SP1-Mediated Glucose Metabolism. Cancer Cell Int. 2020, 20, 69. [Google Scholar] [CrossRef] [Green Version]
- Kang, K.; Zhang, J.; Zhang, X.; Chen, Z. MicroRNA-326 Inhibits Melanoma Progression by Targeting KRAS and Suppressing the AKT and ERK Signalling Pathways. Oncol. Rep. 2018, 39, 401–410. [Google Scholar] [CrossRef] [Green Version]
- Luan, W.; Wang, Y.; Chen, X.; Shi, Y.; Wang, J.; Zhang, J.; Qian, J.; Li, R.; Tao, T.; Wei, W.; et al. PKM2 Promotes Glucose Metabolism and Cell Growth in Gliomas through a Mechanism Involving a Let-7a/c-Myc/HnRNPA1 Feedback Loop. Oncotarget 2015, 6, 13006–13018. [Google Scholar] [CrossRef] [Green Version]
- Gu, J.J.; Gao, G.Z.; Zhang, S.M. MIR-218 Inhibits the Tumorgenesis and Proliferation of Glioma Cells by Targeting Robo1. Cancer Biomark. 2016, 16, 309–317. [Google Scholar] [CrossRef]
- Adhami, M.; MotieGhader, H.; Haghdoost, A.A.; Afshar, R.M.; Sadeghi, B. Gene Co-Expression Network Approach for Predicting Prognostic MicroRNA Biomarkers in Different Subtypes of Breast Cancer. Genomics 2020, 112, 135–143. [Google Scholar] [CrossRef]
- Liu, Z.; Jiang, Z.; Huang, J.; Huang, S. MiR-7 Inhibits Glioblastoma Growth by Simultaneously Interfering with the PI3K/ATK and Raf / MEK / ERK Pathways. Int. J. Oncol. 2014, 44, 1571–1580. [Google Scholar] [CrossRef] [Green Version]
- Aloizou, A.; Pateraki, G.; Siokas, V.; Mentis, A.A.; Liampas, I.; Lazopoulos, G.; Kovatsi, L.; Mitsias, P.D.; Bogdanos, D.P.; Paterakis, K.; et al. The Role of MiRNA-21 in Gliomas: Hope for a Novel Therapeutic Intervention? Toxicol. Rep. 2020, 7, 1514–1530. [Google Scholar] [CrossRef]
- Wang, G.B.; Liu, J.H.; Hu, J.; Xue, K. MiR-21 Enhanced Glioma Cells Resistance to Carmustine via Decreasing Spry2 Expression. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 5065–5071. [Google Scholar] [CrossRef]
- Bhere, D.; Arghiani, N.; Lechtich, E.R.; Yao, Y. Simultaneous Downregulation of MiR-21 and Upregulation of MiR-7 Has Anti-Tumor Efficacy. Sci. Rep. 2020, 10, 1779. [Google Scholar] [CrossRef] [Green Version]
- Müller, P.; Abdel Gaber, S.A.; Zimmermann, W.; Wittig, R.; Stepp, H. ABCG2 Influence on the Efficiency of Photodynamic Therapy in Glioblastoma Cells. J. Photochem. Photobiol. B Biol. 2020, 210, 111963. [Google Scholar] [CrossRef]
- Jeon, H.M.; Sohn, Y.W.; Oh, S.Y.; Kim, S.H.; Beck, S.; Kim, S.; Kim, H. ID4 Imparts Chemoresistance and Cancer Stemness to Glioma Cells by Derepressing MiR-9*-Mediated Suppression of SOX2. Cancer Res. 2011, 71, 3410–3421. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.; Wang, Z.; Liu, Q.; Liu, X.; Wu, M.; Li, G. Disturbing MiR-182 and -381 Inhibits BRD7 Transcription and Glioma Growth by Directly Targeting LRRC4. PLoS ONE 2014, 9, e84146. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, J.; Xu, G.; Wang, W.; Liu, C.; Yang, H.; Yu, Z.; Lei, Q.; Xiao, L.; Xiong, J.; et al. Targeting MiR-381-NEFL Axis Sensitizes Glioblastoma Cells to Temozolomide by Regulating Stemness Factors and Multidrug Resistance Factors. Oncotarget 2015, 6, 3147–3164. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Liu, Y.; Ren, J.; Deng, S.; Yi, G.; Guo, M.; Shu, S.; Zhao, L.; Peng, Y.; Qi, S. MiR-1268a Regulates ABCC1 Expression to Mediate Temozolomide Resistance in Glioblastoma. J. Neurooncol. 2018, 138, 499–508. [Google Scholar] [CrossRef]
- Wang, L.; Jiang, C.F.; Li, D.M.; Ge, X.; Shi, Z.M.; Li, C.Y.; Liu, X.; Yin, Y.; Zhen, L.; Liu, L.Z.; et al. MicroRNA-497 Inhibits Tumor Growth and Increases Chemosensitivity to 5-Fluorouracil Treatment by Targeting KSR1. Oncotarget 2016, 7, 2660–2671. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Li, N.; Yan, Z.; Li, C.; Zhao, Z. MiR-29a-Mediated CD133 Expression Contributes to Cisplatin Resistance in CD133+ Glioblastoma Stem Cells. J. Mol. Neurosci. 2018, 66, 369–377. [Google Scholar] [CrossRef]
- Li, D.M.; Chen, Q.D.; Wei, G.N.; Wei, J.; Yin, J.X.; He, J.H.; Ge, X.; Shi, Z.M. Hypoxia-Induced MiR-137 Inhibition Increased Glioblastoma Multiforme Growth and Chemoresistance Through LRP6. Front. Oncol. 2021, 10, 611699. [Google Scholar] [CrossRef]
- Miroshnichenko, S.; Patutina, O. Enhanced Inhibition of Tumorigenesis Using Combinations of MiRNA-Targeted Therapeutics. Front. Pharmacol. 2019, 10, 488. [Google Scholar] [CrossRef] [PubMed]
- Jesionek-Kupnicka, D.; Braun, M.; Trąbska-Kluch, B.; Czech, J.; Szybka, M.; Szymańska, B.; Kulczycka-Wojdala, D.; Bieńkowski, M.; Kordek, R.; Zawlik, I. MiR-21, miR-34a, miR-125b, miR-181d and miR-648 levels inversely correlate with MGMT and TP53 expression in primary glioblastoma patients. Arch. Med Sci. 2019, 15, 504–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Yu, J.; Zhao, C.; Ren, H.; Yuan, Z.; Zhang, B.; Zhuang, J.; Wang, J.; Feng, B. MiR-181b-5p Modulates Chemosensitivity of Glioma Cells to Temozolomide by Targeting Bcl-2. Biomed. Pharmacother. 2019, 109, 2192–2202. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.H.; Yao, Y.; Yun, Y.L.; Wang, M.L.; Zhu, R.X. MicroRNA-302c Enhances the Chemosensitivity of Human Glioma Cells to Temozolomide by Suppressing P-Gp Expression. Biosci. Rep. 2019, 39, BSR20190421. [Google Scholar] [CrossRef] [Green Version]
