
SIRT1-SIRT7 Expression in Patients with Lymphoproliferative Disorders Undergoing Hematopoietic Stem Cell Mobilization

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. RNA Extraction and cDNA Synthesis
2.2. Absolute Gene Expression with Digital Quantitative PCR
2.3. Statistical Analysis
3. Results
3.1. Mobilization Data
3.2. Kinetics of Sirtuins
3.3. Sirtuin Level and Mobilization Efficacy
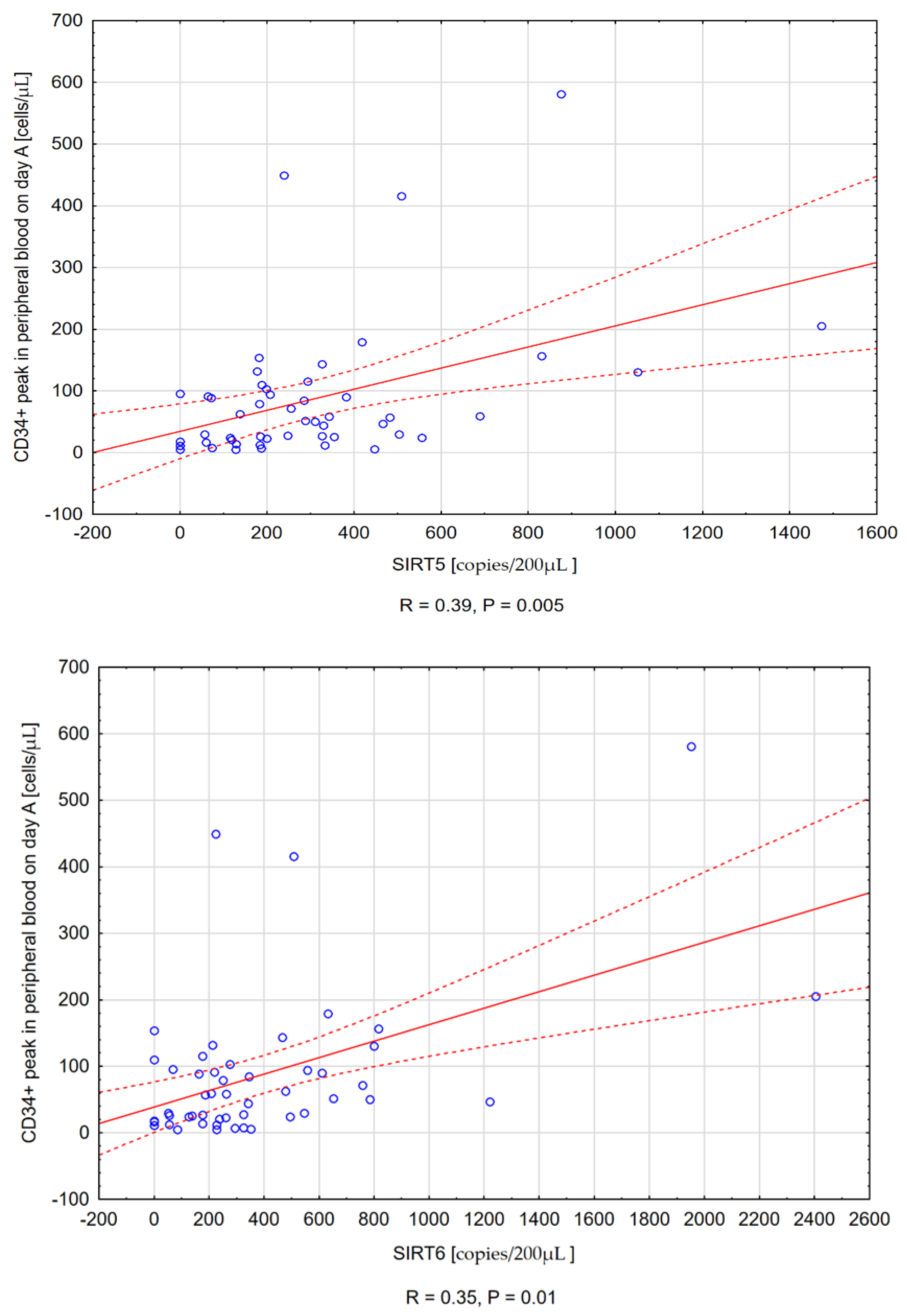
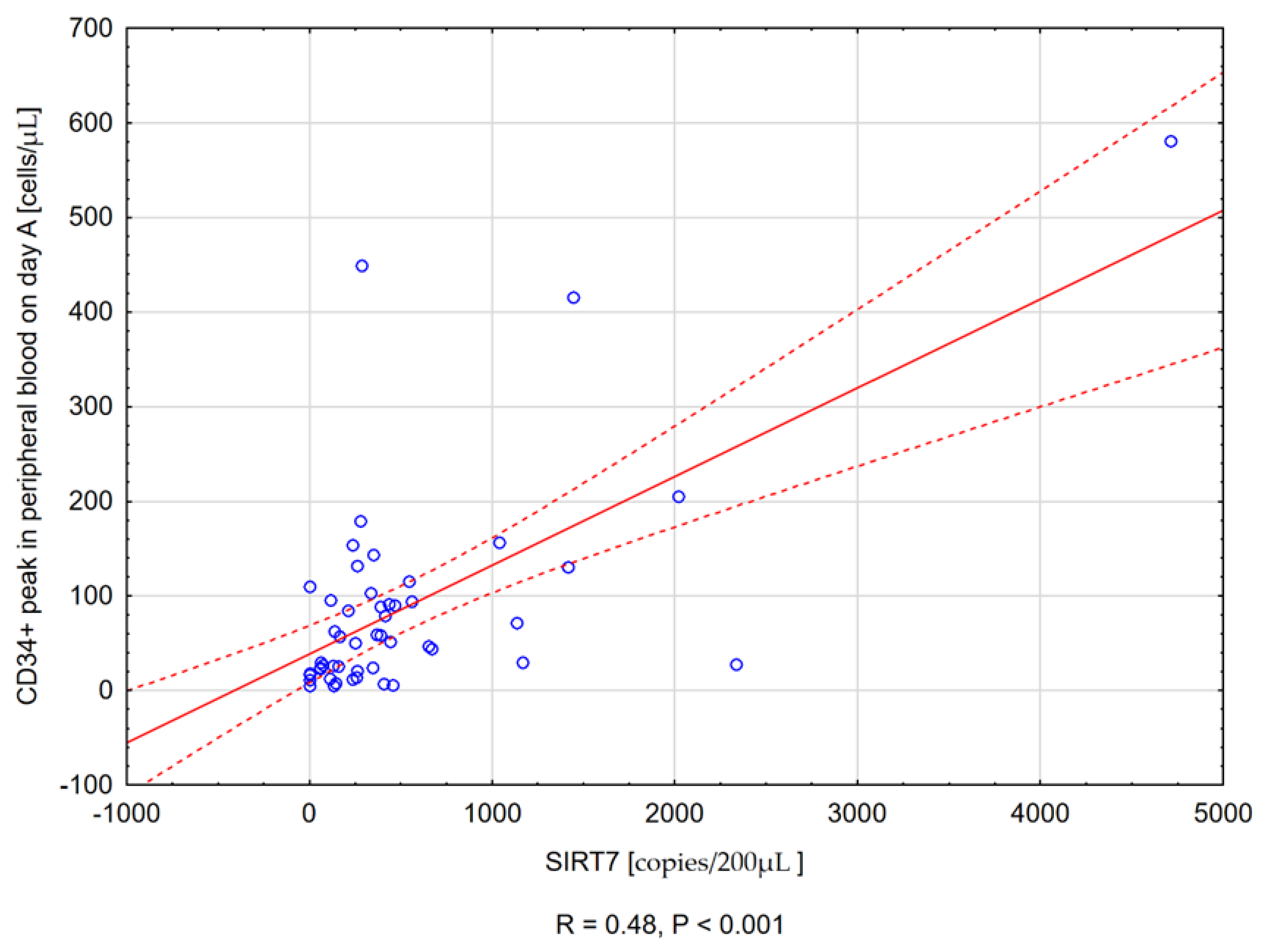
3.3.1. Sirtuin Expression and the CD34+ Peak Value in Peripheral Blood at First Apheresis
3.3.2. Sirtuin Concentration and the Number of Collected CD34+ Cells on the Day of First Apheresis
3.3.3. Sirtuin Expression and the Total Number of CD34+ Cells Collected during Mobilization
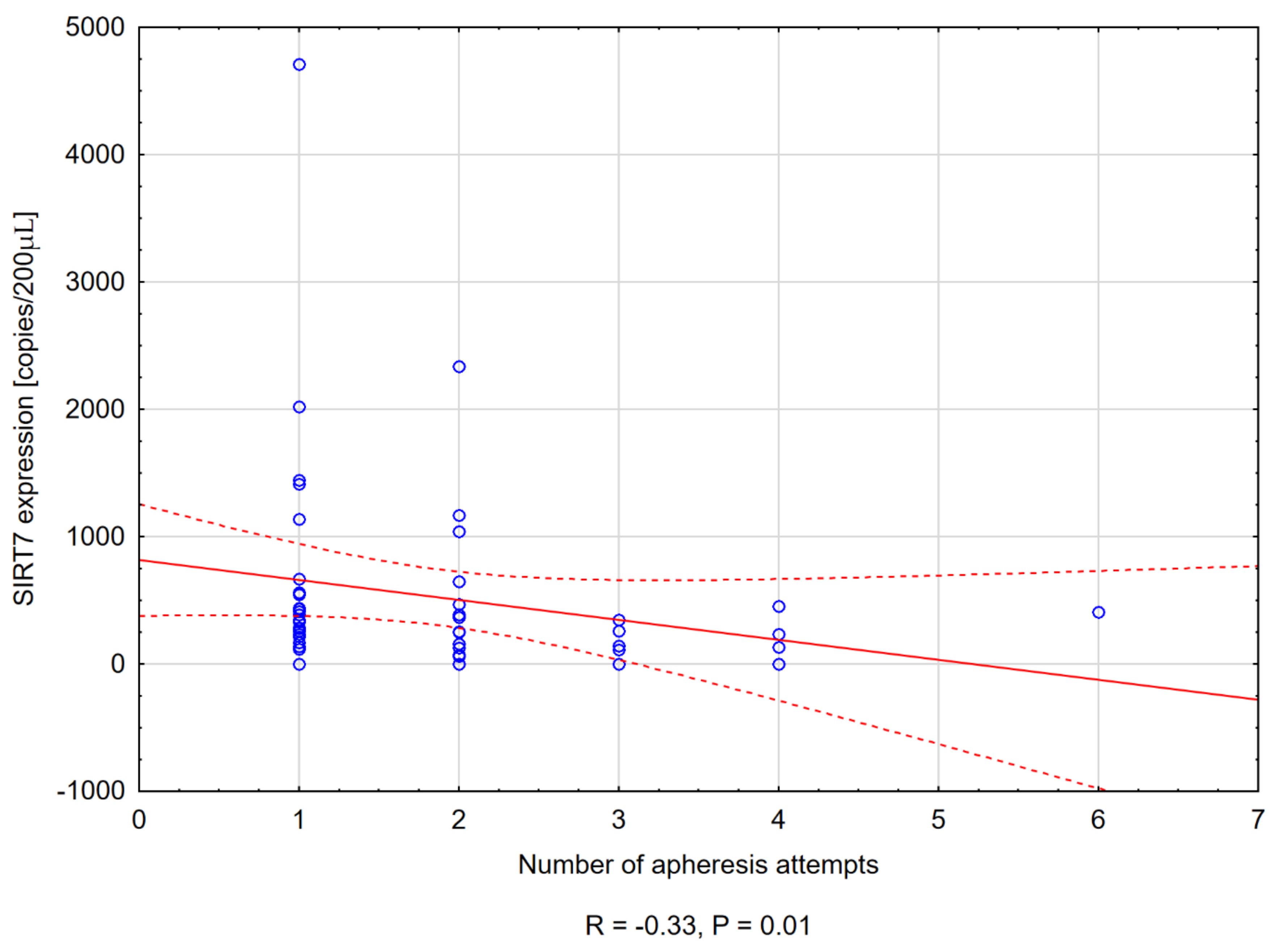
3.3.4. Sirtuin Level and the Number of Apheresis Attempts
3.4. Relationship between WBC and Sirtuins
3.5. Sirtuin Levels and Remission Status
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goldin, L.R.; McMaster, M.L.; Caporaso, N.E. Precursors to Lymphoproliferative Malignancies. Cancer Epidemiol. Biomark. Prev. 2013, 22, 533–539. [Google Scholar] [CrossRef] [Green Version]
- Ganapathi, K.A.; Pittaluga, S.; Odejide, O.O.; Freedman, A.S.; Jaffe, E.S. Early lymphoid lesions: Conceptual, diagnostic and clinical challenges. Haematologica 2014, 99, 1421–1432. [Google Scholar] [CrossRef] [Green Version]
- Ria, R.; Reale, A.; Melaccio, A.; Racanelli, V.; Dammacco, F.; Vacca, A. Filgrastim, lenograstim and pegfilgrastim in the mobilization of peripheral blood progenitor cells in patients with lymphoproliferative malignancies. Clin. Exp. Med. 2014, 15, 145–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopman, R.K.; DiPersio, J.F. Advances in stem cell mobilization. Blood Rev. 2014, 28, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Statkute, L.; Verda, L.; Oyama, Y.; Traynor, A.; Villa, M.; Shook, T.; Clifton, R.; Jovanovic, B.; Satkus, J.; Loh, Y.; et al. Mobilization, harvesting and selection of peripheral blood stem cells in patients with autoimmune diseases undergoing autologous hematopoietic stem cell transplantation. Bone Marrow Transplant. 2007, 39, 317–329. [Google Scholar] [CrossRef] [Green Version]
- Mendelson, A.; Frenette, P.S. Hematopoietic stem cell niche maintenance during homeostasis and regeneration. Nat. Med. 2014, 20, 833–846. [Google Scholar] [CrossRef] [Green Version]
