
The Large GTPase, GBP-2, Regulates Rho Family GTPases to Inhibit Migration and Invadosome Formation in Breast Cancer Cells



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells and Cell Culture
2.2. Western Blots
2.3. Antibodies
2.4. Colony Assay
2.5. Click-It EdU
2.6. Boyden Chamber
2.7. Wound Healing Assay
2.8. Generation of Control and mGBP-2-Directed shRNAs
2.9. Transfection and Cell Selection
2.10. Lentivirus Generation and Selection of GBP-2-Expressing 4T1 Cells
2.11. Immunofluorescence
2.12. Image Analysis
2.13. Rho GTPase Activity Assays
2.14. Gene Expression Profiling and Data Processing
2.15. Invadopodia Analysis
2.16. Statistical Analysis
3. Results
3.1. GBP-2 Correlates with Improved Distant-Metastasis Free Survival
3.2. GBP-2 Expression Inversely Correlates with Breast Cancer Migration
3.3. mGBP-2 Does Not Alter 4T1 or 67NR Cell Proliferation
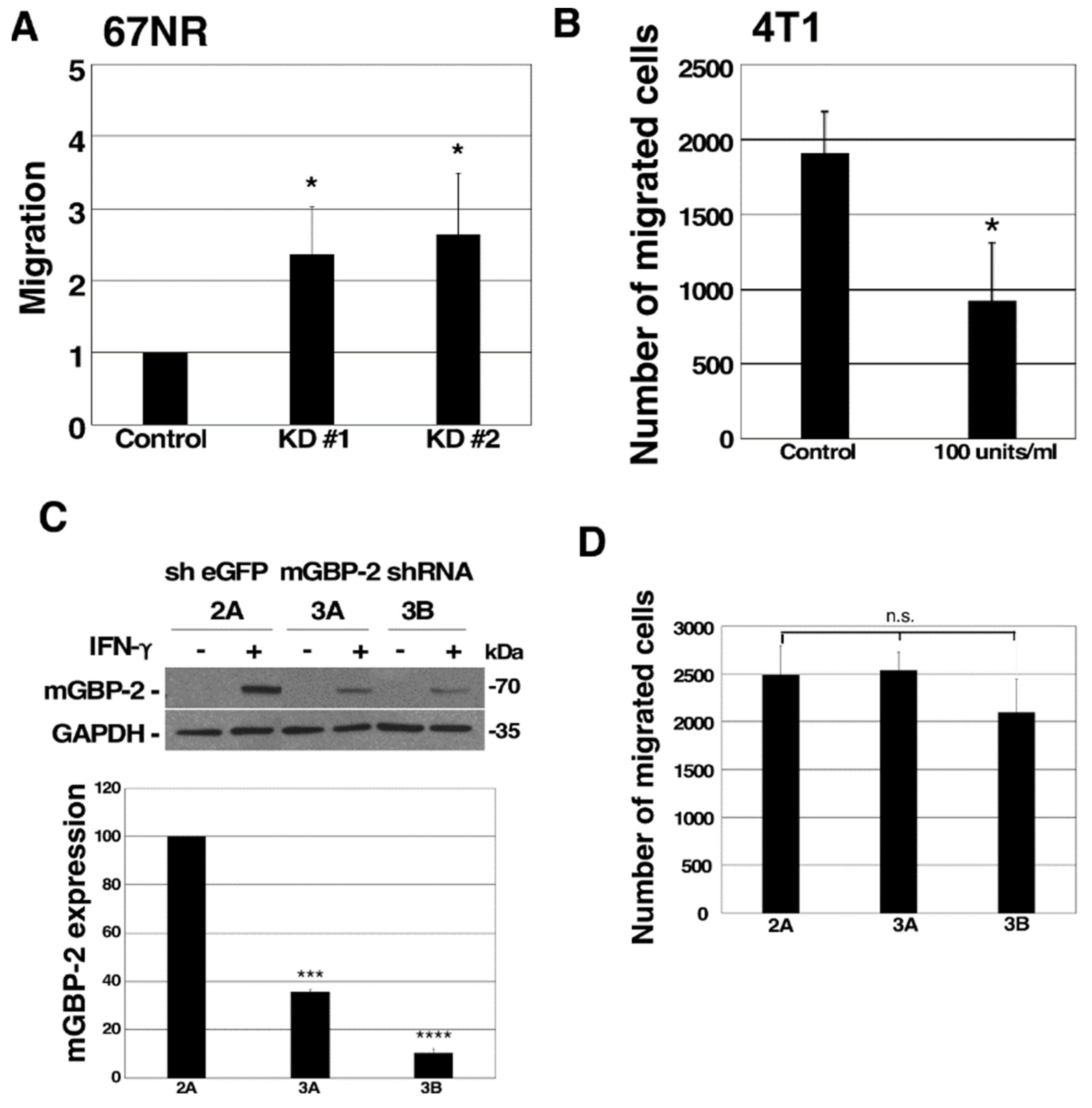
3.4. mGBP-2 Inhibits Murine TNBC Cell Migration
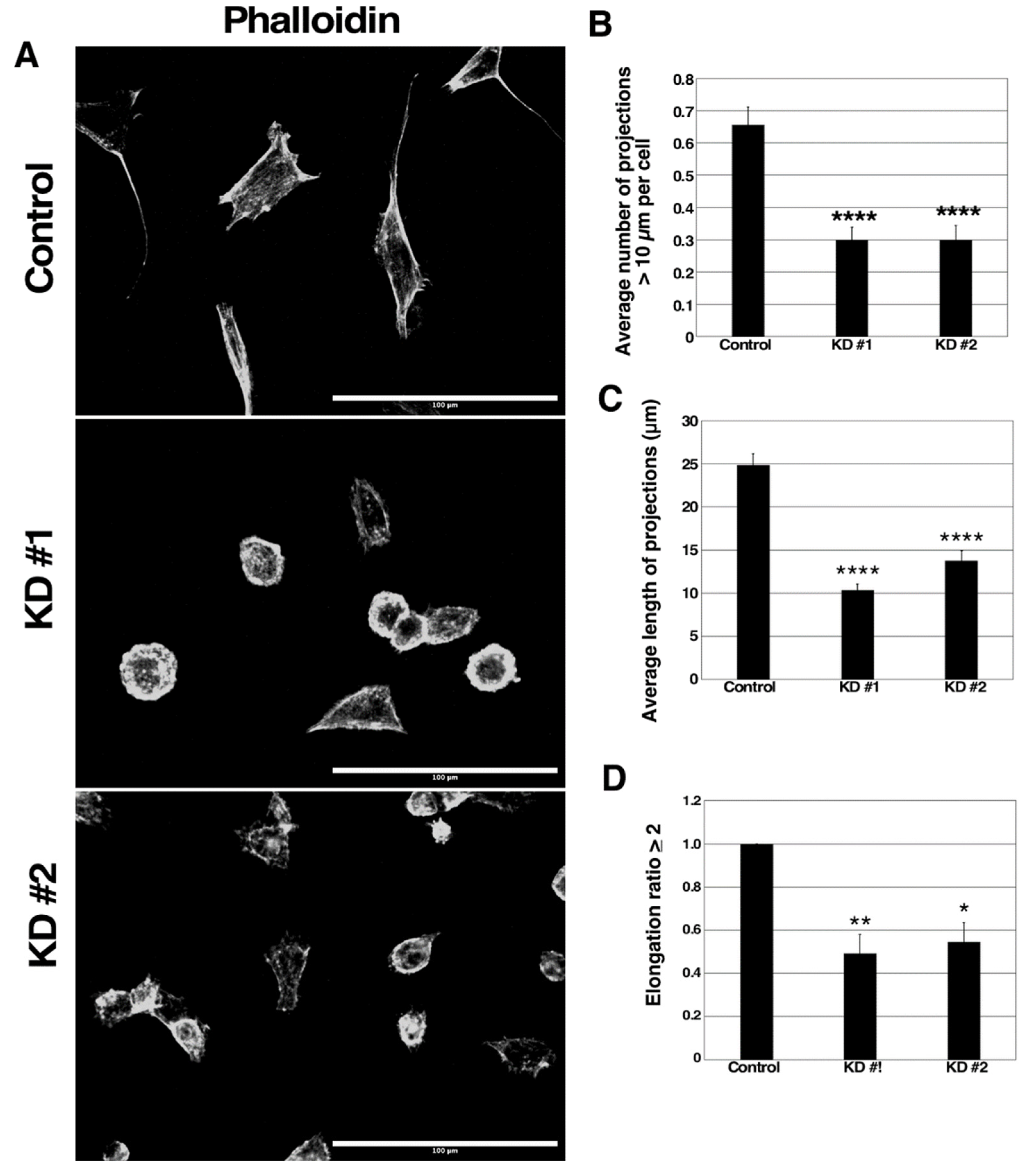
3.5. GBP-2 Alters 67NR Actin Cytoskeleton and Morphology
3.6. GBP-2 Promotes the Activation of CDC42 and RhoA and Inhibits the Activation of Rac1
3.7. mGBP-2 Inhibition of Rac Is Not Accompanied by Inhibition of Activation of Akt
3.8. mGBP-2 Inhibits the Generation of Invadosomes in 67NR Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Breastcancer.org. U.S. Breast Cancer Statistics. Available online: www.breastcancer.org/symptoms/understand_bc/statistics2018 (accessed on 3 January 2019).