- Milani, R.; Brognara, E.; Fabbri, E.; Manicardi, A.; Corradini, R.; Finotti, A.; Gasparello, J.; Borgatti, M.; Cosenza, L.C.; Lampronti, I.; et al. Targeting MiR-155-5p and MiR-221-3p by Peptide Nucleic Acids Induces Caspase-3 Activation and Apoptosis in Temozolomide-resistant T98G Glioma Cells. Int. J. Oncol. 2019, 55, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Ananta, J.S.; Paulmurugan, R.; Massoud, T.F. Nanoparticle-Delivered Antisense MicroRNA-21 Enhances the Effects of Temozolomide on Glioblastoma Cells. Mol. Pharm. 2015, 12, 4509–4517. [Google Scholar] [CrossRef]
- Ma, W.; Cui, Y.; Liu, M.; Tan, Z.; Jiang, Y. Downregulation of MiR-125b Promotes Resistance of Glioma Cells to TRAIL through Overexpression of Tafazzin Which Is a Mitochondrial Protein. Aging 2019, 11, 2670–2680. [Google Scholar] [CrossRef]
- Shirjang, S.; Mansoori, B.; Asghari, S.; Duijf, P.H.; Mohammadi, A.; Gjerstorff, M.; Baradaran, B. Corrigendum to “MicroRNAs in Cancer Cell Death Pathways: Apoptosis and Necroptosis” [Free Radic. Biol. Med. 139 (2019) 1–15]. Free Radic. Biol. Med. 2020, 146, 402. [Google Scholar] [CrossRef]
- Rezaei, T.; Hejazi, M.; Mansoori, B.; Mohammadi, A.; Amini, M.; Mosafer, J.; Rezaei, S.; Mokhtarzadeh, A.; Baradaran, B. MicroRNA-181a Mediates the Chemo-Sensitivity of Glioblastoma to Carmustine and Regulates Cell Proliferation, Migration, and Apoptosis. Eur. J. Pharmacol. 2020, 888, 173483. [Google Scholar] [CrossRef]
- Mondal, P.; Meeran, S.M. MicroRNAs in Cancer Chemoresistance: The Sword and the Shield. Non-Coding RNA Res. 2021, 6, 200–210. [Google Scholar] [CrossRef]
- Lee, S.Y. Temozolomide Resistance in Glioblastoma Multiforme. Genes Dis. 2016, 3, 198–210. [Google Scholar] [CrossRef] [Green Version]
- Tan, A.C.; Ashley, D.M.; López, G.Y.; Malinzak, M.; Friedman, H.S.; Khasraw, M. Management of Glioblastoma: State of the Art and Future Directions. CA Cancer J. Clin. 2020, 70, 299–312. [Google Scholar] [CrossRef]
- Alnahhas, I.; Alsawas, M.; Rayi, A.; Palmer, J.D.; Raval, R.; Ong, S.; Giglio, P.; Murad, M.H.; Puduvalli, V. Characterizing Benefit from Temozolomide in MGMT Promoter Unmethylated and Methylated Glioblastoma: A Systematic Review and Meta-Analysis. Neuro-Oncol. Adv. 2020, 2, vdaa082. [Google Scholar] [CrossRef]
- Singh, N.; Miner, A.; Hennis, L.; Mittal, S. Mechanisms of Temozolomide Resistance in Glioblastoma—A Comprehensive Review. Cancer Drug Resist. 2021, 4, 17–43. [Google Scholar] [CrossRef]
- Yang, L.; Ma, Y.; Xin, Y.; Han, R.; Li, R.; Hao, X. Role of the MicroRNA 181 Family in Glioma Development. Mol. Med. Rep. 2018, 17, 322–329. [Google Scholar] [CrossRef] [Green Version]
- Nie, E.; Jin, X.; Wu, W.; Yu, T.; Zhou, X.; Shi, Z.; Zhang, J.; Liu, N.; You, Y. MiR-198 Enhances Temozolomide Sensitivity in Glioblastoma by Targeting MGMT. J. Neurooncol. 2017, 133, 59–68. [Google Scholar] [CrossRef]
- Gao, Y.T.; Chen, X.B.; Liu, H.L. Up-Regulation of MiR-370-3p Restores Glioblastoma Multiforme Sensitivity to Temozolomide by Influencing MGMT Expression. Sci. Rep. 2016, 6, 32972. [Google Scholar] [CrossRef] [Green Version]
- Serrano-Heras, G.; Castro-Robles, B.; Romero-Sánchez, C.M.; Carrión, B.; Barbella-Aponte, R.; Sandoval, H.; Segura, T. Involvement of N-Methylpurine DNA Glycosylase in Resistance to Temozolomide in Patient-Derived Glioma Cells. Sci. Rep. 2020, 10, 22185. [Google Scholar] [CrossRef]
- Zeng, A.; Wei, Z.; Yan, W.; Yin, J.; Huang, X.; Zhou, X.; Li, R.; Shen, F.; Wu, W.; Wang, X.; et al. Exosomal Transfer of MiR-151a Enhances Chemosensitivity to Temozolomide in Drug-Resistant Glioblastoma; Elsevier B.V.: Amsterdam, The Netherlands, 2018; Volume 436, ISBN 8613813879. [Google Scholar]
- Asadzadeh, Z.; Duijf, P.H.G.; Asgarzadeh, K.H. MicroRNAs in Cancer Stem Cells: Biology, Pathways, and Therapeutic Opportunities. J. Cell. Physiol. 2018, 234, 10002–10017. [Google Scholar] [CrossRef]
- Jiapaer, S.; Furuta, T.; Tanaka, S.; Kitabayashi, T.; Nakada, M. Potential Strategies Overcoming the Temozolomide Resistance for Glioblastoma. Neurol. Med. Chir. 2018, 58, 405–421. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Z.X.; Yin, W.B.; Wang, Z.Y. MicroRNA-132 Induces Temozolomide Resistance and Promotes the Formation of Cancer Stem Cell Phenotypes by Targeting Tumor Suppressor Candidate 3 in Glioblastoma. Int. J. Mol. Med. 2017, 40, 1307–1314. [Google Scholar] [CrossRef] [Green Version]
