
Polymorphism of VDR Gene and the Sensitivity of Human Leukemia and Lymphoma Cells to Active Forms of Vitamin D

 , , , , , , , and
, , , , , , , and
Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Human Cell Lines
2.2. Normal Human Blood
2.3. Compounds
2.4. Anti-Proliferative Assay In Vitro
2.5. Cell Morphology–Cytospin
2.6. Cell Surface Markers
2.7. Vitamin D Receptor and 1,25-D3-MARRS Protein Level Analysis Using Western Blot Method
2.8. Detection of Vitamin D Receptor (VDR) Polymorphisms
- FokI (rs2228570)
- 5′ AGCTGGCCCTGGCACTGACTCTGCTCT 3′
- 5′ ATGGAAACACCTTGCTTCTTCTCCCTC 3′
- BsmI (rs1544410)
- 5′CAACCAAGACTACAAGTACCGCGTCAGTGA′
- 5′ AACCAGCGGGAAGAGGTCAAGGG 3′
- ApaI (rs7975232) and TaqI (rs731236)
- 5′-CAG AGC ATG GAC AGG GAG CAA G-3′
- 5′-GCA ACT CCT CAT GGC TGA GGT CTC A-3′
2.9. Statistical Analysis
3. Results
3.1. Leukemia and Lymphoma Cells Respond to Calcitriol and Tacalcitol
3.2. Cell Morphology
3.3. Analysis of Surface Markers
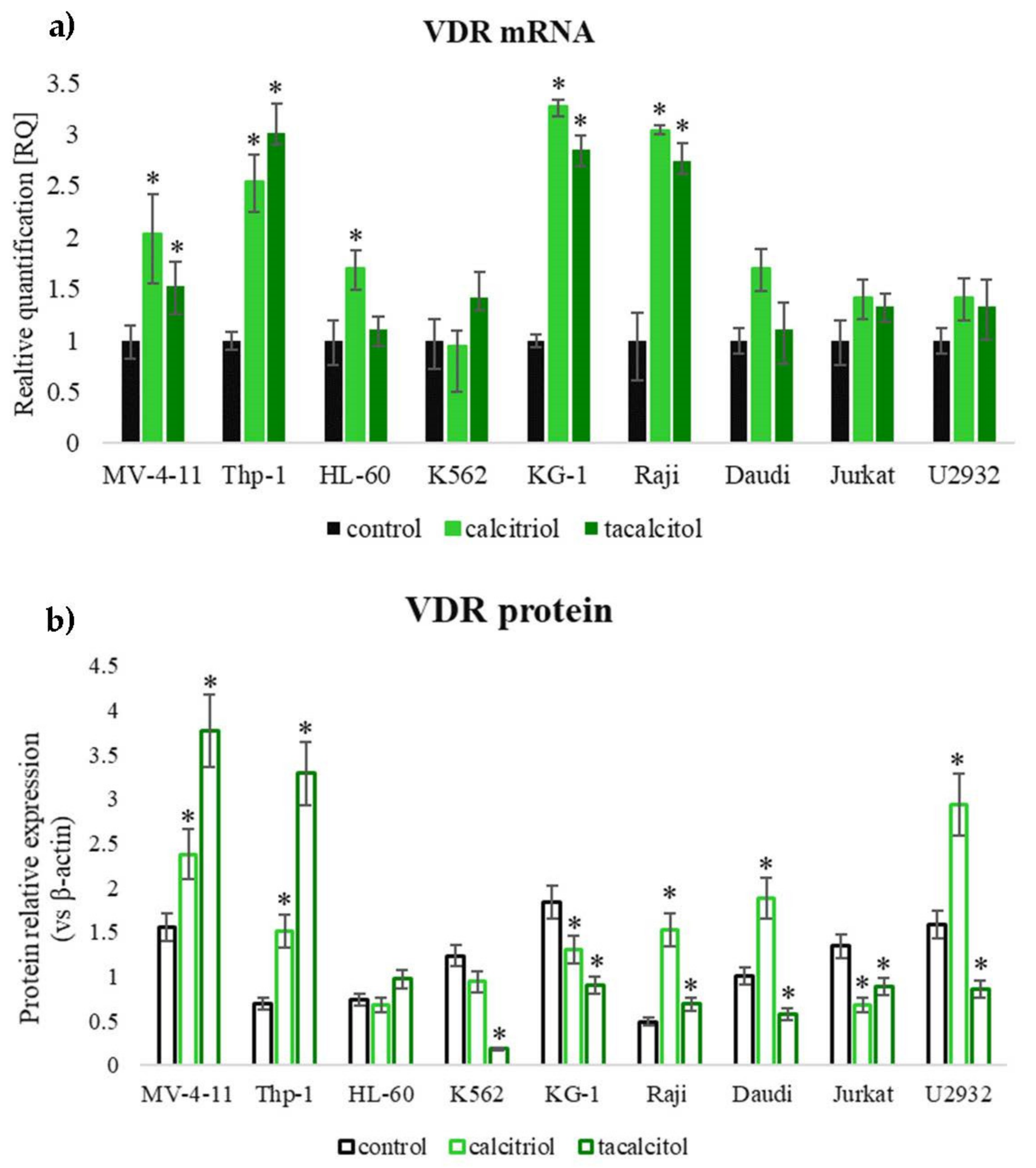
3.4. Assessment of Calcitriol and Tacalcitol Binding Receptors in Human Leukemia and Lymphoma Cells
3.5. Polymorphism of Vitamin D Receptor Gene in Human Leukemia and Lymphoma Cells
3.5.1. FokI Polymorphism
3.5.2. BsmI-ApaI-TaqI Polymorphism
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Campo, E.; Swerdlow, S.H.; Harris, N.L.; Pileri, S.; Stein, H.; Jaffe, E.S. The 2008 WHO classification of lymphoid neoplasms and beyond: Evolving concepts and practical applications. Blood 2011, 117, 5019–5032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, N.N.; Fry, T.J. Mechanisms of resistance to CAR T cell therapy. Nat. Rev. Clin. Oncol. 2019, 16, 372–385. [Google Scholar] [CrossRef] [PubMed]