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Vestal, D.J.; Jeyaratnam, J.A. The Guanylate-Binding Proteins: Emerging Insights into the Biochemical Properties and Functions of This Family of Large Interferon-Induced Guanosine Triphosphatase. J. Interf. Cytokine Res. 2011, 31, 89–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Britzen-Laurent, N.; Lipnik, K.; Ocker, M.; Naschberger, E.; Schellerer, V.S.; Croner, R.S.; Vieth, M.; Waldner, M.; Steinberg, P.; Hohenadl, C.; et al. GBP-1 acts as a tumor suppressor in colorectal cancer cells. Carcinogenesis 2012, 34, 153–162. [Google Scholar] [CrossRef] [Green Version]
- Lipnik, K.; Naschberger, E.; Gonin-Laurent, N.; Kodajova, P.; Petznek, H.; Rungaldier, S.; Astigiano, S.; Ferrini, S.; Sturzl, M.; Hohenadl, C. Interferon g-induced human guanylate-binding protein 1 inhibits mammary tumor growth in mice. Mol. Med. 2010, 16, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Ascierto, M.L.; Kmieciak, M.; Idowu, M.O.; Manjili, R.; Zhao, Y.; Grimes, M.; Dumur, C.; Wang, E.; Ramakrishnan, V.; Wang, X.-Y.; et al. A signature of immune function genes associated with recurrence-free survival in breast cancer patients. Breast Cancer Res. Treat. 2011, 131, 871–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehmann, B.; Bauer, J.A.; Chen, X.; Sanders, M.E.; Chakravarthy, A.B.; Shyr, Y.; Pietenpol, J.A. Identification of human triple-negative breast cancer subtypes and preclinical models for selection of targeted therapies. J. Clin. Investig. 2011, 121, 2750–2767. [Google Scholar] [CrossRef] [Green Version]
- Chan, S.R.; Vermi, W.; Luo, J.; Lucini, L.; Rickert, C.; Fowler, A.M.; Lonardi, S.; Arthur, C.; Young, L.J.T.; Levy, D.E.; et al. STAT1-deficient mice spontaneously develop estrogen receptor a-positive luminal mammary carcinomas. Breast Cancer Res. 2012, 14, R16. [Google Scholar] [CrossRef] [Green Version]
- Burstein, M.D.; Tsimelzon, A.; Poage, G.M.; Covington, K.R.; Contreras, A.; Fuqua, S.A.; I Savage, M.; Osborne, C.K.; Hilsenbeck, S.G.; Chang, J.C.; et al. Comprehensive Genomic Analysis Identifies Novel Subtypes and Targets of Triple-Negative Breast Cancer. Clin. Cancer Res. 2014, 21, 1688–1698. [Google Scholar] [CrossRef] [Green Version]
- Godoy, P.; Cadenas, C.; Hellwig, B.; Marchan, R.; Stewart, J.D.; Reif, R.; Lohr, M.; Gehrmann, M.; Rahnenführer, J.; Schmidt, M.; et al. Interferon-inducible guanylate binding protein (GBP2) is associated with better prognosis in breast cancer and indicates an efficient T cell response. Breast Cancer 2012, 21, 491–499. [Google Scholar] [CrossRef]
- Vestal, D.J. The guanylate binding proteins (GBPs): Pro-inflammatory cytokine-induced members of the dynamin superfamily with unique GTPase activity. J. Interferon Cytokine Res. 2005, 25, 435–443. [Google Scholar] [CrossRef]
- Forster, F.; Paster, W.; Supper, V.; Schatzlmaier, P.; Sunzenauer, S.; Ostler, N.; Saliba, A.; Eckerstorfer, P.; Britzen-Laurent, N.; Schütz, G.; et al. Guanylate Binding Protein 1–Mediated Interaction of T Cell Antigen Receptor Signaling with the Cytoskeleton. J. Immunol. 2013, 192, 771–781. [Google Scholar] [CrossRef] [Green Version]