- Nowicki, M.; Szemraj, J.; Wierzbowska, A.; Misiewicz, M.; Małachowski, R.; Pluta, A.; Grzybowska-Izydorczyk, O.; Robak, T.; Szmigielska-Kapłon, A. miRNA-15a, miRNA-16, miRNA-126, miRNA-146a, and miRNA-223 expressions in autologous hematopoietic stem cell transplantation and their impact on engraftment. Eur. J. Haematol. 2018, 100, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Nowicki, M.; Szemraj, J.; Wierzbowska, A.; Pluta, A.; Grzybowska-Izydorczyk, O.; Nowicka, A.; Stelmach, P.; Czemerska, M.; Szmigielska-Kapłon, A. Alterations in microRNA Expression during Hematopoietic Stem Cell Mobilization. Biology 2021, 10, 668. [Google Scholar] [CrossRef] [PubMed]
- Gattazzo, F.; Urciuolo, A.; Bonaldo, P. Extracellular matrix: A dynamic microenvironment for stem cell niche. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2014, 1840, 2506–2519. [Google Scholar] [CrossRef]
- Skokowa, J.; Lan, D.; Thakur, B.K.; Wang, F.; Gupta, K.; Cario, G.; Brechlin, A.M.; Schambach, A.; Hinrichsen, L.; Meyer, G.; et al. NAMPT is essential for the G-CSF–induced myeloid differentiation via a NAD+–sirtuin-1–dependent pathway. Nat. Med. 2009, 15, 151–158. [Google Scholar] [CrossRef]
- Roth, M.; Wang, Z.; Chen, W.Y. Sirtuins in hematological aging and malignancy. Crit. Rev. Oncog. 2013, 18, 531–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Callaghan, C.; Vassilopoulos, A. Sirtuins at the crossroads of stemness, aging, and cancer. Aging Cell 2017, 16, 1208–1218. [Google Scholar] [CrossRef]
- Carafa, V.; Rotili, D.; Forgione, M.; Cuomo, F.; Serretiello, E.; Hailu, G.S.; Jarho, E.; Lahtela-Kakkonen, M.; Mai, A.; Altucci, L. Sirtuin functions and modulation: From chemistry to the clinic. Clin. Epigenetics 2016, 8, 61. [Google Scholar] [CrossRef]
- Kozako, T.; Suzuki, T.; Yoshimitsu, M.; Arima, N.; Honda, S.-I.; Soeda, S. Anticancer Agents Targeted to Sirtuins. Molecules 2014, 19, 20295–20313. [Google Scholar] [CrossRef] [Green Version]
- Bosch-Presegué, L.; Vaquero, A. The Dual Role of Sirtuins in Cancer. Genes Cancer 2011, 2, 648–662. [Google Scholar] [CrossRef] [PubMed]
- Saunders, L.R.; Verdin, E. Sirtuins: Critical regulators at the crossroads between cancer and aging. Oncogene 2007, 26, 5489–5504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.Y.; Wang, D.H.; Yen, R.C.; Luo, J.; Gu, W.; Baylin, S.B. Tumor Suppressor HIC1 Directly Regulates SIRT1 to Modulate p53-Dependent DNA-Damage Responses. Cell 2005, 123, 437–448. [Google Scholar] [CrossRef]
- Jang, K.Y.; Hwang, S.H.; Kwon, K.S.; Kim, K.R.; Na Choi, H.; Lee, N.-R.; Kwak, J.-Y.; Park, B.-H.; Park, H.S.; Chung, M.J.; et al. SIRT1 Expression is Associated with Poor Prognosis of Diffuse Large B-Cell Lymphoma. Am. J. Surg. Pathol. 2008, 32, 1523–1531. [Google Scholar] [CrossRef]
- Lu, B.; Zhang, D.; Wang, X.; Lin, D.; Chen, Y.; Xu, X. Targeting SIRT1 to inhibit the proliferation of multiple myeloma cells. Oncol. Lett. 2021, 21, 306. [Google Scholar] [CrossRef] [PubMed]