- Mahinfar, P.; Baradaran, B.; Davoudian, S.; Vahidian, F.; Cho, W.C.S.; Mansoori, B. Long Non-Coding RNAs in Multidrug Resistance of Glioblastoma. Genes 2021, 12, 455. [Google Scholar] [CrossRef]
- Wang, L.; Su, J.; Zhao, Z.; Hou, Y.; Yin, X.; Zheng, N.; Zhou, X.; Yan, J.; Xia, J.; Wang, Z. MiR-26b Reverses Temozolomide Resistance via Targeting Wee1 in Glioma Cells. Cell Cycle 2017, 16, 1954–1964. [Google Scholar] [CrossRef]
- Li, X.; Wang, H.; Wu, Z.; Yang, T.; Zhao, Z.; Chen, G.; Xie, X.; Li, B.; Wei, Y.; Huang, Y.; et al. MiR-494-3p Regulates Cellular Proliferation, Invasion, Migration, and Apoptosis by PTEN/AKT Signaling in Human Glioblastoma. Cell. Mol. Neurobiol. 2015, 10, 679–687. [Google Scholar] [CrossRef] [Green Version]
- Qiao, W.; Guo, B.; Zhou, H.; Xu, W.; Chen, Y.; Liang, Y.; Dong, B. MiR-124 Suppresses Glioblastoma Growth and Potentiates Chemosensitivity by Inhibiting AURKA. Biochem. Biophys. Res. Commun. 2017, 486, 43–48. [Google Scholar] [CrossRef]
- Munoz, J.L.; Rodriguez-Cruz, V.; Ramkissoon, S.H.; Ligon, K.L.; Greco, S.J.; Rameshwar, P. Temozolomide Resistance in Glioblastoma Occurs by MiRNA-9-Targeted PTCH1, Independent of Sonic Hedgehog Level. Oncotarget 2015, 6, 1190–1201. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.H.; Cheng, C.H.; Shih, C.M.; Ho, K.H.; Lin, C.W.; Lee, C.C.; Liu, A.J.; Chang, C.K.; Chen, K.C. The Inhibition of MicroRNA-128 on IGF-1-Activating MTOR Signaling Involves in Temozolomide-Induced Glioma Cell Apoptotic Death. PLoS ONE 2016, 11, e167096. [Google Scholar] [CrossRef]
- Toraih, E.A.; Aly, N.M.; Abdallah, H.Y.; Al-Qahtani, S.A.; Shaalan, A.A.M.; Hussein, M.H.; Fawzy, M.S. MicroRNA–Target Cross-Talks: Key Players in Glioblastoma Multiforme. Tumor Biol. 2017, 39. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Yang, L.; Liao, F.; Wang, W.; Wang, Z.F. MiR-450a-5p Strengthens the Drug Sensitivity of Gefitinib in Glioma Chemotherapy via Regulating Autophagy by Targeting EGFR. Oncogene 2020, 39, 6190–6202. [Google Scholar] [CrossRef]
- Oprita, A.; Baloi, S.C.; Staicu, G.A.; Alexandru, O.; Tache, D.E.; Danoiu, S.; Micu, E.S.; Sevastre, A.S. Updated Insights on EGFR Signaling Pathways in Glioma. Int. J. Mol. Sci. 2021, 22, 587. [Google Scholar] [CrossRef]
- Chen, Y.; Li, R.; Pan, M.; Shi, Z.; Yan, W.; Liu, N.; You, Y.; Zhang, J.; Wang, X. MiR-181b Modulates Chemosensitivity of Glioblastoma Multiforme Cells to Temozolomide by Targeting the Epidermal Growth Factor Receptor. J. Neurooncol. 2017, 133, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Zeng, A.; Zhang, Z.; Shi, Z.; Yan, W.; You, Y. Exosomal Transfer of MiR-1238 Contributes to Temozolomide-Resistance in Glioblastoma. EBioMedicine 2019, 42, 238–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Zhang, J.; Zhang, J.; Qiu, W.; Xu, S.; Yu, Q.; Liu, C.; Wang, Y.; Lu, A.; Zhang, J.; et al. MicroRNA-625 Inhibits the Proliferation and Increases the Chemosensitivity of Glioma by Directly Targeting AKT2. Am. J. Cancer Res. 2017, 7, 1835–1849. [Google Scholar] [PubMed]
- Li, S.; Zeng, A.; Hu, Q.; Yan, W.; Liu, Y.; You, Y. MiR-423-5p Contributes to a Malignant Phenotype and Temozolomide Chemoresistance in Glioblastomas. Neuro. Oncol. 2017, 19, 55–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, W.; Yan, D.; Song, Z.; Zhu, X.; Liu, X.; Li, X.; Zhao, S. MiR-126-3p Sensitizes Glioblastoma Cells to Temozolomide by Inactivating Wnt/β-Catenin Signaling via Targeting SOX2. Life Sci. 2019, 226, 98–106. [Google Scholar] [CrossRef]
- Zhou, Y.; Liu, Y.; Hu, C.; Jiang, Y. MicroRNA-16 Inhibits the Proliferation, Migration and Invasion of Glioma Cells by Targeting Sal-like Protein 4. Int. J. Mol. Med. 2016, 38, 1768–1776. [Google Scholar] [CrossRef] [Green Version]
- Ye, Q.; Liu, K.; Shen, Q.; Li, Q.; Hao, J.; Han, F.; Jiang, R.W. Reversal of Multidrug Resistance in Cancer by Multi-Functional Flavonoids. Front. Oncol. 2019, 9, 487. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Li, H.; Li, Y.; Ruan, Y.; Quan, C. Identification of Genes and Pathways Associated with MDR in MCF-7/MDR Breast Cancer Cells by RNA-Seq Analysis. Mol. Med. Rep. 2018, 17, 6211–6226. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Mechanisms of MDR development in glioblastoma. A. MutantTP53; the two accepted pathways leading to TP53-mediated apoptosis are exogenous and endogenous. In endogenous apoptosis, TP53 activates pro-apoptotic proteins PUMA, FAS, and BAX. In response to activation of these proteins, TP53 binds to apoptotic proteins from the Bcl-2 family and finally induces cytochrome c/Apaf-1-dependent endogenous apoptosis. In contrast, mutations in the TP53 gene in cancers such as glioblastoma can cause PUMA, FAS, and BAX proteins to bind to anti-apoptotic proteins and result in the development of MDR. B. EMT; EMT is a subprocess that occurs during the progression of cancer which alters the morphology of cancer cells into highly motile and elongated mesenchymal-like cells which increases the capacity of tumor cells to resist chemotherapy. C. DNA repair; many chemotherapeutic drugs damage DNA in a manner that causes cell cycle arrest and cell death. Therefore, DNA repair involves an intricate network of repairing the cell, and that process leads to the development of MDR. D. P-gp /ABCB1 as a transmembrane protein causing lowered drug accumulation inside cells and consequently diminished drug efficacy. E. In glioblastoma stem-like cells (GSC), Notch1 and PRKCI are overexpressed and are associated with MDR. F. CYP (17A1) as a drug metabolizer overexpressed in glioblastoma and efflux the chemotropic drug through ABCC1 transporter.
Figure 1.
Mechanisms of MDR development in glioblastoma. A. MutantTP53; the two accepted pathways leading to TP53-mediated apoptosis are exogenous and endogenous. In endogenous apoptosis, TP53 activates pro-apoptotic proteins PUMA, FAS, and BAX. In response to activation of these proteins, TP53 binds to apoptotic proteins from the Bcl-2 family and finally induces cytochrome c/Apaf-1-dependent endogenous apoptosis. In contrast, mutations in the TP53 gene in cancers such as glioblastoma can cause PUMA, FAS, and BAX proteins to bind to anti-apoptotic proteins and result in the development of MDR. B. EMT; EMT is a subprocess that occurs during the progression of cancer which alters the morphology of cancer cells into highly motile and elongated mesenchymal-like cells which increases the capacity of tumor cells to resist chemotherapy. C. DNA repair; many chemotherapeutic drugs damage DNA in a manner that causes cell cycle arrest and cell death. Therefore, DNA repair involves an intricate network of repairing the cell, and that process leads to the development of MDR. D. P-gp /ABCB1 as a transmembrane protein causing lowered drug accumulation inside cells and consequently diminished drug efficacy. E. In glioblastoma stem-like cells (GSC), Notch1 and PRKCI are overexpressed and are associated with MDR. F. CYP (17A1) as a drug metabolizer overexpressed in glioblastoma and efflux the chemotropic drug through ABCC1 transporter.

Figure 2.
Treatment strategy based on miR-21 and miR-7 in glioblastoma. MiR-targeting therapy may involve activating or upregulating tumor suppressor miRs and inhibiting the function of oncomiRs. MiR-21 is a key oncomiR, which is overexpressed in Glioblastoma. In contrast, miR-7 is downregulated in Glioblastoma. Suppression of miR-21 and upregulation of miR-7 are crucial to targeting complementary pathways which inhibit glioblastoma growth and development. Assembling miRNA inhibitors or mimicking miRNAs to appropriate carriers including liposomes, extracellular vesicle (EVs), polymer-mediated delivery systems, viral vectors (VV) such as adenoviruses, and cell-based delivery systems and bacteriophage-based virus-like particles (VLPs) potentially inhibiting the function of the oncomiRs.
Figure 2.
Treatment strategy based on miR-21 and miR-7 in glioblastoma. MiR-targeting therapy may involve activating or upregulating tumor suppressor miRs and inhibiting the function of oncomiRs. MiR-21 is a key oncomiR, which is overexpressed in Glioblastoma. In contrast, miR-7 is downregulated in Glioblastoma. Suppression of miR-21 and upregulation of miR-7 are crucial to targeting complementary pathways which inhibit glioblastoma growth and development. Assembling miRNA inhibitors or mimicking miRNAs to appropriate carriers including liposomes, extracellular vesicle (EVs), polymer-mediated delivery systems, viral vectors (VV) such as adenoviruses, and cell-based delivery systems and bacteriophage-based virus-like particles (VLPs) potentially inhibiting the function of the oncomiRs.

Figure 3.
Treatment strategies for CD133+ cisplatin-resistant glioblastoma cells. CD133 as pentaspan transmembrane glycoprotein localized in protrusions of normal and cancer cells. Positive (CD133+) cells are regarded as tumor initiation cells in GBM. High CD133 expression confers resistance to glioma therapy and causes tumor growth. MiR-29a as a specific miRNA directly regulates CD133 expression. In fact, miR-29a promotes cisplatin-induced apoptosis via suppressing CD133 expression. Furthermore, using combination therapy including overexpression of miR-29a and cisplatin treatment substantially suppressed tumor growth in GBM than cisplatin treatment alone.
Figure 3.