- Guenzi, E.; Topolt, K.; Cornali, E.; Lubeseder-Martellato, C.; Jorg, A.; Matzen, K.; Zietz, C.; Kremmer, E.; Nappi, F.; Schwemmle, M.; et al. The helical domain of GBP-1 mediates the inhibition of endo-thelial cell proliferation by inflammatory cytokines. The EMBO J. 2001, 20, 5568–5577. [Google Scholar] [CrossRef] [Green Version]
- Guenzi, E.; Topolt, K.; Lubeseder-Martellato, C.; Jorg, A.; Naschberger, E.; Benelli, R.; Albini, A.; Sturzl, M. The guanylate binding protein-1 GTPase controls the invasive and angiogenic capabiliy of endothelial cells through inhibition of MMP-1 expression. EMBO J. 2003, 22, 3772–3782. [Google Scholar] [CrossRef]
- Capaldo, C.T.; Beeman, N.; Hilgarth, R.S.; Nava, P.; Louis, N.A.; Naschberger, E.; Sturzl, M.; Parkos, C.A.; Nusrat, A. IFN-g and TNF-a-induced GBP-1 inhibits epithelial cell proliferation through suppression of b-catenin/TCF signaling. Mucosal Immunol. 2012, 5, 681–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostler, N.; Britzen-Laurent, N.; Liebl, A.; Naschberger, E.; Lochnit, G.; Ostler, M.; Forster, F.; Kunzelmann, P.; Ince, S.; Supper, V.; et al. Gamma Interferon-Induced Guanylate Binding Protein 1 Is a Novel Actin Cytoskeleton Remodeling Factor. Mol. Cell. Biol. 2014, 34, 196–209. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhang, Y.; Wu, W.; Wang, F.; Liu, X.; Shui, G.; Nie, C. Guanylate-binding protein 2 regulates Drp1-mediated mitochondrial fission to suppress breast cancer cell invasion. Cell Death Dis. 2017, 8, e3151. [Google Scholar] [CrossRef] [PubMed]
- Gorbacheva, V.Y.; Lindner, D.; Sen, G.C.; Vestal, D.J. The IFN-induced GTPase, mGBP-2: Role in IFN-g induced murine fibroblast proliferation. J. Biol. Chem. 2002, 277, 6080–6087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messmer-Blust, A.F.; Balasubramanian, S.; Gorbacheva, V.Y.; Jeyaratnam, J.A.; Vestal, D.J. The interferon-g-induced murine Guanylate-Binding Protein-2 (mGBP-2) inhibits Rac activation during cell spreading on fibronectin and after platelet-derived growth factor (PDGF) treatment: Role for Phosphatidylinositol 3-Kinase. Mol. Biol. Cell. 2010, 15, 2514–2528. [Google Scholar] [CrossRef] [Green Version]
- Aslakson, C.J.; Miller, F.R. Selective events in the metastatic process defined by analysis of the sequential dissemination of sub-populations of a mouse mammary tumor. Cancer Res. 1992, 52, 1399–1405. [Google Scholar] [PubMed]
- Vestal, D.J.; Gorbacheva, V.Y.; Sen, G.C. Different subcellular localizations for the related interferon-induced GTPases, mGBP-1 and mGBP-2: Implications for different functions? J. Interferon Cytokine Res. 2000, 20, 991–1000. [Google Scholar] [CrossRef]
- Vestal, D.J.; Buss, J.E.; McKercher, S.R.; Jenkins, N.A.; Copeland, N.G.; Kelner, G.S.; Asundi, V.K.; Maki, R.A. Murine GBP-2: A new IFN-γ-induced member of the GBP family of GTPases isolated from macrophages. J. Interferon Cytokine Res. 1998, 18, 977–985. [Google Scholar] [CrossRef]