- McGlynn, L.M.; Zino, S.; MacDonald, A.I.; Curle, J.; Reilly, J.E.; Mohammed, Z.M.; McMillan, D.C.; Mallon, E.; Payne, A.P.; Edwards, J.; et al. SIRT2: Tumour suppressor or tumour promoter in operable breast cancer? Eur. J. Cancer 2014, 50, 290–301. [Google Scholar] [CrossRef]
- Deng, A.; Ning, Q.; Zhou, L.; Liang, Y. SIRT2 is an unfavorable prognostic biomarker in patients with acute myeloid leukemia. Sci. Rep. 2016, 6, 27694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Villani, R.M.; Wang, H.; Simpson, M.J.; Roberts, M.S.; Tang, M.; Liang, X. The role of cellular reactive oxygen species in cancer chemotherapy. J. Exp. Clin. Cancer Res. 2018, 37, 266. [Google Scholar] [CrossRef]
- Wang, J.; Yi, J. Cancer cell killing via ROS: To increase or decrease, that is the question. Cancer Biol. Ther. 2008, 7, 1875–1884. [Google Scholar] [CrossRef]
- Ma, C.; Sun, Y.; Pi, C.; Wang, H.; Sun, H.; Yu, X.; Shi, Y.; He, X. Sirt3 Attenuates Oxidative Stress Damage and Rescues Cellular Senescence in Rat Bone Marrow Mesenchymal Stem Cells by Targeting Superoxide Dismutase 2. Front. Cell Dev. Biol. 2020, 8. [Google Scholar] [CrossRef]
- Brown, K.; Xie, S.; Qiu, X.; Mohrin, M.; Shin, J.; Liu, Y.; Zhang, D.; Scadden, D.T.; Chen, D. SIRT3 Reverses Aging-Associated Degeneration. Cell Rep. 2013, 3, 319–327. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Liu, B.; Yu, D.; Zuo, Y.; Cai, R.; Yang, J.; Cheng, J. SIRT3 deacetylase activity confers chemoresistance in AML via regulation of mitochondrial oxidative phosphorylation. Br. J. Haematol. 2019, 187, 49–64. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, K.; Araki, K.Y.; Naka, K.; Arai, F.; Takubo, K.; Yamazaki, S.; Matsuoka, S.; Miyamoto, T.; Ito, K.; Ohmura, M.; et al. Foxo3a Is Essential for Maintenance of the Hematopoietic Stem Cell Pool. Cell Stem Cell 2007, 1, 101–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsui, K.; Ezoe, S.; Oritani, K.; Shibata, M.; Tokunaga, M.; Fujita, N.; Tanimura, A.; Sudo, T.; Tanaka, H.; McBurney, M.W.; et al. NAD-dependent histone deacetylase, SIRT1, plays essential roles in the maintenance of hematopoietic stem cells. Biochem. Biophys. Res. Commun. 2012, 418, 811–817. [Google Scholar] [CrossRef]
- Edatt, L.; Poyyakkara, A.; Raji, G.R.; Ramachandran, V.; Shankar, S.; Kumar, V.B.S. Role of Sirtuins in Tumor Angiogenesis. Front. Oncol. 2020, 9, 1516. [Google Scholar] [CrossRef] [Green Version]
- Peled, T.; Shoham, H.; Aschengrau, D.; Yackoubov, D.; Frei, G.; Rosenheimer, G.N.; Lerrer, B.; Cohen, H.Y.; Nagler, A.; Fibach, E.; et al. Nicotinamide, a SIRT1 inhibitor, inhibits differentiation and facilitates expansion of hematopoietic progenitor cells with enhanced bone marrow homing and engraftment. Exp. Hematol. 2012, 40, 342–355.e1. [Google Scholar] [CrossRef]