- Hamadani, M.; Awan, F.T. Remission induction, consolidation and novel agents in development for adults with acute myeloid leukaemia. Hematol. Oncol. 2010, 28, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Trémezaygues, L.; Reichrath, J. Vitamin D analogs in the treatment of psoriasis: Where are we standing and where will we be going? Dermatoendocrinology 2011, 3, 180–186. [Google Scholar] [CrossRef]
- Rodríguez-Martín, M.; García Bustínduy, M.; Sáez Rodríguez, M.; Noda Cabrera, A. Randomized, double-blind clinical trial to evaluate the efficacy of topical tacalcitol and sunlight exposure in the treatment of adult nonsegmental vitiligo. Br. J. Dermatol. 2009, 160, 409–414. [Google Scholar] [CrossRef]
- Wietrzyk, J.; Pełczyńska, M.; Madej, J.; Dzimira, S.; Kuśnierczyk, H.; Kutner, A.; Szelejewski, W.; Opolski, A. Toxicity and antineoplastic effect of (24R)-1,24-dihydroxyvitamin D3 (PRI-2191). Steroids 2004, 69, 629–635. [Google Scholar] [CrossRef]
- Study of High-Dose Pulse Administration DN-101 (Calcitriol) in Patients with Myelodysplastic Syndrome (MDS)-Full Text View-ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT00057031?term=calcitriol&cond=MDS&draw=2&rank=1 (accessed on 30 June 2020).
- Rituximab, Romidepsin, and Lenalidomide in Treating Patients with Recurrent or Refractory B-Cell Non-Hodgkin Lym-phoma. Available online: https://clinicaltrials.gov/ct2/show/NCT02281279 (accessed on 30 June 2020).