- Salic, A.; Mitchison, T.J. A chemical method for fast and sensitive detection of DNA synthesis in vivo. Proc. Natl. Acad. Sci. USA 2008, 105, 2415–2420. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.-C. Boyden Chamber Assay. Methods Mol. Biol. 2005, 294, 15–22. [Google Scholar] [CrossRef]
- Doyle, W. Operations Useful for Similarity-Invariant Pattern Recognition. J. ACM 1962, 9, 259–267. [Google Scholar] [CrossRef]
- Gyorffy, B.; Lanczky, A.; Eklund, A.C.; Denkert, C.; Budczies, J.; Li, Q.; Szallasi, Z. An online survival analysis tool to rapdily assess the effect of 22, 277 genes on breast cancer prognosis using microarray data of 1809 patients. Breast Cancer Res. Treat. 2010, 123, 725–731. [Google Scholar] [CrossRef] [Green Version]
- Unterer, B.; Wiesmann, V.; Gunasekaran, M.; Sticht, H.; Tenkerian, C.; Behrans, J.; Leone, M.; Engel, F.B.; Britzen-Laurent, N.; Nas-chberger, E.; et al. IFN-g-response mediator GBP-1 represses human cell proliferation by inhibiting the Hippo signaling transcription factor TEAD. Biochem. J. 2018, 475, 2955–2967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, S.; Mu, Z.; Huang, Y.; Ji, P. Guanylate Binding Protein 1 Inhibits Osteogenic Differentiation of Human Mesenchymal Stromal Cells Derived from Bone Marrow. Sci. Rep. 2018, 8, 1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, Q.; Wang, A.; Cheng, Y.; Mukasa, A.; Ma, J.; Hong, L.; Yu, S.; Sun, L.; Huang, Q.; Purow, B.; et al. Guanylate binding protein-1 mediates EGFRvIII and promotes glioblastoma growth in vivo but not in vitro. Oncotarget 2016, 7, 9680–9691. [Google Scholar] [CrossRef]
- Wang, Y.; Shenouda, S.; Baranwal, S.; Rathinam, R.; Jain, P.; Bao, L.; Hazari, S.; Dash, S.; Alahari, S.K. Integrin subunits alpha5 and alpha6 regulate cell cycle by modulating the chk1 and Rb/E2F pathways to affect breast cancer metastasis. Mol. Cancer 2011, 10, 84. [Google Scholar] [CrossRef] [Green Version]
- Bao, L.; Haque, A.; Jackson, K.; Hazari, S.; Moroz, K.; Jetly, R.; Dash, S. Increased Expression of P-Glycoprotein Is Associated with Doxorubicin Chemoresistance in the Metastatic 4T1 Breast Cancer Model. Am. J. Pathol. 2011, 178, 838–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Ma, G.; Jing, C.; Liu, Z. Guanylate-binding protein 1 (GBP1) promotes lymph node metastasis in human esophageal squamous cell carcinoma. Discov. Med. 2015, 20, 369–378. [Google Scholar] [PubMed]
- Li, M.; Mukasa, A.; Inda, M.D.-M.; Zhang, J.; Chin, L.; Cavenee, W.; Furnari, F. Guanylate binding protein 1 is a novel effector of EGFR-driven invasion in glioblastoma. J. Exp. Med. 2011, 208, 2657–2673. [Google Scholar] [CrossRef] [Green Version]
- Weinländer, K.; Naschberger, E.; Lehmann, M.H.; Tripal, P.; Paster, W.; Stockinger, H.; Hohenadl, C.; Stürzl, M. Guanylate binding protein-1 inhibits spreading and migration of endothelial cells through induction of integrin α4expression. FASEB J. 2008, 22, 4168–4178. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.-J.; Chang, K.-P.; Chang, Y.-J.; Hsu, C.-W.; Liang, Y.; Yu, J.-S.; Chi, L.-M.; Chang, Y.-S.; Wu, C.-C. Identification of Guanylate-Binding Protein 1 as a Potential Oral Cancer Marker Involved in Cell Invasion Using Omics-Based Analysis. J. Proteome Res. 2011, 10, 3778–3788. [Google Scholar] [CrossRef]