- Potente, M.; Ghaeni, L.; Baldessari, D.; Mostoslavsky, R.; Rossig, L.; Dequiedt, F.; Haendeler, J.; Mione, M.; Dejana, E.; Alt, F.W.; et al. SIRT1 controls endothelial angiogenic functions during vascular growth. Genes Dev. 2007, 21, 2644–2658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Dang, W. SIRT7 slows down stem cell aging by preserving heterochromatin: A perspective on the new discovery. Protein Cell 2020, 11, 469–471. [Google Scholar] [CrossRef] [PubMed]
- Peck, B.; Chen, C.-Y.; Ho, K.-K.; Di Fruscia, P.; Myatt, S.S.; Coombes, R.C.; Fuchter, M.; Hsiao, C.-D.; Lam, E. SIRT Inhibitors Induce Cell Death and p53 Acetylation through Targeting Both SIRT1 and SIRT2. Mol. Cancer Ther. 2010, 9, 844–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, H.S.; McBurney, M.; Robbins, P.D. SIRT1 Negatively Regulates the Mammalian Target of Rapamycin. PLoS ONE 2010, 5, e9199. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Su, P.; Yin, C.; Lin, X.; Wang, X.; Gao, Y.; Patil, S.; War, A.R.; Qadir, A.; Tian, Y.; et al. The Roles of FoxO Transcription Factors in Regulation of Bone Cells Function. Int. J. Mol. Sci. 2020, 21, 692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imperatore, F.; Maurizio, J.; Aguilar, S.V.; Busch, C.J.; Favret, J.; Kowenz-Leutz, E.; Cathou, W.; Gentek, R.; Perrin, P.; Leutz, A.; et al. SIRT1 regulates macrophage self-renewal. EMBO J. 2017, 36, 2353–2372. [Google Scholar] [CrossRef]
- Sun, X.; Cao, B.; Naval-Sanchez, M.; Pham, T.; Sun, Y.B.Y.; Williams, B.; Heazlewood, S.Y.; Deshpande, N.; Li, J.; Kraus, F.; et al. Nicotinamide riboside attenuates age-associated metabolic and functional changes in hematopoietic stem cells. Nat. Commun. 2021, 12, 2665. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Diao, D.; Shi, Z.; Zhu, X.; Gao, Y.; Gao, S.; Liu, X.; Wu, Y.; Rudolph, K.L.; Liu, G.-H.; et al. SIRT6 Controls Hematopoietic Stem Cell Homeostasis through Epigenetic Regulation of Wnt Signaling. Cell Stem Cell 2016, 18, 495–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinonen, T.; Ciarlo, E.; Le Roy, D.; Roger, T. Impact of the Dual Deletion of the Mitochondrial Sirtuins SIRT3 and SIRT5 on Anti-microbial Host Defenses. Front. Immunol. 2019, 10, 2341. [Google Scholar] [CrossRef]
- Yang, J.; Li, Y.; Zhang, Y.; Fang, X.; Chen, N.; Zhou, X.; Wang, X. Sirt6 promotes tumorigenesis and drug resistance of diffuse large B-cell lymphoma by mediating PI3K/Akt signaling. J. Exp. Clin. Cancer Res. 2020, 39, 142. [Google Scholar] [CrossRef] [PubMed]