- Gocek, E.; Kiełbiński, M.; Baurska, H.; Haus, O.; Kutner, A.; Marcinkowska, E. Different susceptibilities to 1,25-dihydroxyvitamin D3-induced differentiation of AML cells carrying various mutations. Leuk. Res. 2010, 34, 649–657. [Google Scholar] [CrossRef]
- Pike, J.W.; Meyer, M.B.; Brown, G.; Kallay, E. New approaches to assess mechanisms of action of selective vitamin D analogues. Int. J. Mol. Sci. 2021, 22, 12352. [Google Scholar] [CrossRef]
- Christakos, S.; Raval-Pandya, M.; Wernyj, R.P.; Yang, W. Genomic mechanisms involved in the pleiotropic actions of 1,25-dihydroxyvitamin D3. Biochem. J. 1996, 316, 361–371. [Google Scholar] [CrossRef]
- Guo, G.G.; Patel, K.; Kumar, V.; Shah, M.; Fried, V.A.; Etlinger, J.D.; Sehgal, P.B. Association of the chaperone glucose-regulated protein 58 (GRP58/ER-60/ERp57) with Stat3 in cytosol and plasma membrane complexes. J. Interf. Cytokine Res. 2002, 22, 555–563. [Google Scholar] [CrossRef]
- Farach-Carson, M.; Nemere, I. Membrane receptors for vitamin d steroid hormones: Potential newdrug targets. Curr. Drug Targets 2005, 4, 67–76. [Google Scholar] [CrossRef]
- An update on vitamin D signaling and cancer. Semin. Cancer Biol. 2020, 30, S1044-579. [CrossRef]
- Uitterlinden, A.G.; Fang, Y.; Van Meurs, J.B.J.; Pols, H.A.P.; Van Leeuwen, J.P.T.M. Genetics and biology of vitamin D receptor polymorphisms. Gene 2004, 338, 143–156. [Google Scholar] [CrossRef] [Green Version]
- Kerr Whitfield, G.; Remus, L.S.; Jurutka, P.W.; Zitzer, H.; Oza, A.K.; Dang, H.T.L.; Haussler, C.A.; Galligan, M.A.; Thatcher, M.L.; Dominguez, C.E.; et al. Functionally relevant polymorphisms in the human nuclear vitamin D receptor gene. Mol. Cell. Endocrinol. 2001, 177, 145–159. [Google Scholar] [CrossRef]
- Rai, V.; Abdo, J.; Agrawal, S.; Agrawal, D.K. Vitamin D receptor polymorphism and cancer: An update. Anticancer Res. 2017, 37, 3991–4003. [Google Scholar]
- Johnson, D.E.; Redner, R.L. An ATRActive future for differentiation therapy in AML. Blood Rev. 2015, 29, 263–268. [Google Scholar] [CrossRef] [Green Version]
- Vardiman, J.W.; Harris, N.L.; Brunning, R.D. The World Health Organization (WHO) classification of the myeloid neoplasms. Blood 2002, 100, 2292–2302. [Google Scholar] [CrossRef]
- Goldman, S.L.; Hassan, C.; Khunte, M.; Soldatenko, A.; Jong, Y.; Afshinnekoo, E.; Mason, C.E. Epigenetic modifications in acute myeloid leukemia: Prognosis, treatment, and heterogeneity. Front. Genet. 2019, 10, 133. [Google Scholar] [CrossRef] [Green Version]
- Bochtler, T.; Fröhling, S.; Krämer, A. Role of chromosomal aberrations in clonal diversity and progression of acute myeloid leukemia. Leukemia 2015, 29, 1243–1252. [Google Scholar] [CrossRef]
- Luong, Q.T.; Koeffler, H.P. Vitamin D compounds in leukemia. J. Steroid Biochem. Mol. Biol. 2005, 97, 195–202. [Google Scholar] [CrossRef]
- Endres, D.B. Investigation of hypercalcemia. Clin. Biochem. 2012, 45, 954–963. [Google Scholar] [CrossRef]