- Balasubramanian, S.; Fan, M.; Messmer-Blust, A.F.; Yang, C.H.; Trendel, J.A.; Jeyaratnam, J.A.; Pfeffer, L.M.; Vestal, D.J. The interfer-on-g-induced GTPase, mGBP-2, inhibits tumor necrosis factor a (TNF-a) induction of matrix metalloproteinase-g (MMP-9) by in-hibiting NF-kB and Rac protein. J. Bio. Chem. 2011, 286, 20054–20064. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Posadas, R.; Sturzl, M.; Atreya, I.; Neurath, M.F.; Britzen-Laurent, N. Interplay of GTPases and cytoskeleton in cellular barrier defeccts during gut inflammation. Front Immunol. 2017, 8, 1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayala, I.; Baldassarre, M.; Caldieri, G.; Buccione, R. Invadopodia: A guided tour. Eur. J. Cell Biol. 2006, 85, 159–164. [Google Scholar] [CrossRef]
- Gimona, M.; Buccione, R.; Courtneidge, S.A.; Linder, S. Assembly and biological role of podosomes and invadopodia. Curr. Opin. Cell Biol. 2008, 20, 235–241. [Google Scholar] [CrossRef]
- Murphy, D.A.; Courtneidge, S.A. The ’ins’ and ’outs’ of podosomes and invadopodia: Characteristics, formation and function. Nat. Rev. Mol. Cell Biol. 2011, 12, 413–426. [Google Scholar] [CrossRef] [Green Version]
- Hoshino, D.; Branch, K.M.; Weaver, A.M. Signaling inputs to invadopodia and podosomes. J. Cell Sci. 2013, 126, 2979–2989. [Google Scholar] [CrossRef] [Green Version]
- Goicoechea, S.M.; Zinn, A.; Awadia, S.S.; Snyder, K.; Garcia-Mata, R. A RhoG-mediated signaling pathway that modulates in-vadopodia dynamics in breast cancer cells. J. Cell Sci. 2017, 130, 1064–1077. [Google Scholar] [PubMed] [Green Version]
- Eckert, M.; Lwin, T.M.; Chang, A.T.; Kim, J.; Danis, E.; Ohno-Machado, L.; Yang, J. Twist1-Induced Invadopodia Formation Promotes Tumor Metastasis. Cancer Cell 2011, 19, 372–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nobes, C.D.; Hall, A. Rho, Rac, and Cdc42 GTPases regulate the assembly of multimolecular focal complexes associated with actin stress fibers, lamellipodia, and filopodia. Cell 1995, 81, 53–62. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nyabuto, G.O.; Wilson, J.P.; Heilman, S.A.; Kalb, R.C.; Kopacz, J.P.; Abnave, A.V.; Vestal, D.J. The Large GTPase, GBP-2, Regulates Rho Family GTPases to Inhibit Migration and Invadosome Formation in Breast Cancer Cells. Cancers 2021, 13, 5632. https://doi.org/10.3390/cancers13225632
Nyabuto GO, Wilson JP, Heilman SA, Kalb RC, Kopacz JP, Abnave AV, Vestal DJ. The Large GTPase, GBP-2, Regulates Rho Family GTPases to Inhibit Migration and Invadosome Formation in Breast Cancer Cells. Cancers. 2021; 13(22):5632. https://doi.org/10.3390/cancers13225632
Chicago/Turabian StyleNyabuto, Geoffrey O., John P. Wilson, Samantha A. Heilman, Ryan C. Kalb, Jonathan P. Kopacz, Ankita V. Abnave, and Deborah J. Vestal. 2021. "The Large GTPase, GBP-2, Regulates Rho Family GTPases to Inhibit Migration and Invadosome Formation in Breast Cancer Cells" Cancers 13, no. 22: 5632. https://doi.org/10.3390/cancers13225632