- Koch, C.; Samareh, B.; Morishima, T.; Mir, P.; Kanz, L.; Zeidler, C.; Skokowa, J.; Welte, K. GM-CSF treatment is not effective in congenital neutropenia patients due to its inability to activate NAMPT signaling. Ann. Hematol. 2016, 96, 345–353. [Google Scholar] [CrossRef]
- Hu, X.; Mei, S.; Meng, W.; Xue, S.; Jiang, L.; Yang, Y.; Hui, L.; Chen, Y.; Guan, M.-X. CXCR4-mediated signaling regulates autophagy and influences acute myeloid leukemia cell survival and drug resistance. Cancer Lett. 2018, 425, 1–12. [Google Scholar] [CrossRef]
- Vaziri, H.; Dessain, S.K.; Eaton, E.N.; Imai, S.-I.; Frye, R.A.; Pandita, T.K.; Guarente, L.; Weinberg, R.A. hSIR2SIRT1 Functions as an NAD-Dependent p53 Deacetylase. Cell 2001, 107, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Guarani, V.; Deflorian, G.; Franco, C.; Krüger, M.; Phng, L.-K.; Bentley, K.; Toussaint, L.; Dequiedt, F.; Mostoslavsky, R.; Schmidt, M.H.H.; et al. Acetylation-dependent regulation of endothelial Notch signalling by the SIRT1 deacetylase. Nature 2011, 473, 234–238. [Google Scholar] [CrossRef]
- Kunhiraman, H.; Edatt, L.; Thekkeveedu, S.; Poyyakkara, A.; Raveendran, V.; Kiran, M.S.; Sudhakaran, P.; Kumar, S.V. 2-Deoxy Glucose Modulates Expression and Biological Activity of VEGF in a SIRT-1 Dependent Mechanism. J. Cell. Biochem. 2016, 118, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Edatt, L.; Haritha, K.; Sruthi, T.V.; Aswini, P.; Kumar, V.B.S. 2-Deoxy glucose regulate MMP-9 in a SIRT-1 dependent and NFkB independent mechanism. Mol. Cell. Biochem. 2016, 423, 197–206. [Google Scholar] [CrossRef]
- Hashimoto-Komatsu, A.; Hirase, T.; Asaka, M.; Node, K. Angiotensin II induces microtubule reorganization mediated by a deacetylase SIRT2 in endothelial cells. Hypertens. Res. 2011, 34, 949–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, A.H.-H.; Wu, L.-H.; Shieh, S.-S.; Wang, D.L. SIRT3 interactions with FOXO3 acetylation, phosphorylation and ubiquitinylation mediate endothelial cell responses to hypoxia. Biochem. J. 2014, 464, 157–168. [Google Scholar] [CrossRef]
- Tang, X.; Shi, L.; Xie, N.; Liu, Z.; Qian, M.; Meng, F.; Xu, Q.; Zhou, M.; Cao, X.; Zhu, W.-G.; et al. SIRT7 antagonizes TGF-β signaling and inhibits breast cancer metastasis. Nat. Commun. 2017, 8, 318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malik, S.; Villanova, L.; Tanaka, S.; Aonuma, M.; Roy, N.; Berber, E.; Pollack, J.R.; Michishita-Kioi, E.; Chua, K.F. SIRT7 inactivation reverses metastatic phenotypes in epithelial and mesenchymal tumors. Sci. Rep. 2015, 5, 9841. [Google Scholar] [CrossRef] [Green Version]