- Choi, M.; Makishima, M. Therapeutic applications for novel non-hypercalcemic vitamin D receptor ligands. Expert Opin. Ther. Pat. 2009, 19, 593–606. [Google Scholar] [CrossRef]
- Purdue, M.P.; Hartge, P.; Davis, S.; Cerhan, J.R.; Colt, J.S.; Cozen, W.; Severson, R.K.; Li, Y.; Chanock, S.J.; Rothman, N.; et al. Sun exposure, vitamin D receptor gene polymorphisms and risk of non-Hodgkin lymphoma. Cancer Causes Control 2007, 18, 989–999. [Google Scholar] [CrossRef]
- Baurska, H.; Kiełbiński, M.; Biecek, P.; Haus, O.; Jaźwiec, B.; Kutner, A.; Marcinkowska, E. Monocytic differentiation induced by side-chain modified analogs of vitamin D in ex vivo cells from patients with acute myeloid leukemia. Leuk. Res. 2014, 38, 638–647. [Google Scholar] [CrossRef]
- Kim, M.; Mirandola, L.; Pandey, A.; Nguyen, D.D.; Jenkins, M.R.; Turcel, M.; Cobos, E.; Chiriva-Internati, M. Application of vitamin D and derivatives in hematological malignancies. Cancer Lett. 2012, 319, 8–22. [Google Scholar] [CrossRef]
- Ladines-Castro, W.; Barragán-Ibañez, G.; Luna-Pérez, M.A.; Santoyo-Sánchez, A.; Collazo-Jaloma, J.; Mendoza-García, E.; Ramos-Peñafiel, C.O. Morphology of leukaemias. Rev. Médica Del Hosp. Gen. México 2016, 79, 107–113. [Google Scholar] [CrossRef]
- Klionsky, D.J.; Emr, S.D. Autophagy as a regulated pathway of cellular degradation. Science 2000, 290, 1717–1721. [Google Scholar] [CrossRef]
- Gewirtz, D.A. Autophagy as a mechanism of radiation sensitization in breast tumor cells. Autophagy 2007, 3, 249–250. [Google Scholar] [CrossRef] [Green Version]
- Pike, J.W.; Meyer, M.B. The vitamin D receptor: New paradigms for the regulation of gene expression by 1,25-dihydroxyvitamin D3. Endocrinol. Metab. Clin. North Am. 2010, 39, 255–269. [Google Scholar] [CrossRef] [Green Version]
- Diaz, D.; Allamnei, K.; Tarrant, J.M.; Lewin-Koh, S.-C.; Pai, R.; Dhawan, P.; Cain, G.R.; Kozlowski, C.; Hiraragi, H.; La, N.; et al. Phosphorous dysregulation induced by MEK small molecule inhibitors in the rat involves blockade of FGF-23 signaling in the kidney. Toxicol. Sci. 2012, 125, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Consiglio, M.; Destefanis, M.; Morena, D.; Foglizzo, V.; Forneris, M.; Pescarmona, G.; Silvagno, F. The vitamin D receptor inhibits the respiratory chain, contributing to the metabolic switch that is essential for cancer cell proliferation. PLoS ONE 2014, 9, e115816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorne, J.; Campbell, M.J. The vitamin D receptor in cancer. Nutr. Soc. 2008, 67, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Vitamin D Receptor Controls Cell Stemness in Acute Myeloid Leukemia and in Normal Bone Marrow | Elsevier Enhanced Reader. Available online: https://reader.elsevier.com/reader/sd/pii/S2211124719317127?token=44B1A4237F242647C695F2F440C635CAC4E6EECD6E0949ACA9C270187557A9E054E198C8D4DA56B60A88413713B0139A&originRegion=us-east-1&originCreation=20211214015034 (accessed on 13 December 2021).