Treatment strategies for CD133+ cisplatin-resistant glioblastoma cells. CD133 as pentaspan transmembrane glycoprotein localized in protrusions of normal and cancer cells. Positive (CD133+) cells are regarded as tumor initiation cells in GBM. High CD133 expression confers resistance to glioma therapy and causes tumor growth. MiR-29a as a specific miRNA directly regulates CD133 expression. In fact, miR-29a promotes cisplatin-induced apoptosis via suppressing CD133 expression. Furthermore, using combination therapy including overexpression of miR-29a and cisplatin treatment substantially suppressed tumor growth in GBM than cisplatin treatment alone.


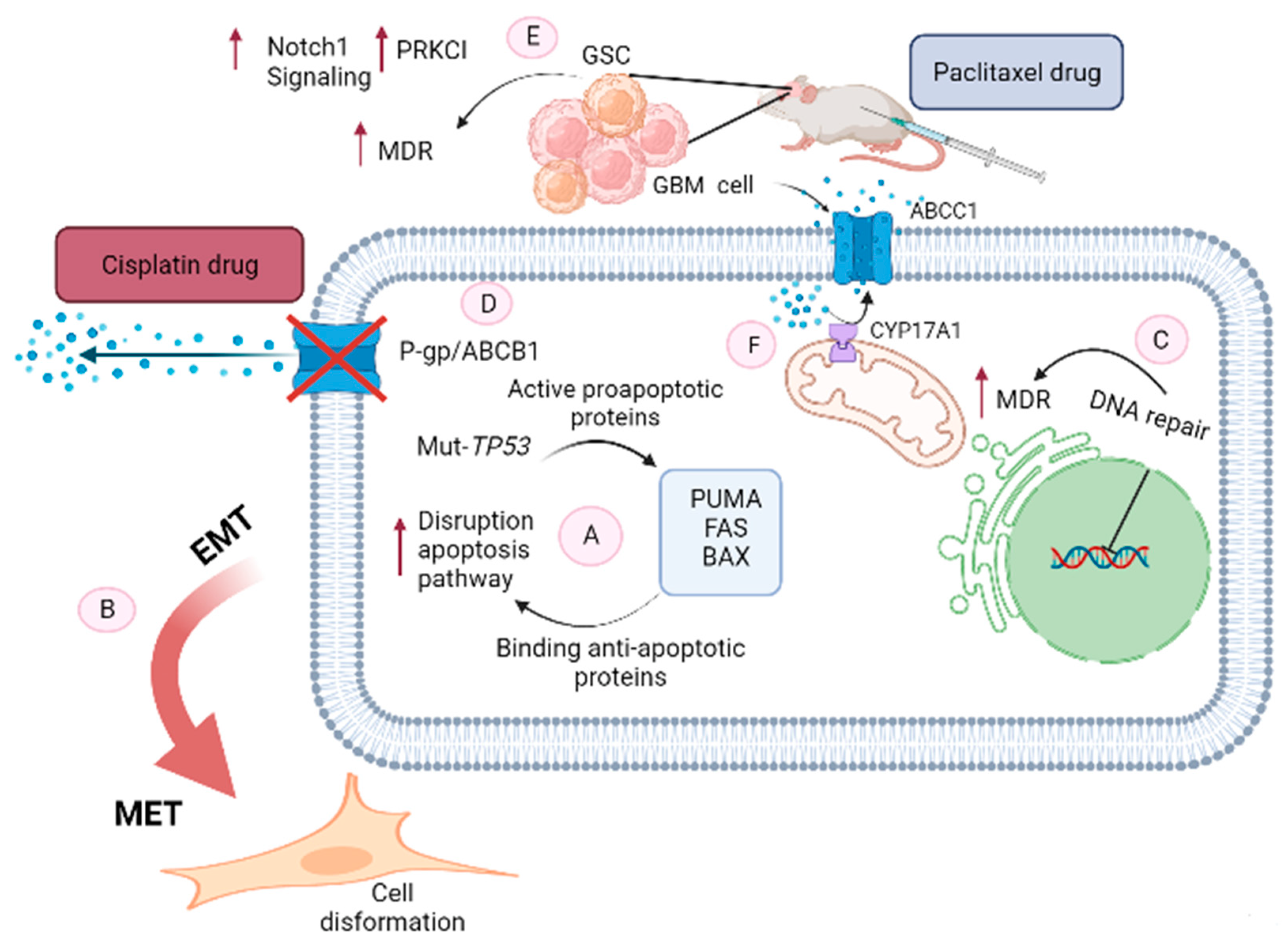{kind=link}
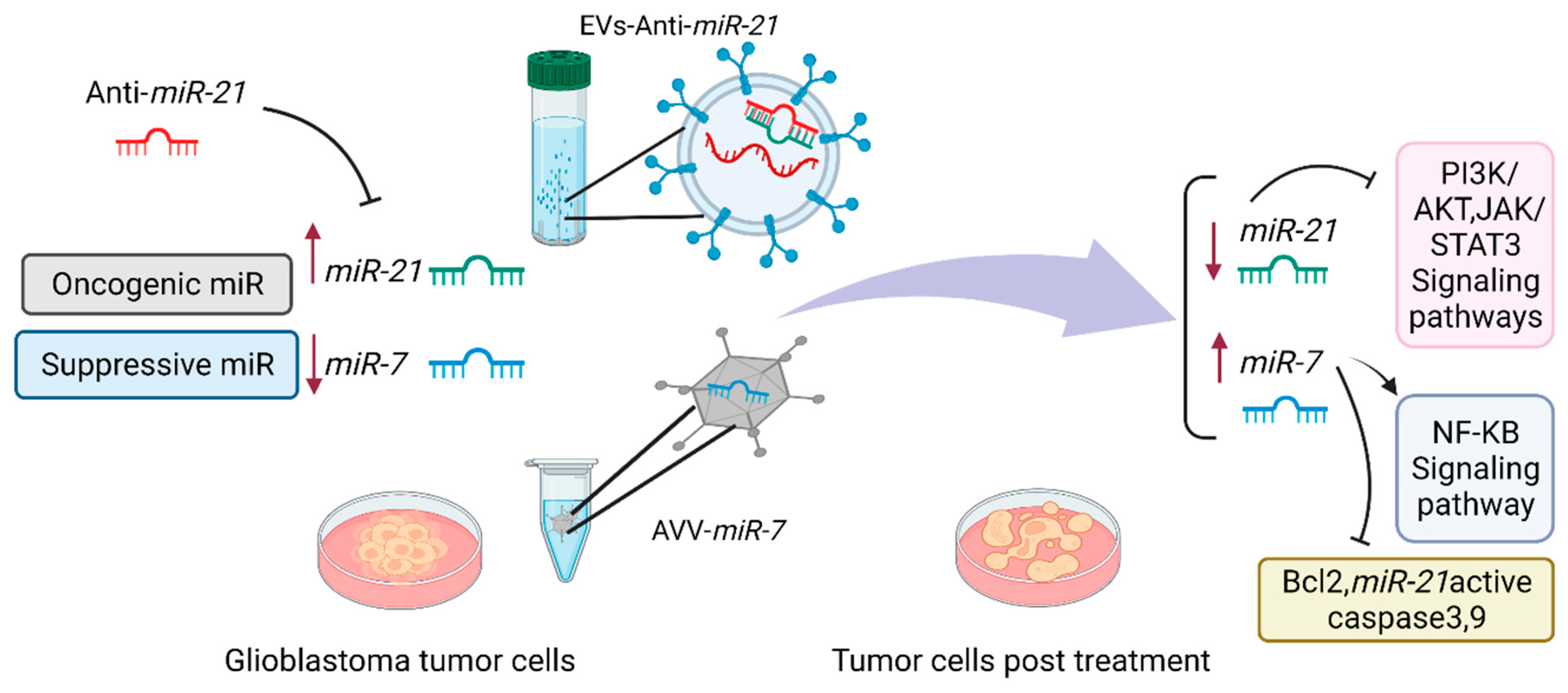{kind=link}
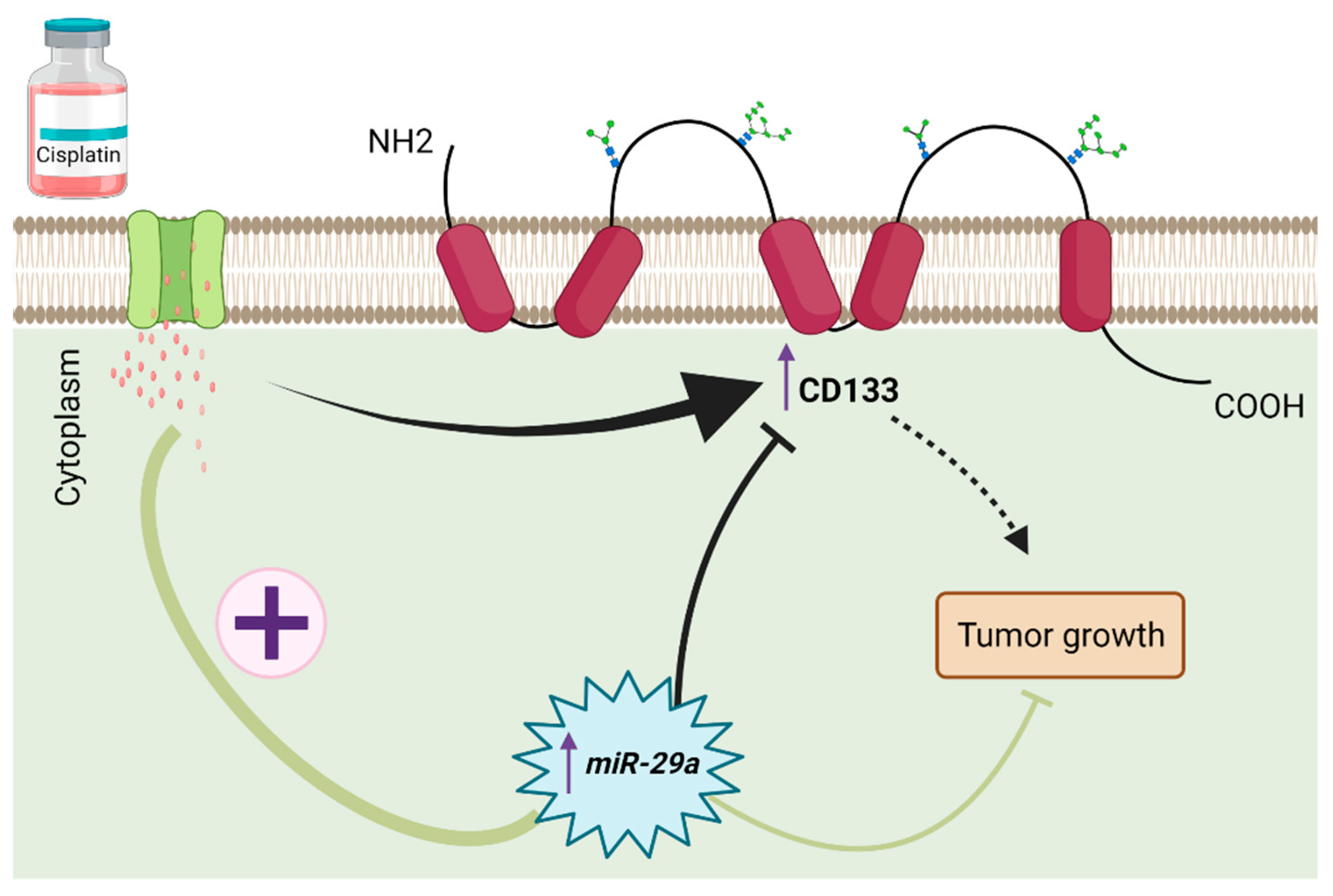{kind=link}
Table 1.