- Olivieri, A.; on behalf of the Italian Group for Stem Cell Transplantation (GITMO); Marchetti, M.; Lemoli, R.; Tarella, C.; Iacone, A.; Lanza, F.; Rambaldi, A.; Bosi, A. Proposed definition of ‘poor mobilizer’ in lymphoma and multiple myeloma: An analytic hierarchy process by ad hoc working group Gruppo ItalianoTrapianto di Midollo Osseo. Bone Marrow Transpl. 2011, 47, 342–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Lei, W.; Chen, X.; Wang, S.; Qian, W. Oxidative stress response induced by chemotherapy in leukemia treatment (Review). Mol. Clin. Oncol. 2018, 8, 391–399. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Shi, L.; Liang, F.; Xu, W.; Li, T.; Gao, L.; Sun, Z.; Yu, J.; Zhang, J. Sirt3 Ameliorates Oxidative Stress and Mitochondrial Dysfunction After Intracerebral Hemorrhage in Diabetic Rats. Front. Neurosci. 2018, 12, 414. [Google Scholar] [CrossRef] [PubMed]
- Wilk, A.; Hayat, F.; Cunningham, R.; Li, J.; Garavaglia, S.; Zamani, L.; Ferraris, D.M.; Sykora, P.; Andrews, J.; Clark, J.; et al. Extracellular NAD+ enhances PARP-dependent DNA repair capacity independently of CD73 activity. Sci. Rep. 2020, 10, 651. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.; Mu, W.-C.; Karki, R.; Chiang, H.-H.; Mohrin, M.; Shin, J.J.; Ohkubo, R.; Ito, K.; Kanneganti, T.-D.; Chen, D. Mitochondrial Stress-Initiated Aberrant Activation of the NLRP3 Inflammasome Regulates the Functional Deterioration of Hematopoietic Stem Cell Aging. Cell Rep. 2019, 26, 945–954.e4. [Google Scholar] [CrossRef] [Green Version]
- Baumann, M.; Frye, T.; Naqvi, T.; Gomez-Cambronero, J. Normal neutrophil maturation is associated with selective loss of MAP kinase activation by G-CSF. Leuk. Res. 2005, 29, 73–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardus, A.; Uryga, A.K.; Walters, G.; Erusalimsky, J.D. SIRT6 protects human endothelial cells from DNA damage, telomere dysfunction, and senescence. Cardiovasc. Res. 2012, 97, 571–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohrin, M.; Shin, J.; Liu, Y.; Brown, K.; Luo, H.; Xi, Y.; Haynes, C.M.; Chen, D. A mitochondrial UPR-mediated metabolic checkpoint regulates hematopoietic stem cell aging. Science 2015, 347, 1374–1377. [Google Scholar] [CrossRef] [Green Version]
- Vakhrusheva, O.; Smolka, C.; Gajawada, P.; Kostin, S.; Boettger, T.; Kubin, T.; Braun, T.; Bober, E. Sirt7 Increases Stress Resistance of Cardiomyocytes and Prevents Apoptosis and Inflammatory Cardiomyopathy in Mice. Circ. Res. 2008, 102, 703–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betsinger, C.N.; Cristea, I.M. Mitochondrial Function, Metabolic Regulation, and Human Disease Viewed through the Prism of Sirtuin 4 (SIRT4) Functions. J. Proteome Res. 2019, 18, 1929–1938. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Huang, C.; Huang, Y.; Hong, L.; Wang, H.; Zhou, Z.; Qiu, Y. SIRT4 Suppresses Inflammatory Responses in Human Umbilical Vein Endothelial Cells. Cardiovasc. Toxicol. 2014, 15, 217–223. [Google Scholar] [CrossRef]