- Nemere, I.; Ray, R.; McManus, W. Immunochemical studies on the putative plasmalemmal receptor for 1,25(OH)2D3. I. Chick intestine. Am. J. Physiol.-Endocrinol. Metab. 2000, 278, E1104–E1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; He, B.; Pan, Y.; Deng, Q.; Sun, H.; Li, R.; Gao, T.; Song, G.; Wang, S. Systematic review and meta-analysis on vitamin D receptor polymorphisms and cancer risk. Tumor Biol. 2014, 35, 4153–4169. [Google Scholar] [CrossRef]
- Simmons, J.D.; Mullighan, C.; Welsh, K.I.; Jewell, D.P. Vitamin D receptor gene polymorphism: Association with Crohn’s disease susceptibility. Gut 2000, 47, 211–214. [Google Scholar] [CrossRef] [Green Version]
- Dresner-Pollak, R.; Ackerman, Z.; Eliakim, R.; Karban, A.; Chowers, Y.; Fidder, H.H. The Bsm I vitamin d receptor gene polymorphism is associated with ulcerative colitis in Jewish Ashkenazi patients. Genet. Test. 2004, 8, 417–420. [Google Scholar] [CrossRef]
- Haussler, M.R.; Whitfield, G.K.; Haussler, C.A.; Hsieh, J.C.; Thompson, P.D.; Selznick, S.H.; Dominguez, C.E.; Jurutka, P.W. The nuclear vitamin D receptor: Biological and molecular regulatory properties revealed. J. Bone Miner. Res. 1998, 13, 325–349. [Google Scholar] [CrossRef]
- Jurutka, P.W.; Remus, L.S.; Whitfield, G.K.; Thompson, P.D.; Hsieh, J.C.; Zitzer, H.; Tavakkoli, P.; Galligan, M.A.; Dang, H.T.L.; Haussler, C.A.; et al. The polymorphic N terminus in human vitamin D receptor isoforms influences transcriptional activity by modulating interaction with transcription factor IIB. Mol. Endocrinol. 2000, 14, 401–420. [Google Scholar] [CrossRef]
- Colin, E.M.; Weel, A.E.A.M.; Uitterlinden, A.G.; Buurman, C.J.; Birkenhager, J.C.; Pols, H.A.P.; van Leeuwen, J.P.T.M. Consequences of vitamin D receptor gene polymorphisms for growth inhibition of cultured human peripheral blood mononuclear cells by 1,25-dihydroxyvitamin D3. Clin. Endocrinol. 2000, 52, 211–216. [Google Scholar] [CrossRef]
- Decker, C.J.; Parker, R. Diversity of cytoplasmic functions for the 3′ untranslated region of eukaryotic transcripts. Curr. Opin. Cell Biol. 1995, 7, 386–392. [Google Scholar] [CrossRef]
- Miyamoto, K.; Kesterson, R.A.; Yamamoto, H.; Taketani, Y.; Nishiwaki, E.; Tatsumi, S.; Inoue, Y.; Morita, K.; Takeda, E.; Pike, J.W. Structural organization of the human vitamin d receptor chromosomal gene and its promoter. Mol. Endocrinol. 1997, 11, 1165–1179. [Google Scholar] [CrossRef]
- Mocellin, S.; Nitti, D. Vitamin D receptor polymorphisms and the risk of cutaneous melanoma: A systematic review and meta-analysis. Cancer 2008, 113, 2398–2407. [Google Scholar] [CrossRef]
- Birke, M.; Schope, J.; Wagenpfeil, S.; Vogt, T.; Reichrath, J. Association of Vitamin D receptor gene polymorphisms with melanoma risk: A meta-analysis and systematic review. Anticancer Res. 2020, 40, 583–595. [Google Scholar] [CrossRef]
- Caini, S.; Gnagnarella, P.; Stanganelli, I.; Bellerba, F.; Cocorocchio, E.; Queirolo, P.; Bendinelli, B.; Saieva, C.; Raimondi, S.; Gandini, S. Vitamin d and the risk of non-melanoma skin cancer: A systematic literature review and meta-analysis on behalf of the italian melanoma intergroup. Cancers 2021, 13, 4815. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line. | Disease | FAB | Sex and Age | Doubling Time | Gene Fusion | Gene Mutation; Zygosity |