Involvement of miRNAs in glioblastoma.
| miRNA | Expression in Glioblastoma | Targets Genes | Effects | Ref. |
|---|---|---|---|---|
| Facilitate cell growth and proliferation in glioblastoma | ||||
| Sustaining Proliferative Signaling | ||||
| miR-7 | Down-regulated | EGFR | transfection with miR-7 decreased viability and invasiveness | [54] |
| AKT | ||||
| miR-128 | Down-regulated | EGFR | correlates with aggressive human glioma subtypes | [45] |
| PDGR | miR-128 inhibits growth and mediates differentiation | |||
| miR-133 | Down-regulated | EGFR | decreased cell growth and increased cell apoptosis | [55] |
| miR-218 | Down-regulated | RTK | decreased tumor burden and reduced survival | [45] |
| HIF2α | ||||
| miR-219-5p | Down-regulated | MAPK | inhibit the proliferation, anchorage-independent growth and migration | [56] |
| PI3K | ||||
| EGFR | ||||
| miR-491-5p and -3p | Down-regulated | EGFR | decrease cellular proliferation and invasion | [57] |
| CDK6 | inhibit the propagation of glioma stem cells | |||
| BCL-XL | ||||
| IGFBP2 | ||||
| CDK6 | ||||
| miR-7 | Down-regulated | PI3K | Inhibit cell cycle and cell growth | [58] |
| AKT | ||||
| RAF-1 | ||||
| MEK 1/2 | ||||
| cyclin D1 | ||||
| EGFR | ||||
| miR-34a | Down-regulated | SMAD4 | Decrease cellular proliferation and invasion | [59] |
| PDGFRA | miR-34a expression level is shown to be prognostic | |||
| miR-218 | Down-regulated | EGFR | Decrease cellular proliferation | [60] |
| PLCγ1 | ||||
| PIK3CA | ||||
| ARAF | ||||
| PDGFRα | ||||
| RSK2 | ||||
| S6K1 | ||||
| STAT3 | ||||
| BCLAF1 | ||||
| miR-410, miR-144-3p, and miR-34a | Down-regulated | C-MET | The overexpression of these miRs produces anti-proliferative effects | [58,61] |
| MiR-126, let-7a, and miR-622 | Up-regulated | KRAS | restrain glioma cells’ proliferation | [62] |
| MiR-124 | Up-regulated | R-RAS | governs glioma growth and angiogenesis and enhances chemosensitivity | [63] |
| N-RAS | ||||
| MiR-143 | Down-regulated | N-RAS | acts as a tumor suppressor | [62] |
| enhances temozolomide-induced apoptosis in glioma | ||||
| let-7e | Down-regulated | N-RAS | Repress tumor function by decreasing proliferation, migration and invasion while promoting apoptosis | [64] |
| miR-17-5p, miR-19a/b, miR-21, miR-1908, miR-494-3p, miR-10a/10b, miR-23a, and miR-26a | Down-regulated | PTEN | Inhibit tumor growth | [65,66] |
| miR-542-3p | Down-regulated | AKT | Suppress tumor cell proliferation and invasion | [67] |
| MiR-199a-3p | Down-regulated | mTORC1 and mTORC2 | decrease glioma cell proliferation | [68] |
| MiR-34a | Down-regulated | RICTOR | Inhibit cell proliferation and tumor growth of glioma stem cells | [69] |
| AKT | ||||
| WNT signaling | ||||
| Evading Growth Suppressors | ||||
| miR-10b | Up-regulated | TP53 | promotes growth, invasiveness, and angiogenesis and inhibits apoptosis | [70] |
| MMP14 | ||||
| UPAR | ||||
| RHOC | ||||
| HOXD10 | ||||
| miR-25 and -32 | Down-regulated | TP53 | inhibited growth of the glioblastoma multiforme cells | [71] |
| MTOR | ||||
| MDM2 | ||||
| TSC1 | ||||
| miR-17 | Up-regulated | PTEN | promoted cell motility, invasion, and tube-like structure formation | [72] |
| HIF1 | ||||
| VEGF | ||||
| MiR-217 | Up-regulated | YWHAG | enhances the proliferation of cells | [73] |
| MDM4 | ||||
| TP53 | ||||
| MiR-26a | Up-regulated | RB | promotes GBM formation | [71] |
| MiR-329 and miR-320 | Down-regulated | E2F1 | inhibit cell proliferation | [74] |
| MiR-195 | Down-regulated | CCND1 | block GBM cell proliferation by inducing G1-S arrest | [75] |
| CCNE1 | ||||
| let-7b, miR-15b, miR-34a, and miR-340 | Down-regulated | CCND1 | block cell cycle and proliferation | [76,77,78] |
| miR-34a, miR-107, miR-138, miR129-3p, miR-29b-1, miR-218, miR-129-1, miR-340, miR-491-3p/5p | Down-regulated | CDK6 | inhibit cell cycle of GBM cells | [45,76,79,80,81] |
| MiR-10b | Down-regulated | CDKN2A/p16INK4A | arrest the cell cycle | [82] |
| miR-138 | Down-regulated | CDK6 | inhibit cell proliferation | [82] |
| EZH2 | ||||
| PRB-E2F1 | ||||
| Resisting Cell Death | ||||
| miR-21 | Up-regulated | FASL | suppressed the apoptosis in GBM stem cells (GSCs) | [83] |
| miR-133a | Up-regulated | DR5 | suppressed the cells’ apoptosis | [84] |
| miR-363 and miR-582-5p | Up-regulated | BIM | inhibit GSC apoptosis to promote GSC growth | [85] |
| CASPASE-3 | ||||
| CASPASE-9 | ||||
| miR-21 and miR-30b/c | Up-regulated | CASPASE-3 | prominently inhibited TRAIL-induced apoptosis | [86] |
| TAP63 | ||||
| miR-148a, miR-363, miR-92a | Up-regulated | BIM | decrease apoptosis | [85] |
| MiR-16, miR-34a, and miR-429 | Down-regulated | BCL-2 | increase apoptosis to suppress proliferation in glioma cells | [87,88,89] |
| MiR-29b | Down-regulated | BCL2L2 | induce apoptosis in GBM cells | [90] |
| MiR-139 | Down-regulated | MCL-1 | promoted apoptosis related to TMZ | [91] |
| miR-153 | Down-regulated | MCL-1, BCL-2, and IRS-2 | inhibited survival and promoted apoptosis | [92] |