- Carafa, V.; Altucci, L.; Nebbioso, A. Dual Tumor Suppressor and Tumor Promoter Action of Sirtuins in Determining Malignant Phenotype. Front. Pharmacol. 2019, 10, 38. [Google Scholar] [CrossRef] [Green Version]
- Jeong, S.M.; Lee, A.; Lee, J.; Haigis, M.C. SIRT4 Protein Suppresses Tumor Formation in Genetic Models of Myc-induced B Cell Lymphoma. J. Biol. Chem. 2014, 289, 4135–4144. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.-S.; Xia, L.; Mills, G.B.; Lowell, C.A.; Touw, I.P.; Corey, S.J. G-CSF induced reactive oxygen species involves Lyn-PI3-kinase-Akt and contributes to myeloid cell growth. Blood 2006, 107, 1847–1856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajkumar, S.V.; Fonseca, R.; Witzig, T.; Gertz, M.; Greipp, P. Bone marrow angiogenesis in patients achieving complete response after stem cell transplantation for multiple myeloma. Leukemia 1999, 13, 469–472. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Shi, X.; Ren, M.; Liu, L.; Qi, H.; Zhang, C.; Zou, J.; Qiu, X.; Zhu, W.-G.; Zhang, Y.E.; et al. SIRT7 Deacetylates STRAP to Regulate p53 Activity and Stability. Int. J. Mol. Sci. 2020, 21, 4122. [Google Scholar] [CrossRef] [PubMed]
- Ianni, A.; Kumari, P.; Tarighi, S.; Simonet, N.G.; Popescu, D.; Guenther, S.; Hölper, S.; Schmidt, A.; Smolka, C.; Yue, S.; et al. SIRT7-dependent deacetylation of NPM promotes p53 stabilization following UV-induced genotoxic stress. Proc. Natl. Acad. Sci. USA 2021, 118, e2015339118. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Characteristics | Numbers |
|---|---|
| Age (years) | Median 60 (range 44–69) |
| Sex (female/male) | 24/26 |
| Multiple myeloma | 39 (7 CR, 24 VGPR, 8 PR) |
| Hodgkin lymphoma | 4 (1 CR, 3 PR) |
| non-Hodgkin lymphoma: Diffuse large B-cell lymphoma Mantle cell lymphoma Anaplastic large-cell lymphoma Hepatosplenic T-cell lymphoma | 7 3 (1 CR, 2 PR) 2 (CR) 1 (PR) 1 (PR) |
| Mobilization chemotherapy: Multiple myeloma Hodgkin and non-Hodgkin lymphoma | Cyclophosphamide (2000 mg/m2) (n = 26) DCEP (Dexamethasone 40 mg, Cyclophosphamide 400 mg/m2, Cisplatin 10 mg/m2, Etoposide 40 mg/m2) (n = 6) Cytarabine (1600 mg/m2) (n = 2) G-CSF (10 µg/kg) in monotherapy (n = 5) ICE (Ifosfamide 3000 mg/m2, Carboplatin AUC = 5, max 800 mg/m2, Etoposide 100 mg/m2) (n = 5)R-ICE (with rituximab 375 mg/m2) (n = 1) DHAP (Dexamethasone 40 mg, Cytarabine 2000 mg/m2, Cisplatin 100 mg/m2) (n = 1) R-DHAP (with rituximab 375 mg/m2) (n = 2) Cyclophosphamide (2000 mg/m2) (n = 1) Cytarabine (1600 mg/m2 (n = 1) |
| Characteristics | Numbers |