|---|---|---|---|---|---|---|
| KG-1 | acute myeloid leukemia | M0 | Male, 59Y | 38 h | FGFR1OP2-FGFR1 | NRAS (p.Gly12Asp (c.35G>A)); unspecified TP53 (c.672+1G>A); homozygous |
| K562 | chronic myeloid leukemia | M1 | Female, 53Y | 47 h | BCR-ABL1 | TP53 (p.Gln136fs*13 (c.406_407insC)); homozygous |
| HL-60 | acute myeloid leukemia | M2 | Female, 36Y | 36–48 h | n/a | TP53, gene deletion; homozygous CDKN2A (p.Arg80Ter (c.238C>T) (p.Pro94Leu, c.281C>T)); homozygous NRAS (p.Gln61Leu (c.182A>T); heterozygous |
| MV-4-11 | acute monoblastic /monocytic leukemia | M5 | Male, 10Y | 40 h | KMT2A-AFF1 MLL-AFF1 ALL-1/AF4 | FLT3, unexplicit, internal tandem duplication; unspecified |
| Thp-1 | acute monoblastic /monocytic leukemia | M5 | Male, 1Y | 60–70 h | CSNK2A1-DDX39B KMT2A-MLLT3 MLL-MLLT3 MLL-AF9 | NRAS (p.Gly12Asp (c.35G>A)); heterozygous TP53 (p.Arg174fs*3 (c.520_545del26)); heterozygous |
| Jurkat | precursor T-cell acute lymphoblastic leukemia | L2 | Male, 14Y | 24 h | n/a | BAX (p.Glu41Argfs*19 (c.121delG) and p.Glu41Glyfs*33 (c.121dupG)); heterozygous FBXW7 (p.Arg505Cys (c.1513C>T)); heterozygous INPP5D (p.Gln345Ter (c.1033C>T) and (c.1097+1065_1097+1112del47)); heterozygous MSH2 (p.Arg711Ter (c.2131C>T)); homozygous MSH6 (p.Phe1088Serfs*2 (c.3261delC)); homozygous NOTCH1 (p.Arg1627His (c.4880G>A)); heterozygous TP53 (p.Arg196Ter (c.586C>T)); heterozygous |
| U2932 | diffuse large B-cell lymphoma | L2 | Female, 29Y | 48–50 h | n/a | n/a |
| Daudi | Burkitt lymphoma | L3 | Male, 16Y | 30–40 h | MYC-IGH | B2M (p.Met1Ile (c.3G>C)); homozygous CTNNB1 (p.Ala5_Ala80del)); homozygous TP53 (p.Gly266Glu (c.797G>A)); heterozygous |
| Raji | Burkitt lymphoma | L3 | Male, 11Y | 24–36 h | MYC-IGH | TP53 (p.Arg213Gln (c.638G>A) and p.Tyr234His (c.700T>C)); heterozygous |
| Cell Line | FokI | BsmI | ApaI | TaqI | Haplotype |
|---|---|---|---|---|---|
| leukemia | |||||
| MV-4-11 | Ff | bb | aa | TT | baT |
| Thp-1 | Ff | bb | aa | TT | baT |
| HL-60 | Ff | BB | AA | Tt | BAt |
| K562 | ff | Bb | Aa | Tt | BAT |
| KG-1 | ff | bb | Aa | TT | bAT |
| lymphoma | |||||
| Raji | FF | Bb | AA | Tt | BAT |
| Daudi | FF | Bb | AA | TT | BAT |
| Jurkat | Ff | Bb | Aa | TT | BAT |
| U2932 | FF | Bb | Aa | Tt | BAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gleba, J.J.; Kłopotowska, D.; Banach, J.; Turlej, E.; Mielko, K.A.; Gębura, K.; Bogunia-Kubik, K.; Kutner, A.; Wietrzyk, J. Polymorphism of VDR Gene and the Sensitivity of Human Leukemia and Lymphoma Cells to Active Forms of Vitamin D. Cancers 2022, 14, 387. https://doi.org/10.3390/cancers14020387
Gleba JJ, Kłopotowska D, Banach J, Turlej E, Mielko KA, Gębura K, Bogunia-Kubik K, Kutner A, Wietrzyk J. Polymorphism of VDR Gene and the Sensitivity of Human Leukemia and Lymphoma Cells to Active Forms of Vitamin D. Cancers. 2022; 14(2):387. https://doi.org/10.3390/cancers14020387
Chicago/Turabian StyleGleba, Justyna Joanna, Dagmara Kłopotowska, Joanna Banach, Eliza Turlej, Karolina Anna Mielko, Katarzyna Gębura, Katarzyna Bogunia-Kubik, Andrzej Kutner, and Joanna Wietrzyk. 2022. "Polymorphism of VDR Gene and the Sensitivity of Human Leukemia and Lymphoma Cells to Active Forms of Vitamin D" Cancers 14, no. 2: 387. https://doi.org/10.3390/cancers14020387