| Enabling Replicative Immortality | ||||
| miR-141 | Down-regulated | JAGGED1 | suppressed the self-renewal of GSCs | [93] |
| miR-181a | Down-regulated | NOTCH2 | suppressed GSCs formation and proliferation and increased apoptosis of GSCs | [94] |
| miR-182 | Down-regulated | HIF2α | reduced the aggressive phenotype of GSCs | [95] |
| miR-148a and miR-31 | Up-regulated | HIF1α | activated Notch signaling to maintain potential of GSCs | [96] |
| HIF1AN | ||||
| miR-128 | Down-regulated | PDGFRα | significantly suppressed proliferation in GSCs | [97] |
| EGFR | ||||
| miR-608 | Down-regulated | MIF | suppressed proliferation, invasion and promoted apoptosis in GSCs | [98] |
| MiR-152 | Down-regulated | KLF4 | exerts tumor-suppressive effects | [99] |
| miR-101 miR-608 | Down-regulated | KLF6 | exerts tumor-suppressive effects | [100] |
| MiR-449a | Down-regulated | MAZ | blocks proliferation and induces apoptosis in GSCs | [101] |
| miR-29a | Down-regulated | QKI-6 | repressed the malignant behavior of GSCs | [102] |
| WTAP | ||||
| miR-663 | Down-regulated | CXCR4 | effectively suppressed the invasion and proliferation of GBM cells | [103] |
| miR-137 | Down-regulated | RTVP-1 | inhibited the self-renewal of GSCs | [104] |
| Activating Invasion and Metastasis | ||||
| miR-663, miRNA-181c, and miR-564 | Down-regulated | TGF-β1 | suppress the invasion. | [105,106,107] |
| MiR-373 and miR-520c | Down-regulated | TGFBR2 | suppress the invasion of GBM | [108,109] |
| miR-211 and miR-491-5p | Down-regulated | MMP-9 | suppress the invasion of GBM | [58] |
| MiR-152 | Down-regulated | MMP-3 | suppresses the invasion of GBM | [110] |
| MiR-16 | Down-regulated | NF-κB1 | inhibits the invasion of GBM cells. | [89] |
| MiR-203 | Down-regulated | ROBO1 | suppresses the migration of glioma cells | [111] |
| ERK | ||||
| MMP-9 | ||||
| miR-218 | Down-regulated | MMP-9 | inhibit the invasion of GBM | [112] |
| LEF1 | ||||
| MiR-7 | Down-regulated | FAK | suppresses the invasion of GBM | [112] |
| MiR-21 | Up-regulated | TIMP-3 and RECK | enhances the expression of MMPs and glioma cell invasion | [113] |
| miR-20a and miR-106a | Up-regulated | TIMP-2 | increase the invasion of GBM | [114] |
| MiR-124 | Down-regulated | CAPN4 | inhibited the invasion of glioma cells. | [115] |
| Let-7g-5p | Down-regulated | VSIG4 | inhibited GBM cell invasion and migration | [116] |
| MiR-203 | Down-regulated | SNAI2 | inhibits the invasive and migratory capacity of GBM cells | [117] |
| MiR-590-3p | Down-regulated | ZEB1 and ZEB2 | suppresses the invasive and migratory capacity of GBM cells | [118] |
| Inducing Angiogenesis | ||||
| MiR-23b, miR-566 | Down-regulated | VEGF-A | decrease angiogenesis capacity in tumor | [112,119] |
| VHL | ||||
| MiR-128 | Down-regulated | P70S6K1 | suppress tumor angiogenesis | [120] |
| MiR-125b | Down-regulated | MAZ | inhibited VEGF-mediated angiogenesis | [121] |
| Evading Immune Destruction | ||||
| miR-124 | Down-regulated | STAT3 | reversed immune suppression associated with T cells | [122] |
| Exhaustion of T cells (CD4+ and CD8+) completely counteracted the role of miR-124 in the inhibiting glioma | ||||
| miR-146b-5p | Down-regulated | TRAF6 | Regulate immunosuppression and macrophage polarization | [123] |
| miR-31 | TRADD | |||
| miR-17-92 | Down-regulated | TGFBR2 | activated CD8+ CD44+ memory T cells (Th1 CD8+ T cells) which then secreted IFN-γ and increased resistance of CD8+ T cells to the immunosuppressive effects of TGF-β1 | [124] |
| miR-138 | Down-regulated | CTLA-4 | reversed the immunosuppressive effects to exert antitumor properties | [125] |
| PD-1 | ||||
| miR-20a, miR-93, and miR-106b | Down-regulated | NKG2DL | improved NK cell-mediated cytotoxicity | [126] |
| Reprogramming Cellular Energetics | ||||
| miR-106a | Down-regulated | GLUT3 | blocked glucose uptake in GBM cells | [127] |
| miR-143 | Down-regulated | HK2 | suppressed glycolysis | [112] |
| miR-326 | Down-regulated | PKM2 | suppressed glycolysis | [128] |
| let-7a | C-MYC | inhibit glucose metabolism and the growth of gliomas | [129] | |
| HNRNPA1 | ||||
| PKM2 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mahinfar, P.; Mansoori, B.; Rostamzadeh, D.; Baradaran, B.; Cho, W.C.; Mansoori, B. The Role of microRNAs in Multidrug Resistance of Glioblastoma. Cancers 2022, 14, 3217. https://doi.org/10.3390/cancers14133217
AMA Style
Mahinfar P, Mansoori B, Rostamzadeh D, Baradaran B, Cho WC, Mansoori B. The Role of microRNAs in Multidrug Resistance of Glioblastoma. Cancers. 2022; 14(13):3217. https://doi.org/10.3390/cancers14133217
Chicago/Turabian StyleMahinfar, Parvaneh, Behnaz Mansoori, Davoud Rostamzadeh, Behzad Baradaran, William C. Cho, and Behzad Mansoori. 2022. "The Role of microRNAs in Multidrug Resistance of Glioblastoma" Cancers 14, no. 13: 3217. https://doi.org/10.3390/cancers14133217
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.