|---|---|
| CD34+ cells collected during mobilization (total number) (×106/kg) | Median 5.23 (range 2.2–35.6) |
| CD34+ collected on Day A (×106/kg) | Median 3.35 (range 0.3–35.6) |
| Number of apheresis needed to collect at least 2 × 106/kg CD34+ | Median 2 (range 1–6) |
| CD34+ peak in peripheral blood before first apheresis (cells/µL) | Median 54.4 (range 4.8–581) |
| WBC count on Day A (×103/µL) | Median 16.91 (range 2.68–47.42) |
| Mobilization efficacy | |
| Good mobilizers | 45 |
| * Poor mobilizers | 5 |
| Sirtuin | Day 0 (Me, Range) | Day A (Me, Range) | p Value |
|---|---|---|---|
| SIRT1 | 68.55 range: 0–394.57 | 368.22 0–16748.56 | p < 0.001 |
| SIRT2 | 80.12 0–425.65 | 730.74 65.6–4787.37 | p < 0.001 |
| SIRT3 | 28.67 0–256.72 | 161.28 0–14167.35 | p < 0.001 |
| SIRT4 | 0 0–71.86 | 0 0–134.96 | p = 0.05 |
| SIRT5 | 0 0–255.69 | 243.6 0–1473.39 | p < 0.001 |
| SIRT6 | 0 0–240.22 | 256.16 0–2404.06 | p < 0.001 |
| SIRT7 | 0 0–206.93 | 282.96 0–4711.99 | p < 0.001 |
| Factor | Coeff | Std Err | t | p > ǀtǀ | [0.025 | 0.975] |
|---|---|---|---|---|---|---|
| Intercept | 1.8104 | 79.591 | 0.023 | 0.982 | −158.594 | 162.215 |
| Age | 1.2544 | 1.380 | 0.909 | 0.368 | −1.527 | 4.036 |
| SIRT5 (Day A) | 0.0452 | 0.074 | 0.608 | 0.546 | −0.105 | 0.195 |
| SIRT6 (Day A) | 0.0355 | 0.058 | 0.615 | 0.542 | −0.081 | 0.152 |
| SIRT 7 (Day A) | 0.0911 | 0.025 | 3.611 | 0.001 | 0.040 | 0.142 |
| WBC count (Day A) | −3.0984 | 1.395 | −2.221 | 0.032 | −5.909 | −0.287 |
| Factor | Coeff | Std Err | t | p > ǀtǀ | [0.025 | 0.975] |
|---|---|---|---|---|---|---|
| Intercept | 38.3839 | 14.983 | 2.562 | 0.014 | 8.258 | 68.510 |
| SIRT7 (Day A) | 0.0939 | 0.016 | 5.861 | 0.000 | −0.105 | 0.195 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nowicki, M.; Wierzbowska, A.; Stec-Martyna, E.; Kulczycka-Wojdala, D.; Nowicki, G.; Szmigielska-Kapłon, A. SIRT1-SIRT7 Expression in Patients with Lymphoproliferative Disorders Undergoing Hematopoietic Stem Cell Mobilization. Cancers 2022, 14, 1213. https://doi.org/10.3390/cancers14051213
Nowicki M, Wierzbowska A, Stec-Martyna E, Kulczycka-Wojdala D, Nowicki G, Szmigielska-Kapłon A. SIRT1-SIRT7 Expression in Patients with Lymphoproliferative Disorders Undergoing Hematopoietic Stem Cell Mobilization. Cancers. 2022; 14(5):1213. https://doi.org/10.3390/cancers14051213
Chicago/Turabian StyleNowicki, Mateusz, Agnieszka Wierzbowska, Emilia Stec-Martyna, Dominika Kulczycka-Wojdala, Grzegorz Nowicki, and Anna Szmigielska-Kapłon. 2022. "SIRT1-SIRT7 Expression in Patients with Lymphoproliferative Disorders Undergoing Hematopoietic Stem Cell Mobilization" Cancers 14, no. 5: 1213. https://doi.org/10.3390/cancers14051213